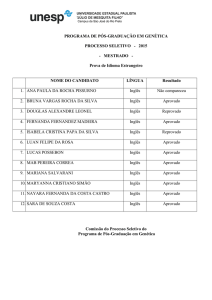
O anticorpo monoclonal contra o antígeno de diagnóstico do Paracoccidioides brasiliensis medeia
proteção inunológica em camundongos BALB/c infectados intratraquealmente com o fungo.
R. Buissa-Filho,1 R. Puccia,2 A. F. Marques,1 F. A. Pinto,1 J. E. Mun˜oz,1,J. D. Nosanchuk,3 L. R. Travassos,2 and C. P. Taborda1*
Institute of Biomedical Sciences, Department of Microbiology, University of Sa˜o Paulo, Sa˜o Paulo, SP, Brazil1; Department of
Microbiology, Immunology and Parasitology, Federal University of Sa˜o Paulo, Sa˜o Paulo, SP, Brazil2; and Departments of Medicine
and Microbiology and Immunology, Albert Einstein College of Medicine, Bronx, New York3
Received 17 March 2008/Returned for modification 8 April 2008/Accepted 24 April 2008
RESUMO: O papel protetor de anticorpos específicos contra o Paracoccidioides brasiliensis é controverso. No presente estudo, foram
analisados os efeitos dos anticorpos monoclonais no antígeno principal de diagnóstico (gp43) utilizando modelos in vitro e in vivo de
infecção por P. brasiliensis. A administração passiva de alguns anticorpos monoclonais (AcM) antes e depois da infecção intratraqueal
ou intravenosa levou a uma diminuição da carga fúngica e diminuiu inflamação pulmonar. A proteção mediada pelo MAB 3E, o MAb
mais eficiente na redução de carga fúngica, foi associado com a fagocitose aumentada de células de leveduras P. brasiliensis por
J774.16, MH-S, ou macrófagos primários. A ingestão de células de leveduras opsonizadas levou a um aumento da produção de NO por
macrófagos. A imunização passiva com MAb 3E induzida aumenta os níveis de interferon gama nos pulmões de ratos infectados. A
reatividade do MAB 3E contra um painel de peptídeos derivados gp43 sugeriu que a seqüência NHVRIPIGWAV contém o resumo de
ligação. O presente trabalho mostra que alguns, mas não todos os AcM contra gp43 podem reduzir a carga fúngica e identifica um
peptídeo novo candidato para o desenvolvimento de vacinas.
INTRODUÇÃO:
Paracoccidioides brasiliensis é um fungo dimórfico térmico que é o agente etiológico da paracoccidioidomicose (PCM),
a micose sistêmica mais prevalente na América Latina que é endêmica em áreas do Brasil, Argentina, Colômbia e
Venezuela. O impacto da doença é demonstrada por Coutinho et al., que relataram que 3.181 casos letais da PCM
ocorreu no período de 1980 a 1995 no Brasil. As formas aguda e subaguda da PCM afetam ambos os sexos e em
primeiro lugar envolvem o sistema reticuloendotelial / linfático. A forma crônica afeta principalmente os homens adultos
com doença pulmonar predominante e/ou envolvimento cutâneo. A ativação do resposta imune celular é o principal
mecanismo eficaz para controlar PCM experimental e humana. Uma correlação foi encontrada entre a severidade da
doença e uma redução da resposta de hipersensibilidade do tipo tardia.
A quimioterapia Antifungica é necessária para o tratamento da PCM, embora mesmo após administração prolongada,
não há garantia da completa destruição do fungo. O período de tratamento depende da droga usada e severidade da
doença. Drogas usadas incluem sulfonamidas, anfotericina B e azólicos.
gp43, primeiramente descrita por Puccia et al., é o principal antígeno diagnóstico de P. brasiliensis usado em uma
variedade de testes sorológicos. Os títulos de anticorpos para gp43 foram usados para monitorar a resposta ao
tratamento em pacientes. gp43 também já mostrou ser imunodominantes como preparado antigênico bruto,
provocando reações de hipersensibilidade tardia em cobaias e humanos, que indicou a presença de T-CD4 reagindo
com epítopos.
