JÉSSICA FERNANDES KASEKER
FORMAS DE POTÁSSIO EM SOLOS DO SUL DO BRASIL E
SUA ABSORÇÃO POR MUDAS DE PINUS, INFLUENCIADAS
PELA CALAGEM
Tese apresentada como requisito parcial para
obtenção do título de Doutor no Curso de
Pós-graduação em Ciência do Solo da
Universidade do Estado de Santa Catarina UDESC
Orientador: Dr. Paulo Roberto Ernani
Co-orientadores: Dr. Jaime Antônio Almeida
Dr. Luciano Colpo Gatiboni
Lages, SC
2016
Kaseker, Jéssica Fernandes
Formas de potássio em solos do Sul do Brasil
e sua absorção por mudas de pinus, influenciadas
pela calagem / Jéssica Fernandes Kaseker. –
Lages, 2016.
109 p.
Orientador: Paulo Roberto Ernani
Inclui bibliografia
Tese (doutorado) – Universidade do Estado de
Santa Catarina, Centro de Ciências
Agroveterinárias, Programa de Pós-Graduação em
Ciência do Solo, Lages, 2016.
1. pinus 2. rizosfera 3. Solução 4. Nãotrocável I. Kaseker, Jéssica Fernandes. II.
Ernani, Paulo Roberto. III. Universidade do
Estado de Santa Catarina. Programa de PósGraduação em Ciência do Solo. IV. Título
Ficha catalográfica elaborada pelo aluno.
JÉSSICA FERNANDES KASEKER
FORMAS DE POTÁSSIO EM SOLOS DO SUL DO BRASIL E
SUA ABSORÇÃO POR MUDAS DE PINUS, INFLUENCIADAS
PELA CALAGEM
Tese apresentada como requisito parcial para obtenção do grau de Doutor
no Curso de Pós-Graduação em Ciência do Solo da Universidade do
Estado de Santa Catarina – UDESC.
Banca Examinadora:
Orientador:_______________________________________
Professor Dr. Paulo Roberto Ernani
Universidade do Estado de Santa Catarina
Membro: _______________________________________
Professora Dra. Carla Maria Pandolfo
Universidade do Oeste de Santa Catarina
Membro: _______________________________________
Dra. Marlise Nara Ciotta
EPAGRI - São Joaquim/SC
Membro: ___________________________________________
Professor Dr. Álvaro Luiz Mafra
Universidade do Estado de Santa Catarina
Membro: ___________________________________________
Professor Dr. Paulo Cézar Cassol
Universidade do Estado de Santa Catarina
Lages, SC, 26 de agosto de 2016.
AGRADECIMENTOS
À Deus, por ter me dado forças para chegar ao final de
mais esta etapa.
Ao meu orientador Paulo Roberto Ernani, pelos
ensinamentos, apoio e compreensão durante os anos de estudo.
Aos bolsistas e colegas que auxiliaram no
desenvolvimento deste trabalho.
Ao professor José Stape e à North Carolina State
University, por me receberem durante o período de Doutorado
Sanduíche.
À minha família e amigos, por todo o apoio e incentivo.
À Capes pela concessão da bolsa de estudos.
RESUMO
KASEKER, J.F. Formas de potássio em solos do sul do Brasil e
sua absorção por mudas de pinus, influenciadas pela
calagem. 2016. 109 p. Tese (Doutorado em Ciência do Solo) Universidade do Estado de Santa Catarina – Programa de Pós graduação
em Ciência do Solo, Lages, 2016.
O potássio é um nutriente requerido em grandes quantidades
pelas culturas. No solo, pode estar nas formas estrutural, não
trocável, trocável e em solução. Todas elas se encontram em
equilíbrio químico dinâmico com a solução do solo, sendo
passível a liberação das reservas quando as formas mais
prontamente disponíveis são removidas. O objetivo deste
trabalho foi avaliar a distribuição das formas de K em diferentes
valores de pH, em cinco solos. Os experimentos foram
conduzidos em Lages, SC, de 2013 a 2015, sendo utilizados:
Latossolo Bruno, Nitossolo Vermelho, Argissolo Amarelo e
dois Cambissolos Húmicos, um destes proveniente de um
plantio de pinus de quarta rotação. No primeiro experimento,
utilizou-se um fatorial, com doses de calcário e aplicação ou não
de potássio. Foram utilizadas 0, 0,25, 0,5, 1,0 e 1,5 vezes a
quantidade de calcário indicada pelo índice SMP para elevar o
pH até 6,0, combinada ou não com a adição de 1000 mg kg-1 de
K. Houve elevação do pH com o aumento das doses de calcário
em todos os solos, porém a adição de K resultou em valores de
pH ligeiramente menores do que nos tratamentos sem a adição
de K. Os teores de K na solução dos solos foram os que
demonstraram mais expressivamente os efeitos da adição de
doses de calcário, diminuindo em todos os solos,
independentemente da adição ou não de K. Houve também
diminuição do K extraível em todos os solos exceto no
Argissolo. Os teores de K não trocável foram os menos afetados,
variando somente no Argissolo e no Nitossolo na ausência da
adição de K. No segundo experimento, os solos foram utilizados
para cultivar uma muda de Pinus taeda, por dois cultivos
consecutivos. Os tratamentos consistiram na aplicação de 50 mg
kg-1 de K, aplicação de calcário em quantidade para elevar o pH
a 6,0, aplicação de calcário e K, além de um tratamento
testemunha. O solo rizosférico foi avaliado separadamente do
restante do volume de solo. A adubação potássica aumentou os
teores de K nas diferentes formas no solo e na planta, porém não
afetou o desenvolvimento inicial das mudas de pinus. A adição
de calcário, combinada ou não com K, prejudicou o crescimento
das mudas em algumas classes de solo. Em geral, a rizosfera
diferiu do restante do volume de solo, sendo a diferença bastante
variável para cada solo. Não foi possível concluir sobre a
contribuição do K não trocável na nutrição do pinus em função
dos altos teores trocáveis iniciais.
