Universidade Castelo Branco
Rainer Alves da Fonseca - 2007000249
FILO ACANTHOCEPHALA
Rio de Janeiro
2008
Rainer Alves da Fonseca - 2007000249
FILO ACANTHOCEPHALA
Trabalho
realizado
cumprimento
para
parcial
o
da
disciplina de Helmintologia e
Metazoários
Inferiores,
graduação
em
Ciências
Biológicas.
Prof. Diego da Silva Ferreira
Rio de Janeiro
2008
da
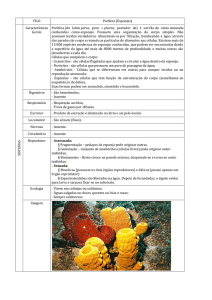
Porífera
Os poríferos, ou espongiários, constituem o filo mais primitivo dos metazoários,
embora sejam pluricelulares, suas células formam agregados frouxos, não
constituindo tecidos verdadeiros. As esponjas são animais aquáticos,
predominantemente marinhos com algumas formas de água doce, são sésseis
(quando adultos) e filtradores. Sua organização é muito simples, no geral as
esponjas têm o aspecto de um vaso, podem ser ramificadas, globulares ou de
formas variadas, tendo um aspecto poroso (o corpo com muitas aberturas), daí o
nome Porífera. As esponjas menores e simples mostram simetria radial, enquanto
a maioria é assimétrica.
Esponja Gigante
Realizam digestão intracelular (não apresentam cavidade digestiva). A respiração e
a excreção ocorrem por difusão direta entre as células e a água que circula através
dos
canais
do
corpo.
A parede do corpo é formada pela epiderme, pelo mesênquima, e pelo
revestimento interno de células flageladas com colarinho transparente, chamadas
coanócitos.
A epiderme é constituída por uma fina camada de células achatadas, os
pinacócitos,
revestindo
externamente
os
espongiários.
No mesênquima encontram-se células móveis e indiferenciadas os amebócitos,
que participam da digestão distribuindo o alimento pelas diversas células do corpo,
podendo também se diferenciar em gametas, além disso são responsáveis pela
regeneração celular. A sustentação também é garantida pelo mesênquima
(gelatinoso), interno à camada de pinacócitos: no mesênquima estão mergulhadas
espículas de calcário ou silício.O endoesqueleto é formado por espículas minerais
(calcárias ou silicosas, secretadas pelas células escleroblastos) e por filamentos da
proteína espongina.
A água entra por numerosos poros (porócitos) na superfície do seu corpo e após
circular na cavidade do átrio ou espongiocele (paragáster) sairá pelo ósculo, que
é a abertura no pólo superior. Esse sentido de movimentação da água é
determinado pelo batimento unidirecional (direção oposta ao corpo celular) de
flagelos dos coanócitos. Outra função destas “células em colarinho” é fagocitar os
alimentos que estão em suspensão na água e realizarem a única forma de digestão
desses
animais:
intracelular.
Os espongiários vivem isolados ou em colônias. Sua coloração é variável devido a
associações com algas: acinzentadas, avermelhadas, transparentes, amareladas,
verdes, azuis - proteção à radiação solar ou coloração de alerta.
Quando industrializadas para a produção de esponjas de banho, as esponjas
perdem totalmente o seu esqueleto mineral, sobrando apenas às fibras de
espongina. As esponjas apresentam simetria radial ou podem ser assimétricas.
Tipos de esponjas, com crescente grau de complexidade: Ascon, Sycon,
Leucon.
Áscon
A forma primitiva dos espongiários é a de um tubo ou vaso, fixado no substrato. Na
extremidade apical aparece uma grande abertura — o ósculo — que serve para a
saída da água que continuamente atravessa o corpo da esponja. A parede do
corpo é provida de um grande número de poros (daí o nome de porífera), através
dos
quais
penetram
água
e
partículas
alimentares.
Nos áscon, bem como nos outros dois tipos, não existem órgão diferenciados, mas,
distinguem-se diversos tipos celulares adaptados a determinadas funções. A
parede do corpo é formada por duas camadas celulares. A camada mais externa é
dermal, de origem ectodérmica, e a mais interna, denominada gastral, tem origem
endodérmica. Entre as duas camadas celulares, há um mesênquima gelatinoso. A
cavidade central do corpo é chamada átrio ou espongiocela. Nas duas camadas
celulares e no mesênquima, encontramos os seguintes tipos celulares:
* Pinacócitos: são células achatadas que, justapostas, formam a camada dermal.
* Coanócitos: são células flageladas e providas de um colarinho, uma formação
membranosa que envolve o flagelo. Revestem a cavidade atrial e constituem a
camada
gastral.
* Porócitos: são células, percorridas por uma perfuração cônica. São estas
perfurações dos porócitos que constituem os numerosos poros que ligam o átrio ao
meio
externo.
* Miócitos: são células alongadas e contrácteis, que formam esfíncter em torno
dos
poros
e
do
ósculo.
* Amebócitos: células, situadas no mesênquima, que possuem movimento
emebóide,
realizando
várias
funções.
Sícon
Observada externamente, apresenta-se como uma urna alongada, fixada pela
extremidade superior, circundado por uma coroa de espículas longas e afiliadas. A
superfície do corpo possui numerosas elevações ou papilas, das quais saem
pequenas
espículas.
Entre
as
papilas
aparecem
os
poros.
Cortada longitudinalmente, apresenta a parede do corpo espessa e com uma série
de dobras, formando curtos canais horizontais. Distinguimos dois tipos de canais:
inalantes e exalantes. Os primeiros abrem-se na superfície externa e terminam em
fundo cego. Os canais exalantes, são internos e desembocam no átrio.
A superfície externa e os canais inalantes são revestidos pela camada dermal,
formada por pinacócitos, ficando os coanócitos limitados aos canais exalantes. O
mesênquima gelatinoso é bem mais desenvolvido do que no áscon, contém
amebócitos e espículas.
Lêucon
É o tipo mais evoluído, o átrio é reduzido, enquanto a parede do corpo é bastante
desenvolvida e percorrida por um complicado sistema de canais e câmaras. Os
coanócitos encontram-se revestindo câmaras esféricas, também denominada
câmaras vibráteis, interpostas num sistema de canais. Os canais partem dos poros
e atingem as câmaras transportando água são denominadas inalantes ou
aferentes. Das câmaras saem os canais exalantes ou eferentes que atingem o
átrio.
Os coanócitos só aparecem nas câmaras vibráteis. Os pinacócitos revestem a
superfície externa, o átrio e os diversos canais. No desenvolvimento do
mesênquima encontramos amebócitos e espículas.