O gene da gp43 foi clonado e seqüenciado. Ele codifica um polipeptídeo de 416 aminoácidos (Mr, 45.947), com um
peptídeo líder de 35 resíduos; a proteína madura possui uma única cadeia glicosilada de N-manose. O epítopo H-2
restrito de célula T foi mapeado de um pepitídio 15-mer chamado P10. Diferentes seqüências de 12-mer, todos
contendo o hexapeptídios HTLAIR, induzem a proliferação de células do linfonodo de camundongos sensibilizados pela
gp43 ou infectados com P. brasiliensis. Linfoproliferação induzida por uma P10 ou gp43 envolve Linfócitos CD4 Thelper que produzem interleucina-2 (IL-2) e interferon gama (IFN-γ). A imunização de camundongos quer com gp43 ou
P10 em adjuvante completo de Freund significativamente os protegeu da infecção intratraqueal (IT) com leveduras
virulentas de P. brasiliensis. Após 3 meses de infecção, UFC no pulmão foram 200 vezes menores do que nos
camundongos tratados do controle.
O papel da imunidade mediada por anticorpos na resistência de P. brasiliensis é menos certo. Em vários sistemas, no
entanto, há evidências consideráveis de que a administração de anticorpos monoclonais (AcM) podem modificar o
curso da doença em camundongos infectados com fungos, como Cryptococcus neoformans, Candida albicans,
Histoplasma capsulatum, Pneumocystis spp., Fonsecaea pedrosoi, Aspergillus spp. e, mais recentemente, P.
brasiliensis.
Os mecanismos de ação do anticorpo no combate a uma doença infecciosas incluem a neutralização do antígeno, os
efeitos de cooperação com a imunidade celular pelo aumento da fagocitose ou mediação da citotoxicidade celular,
ativação do complemento, inibição de crescimento, aderência, direta ou biofilme e ação antimicrobiana. Com C.
neoformans, é claro que a imunidade mediada por anticorpos pode ser decisiva para a defesa do hospedeiro. Os
mecanismos envolvidos são complexos e dependentes de vários parâmetros, tais como o isótipo de anticorpo, a função
das células T, cepa do C. neoformans, a quantidade de anticorpos, expressão do NO sintase, a região Fc e ativação do
complemento.
O controle da PCM é classicamente associada a uma vigorosa resposta Th1 e formação de granuloma. Há evidências,
no entanto, que a imunidade mediada por anticorpos podem contribuir para depuração da infecção. No presente
estudo, um painel de AcM contra a gp43 foi utilizado para avaliar o efeito da imunização passiva em camundongos por
via intravenosa (i.v.) ou infectados Intrataquealmente (I.T.) com uma cepa virulenta de P. brasiliensis.
RESULTADOS
In vitro, a fagocitose mediada por anticorpos monoclonais.
Nós investigamos o efeito de AcM para gp43 de P. brasiliensis na fagocitose in vitro utilizando ambos macrófagos
primários (pulmão e peritoneal) e linhagens de células J774.16 e MH-S. As células de levedura opsonizadas com MAbs
19G, 21F, 10D, 17D, e 3E foram pelo menos duas vezes mais internalizada pelas células J774.16 do que pelas não
opsonizadas ou células de leveduras opsonizadas por 32-H (Fig. 1). O mesmo resultado foi obtido com macrófagos
primários. A linhagem de célula MH-S e de macrógafagos primários de pulmão mostraram resultados semelhantes. As
fagocitoses foram observadas, independentemente se os macrófagos foram ou não ativados com IFN-γ, mas os
índices foram significativamente mais elevados em células estimuladas pelas citocinas (dados não mostrados).
