Gametogênese
A espermatogênese e a oogênese, são os processos de formação do gameta masculino
(espermatozoide) e do gameta feminino (oócito). O espermatozoide e o oócito são
células muito especializadas. Elas contêm a metade do número de cromossomos das
outras células do corpo (as células somáticas). Essa redução no numero de
cromossomos se deve a divisão celular chamada meiose, específica para a formação dos
gametas. A restauração do número de cromossomos (46) ocorre quando essas duas
células se unem na fecundação e formam o zigoto.
A espermatogênese acontece nos testículos, enquanto a oogênese ocorre nos ovários.
Após serem formados, os espermatozóides são armazenados no epidídimo, de onde
podem ser expelidos pela uretra e os oócitos são captados pelas tubas uterinas, local de
encontro dos dois gametas.
Começaremos pela espermatogênese. Ela acontece nos túbulos seminíferos, estrutura
tubular dentro dos testículos. Em um corte histológico dos túbulos seminíferos,
podemos visualizar as células da linhagem espermatogênica em diferentes fases de
desenvolvimento. Na puberdade, por influência da testosterona, inicia-se a
espermatogênese.
Túbulo seminífero:
As espermatogônias são as células mais imaturas e estão localizadas no compartimento
basal do túbulo seminífero. São elas que sofrem sucessivas mitoses e assim, mantém a
espermatogênese ativa por toda vida. Algumas células permanecem como
espermatogônias e outras se diferenciam em espermatócito I. Essa célula sofrerá a
primeira meiose, originando dois espermatócitos II. Em seguida, os espermatócitos II
completam a segunda meiose e originam 4 espermátides (células haploides). As
espermátides passarão por um processo chamado espermiogênese até se transformarem
em espermatozoides, encontrados no lúmen do túbulo. Na espermiogênese não há
divisão celular, apenas um rearranjo das organelas e da forma da célula, adquirindo um
acrossomo, que contém enzimas hidrolíticas, e um flagelo, estruturas importantes para a
fecundação.
Diferentemente da espermatogênese, a ovogênese acontece da puberdade à menopausa.
Milhares de oogônias (análogas às espermatogônias), são formadas na vida intra-uterina
da mulher e se diferenciam em oócitos primários, sendo que ao nascimento, a mulher
tem aproximadamente apenas 300.000 oócitos primários, ou seja, a maioria degenera.
Os oócitos primários estacionam na primeira meiose até a puberdade, quando então, em
cada ciclo, a cada mês, um matura e é liberado do ovário (oocitação ou ovulação). Nesse
processo ele completa a primeira meiose e dá origem ao oócito II e ao primeiro corpo
polar, célula não funcional. A segunda meiose só se completa no momento da
fecundação e dá origem ao oócito que será o gameta fecundado e o segundo corpo polar
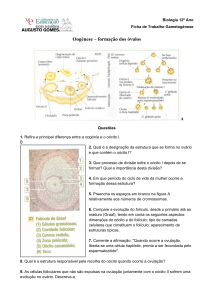
(célula também não funcional). Os oócitos primários se desenvolvem dentro de
estruturas que chamamos de folículos ovarianos. Então, um folículo é o oócito mais as
células foliculares em torno dele. Os folículos sofrem várias modificações até chegar ao
estágio maduro ou de Graff (folículo grande, que faz volume na parede do ovário). É
esse folículo que liberará o ovócito II.
A regulação hormonal é mais complexa no sistema reprodutor feminino do que no
masculino. Em um ciclo de 28 dias, na primeira metade, o FSH estimula a maturação
dos folículos que por sua vez secretam estrogênio. O aumento do estrogênio, causa um
pico de LH que induz a liberação do oócito, a ovulação, e assim o organismo feminino
se prepara para uma possível gestação, com liberação de progesterona também pelo
corpo lúteo, estrutura glandular formada à partir do folículo maduro que liberou o oócito
que poderá ser fecundado.