REPRODUÇÃO NAS ESPONJAS
Reprodução Assexuada
Fragmentação
A elevada capacidade de regeneração apresentada pelas esponjas permite que
fragmentos eventualmente separados de um indivíduo originem novas esponjas.
Brotamento
Muitas esponjas reproduzem-se assexuadamente por brotamento. Formam-se
expansões ou brotos, na superfície da esponja-mãe, que podem se separar do
organismo genitor e constituir novos indivíduos. O brotamento, sem a posterior
separação dos brotos, leva à formação e ao crescimento de colônias.
Gemulação
Certas esponjas de água doce formam estruturas denominadas gêmulas. Estas
consistem de um pequeno "pacote" de amebócitos, abrigados em um envoltório
resistente que contém espículas.
As gêmulas são capazes de resistir à falta de água durante uma estação seca
prolongada, em que o lago ou rio onde a esponja vive seca por completo, até que
retornem as condições de umidade favoráveis, quando os amebócitos se libertam
do
envoltório
e
originam
uma
nova
esponja.
Reprodução Sexuada
A maioria das esponjas apresenta reprodução sexuada. Algumas espécies são
hermafroditas ou monóicas, isto é, o mesmo indivíduo forma gametas masculinos e
femininos, enquanto outras são dióicas, isto é, os indivíduos apresentam sexos
separados.
Tanto os óvulos como os espermatozóides se formam a partir dos amebócitos. Os
espermatozóides são liberados na água, enquanto os óvulos ficam presos à parede
do corpo da esponja.
Os espermatozóides nadam ativamente para dentro do átrio da esponja fêmea,
onde penetram na parede do corpo e fecundam os óvulos ali presentes. O zigoto
assim formado se multiplica, originando uma pequena bola de células flageladas,
que se transforma em uma larva, que em algumas espécies é a parenquímula e,
em
outras,
a
anfiblástula.
A larva anfiblástula, um estágio jovem da futura esponja, se liberta da parede do
corpo, atingindo o exterior através do ósculo. Após nadar durante certo tempo, a
anfiblástula se fixa à um substrato submerso, onde origina uma nova esponja.
Como há um estágio larval entre o zigoto e o adulto, diz-se que as esponjas
apresentam
desenvolvimento
indireto.
Ciclo sexuado de uma esponja
Ecologia
Esponjas são organismos dominantes em muitos habitats marinhos bentônicos. A
maioria dos litorais rochosos abrigam um grande número de espécies, e elas são
também abundantes em tôrno da Antártida, onde boa parte do substrato
consolidado deve-se a uma espessa camada de espículas silicosas. Pouco se sabe
sobre suas taxas de crescimento, mas os dados disponíveis sugerem uma grande
variação entre as espécies. Por exemplo, a esponja Terpios sp. das Filipinas
cresce até 2.3 cm por mês sobre corais, hidrocorais, moluscos e algas, matando-os
através da liberação de uma toxina e por sufocamento. De modo geral as esponjas
parecem ser animais bastante estáveis e de vida longa. Embora algumas espécies
tenham um ciclo de vida anual (por exemplo, Sycon sp), estimativas baseadas em
taxas de crescimento conferem idades acima de 1500 anos a indivíduos algumas
espécies (Lehnert & Reitner, 1997). Se confirmadas estas estimativas as esponjas
seriam os animais com tempo de vida mais longa do planeta.
Vários animais se alimentam de esponjas, embora o dano causado por estes
predadores seja geralmente pequeno. Alguns moluscos, ouriços e estrelas-do-mar,
além de peixes tropicais (donzelas, peixes-borboleta) e tartarugas, comem
esponjas. Muitas espécies são totalmente expostas aos predadores, e na
impossibilidade de bater em retirada apresentam mecanismos alternativos de
defesa contra a predação excessiva. O mecanismo primário de defesa das
esponjas é de natureza química. As esponjas produzem uma ampla gama de
compostos tóxicos, alguns bastante potentes. Espécies de alguns gêneros como
Tedania e Neofibularia, podem mesmo causar dermatites dolorosas em seres
humanos. Muitas espécies produzem compostos com atividade antimicrobiana
(antibacteriana, antifúngica, antiviral). Além de defesas antipredação e contra
infecções microbianas, as toxinas de esponjas servem também para a competição
por espaço com outros invertebrados, como briozoários, ascídias, corais e até
mesmo outras esponjas. Isto permite a algumas esponjas crescer rapidamente e
recobrir
a
fauna
e
a
flora
adjacentes.
Relações de comensalismo envolvendo esponjas são muito comuns. O intrincado
sistema de canais das esponjas e suas defesas antipredação as tornam excelentes
refúgios para uma horda de invertebrados menores (crustáceos, ofiuróides,
poliquetos) e alguns peixes (gobídeos e blennídeos). Várias espécies dependem
dessa proteção das esponjas em sua fase de juvenis para manterm suas
populações em níveis estáveis. Por exemplo, conhece-se um peixe no Japão que
desova dentro de uma esponja (Mycale adhaerens), valendo-se da química desta
espécie para a proteção de seus ovos. Outros organismos usam as esponjas como
cobertura ou camuflagem, como os caranguejos do gênero Dromia, que recortam
pedaços de esponjas de diversas espécies e os posicionam sobre sua carapaça.
Outras associações muito comuns são aquelas envolvendo esponjas e
microorganismos endossimbiontes, principalmente bactérias e cianofíceas.
Presumivelmente, a matriz extracelular das esponjas provê um meio rico para o
crescimento das bactérias, e o hospedeiro se beneficia de um estoque de bactérias
utilizável em sua nutrição. As esponjas são os únicos metazoários conhecidos a
manter relações simbióticas com cianofíceas, que produzem glicerol e compostos
fosfatados para a nutrição das esponjas. Esponjas portadoras de cianobactérias
funcionam como produtores primários, e apresentam um crescimento rápido e alta
produtividade primária em recifes de coral.
Histórico
e
Classificação
Conhecidas e utilizadas desde a antiguidade pelos povos primitivos as esponjas
foram incluídas já no primeiro tratado sobre classificação de organismos, escrito
em 350 A.C. na Grécia clássica por Aristóteles. Consideradas inicialmente como
plantas, sua natureza animal só foi reconhecida no final do século XVIII, quando se
observaram as correntes de água no seu corpo. No entanto, os grandes
naturalistas da época (Lammarck, Lineu, Cuvier) classificavam as esponjas como
Zoophyta (animais-plantas) ou Pólipos (considerando-as como próximas dos
cnidários). Foi o naturalista inglês R.E. Grant quem primeiro compreendeu a
anatomia e fisiologia das esponjas e criou o nome Porifera. A elevação de Porifera
ao nível de Filo, sugerida por Huxley em 1875 e por Sollas em 1884, só foi aceita
no início do século XX. Ainda assim, os debates a respeito de sua posição em
relação aos protozoários e metazoários permaneceram até recentemente.