FIG. 1. Fagocitose de células de levedura por células J774.16 após 12 h, na presença de anticorpos monoclonais contra a gp43 (3E,
10D, 17D, 19G, 21F e 32H), comparada com a de controles de leveduras incubadas sem MAB (PBS) ou com um anticorpo monoclonal
irrelevante (MAb A4). Cada barra representa a média das três medidas, e as barras de erro indicam SD. Os experimentos foram feitos
em triplicata, e diferentes campos foram contados. Significativa P (0,05, determinada pela análise de variância e Tukey´s) honestamente
teste de diferença) em relação ao controle sem MAB.
As células de levedura opsonizadas com MAbs de gp43 para promover atividade fungicida in vitro em macrógafo.
MAbs que aumentam fagocitose de P. brasiliensis pelos macrófagos também induzem um aumento metabolito de óxido
nítrico (nitrito) nos sobrenadantes das células fagocíticas (Fig. 2).
FIG. 2. Medições Nitrito em sobrenadantes de ensaios de fagocitose. Levedura P. brasiliensis foi cultivada com células J774.16 durante
12h, na presença de anticorpos monoclonais contra a gp43 (3E, 10D, 17D, 19G, 21F e 32H), PBS (sem MAB), ou o MAb irrelevante
(A4). Níveis de nitrito foram detectados utilizando um método de Griess. Diferença significativa (P 0,05, determinada pela análise de
variância e Tukey´s honestamente teste de diferença) em relação para o controle PBS. As barras de erro indicam SD.
A imunização passiva reduz a carga fúngica.
Para avaliar se a administração de MAbs poderia reduzir UFC de pulmão de camundongos infectados com P.
brasiliensis,foram seguidos dois protocolos. No primeiro protocolo, camundongos BALB/ c foram imunizados com
diferentes AcM contra gp43 24h antes da infecção com 3x105 células de levedura do P. brasiliensis. Após 15 dias de
infecção, o número de UFC nos pulmões do ratos que receberam MAbs 19G, 21F, 10D, 3E, mas não 17D e 32H
mostrou uma significativa redução quando comparada com a dos controles (Fig. 3).
FIG. 3. UFC pulmão de camundongos infectados I.T. com 3x105 células de levedura e tratados com anticorpos monoclonais contra a
gp43 (3E, 10D, 17D, 19G, 21F e 32H) 24h antes da infecção. Os ratos foram sacrificados após 15 (barras pretas) ou 30 (barras cinza)
dias de infecção. Controle de ratos foram infectados e receberam PBS (infectados somente) ou um MAb irrelevante (A4). Cada barra
representa a média de contagem de fungos no pulmão, e as barras de erro indicam SD.
Além disso, MAbs 19G, 10D, e 3E mantiveram significativas reduções UFC após 30 dias de infecção (Fig. 3). No
segundo protocolo, foram selecionados dois AcM, 32H, que não reduziu a UFC do pulmão e 3E, que reduziu a UFC.
BALB / c foram infectados, como indicado acima, e após 30 dias, 1mg de MAbs 32H ou 3E foi injetado I.P. As UFC
foram medidos após 45 e 60dias de infecção, e MAb 3E mas não o 32H, foi capaz de reduzir o UFC dos pulmões de
ratos em 60 dias. (Fig. 4)
FIG. 4. UFC pulmão de camundongos infectados I.T. 3 105 com células de levedura e tratados com anticorpos monoclonais contra a
gp43 (3E ou 32H) 30dias após a infecção. Ratos foram sacrificados após 45 (barras pretas) ou 60 (barras cinza) dias de infecção.
Controle de ratos foram infectados e recebeu PBS (infectados somente) ou um irrelevante MAB (A4). Cada barra representa a média de
contagem de fungos no pulmão, e as barras de erro indicam SD.
Para avaliar o efeito da MAbs em camundongos, um modelo de infecção intravenoso foi usado. Grupos de animais
foram passivamente i.p. imunizados com 1 mg de MAb 3E, MAb 32H, ou o controle irrelevante mAb, 24h antes da
infecção com 5x106 células de levedura e receberam 0,1mg de MAbs cada semana,1 mês antes de os animais serem
sacrificados. Para determinações de UFC, o pulmão, baço, fígado e tecidos foram homogeneizados separadamente e
semeadas em meio sólido. Os camundongos imunizados com o MAb 3E teve significativamente UFC mais baixos no
pulmão e baço em comparação com o UFC de ratos tratados com o MAb 32H, o MAb irrelevantes, ou PBS (Fig. 5).