Fecundação
Fecundação é uma sequencia complexa de eventos coordenados, que se inicia com o
contato dos gametas, oócito e espermatozoide na tuba uterina. Termina com a mistura
dos cromossomos maternos e paternos, assim originando o embrião, unicelular,
diploide, o zigoto.
O espermatozoide, ao chegar ao sistema genital feminino, sofre a capacitação, que é a
perda de glicoproteínas e proteínas que cobrem a superfície do acrossomo, organela em
forma de capuz que contém enzimas que propiciam a fecundação. Os espermatozoides
são quimio-atraídos para a tuba uterina, se tornam hiperativos e fazem contato e
reconhecimento do oócito, o que propicia a reação acrossômica. A reação acrossômica é
a liberação das enzimas do acrossoma, por formação de poros entre a membrana
plasmática do espermatozoide e do acrossomo. A liberação dessas enzimas permite que
o espermatozoide penetre a corona radiata, que são células foliculares ao redor do
oócito. Também propicia a penetração na zona pelúcida, camada glicoproteica ao redor
do oócito, entre a corona radiata e o oócito, então, há a fusão das membranas
plasmáticas do oócito com o espermatozoide, com a entrada do núcleo do
espermatozoide no oócito, deixando para trás seu flagelo e suas mitocôndrias.
Com a entrada do núcleo do espermatozoide, ocorre o término da segunda divisão
meiótica do ovócito, formação do pronúcleo feminino e do segundo corpo polar, célula
não funcional. Forma-se o pronúcleo masculino, e, então, o zigoto. À medida que o
zigoto passa ao longo da tuba em direção ao útero, sofre as clivagens, também
chamadas de segmentação: uma série de divisões mitóticas que formam várias células
menores, os blastômeros. Aproximadamente 3 dias após a fecundação, uma massa de 12
ou mais blastômeros, a mórula, entra no útero. Reparem que a zona pelúcida ainda
permanece, impedindo o crescimento em tamanho dessa estrutura. Uma cavidade se
forma na mórula, chamando-se agora de blastocisto. O blastocisto perde a zona pelúcida
e irá começar o processo de implantação ou nidação no endométrio uterino.
A fecundação 1) reestabelece o número diplóide de cromossomos; 2) determina o sexo
do embrião (por parte do cromossomo X ou Y do espermatozoide), que combinado ao
oócito X, originará XX: feminino ou XY: masculino; 3) proporciona a variabilidade da
espécie pela combinação de cromossomos maternos e paternos; 4) produz a ativação do
zigoto: início da segmentação (clivagens).
Apenas 1 espermatozoide penetra o oócito e o fecunda, embora muitos tentem. Se mais
de um espermatozoide participar da fecundação, o zigoto terá um conjunto extra de
cromossomos, uma triplodia, que é responsável pela maioria dos abortos espontâneos.
No final da primeira semana após a fecundação, o blastocisto, formado a partir das
sucessivas mitoses do zigoto, encontra-se solto na cavidade uterina e é formado pelo 1)
trofoblasto, trofo significa nutrição, porque ele será a contribuição embrionária para a
formação da placenta e 2) um aglomerado de células, a massa celular interna, ou
embrioblasto, que formará o embrião propriamente dito.
No início da segunda semana o blastocisto está parcialmente incrustado no endométrio.
O trofoblasto diferencia-se em 2 camadas, a mais interna, o citotrofoblasto, cito porque
as células mantêm sua identidade e o sinciciotrofoblasto, sincício porque as células
perdem sua identidade, e atuam como uma massa multinucleada que invade o tecido
materno.
Normalmente a implantação do blastocisto ocorre na parede posterior do útero, porém,
algumas vezes o blastocisto pode se implantar fora do útero, são as gestações ectópicas.
A mais frequente entre elas é na tuba uterina, podendo ocorrer fora do útero, no abdome
ou até no ovário. Esses locais são inviáveis de sustentar uma gestação.