Do nível de ordem até espécie, os principais caracteres de importância sistemática
ou seja, auxiliares no reconhecimento de grupos de parentesco, são a cor, a forma,
a textura da superfície, a forma das aberturas inalantes e exalantes, a consistência,
o esqueleto e os tipos de espículas. Estes caracteres são muito importantes para a
distinção a nível específico e devem ser obrigatoriamente anotados durante a
coleta, pois frequentemente se perdem após a fixação. Portanto, a fotografia
submarina tem uma importância fundamental para a identificação das espécies de
esponjas, já que é o único meio de preservar as caraterísticas externas da esponja
como elas eram em vida. A utilização exclusiva dos dados de forma, cor e
características da superfície raramente no entanto permite uma identificação
completa das espécies. Para que uma espécie de esponja seja identificada com
segurança é necessário que se façam preparações do esqueleto e das espículas
dissociadas, montados em lâminas de microscopia para observação em
microscópio ótico (100-400x de aumento). Para a observação do esqueleto um
método simples é fazer dois cortes finos com lâmina de barbear ou bisturi: um
tangencial à superfície e outro perpendicular. Em seguida, imergir estes cortes em
etanol 96% por alguns minutos, secá-los e montá-los em lâmina de microscopia
com bálsamo do Canadá ou outro meio de montagem. Para observar as espículas
dissociadas, pode-se banhar um pedaço pequeno da esponja em água sanitária
(hipoclorito de sódio) para remover todo o material orgânico. Em seguida o resíduo,
que contém as espículas e outras partículas inorgânicas, é lavado várias vezes por
decantação em água doce. Por último, as espículas são então colocadas nas
lâminas, secas e montadas com bálsamo do Canadá, podendo assim ser
observadas
em
maior
detalhe.
A classificação do Filo Porifera é ainda hoje muito debatida, desde a definição das
espécies até as relações entre as classes e subclasses. Os caracteres
morfológicos e esqueléticos, insuficientes para um estudo filogenético amplo do
grupo, vêm sendo complementados por uma diversidade cada vez maior de
critérios alternativos (incluindo métodos bioquímicos, moleculares, embriológicos,
histológicos e citológicos) para a taxonomia de Porifera. A grande variabilidade
morfológica das esponjas e a dificuldade de se estabelecerem os limites das
espécies ainda hoje leva muitos estudantes de taxonomia de esponjas à frustração
muito
cedo
em
suas
carreiras.
A fauna de poríferos da costa Atlântica da América do Sul é uma das menos
conhecidas do mundo, e a maior parte do conhecimento sobre a espongofauna
brasileira vêm de dragagens na plataforma continental efetuadas por expedições
estrangeiras (p.ex. navios "Challenger" e "Calypso"). Das espécies de
Demospongiae conhecidas para a Costa Brasileira, cerca de 73% (194/267) foram
registradas por pesquisadores estrangeiros. As espécies litorais, de fácil acesso
por mergulho livre, têm sido pouco estudadas, com coletas esparsas ao longo da
costa. Recentemente essa tendência vem se invertendo, e os poucos grupos de
pesquisa dedicados ao estudo das esponjas marinhas do Brasil (coordenados por
Beatriz Mothes e Rosária de Rosa Barbosa na Fundação Zoobotânica de Porto
Alegre; por Eduardo Hajdu e Guilherme Muricy no Museu Nacional, e por Solange
Peixinho na Universidade Federal da Bahia) começam a intensificar o estudo
taxonômico das espécies litorais, principalmente através de coletas por mergulho
autôno.
As esponjas perfurantes (gêneros Cliona e Aka) escavam galerias complexas em
substratos calcáreos como corais e conchas de moluscos. Apesar de causarem
danos significativos às culturas comerciais de ostras, a biorosão causada por estas
esponjas em recifes de coral auxiliam no processo de crescimento do recife. O
processo de perfuração envolve a remoção química de pastilhas de carbonato de
cálcio que contribuem para a deposição de sedimentos no local. Esse sedimento
será depois reincorporado ao recife pelo processo de cimentação, que envolve
fatores físicos, químicos e biológicos e que é fundamental para o crescimento do
recife de coral. As esponjas auxiliam também por crescerem sobre os grãos de
sedimento, mantendo-os agregados e estabilizados por mais tempo e facilitando a
ação de microorganismos cimentadores.
Características dos principais grupos de corais fósseis
RUGOSA
TABULATA
Redondo ou polígono Redondo, polígono
ou oval
Exclusivamente
Corallum
Solitário ou colonial
colonial
Sempre pequenos
Coralitos
Relativamente
grandes
Bem desenvolvida
Rugosidade
Presente
ocasionalmente
Composição
Calcita
Calcita
Tábulas
Presentes,
exceto Sempre presentes.
quando substituídas Planas,
convexas
por dissepimentos
ou côncavas
Dissepimentos
Distintas, parecidas Partes indistintas do
com bolhas no cálice; cenósteo
solitários ou juntos
com septos e tábulas
Septos
Presentes,
exceto Ausentes
quando substituídas
por dissepimentos
Extensão
dos Confinados ao cálice --septos
Organização dos Inserção tetrâmera, --septos
em quatro compartimentos (Tetracorais)
Forma dos septos Comumente curvos
--Bossa
axial
/ Presente em alguns
Ausente
columela
Poros nas paredes Ausentes
Presente em alguns
Paleozóico:
Paleozóico:
Distribuição
Ordoviciano
a Ordoviciano
a
geológica
Permiano
Permiano
(temporal)
Cálice
SCLERACTINIA
Símples a complexo
Solitário ou colonial
Relativamente grandes
Rara
Aragonita
Ausentes
Partes indistintas
cenósteo
do
Presentes em todos
Podem estender além
do cálice
Inserção hexâmera, em
seis comparti-mentos
(Hexacorais)
Geralmente retos
Presente em alguns
Ausentes
Mesozóico-Cenozóico:
Triássico ao Recente
PHYLUM CNIDARIA
Características gerais:
Distribuição geológica (temporal): Ediacarano ao Recente
Grau de complexidade:
Metazoários
Simetria bilateral / birradial
Diploblásticos:
Ectoderma / mesogléia / endoderma
Cavidade do corpo única = ênteron
Boca e ânus não separados
Sistema nervoso presente mas sistemas circulatório e excretório não
Caracterizdos por nematocistos
Solitários / coloniais / polimorfos / pólipos / medusas
Exoesqueleto calcário ou orgânico
Classificação taxonômica:
Filo Cnidaria
Classe Hydrozoa (Ediacarano – Recente)
Alternância de gerações com predominância de fase de pólipo
Exemplo: Obelia (Recente)
Classe Scyphozoa (Ediacarano – Recente)
Alternância de gerações com predominância de fase de medusa
Exemplos: Brooksella (medusa cambriano)
Ou Corumbella werneri ( pólipo? ediacarano do
Brasil)
Classe Anthozoa (Ediacarano – recente)
Pólipos
Subclasse Cerianpatharia (Mioceno a Recente)
Subclasse Octocorallia (Ediacarano a Recente)
Subclasse Zoantharia (Cambriano a Recente)
Ordem Tabulata (Ordoviciano Inferior a Permiano)
Ordem Rugosa (Ordoviciano Médio a Permiano)
Ordem Scleractinia (Triássico Médio a Recente)
CNIDÁRIOS
Desde o Ediacarano:
3 gen de Hydrozoa,
2 gen Scyphozoa,
penas do mar Anthozoa
Vantagens do hábito colonial (Clarkson, 3rd ed, cap. 5):
Reprodução assexuada (clones) permite crescimento rápido tanto em
número de indivíduos como em tamanho da colônia; mais segura porque não
envolve fases larval e juvenil.