UFC do fígado estavam abaixo do limiar de detecção.
FIG. 5. UFC dos pulmões (barras pretas) ou baço (barras cinzas) de camundongos infectados iv com 3x105 células de levedura e
tratados com 1mg de AcMs contra gp43 (3E ou 32H) 24h antes da infecção e com 100 g cada semana durante um mês como uma dose
de manutenção. Os camundongos foram sacrificados após 30 dias de infecção. Controle de ratos foram infectados e recebeu PBS
(infectados somente) ou um MAb irrelevante (A4). Cada barra representa a média de contagem de fungos no órgão, e as barras de erro
indicam SD.
Viabilidade de células de levedura P. brasiliensis em presença de AcMs 3E e 32H.
A viabilidade das células de leveduras incubadas com anticorpos monoclonais 3E ou 32H ou o MAb irrelevante foi
avaliada durante 30 dias; nenhum efeito inibitório direto pelo MAbs foram observados (dados não mostrados).
Histopatologia do pulmão tratados, infectados I.T. com camundongos BALB/c (30 dias).
Os pulmões dos animais controle não tratados apresentaram infiltração intensa, principalmente por macrófagos,
linfócitos e células epitelióides. Em torno dos focos de granulomas epitelióides, células gigantes também foram
observadas. Células fúngicas em multiplicação foram vistos. Usando o protocolo 1 (no qual os ratos receberam MAbs
24h antes infecção I.T.), observou-se que os animais imunizados com MAb 32H foram patologicamente semelhante ao
camundongos infectados controle. Em contraste, os animais que receberam MAb 3E tinham menos granulomas
epitelióides com um reduzido número de células de levedura e grandes áreas de tecido pulmonar preservado (Fig. 6).
FIG. 6. Histopatologia das secções de pulmão dos ratos infectados de acordo com o protocolo 1 e sacrificados em 30 dias após a
infecção. (A) secção de pulmão de um rato infectada não tratada; (secção B do pulmão) de um rato infectados e tratados com o MAb
32H; (C) seção de pulmão um mouse infectados e tratados com MAb 3E. Hematoxilina-eosina, 10 de ampliação.
Tratamento MAb altera a expressão de citocinas.
Níveis de citocinas foram detectadas no tecido pulmonar de camundongos infectados I.T. e tratados com o MAb 3E e
32H em ambos os protocolos. Grupos de 10 ratos foram utilizadas em cada protocolo, todos os experimentos foram
repetidos duas vezes com resultados semelhantes. Conforme demonstrado na Tabela 1, com o primeiro protocolo, os
animais imunizados com MAb 3E tinham níveis mais elevados de IFN-γ e redução dos níveis de IL-4 após 15 e 30 dias
de infecção em comparação com os dos animais imunizados com MAb 32H. As outras citocinas (IL-10, IL-12 e TNF-α)
apresentaram menor variabilidade. Com o segundo protocolo, os níveis de IL-12 após 45 dias de infecção e IFN-γ após
45 e 60 dias de infecção foram superiores aos do MAB 32H (Tabela 2).
TABELA 1. Protocolo nº 1 os níveis de citocinas em pulmões de ratos em resposta à transferência passiva de anticorpos monoclonais
24h antes infecção com P. brasiliensis.
a Values are means standard deviations of measurements from 10 animals per group. The experiment was repeated twice with reproducible results.
b Statistically significant (P _ 0.05, determined by analysis of variance and Tukey’s honestly significant difference test) relative to the value for the untreated,
infected mice.
TABELA 2. Protocolo nº2 níveis de citocinas em pulmões de ratos em resposta à transferência passiva de anticorpos monoclonais 30
dias após a infecção com P. brasiliensis.
Identificação do epítopo do MAB 3E.