Com aproximadamente 8 dias, o blastocisto está parcialmente implantado e a massa
celular interna se segrega em duas camadas embrionárias: o epiblasto, voltado para a
região dorsal e o hipoblasto, voltado para a região ventral. Forma-se uma cavidade, a
cavidade amniótica – acima do epiblasto, que se enche de líquido, o líquido amniótico.
Temos então, o disco embrionário bilaminar.
Aos 9 dias de desenvolvimento o hipoblasto formará uma camada de células achatadas,
que reveste o saco vitelino primitivo, e um tecido conjuntivo frouxo chamado
mesoderma extraembrionária, que se interpõe entre o trofoblasto de um lado, e o saco
amniótico e o saco vitelino, de outro.
Ao final da segunda semana, formam-se vacúolos no mesoderma extraembrionário,
formando a cavidade coriônica.
A segunda semana de desenvolvimento é denominada de semana dos 2: o embrioblasto
forma dois folhetos embrionários: o epiblasto e o hipoblasto. Formam-se duas
cavidades, a cavidade amniótica e o saco vitelino. O trofoblasto, agora chamado córion,
se diferencia em duas camadas o citotrofoblasto e o sinciciotrofoblasto.
Terceira e quarta semanas
A terceira semana caracteriza-se pela gastrulação, que é o estabelecimento dos 3
folhetos germinativos que darão origem a todos os tecidos do embrião. Esses folhetos se
dispõem em camadas, o ecto, o meso e o endoderma. Eles se formam a partir do
epiblasto do embrião didérmico. Olhando o disco didérmico de cima em que foi retirado
o saco amniótico, vemos a formação da linha primitiva no epiblasto. Posteriormente,
por um mecanismo de invaginação celular, a linha primitiva se aprofunda e se
transforma em um sulco primitivo até uma depressão, chamada fosseta primitiva.O
sulco primitivo aparece, então, como consequência de deslocamentos celulares que, em
conjunto, caracterizam a etapa de gastrulação.
À medida que as células epiblásticas se introduzem na linha primitiva entre as células
do ecto e as células do hipoblasto, as células do ecto substituem as do hipoblasto,
formando a endoderme. Posteriormente, outras células epiblásticas continuam
ingressando pela linha primitiva em sentido lateral e se introduzem entre o epiblasto
(agora denominado ectoderma) e o endoderma para formar uma camada intermediária,
chamada mesoderma intraembrionária.
Assim, as primeiras células que entram pela linha primitiva substituem as células do
hipoblasto e formam a endoderme, e as ondas celulares posteriores formam a camada
mesodérmica.
Temos então, a formação do embrião tridérmico, originado pelo processo de gastrulação
Posteriormente, ocorre a formação da notocorda, que se dá pela formação de um tubo
celular rígido no mesoderma, que cresce em direção cefálica. A notocorda é uma
estrutura maciça, com forma de barra, que é transitória no ser humano. Ela tem um
papel muito importante na indução do desenvolvimento de outras estruturas
embrionárias, como o SN.
Em torno dos 20 dias há um engrossamento da região medial da ectoderme
(neuroectoderme) provocado por sinais moleculares da notocorda que induzem a
formação da placa neural.
As células ectodérmicas da região medial do disco embrionário aumentam em altura, e
as células ectodérmicas destinadas a formar a epiderme da pele permanecem achatadas.
Como consequência da mudança de forma desta população celular na parte média da
placa neural, ocorre uma invaginação sobre seu eixo longitudinal e a consequente
formação do tubo neural ou sulco neural, limitado lateralmente pelas pregas neurais
As bordas do tubo neural (pregas neurais), por volta do final da terceira semana
aproximam-se entre si e se fundem. O tubo neural é o primórdio do sistema nervoso
central.
O fechamento do tubo neural começa no meio do embrião e avança de forma
progressiva para as extremidades cranial e caudal.
Um grupo de células da neuroectoderme, correspondentes à zona das pregas neurais, são
segregadas do tubo neural e iniciam uma nova etapa migratória: são as células da crista
neural. Essas células se diferenciarão no sistema nervoso periférico, entre outros.