Crescimento em qualquer direção é vantajosa quando da competição
por espaço: reptante, folhoso, como lençóis, arbustiforme, em montículos.
Estratégia depende de fatores como iluminação, energia do ambiente,
perímetro de defesa
Esqueletos podem fixar organismos ao substrato, permitem elevar os
indivíduos acima do substrato (vantagem para suspensívoros), protegem.
Permite cooperação entre indivíduos.
Desvantagem:
Baixa adaptabilidade genética a mudanças ambientais (a qual pode ser
superada entrando em fase sexuada de reprodução – soltando gametas).
BIOLOGIA
Mais simples dos verdadeiros metazoários
Simetria radial, biradial ou radiobilateral bilateral
Diploblásticos: Células organizadas em dois tipos de tecidos que
constituem duas camadas distintas
Corpo consiste de uma cavidade simples, o ênteron, onde ocorre a
digestão
Em alguns grupos, esta cavidade apresenta divisórias, designadas
mesentérios, que aumentam a superfície disponível para digestão
Ectoderme (camada externa) e endoderme (camada interna)
São separadas pela mesogléia, uma substância gelatinosa
permeada por uma rede nervosa simples
Todos os membros do filo moderno apresentam células urticantes,
nematocistos
Abertura única que serve tanto de boca como de ânus
Marinhos ou de água doce
Solitários ou coloniais
Exosqueleto orgânico ou de carbonato de cálcio (aragonita ou calcita):
corallum (pl., coralla) = o esqueleto total, solitário ou colonial, coralito =
esqueleto individual dentro da colônia
Taxas de crescimento em corais modernos: Formas ramificadas – até 26
cm/ano; formas compactas em montículos – até 1 cm/ano
DISTRIBUIÇÃO DOS CORAIS ATUAIS
Formadores de recifes (corais coloniais hermatípicos)
Águas marinhas de salinidade normal: 34-36‰ (27-48‰)
Águas rasas e agitadas: melhor desenvolvimento a menos de 20
m, comuns até 50 m, alguns até 120 m
Águas de poucos nutrientes (oligotróficas); microalgas como
endossimbiontes (zooxantelas no endoderme) que fixam C e fornecem
O2 para os hospedeiros
Águas limpas, bem iluminadas,
Águas quentes: ótimo entre 25° e 29°C (sobrevivem entre 16° e
40°C)
Águas tropicais (entre latitudes 30°S e 30°N)
Comumente em ambientes de deposição de carbonato
Tipos de recifes (Darwin e a Beagle):
Franja – Barreira - Atol
Corais que não formam recifes: corais solitários (e coloniais)
anermatípicos
Abundantes até 500 m; mas encontrados até 6.000 m de
profundidade
Temperaturas de 1 a 28°C
Tropical a boreal (Noruega)
Presentes em recifes ou em bancos ou montes em águas mais
profundas de salinidade marinha normal
CICLO DE VIDA
Alguns cnidários apresentam alternância de gerações, com formas muito
distintas ao longo de seu ciclo de vida (polimorfismo):
Organismos fixos – pólipos, com reprodução assexuada por brotamento.
Organismos livres – medusas, com reprodução sexuada
Hydrozoa: geralmente polimórficos, geralmente dominância da
fase de pólipos
Scyphozoa: dominância da fase de medusóides
Anthozoa: somente pólipos
Ancestral: planulóide, diploblástico, radialmente simêtrico, ciliado, livre
Hydrozoa:
Ancestral
Hydrozoa
Scyphozoa
Anthozoa
Planulóide
Polimórfica
Pólipos e medusas
Pólipos
Simetria
Radial
Radial, tetrâmera
Radial, tetrâmera
Bilateral
Ênteron
Simples
Dividido
Ênteron
mesentérios
com
CLASSE ANTHOZOA
Subclasse Zoantharia (corais): corais, anêmonas, gorgônias, penas-domar
Ordem Scleractinia: corais modernos, também conhecidos como
hexacorais, Triássico – Recente, 600 gêneros
Ordem Rugosa (†): corais extintos, os corais rugosos ou
tetracorais, Ordoviciano – Permiano, 800 gêneros
Ordem Tabulata (†): corais extintos, os corais tabulados,
Ordoviciano – Permiano, 280 gêneros
ORDEM TABULATA: Os corais tabulados
Nome derivado da presença, quase sempre marcante, de tábulas
(tabulae) como assoalho dos coralitos
Exclusivamente paleozóicos, surgindo no Ordoviciano Inferior um pouco
antes dos corais rugosos
Septos ausentes ou, quando raramente presentes, são poucos e mal
desenvolvidos
Exclusivamente coloniais, com coralitos pequenos, calcíticos; alguns
capazes de se cimentarem ao substrato
Forma do corallum: eretos ou reptantes
Importantes participantes em recifes paleozóicos, junto com algas
calcárias e estromatoporóides (grupo extinto atribuído às esponjas)
ORDEM RUGOSA: Os corais rugosos (ou tetracorais)
Nome derivado da rugosidade marcante exibida pelos coralla,
interpretada como refletindo flutuações cíclicas no crescimento dos corais
Exclusivamente paleozóicos, surgindo no Ordoviciano Médio um pouco
depois dos corais tabulados
Coloniais e solitários, exosqueletos calcíticos, fortes e densos
Septos bem desenvolvidos, inseridos continuamente, de modo
tetrâmera, ao longo da vida (daí, o nome de tetracorais); epiteca bem
desenvolvida
Simetria radial- “bilateral”. Corallum das formas solitárias comumente
curvo, lembrando chifre de boi (daí, o nome “horn coral” em inglês)
Não foram formadores de recifes embora tenham habitado recifes
paleozóicos
Modos de crescimento sobre substrato: preferência por substratos
moles; incapazes de se fixarem ao fundo
Homeomorfia: convergência morfológica de diversos gêneros
ORDEM SCLERACTINIA: Os corais escleractíneos (ou hexacorais)
Nome significa “raios duros”, em referência ao aspecto dos septos
Exclusivamente pós-paleozóicos, surgindo no Triássico Médio bem
depois da extinção dos corais paleozóicos
Coloniais e solitários, exosqueletos aragoníticos, fortes, mas leves
Septos bem desenvolvidos, inseridos continuamente, de modo
hexâmera, ao longo da vida (daí, o nome hexacorais). Epiteca comumente mal
desenvolvida ou ausente
Simetria
Formadores de recifes ou não (hermatípicos ou anermatípicos). Formas
hermatípicas capazes de se cimentar a substratos duros
ESTRATÉGIAS DE CRESCIMENTO
Em formas solitárias
Em formas coloniais:
Crescimento periférico – conquistar espaço
Crescimento mediano – aumentar altura acima do substrato
DIFERENÇAS ENTRE OS TRÊS GRUPOS DE CORAIS
Morfologia:
Estratégias de crescimento/desenvolvimento:
Escleractíneos:
Inserção de septos
Maior integração entre pólipos
Exosqueleto aragonítico, poroso e leve, mas forte e de
crescimento rápido
Capacidade de se cimentar ao substrato
Importância das zooxantelas?