A reatividade dos MAb 3E foi avaliado contra um painel de peptídeos derivados gp43 descrito anteriormente. A análise
dos resultados sugeriu que o epítopo é reconhecido dentro da seqüência NHVRIPIGWAV (dados não mostrados). O
MAB 3E, mas não 32H, reconhece esse peptídeo pelo ELISA (Fig. 7). Sequencia deste peptídeo foi encontrada para
ser conservada na sequências internas de β-1,3-glucanases de Aspergillus fumigatus, Aspergillus oryzae, e Blumeria
graminis.
FIG. 7. Reatividade dos peptídeos NHVRIPIGYWAV (R-CONH2),- R (-COOH) e P10 (QTLAIAHTLAIRYAN) com MAb 3E por ELISA.
Microplacas foram sensibilizadas com 100ng de cada peptídeo, e a reação foi desenvolvida com o MAb 3E. O símbolo do diamante
indica a medida de fundo com tampão sozinho, e os símbolo quadrado corresponde à reatividade dos peptídeos contra o anticorpo
irrelevante.
DISCUSSÃO
O papel protetor de anticorpos contra infecções fúngicas é em muitos casos controversos. Recentemente, vários
estudos têm provado que alguns anticorpos são protetores contra fungos. MAbs ou fragmentos de anticorpos
monoclonais foram aprovados para a avaliação clínica da criptococose e candidíase. Murinos MAb 18B7 dirigidos
contra a cápsula de polissacarídeo de C. neoformans foi utilizada para avaliar a segurança da dose máxima tolerada de
um MAb terapêutico. O estudo utilizou vírus da imunodeficiência humana em pacientes infectados que tinham sido
tratados com sucesso para a meningite criptocócica. A infusão MAb tinha uma meia-vida no soro de aproximadamente
53h, e reduzia o antígeno circulante de fungos. Em outro estudo, um anticorpo monoclonal humano recombinante de
90 proteínas de choque térmico foram utilizadas no tratamento de pacientes com candidíase invasiva. Um consenso
surgiu agora que a incapacidade de soros imunes para mediar proteção contra fungos reflete quantidades inadequadas
de anticorpos protetores e/ou da simultânea presença de anticorpos protetores e falta de proteção ao invés de uma
incapacidade fundamental de anticorpos para proteger contra fungos.
A primeira evidência de um anticorpo mediador protecção contra o P. brasiliensis foi descrito por Mattos Grosso et al..
Os autores mostraram que a transferência passiva de dois anticorpos monoclonais murinos contra uma glicoproteína
de 70kDa, que é reconhecido em 96% dos soros de pacientes com PCM, levou a uma significativa redução do número
de UFC no pulmão e menos granulomas nos órgãos de camundongos infectados experimentalmente. Mais
recentemente, um outro anticorpo monoclonal que reconhece um secretado 75-kDa com atividade da fosfatase inibiu
crescimento in vitro de P. brasiliensis e UFC pulmonar reduzida in vivo).
Embora o MAbs de gp43 não tenham afetado diretamente a crescimento de P. brasiliensis, a ação protetora dos AcMs
foi diretamente associado com a sua capacidade para aumentar a fagocitose. Uma vez que os macrófagos são células
efetoras capazes de exercer o aumento do efeito fungicida, foi investigada a produção de NO em macrófagos que
tinham fagocitado células de levedura MAb-opsonizadas. Os resultados de NO mostraram um aumentoda "protecção"
para todos os AcM, mas a resposta foi variável, com MAb 3E demonstrando um efeito mais impressionante. Dados
prévios do nosso grupo e outros claramente demonstraram que macrófagos ativados matam o fungo in vitro pela
produção de óxido nítrico e hidrogênio peróxido.
Usando um painel de seis AcM contra a gp43, incluindo IgG2a (10D, 17D, 19G e 32H) e IgG2b (3E e 21F), foram
identificadas a protecção e não proteção MAbs pela administração passiva ou i.v. de anticorpos em ratos infectados
com P. brasiliensis. A presença de proteção e de falta de proteção anticorpos contra o antígeno mesmo é
conceitualmente relevante tendo sido descrito previamente para criptococose. A histopatologia das secções do pulmão
de animais infectados I.T. e tratados com proteção MAb mostrou uma redução do número de células de levedura e
granulomas epitelióides em comparação com o controle de ratos não tratados.