Rugosos e tabulados:
Participação passiva na construção de recifes paleozóicos
Ambientes de deposição preferidos geralmente não tão
rasos, nem tão turbulentos quanto aqueles habitados pelos
escleractíneos
Origem dos escleractíneos
Aplicabilidade do atualismo a organismos extintos?
Os corais extintos tinham nematocistos?
Os corais extintos tinham zooxantelas?
Delta C-13 dos esqueletos?
Adaptações à exposição ao sol: exosqueletos leves,
epiteca reduzida ou ausente, forma achatada e fina
Ambiente de deposição inferido para os corais extintos
Conclusão (Coates & Jackson, 1944):
Tabulata, sim, podem ter tido zooxantelas
Rugosa, não
USOS DOS CORAIS FÓSSEIS
Paleoambiente: condições físico-químicas do meio
Para todos: marinho, águas relativamente rasas, faixa latitudinal
intertropical
Recifes de escleractíneos
Morfologia vs. parâmetros físico-químicos diversos
Agitação
Profundidade
Taxa de sedimentação
Grau de exposição (emergência)
Platyhelminthes
CARACTERIZAÇÃO
Os membros do filo Platyhelminthes, conhecidos como vermes planos, são
animais de vida livre e parasitas. Aprersentam um achatamento no sentido
dorso-ventral e uma constituição corporal acelomada. Muitos possuem túbulos
protonefridiais e são hermafroditas simultâneos.
CLASSIFICAÇÃO
Os Platyhelminthes de vida livre, membros da classe Turbellaria, são
provavelmente os mais primitivos dentre todos os animais bilaterais. Seu
pequeno tamanho, baixo nível de cefalização, condição acelomada, e a
ausência de um ânus são provavelmente características primitivas. Além do
mais, existem algumas espécies com células epitélio-musculares e um sistema
nervoso na forma de uma rede nervosa.
A grande maioria dos turbelários é marinha, mas existem espécies de água
doce e algumas formas terrestres em ambientes úmidos. Os turbelários são
animais bentônicos, vivendo sobre ou embaixo de pedras, algas e outros
objetos. São membros comuns da fauna intersticial.
A maioria dos turbelàrios move-se inteiramente através de cilios; as espécies
grandes (policladidos) são marcantemente achatados e movem-se através dos
cilios mais ondulação muscular sobre a grande superfície ventral. Sistemas de
glândulas duplas possibilitam a fixação temporária em muitas espécies.
Os turbelários são predadores e saprófagos. A digestão é inicialmente
extracelular e posteriormente intracelular. As espécies pequenas possuem um
intestino simples com formato de saco com uma faringe simples ou bulbosa. As
espécies grandes têm um intestino ramificado e uma faringe plicada,
usualmente tubular.
O muco produzido pelos rabdóides e pelas flândulas epidérmicas desempenha
um papel importante na vida dos turbelários, revestindo o substrato sobre o
qual o animal rasteja e envolvendo a presa. As flândulas auxiliam o
aprisionamento da presa e no ato de engolir.
O pequeno tamanho, o formato achatado e o intestino ramificado (nas formas
maiores) formam sistemas especiais para o transporte interno, troca de gases e
excreção. Os protonefrídios estão presentes em muitos platielmintes e estão
provavelmente envolvidos no balanço do fluido interno e na regulação
osmótica.
Um arranjo radial de quatro pares de cordões nervosos longitudinais é
provavelmente primitivo, e os arranjos com números menores provavelmente
evoluíram através de perda. Ocelos em cálices pigmentares, que podem ser
numerosos, são os principais órgãos sensoriais.
Os turbelários são hermafroditas simultâneos com sistemas reprodutores
adaptados à fertilização interna e à deposição de ovos.
Primitivamente (nível oncóforo), os ovos são endolécitos, a clivagem é espiral e
há uma larva de vida livre. Entretanto, na maioria das espécies oncóforos o
desenvolvimento é direto. Muitos turbelários desenvolveram uma divisão
ovariana de trabalho entre a produção de óvulos e a produção de vitelo (nível
neóforo), levando a ovos ectolécitos. A clivagem espiral foi perdida e o
desenvolvimento é invariavelmente direto.
Duas classes de platielmintes, os Trematoda que contém as fascíolas, e os
Cestoda que contém as tênias, são inteiramente parasitárias. Em contraste
com os turbelários, ambas apresentam um revestimento do corpo não-ciliado,
ou tegumento.
Os trematódeos adultos são parasitas externos ou internos de vertebrados.
Partindo da condição turbelária ancestral, apresentam-se menos modificados
se comparados às tênias. O corpo oval a alongado é achatado e provido de
uma ventosa ou de outros órgãos de fixação. O intestino está presente e, em
algumas espécies (Digenea), a boca anterior está associada a uma segunda
ventosa.
Os trematódeos são hermafroditas, e os seus sistemas reprodutores estão
adaptados à copulação, à fertilização interna, ao desenvolvimento ectolécito e
à formação de cascas do ovo.
Os trematódeos monogêneos são principalmente ectoparasitas de peixes. O
ciclo de vida envolve apenas um hospedeiro para o adulto, e um novo
hospedeiro é atingido através de uma larva de vida livre denominada
oncomiracídio.
Os trematódeos digêneos são endoparasitas e constituem o maior grupo de
platielmintes parasitas. O ciclo de vida envolve de dois a quatro hospedeiros e
um certo número de distintos estágios de desenvolvimento, incluindo dois tipos
de larvas (miracídio e cercária). O hospedeiro primário é invariavelmente um
vertebrado e os caracóis são comumente os hospedeiros intermediários. As
espécies de esquistossomos (Schistosoma) estão entre os grupos de parasitas
mais disseminados e mais danosos para o homem.
Cestóides, ou tênias, são parasitas intestinais de vertebrados. São
estruturalmente mais especializados que as fascíolas, tendo um corpo
composto de um escólex provido de órgãos de fixação, de um colo e de um
estróbilo, o qual consiste de uma cadeia de segmentos (proglótides) que
brotam da região do colo.
O intestino está ausente e o sistema reprodutor, que é algo semelhante ao dos
trematódeos, repete-se em cada segmento.
O filo Mesozoa contém dois pequenos grupos de diminutos endoparasitas de
invertebrados. O grupo que habita órgãos excretores de lulas e polvos é o mais
conhecido.
O corpo ciliado, simples e alongado, não possui os tipos celulares usuais e os
órgãos típicos. Supõe-se que os Mesozoa ou tenham derivado dos
platielmintes através de regressão, ou que sejam representativos de um ramo
de algum metazoário primitivo de vida livre. Os dois grupos de mesozoários
poderiam ter tido origens distintas.
PSEUDOCELOMADOS
Os membros dos fitos Gastrotricha, Rotifera, Nematoda e vários outros
compartilham feições como uma cutícula colágena, tubos adesivos, faringe
muscular, protonefridios, pequeno tamanho e eutelia. Quando está presente
uma cavidade do corpo, como nos nematódeos, ela deriva-se do blastocelo,
embrionário e é denominada pseudocelo.
Estas feições em comum constituem a base pela qual se coloca a maioria
destes grupos no filo Aschelminthes. Entretanto, o filo carece de uma
característica distintiva unificadora, pois em outros aspectos de forma e
estrutura os seus membros-grupos são muitos distintos entre si. O nome
Aschelminthes é, portanto, também utilizado como um nome coletivo comum
ou como superfilo.
Os gastrótricos são habitantes da água doce e do mar, sendo as espécies
marinhas principalmente intersticiais. Eles deslizam sobre o substrato através
de cílios ventrais e ancoram-se temporariamente através de tubos adesivos
que contêm sistemas de glândulas duplas. Supõe-se que o conjunto ciliar
ventral seja primitivo e que os gastrótricos tenham-se derivado a partir de
alguma forma ancestral que tivesse sido unifomiemente ciliada.
Filogeneticamente, os gastrótricos constituem provavelmente um grupo central
dentro do conjunto dos asquelmintes. As características significativas são a
estrutura da cutícula externa, o epitélio monociliado de algumas espécies e o
mioepitélio da faringe.
Os rotíferos são distinguíveis por sua coroa ciliada, pé e mástax. A cutícula não
é externa. Eles são principalmente habitantes de água doce. As espécies
bentônicas nadam intermitentemente por meio da coroa ciliada e fixam-se
temporariamente através das glândulas adesivas que se abrem nas
extremidades dos artelhos do pé. Os rotíferos planctônicos nadam
continuamente e têm um pé reduzido. As peças cuticulares faríngeas que
formam o mástax estão adaptadas para a ação predatória ou para comer
suspensões.
Os nematódeos de vida livre ocorrem em grande número no mar, na água doce
e no solo. Seu corpo cilíndrico, longo e afilado, está adaptado à vida em
pequenos espaços, particularmente nos espaços intersticiais de sedimentos
aquáticos e tapetes de algas. As espécies terrestres vivem em películas de
água que circundam as partículas do solo e são, na verdade, aquáticas.
O cilindro da musculatura longitudinal da parede do corpo, a complexa cutícula
elástica e a pressão hidrostática do fluido no interior do pseudocelo possibilitam
os movimentos ondulatórios que dirigem os nematódeos através dos espaços
intersticiais.
Os nematódeos exibem uma grande variedade de hábitos alimentares, a
maioria dos quais envolve uma faringe muscular para a ingestão. Dentes ou um
estilete estão comumente presentes nos nematódeos carnívoros e herbívoros.
O grande número de nematódeos parasitas inclui muitas espécies de
importância médica e econômica, como os vermes ancilóstomos, ascarídeos,
oxiúros, Trichinella e filarioideos. As espécies parasitas variam muito quanto ao
seu ciclo de vida.
O filo Kinorhyncha é composto de um pequeno número de espécies intersticiais
marinhas, nas quais a cutícula é segmentada e a boca fica na extremidade de
um cone retrátil espinhoso na porção anterior da cabeça.
Os membros do filo Nematomorpha possuem corpo filiforme com comprimento
de até 36 cm. Os adultos de vida livre na maioria das espécies vivem em água
doce ou no solo úmido. Os estágios juvenis são parasitas de artrópodos.
Os membros do filo Acanthocephala são parasitas intestinais de vertebrados
aquáticos e terrestres. O alongado parasita ancora-se à parede intestinal
através de uma probóscide retrátil anterior provida de espinhos recurvados.
Não existe um trato digestivo e o alimento é absorvido através da complexa
parede do corpo. O ciclo de vida requer um crustáceo ou um inseto como
hospedeiro intermediário.
Os Gnathostomulida formam um pequeno filo de animais intersticiais marinhos
alongados, os quais são similares aos platielmintes por possuírem uma
superfície externa ciliada e um intestino sem ânus. O epitélio é monociliado,
embora a região bucal possua um par de mandíbulas e uma placa ventral em
forma de pente.
Os nematódeos e rotíferos que habitam os ambientes desfavoráveis das
películas de água que circundam partículas do solo e musgos são capazes de
notáveis graus de anabiose.
Partenogênese e dois tipos de ovos – ovos de rápida eclosão e ovos
dormentes - são adaptações comuns de animais que vivem em massas
temporárias de água doce. Elas permitem rápido aumento de populações e
sobrevivência durante os períodos de condições adversas. Uma ou ambas as
adaptações são encontradas em algumas espécies de platielmintes,
gastrótricos, rotíferos e crustáceos.
Anelídeos
Anatomia e Fisiologia
O filo annelida é constituído aproximadamente de 8700 espécies,
agrupadas em três classes: Polychaeta, Oligochaeta e Hirudinea e é
constituído por organismos que apresentam corpo mais ou menos cilíndrico e
alongado. A grande diferença entre os vermes anelídeos e os outros vermes
estudados é a organização, do corpo nos primeiros, em segmentos ou anéis,
que se repetem no sentido do comprimento do animal. Cada um dos
segmentos ou anéis que constituem o corpo dos anelídeos é chamado
metâmero. A metamerização geralmente se evidencia tanto nos aspectos
internos com no externos, incluindo músculos, nervos, sistemas circulatórios,
excretor e reprodutor.