O papel de IFN-γ mediando macrófagos ativados em uma infecção fúngica sistêmica tem sido relatada. Macrófagos
peritoneais de murinos ativados por IFN-γ apresentaram melhor atividade fungicida com as leveduras e as formas de
conídios de P. brasiliensis. A produção de IL-10 tem sido associada com doença mais severa em camundongos
suscetíveis. Macrófagos alveolares infecção por I.T de camundongos deficientes de IL-4 controla de forma mais
eficiente crescimento de fungos do que os macrófagos do tipo selvagem, e também ratos IL-4-deficientes aumentaram
os níveis de IFN-γ pulmonar. A administração do MAB passivo irrelevante, 3E, ou 32H em infecção por I.T. do
camundongo BALB/c mostraram um padrão misto Th1/Th2 de ativação. No entanto, as alterações de citocinas
associados com a administração de proteção MAb foram mais como Th1, dado o aumento dos níveis de IFN-γ e IL-12
nos pulmões de camundongos I.T. infectados. Um aumento discreto, mas estatisticamente significativo de IL-10 e uma
redução de IL-4 e TNF-α foram anotados em comparação com os níveis de IL-10, IL-4 e TNF no controle camundongos
infectados. A transferência passiva de MAb 32H induziu menores níveis de IFN no pulmão e IL-12 que tratamento com
MAb 3E. Em doença fúngica, os efeitos do tratamento com anticorpos na produção de citocinas são mal
compreendidos e pode depender de diversas características, incluindo o fungo, em órgãos-alvo, tempo disseminação
dos fungos, e a quantidade de anticorpos, isotipo, e especificidade. Proteção mediada por anticorpo tem sido associada
com menores níveis de IFN-γ em camundongos A/J infectados com C. neoformans, mas ambos Th1 e Th2 de citocinas
relacionadas são necessárias para a proteção de anticorpos para criptococose. Forte respostas Th1 pode resultar em
sobrevivência reduzida e danificam o tecido. Em nossos estudos, descobrimos que o tratamento MAb não têm a
polaridade da resposta Th1/Th2 e que IFN-γ parece ser benéfico.
As tentativas de determinar o resumo da protecção MAB 3E levou à seqüência NHVRIPIGYWAV compartilhada com o
A. fumigatus, A. oryzae e seqüências internas de β-1,3-glucanases B. graminis O efeito opsonizantes do MAB 3E
poderia então envolvem o reconhecimento desta seqüência do peptídeo gp43 acumulado na parede celular do P.
brasiliensis. Na verdade, é gp43 armazenados dentro de um vacúolo, migra para a membrana plasmática, e espalha na
parede celular de P. brasiliensis. A secreção deste antígeno ocorre em locais distintos ao longo da superfície celular.
Historicamente, a proteção contra a PCM tem sido atribuída a uma resposta imune celular vigorosa, enquanto
específica elevada níveis de anticorpos têm sido associados com a severidade da doença, como observado em
pacientes com formas aguda e subaguda. Aqui nós mostram que há falta de proteção e de não ptoteção via anticorpos
anti-gp43 e que desempenham um papel importante no controle da doença. Anticorpos contra gp43 são encontrados
em quase 100% dos pacientes com PCM. Provavelmente, existe uma baixa concentração de anticorpos nas amostras
de soro de desses pacientes, e eles não são suficientes para controlar a doença. Para superar isso, a imunização
passiva com anticorpos monoclonais para gp43 foi utilizada e mostrou-se protetora em modelos do rato com
PCM. O presente trabalho também aumenta o potencial terapêutico uso do peptídeo carregando o epítopo B do MAB
3E no adjuvante ativo de imunização à quimioterapia e / ou em associação com uma seqüência de peptídeo contendo
a epítopos de células T de gp43, conhecido como P10, que provoca uma forte imunidade mediada por célula e
protege contra a infecção experimental.