A maioria dos anelídeos apresentam o corpo vermiforme; ao longo
deste há pequenas cerdas de natureza quitinosa e de distribuição característica
nas classes onde elas ocorrem; estas cerdas são elementos auxiliares na
locomoção do animais.
O sistema nervoso é constituído por uma cadeia nervosa ganglionar e
ventral formada por um anel nervoso anterior e um par de gânglios nervosos
para cada segmento do corpo de onde partem ramificações nervosas para as
diferentes partes do organismo.
O sistema sensorial é pouco desenvolvido; raramente são encontrados
órgãos visuais; na maioria dos representantes existem apenas células
sensoriais difusas.
O tubo digestivo dos anelídeos é completo: iniciado pela boca; depois a
faringe, que se comunica com o esôfago; depois o intestino e depois o intestino
terminal que geralmente é curto.
O sistema digestivo da minhoca é completo, isto é, possui duas
aberturas. a boca e o ânus. A boca, localizada no primeiro segmento da região
anterior, abre-se sob um lábio musculoso, o prostômio, à boca segue-se uma
faringe curta, ligada à parede do corpo por muitos feixes musculares. Após a
faringe encontra-se a moela que é uma porção dilatada do tubo digestivo; a
moela é muito musculosa e sua função é, graças às contrações de suas
paredes, triturar o alimento que por ali passa. A dieta alimentar da minhoca
consiste em detritos vegetais em decomposição e pequenos microorganismos.
0 alimento juntamente com a terra é ingerido graças à sucção provocada pela
contração dos músculos da faringe. Na moela, ocorre a digestão mecânica do
alimento.
As contrações da moela atritam partículas de terra contra o
alimento, o qual, dessa forma, fica finamente fragmentado. Após a triturarão na
moela, o alimento passa para o intestino, onde enzimas digestivas são
secretadas .A digestão ocorre extracelularmente. Após a digestão extracelular,
o alimento é absorvido. No intestino de Pheretima, mais ou menos na altura do
trigésimo segmento, existem duas expansões laterais: os cecos intestinais. No
intestino, na porção que continua por trás dos cecos, existe uma prega
longitudinal interna, o tiflossole .Ambas as estruturas, cecos e tiflossole, têm o
mesmo significado funcional: aumentar a superfície de contato com o alimento
digerido, fornecendo assim uma maior superfície de absorção. Os restos não
aproveitados são eliminados pelo ânus.
O sistema excretor dos anelídeos é constituído por metanefrídeos,
distribuídos aos pares em cada segmento; estes canais começam dentro de
uma câmara celômica.
O sistema circulatório dos anelídeos é diferente do encontrado nos
moluscos. Nos moluscos, o sangue abandona os vasos e cai em lacunas dos
tecidos, sendo por isso chamado sistema circulatório aberto. Nos anelídeos o
sangue circula sempre dentro dos vasos, sendo um sistema circulatório
fechado.
O sistema circulatório dos anelídeos é totalmente separado da
cavidade do corpo e consiste principalmente em dois vasos sangüíneos
longitudinais; o vaso dorsal é contrátil; pode ocorrer o desenvolvimento de mais
vasos tanto longitudinais como transversais, o que torna o sistema circulatório
mais complexo.
O sistema circulatório da minhoca ‚ composto por dois grandes vasos
que percorrem o animal no sentido do comprimento; um sobre o intestino, o
vaso dorsal e um abaixo do intestino, o vaso ventral. Existe também um vaso
longitudinal menor que estes dois, que corre ventralmente ao sistema nervoso
e é chamado de vaso subneural. Os vasos longitudinais ligam-se entre si
através de 4 grandes vasos circulares situados um antes, um sobre e dois
depois da moela. Esses vasos circulares possuem elevada capacidade de
contração e funcionam como "corações”, que impulsionam o sangue pelo
sistema circulatório.
A respiração dos vermes segmentados é branquial ou cutânea; as
brânquias são órgãos de forma variável, servindo para retirar o oxigênio e
eliminar o gás carbônico nos anelídeos de vida aquática; nos de vida terrestre
as trocas gasosas ocorrem diretamente através da pele.
Em relação ao habitat dos anelídeos podem ser aquáticos, marinhos ou
de água doce e terrestres vivendo em lugares sombrios, debaixo de pedras e
folhas ou escavando galerias no solo, onde passam a viver. Alguns anelídeos
são de importância econômica, como o caso das minhocas, que são usadas
como iscas para a pesca.
A classe Polychaeta é formada de organismos com o corpo alongado e
cilíndrico com segmentações externa e interna bem nítida.
A classe Oligochaeta é formada de organismos com o corpo alongado
e cilíndrico, com segmentações externa e interna bem nítida.
Na classe Oligochaeta estão agrupados organismos de corpo alongado
e fino. 0 nome da classe deve-se ao fato de existirem, nos seus
representantes, poucos apêndices ou cerdas no corpo. (oligos = pouco e
chaete = cerdas). As minhocas comuns são animais terrestres e um gênero
bastante encontrado no Brasil é Pheretima, cujo tamanho é cerca de 10 cm de
comprimento. Lumbricus é um gênero muito freqüente na Europa e América do
Norte e mede cerca de 30 cm de comprimento. Megascolecides australis ‚ uma
minhoca que vive em regiões tropicais, principalmente na Austrália e pode
medir mais de 2 metros de comprimento.
Descreveremos, neste capítulo, a anatomia e fisiologia da minhoca Pheretima,
encontrada facilmente na terra em lugares úmidos.
Observando extremamente um animal adulto, podemos perceber que
seu corpo ‚ formado por 85 a 95 segmentos, todos aproximadamente do
mesmo tamanho. Se numerarmos os segmentos a partir da região anterior,
notaremos que, ocupando os segmentos de número 14,15 e 16, existe uma
região mais dilatada e mais clara que o resto do corpo, o clitelo. A face superior
do animal é normalmente mais escura que a face inferior.
No primeiro segmento localiza-se a boca e, no último, abre-se o ânus.
Externamente, a minhoca é revestida por uma cutícula abaixo da qual esta a
epiderme formada por uma camada de células. Abaixo da epiderme
encontrasse a musculatura do animal. A camada muscular mais externa é
circular ao corpo e a camada muscular mais interna é mais espessa que a
circular, com fibras dispostas no sentido longitudinal. Abaixo da camada
muscular longitudinal, revestindo a cavidade celomática do corpo, encontra-se
um epitélio fino, de origem mesodérmica, o peritônio.
A locomoção da minhoca faz-se através da contração dos músculos
circulares e longitudinais e graças à presença do fluido celomático, que
preenche os metâmeros. e serve de esqueleto hidrostático. Auxiliando os
movimentos da minhoca existe, ao lado de cada metâmero (com exceção do
primeiro e do último) um par de cerdas quitinosas . Essas cerdas, quando o
segmento esta distendido, são recolhidas. No entanto, quando o segmento
esta contraído, as cerdas eriçam-se. Como o deslocamento é efetuado graças
as distensões e contrações que se propagam do primeiro ao último segmento,
as cerdas eriçam-se e recolhem-se alternadamente, apoiando-se as saliências
do terreno e auxiliando a locomoção.
Cada segmento da minhoca, com exceção dos 3 primeiros e do último,
têm pares de estruturas enoveladas, os nefrídios. 0 nefrídio consiste de um
funil ciliado chamado nefróstoma; a este segue-se um túbulo fino que se
ramifica e, na última porção volta a ser único, alargando-se e formando a
vesícula, que se abre ou para o exterior ou para o intestino pelo poro excretor
ou nefridióporo.
Os nefrídios retiram produtos de excreção (amônia, uréia) do líquido
celomático e do sangue. Os excretas do fluido celomático são retirados pelo
nefróstoma graças ao batimento dos cílios. O túbulo nefridial é circundado por
capilares sangüíneos, dos quais retiram os excretas do sangue e os eliminam
pelo nefridiopóro.
A classe Hirudinea é formada de organismos com o corpo de forma
achatada e segmentado, porém a segmentação externa não corresponde à
interna; a maior parte das estrias que limitam externamente os anéis são sulcos
superficiais; os limites de cada segmento interno podem ser percebidos na face
dorsal em virtude da existência das chamadas papilas segmentares.
Uma das características anatômica e funcionalmente importante é a
presença de segmentação do corpo. A segmentação é a presença de
constricções transversais sucessivas, no corpo, os segmentos. A segmentação
pode ser de dois tipos: superficial e metamérica.
A segmentação superficial acontece apenas na ectoderme, atingindo,
portanto, somente a parede do corpo.
O outro tipo de segmentação, a metamérica, tem origem na
mesoderme e não atinge só esta como também muitas partes da ectoderme.
Diversidade e Ecologia
Os anelídeos podem viver em ambientes aquáticos (de água doce ou salgada)
e terrestres. Os oligoquetos tem poucas cerdas, os poliquetos tem muitas
cerdas implantadas nos parapódios e os hirudíneos não tem cerdas. Os
poliquetos podem viver fixos ou explorar o ambiente à procura de alimentos. Os
hirudíneos (sanguessugas) possuem ventosas para fixação e alimentam-se de
sangue.
Os oligoquetos vivem em ambientes aquáticos, principalmente de água
doce ou na terra. Os vermes terrestres necessitam habitar lugares úmidos, pois
sua respiração é feita por difusão de gases pela epiderme, que necessita estar
sempre úmida para permitir a entrada dos gases para os capilares sangüíneos
da parede do corpo. Muitos oligoquetos aquáticos vivem nas margens de rios e
lagos de águas limpas. Outros, como Tubifex, vivem fixos ao fundo de lagos e
rios de águas poluídas, alimentando-se dos detritos orgânicos poluentes.
Reprodução dos anelídeos
A reprodução dos anelídeos pode ser por assexuada por bipartição ou
brotamento, mas a maioria apresenta reprodução sexuada por fecundação
cruzada.
Os anelídeos podem ser monóicos (oligoquetos e hirudíneos) ou dióicos
(a maioria dos poliquetos). O desenvolvimento é direto em oligoquetos e
hirudíneos e indireto nos poliquetos, que apresentam uma forma larval ciliada,
a trocófora.
A minhoca é monóica. Externamente podemos notar na face inferior, 3
pares de orifícios nos segmentos de números 6, 7 e 8. Cada orifício abre-se,
internamente, para um receptáculo seminal. Na região do clitelo existe um par
de orifícios, os poros genitais femininos. Cada um deles abre-se, internamente,
para um oviduto em forma de funil. Na altura do 189 segmento existe ainda um
par de orifícios. Cada orifício é abertura do duto deferente.
O aparelho reprodutor feminino é composto por um par de ovários.
Próximo aos ovários existem dois ovidutos em forma de funil, que abremúse no
par de orifícios do clitelo. Fazem parte também do aparelho reprodutor feminino
os 3 pares de receptáculos seminais, que se abrem nos segmentos de
números 6, 7 e 8.
O aparelho reprodutor masculino é composto por 2 pares de testículos,
cada par situado nos segmentos de números 10 e 15. Próximo a cada testículo
existe um funil espermático. Os dois funis espermáticos de cada lado unem-se,
formando dois ducos deferentes cada um deles, desembocando no poro genital
masculino do 18 metâmero. Cada testículo com seu respectivo funil
espermático esta contido no interior de uma vesícula seminal. Existe também
um par de glândulas prostáticas que desembocam, cada uma em um duto
deferente.
Duas minhocas sexualmente maduras aproximam-se e unem suas
superfícies ventrais, com suas extremidades anteriores opostas. 0 orifício
genital masculino de uma fica em contato com os orifícios dos receptáculos
seminais da outra e vice-versa. Cada um dos copulantes elimina seus
espermatozóides nos receptáculos seminais do outro, onde ficam
armazenados. Ocorre, então, a separação; cada minhoca carrega, agora, os
espermatozóides da outra. Os óvulos amadurecem nos ovários e passam para
o oviduto e são eliminados, através dos poros genitais femininos, em um
casulo, que foi secretado pelo clitelo. 0 casulo, que é um tubo que envolve a
região clitelar, desloca-se para a extremidade anterior do animal. Quando
passa pelos poros dos receptáculos, os espermatozóides do parceiro são
eliminados sobre os óvulos que são fecundados. O casulo continua deslizando,
sai do corpo da minhoca e fecha-se nas extremidades. Mais tarde os ovos
desenvolver-se-ão diretamente em minhocas jovens. Não há estágio larval.
Os vermes poliquetos, podem apresentar formas de reprodução
diferentes da que foi descrita acima. Nereis é um poliqueto marinho que
apresenta sexos separados. As gônadas deste animal não estão presentes
durante toda a vida; aparecem somente na época da reprodução, originando-se
do peritônio mesodérmico. Os óvulos e espermatozóides são eliminados na
água, onde ocorre a fecundação. O ovo desenvolve-se em uma forma larval
ciliada, chamada trocófora. A partir desse estágio larval, forma-se um verme
jovem.