Palavras-chave: pinus, rizosfera, solução, não-trocável.
ABSTRACT
KASEKER, J.F. Forms of potassium in southern Brazilian soils and
effect of liming on K uptake by pine seedlings. 2016. 109 p. Thesis
(Doctorate in Soil Science) - Santa Catarina State University – Post
Graduation Program in Soil Science, Lages, 2016.
Potassium is a nutrient required in large quantities by crops. On
soils, it may be in the structural, non-exchangeable and
exchangeable forms. All these forms are in dynamic chemical
equilibrium with K in the soil solution and K is released from
the reserves when the most readily available forms are removed.
The objective of this study was to evaluate the dynamics
between the K forms as affected by liming and K addition in five
soils. The experiments were conducted in Lages, SC, from 2015
to 2015, using: an Oxisol, a Nitosol, an Argisol and two
Cambisols, being one from a fourth rotation pine plantation area.
In the first experiment, a factorial including rates of limestone
and addition or not of K was used. Liming levels were 0, 0.25,
0.5, 1.0 and 1.5 times the amount of lime indicated by the SMP
method to raise soil pH to 6.0, combined or not with 1000 mg
kg-1 K. Soil pH increased with the increase of lime application
on all soils, but the addition of K slightly decreased pH values.
Solution K was the most sensitive K form affected by liming,
which decreased in all soils, regardless of K addition.
Exchangeable K also decreased in all soils except on Argisol.
The non-exchangeable K levels were the least affected, varying
only in Argisol and Nitosol in the absence of K addition. In the
second experiment, the soils were used to grow a seedling of
Pinus taeda for two consecutive growing seasons. Treatments
consisted in the application of 50 mg kg-1 K, liming, in a quantity
to raise the pH to 6.0, liming + K, and a control treatment.
Rhizosphere soil was evaluated separately from bulk soil.
Potassium fertilization increased K contents in different forms
in the soil and in the plant, but did not affect the early
development of pine seedlings. Liming, combined or not with K,
negatively affected the growth of seedlings in some soil types.
In general, rhizosphere soil differs from bulk soil with quite
variable difference for each soil. It was not possible to conclude
on non-exchangeable K contribution in pine nutrition due to the
high initial exchangeable K content.
Keywords: pine, rhizosphere, solution, non-exchangeable.
LISTA DE TABELAS
Tabela 1.1Tabela 1.2 –
Tabela 2.1 –
Tabela 2.2 –
Tabela 2.3 –
Tabela 2.4 -
Tabela 2.5 -
Tabela 2.6 -
Caracterização química e física dos solos antes
da implantação do experimento.......................26
Dose equivalente de calcário aplicada em cada
tratamento para cada um dos solos..................27
Caracterização química e física dos solos antes
da implantação de experimento.......................63
Altura, diâmetro de mudas e massa seca de raiz
e parte aérea de mudas de pinus no primeiro
cultivo, nos tratamentos testemunha, aplicação
de potássio, aplicação de calcário e calcário mais
potássio avaliados para cada classe de solo.....66
Altura, diâmetro de mudas e massa seca de raiz
e parte aérea de mudas de pinus no segundo
cultivo, nos tratamentos testemunha, aplicação
de potássio, aplicação de calcário e calcário mais
potássio avaliados para cada classe de solo.....69
Valores de pH e teores de Al (cmolc dm-3) nas
porções rizosférica (R) e não rizosférica (NR) do
solo após o primeiro cultivo de pinus nos
tratamentos testemunha, aplicação de potássio
(K), aplicação de calcário e calcário mais
potássio avaliados para cada classe de solo.....71
Valores de pH e teores de Al (cmolc dm-3) nas
porções rizosférica (R) e não rizosférica (NR) do
solo após o segundo cultivo de pinus nos
tratamentos testemunha, aplicação de potássio
(K), aplicação de calcário e calcário mais
potássio avaliados para cada classe de solo.....73
Teores de Ca e Mg (cmolc dm-3) nas porções
rizosférica (R) e não rizosférica (NR) do solo
após o primeiro cultivo de pinus nos tratamentos
testemunha, aplicação de potássio (K), aplicação
Tabela 2.7 -
Tabela 2.8 –
Tabela 2.9 -
Tabela 2.10 -
Tabela 2.11–
Tabela 2.12 -
de calcário e calcário mais potássio avaliados
para cada classe de solo...................................75
Teores de Ca e Mg (cmolc dm-3) nas porções
rizosférica e não rizosférica do solo após o
segundo cultivo de pinus nos tratamentos
testemunha, aplicação de potássio, aplicação de
calcário e calcário mais potássio avaliados para
cada classe de solo...........................................77
Valores de K na solução nas porções rizosférica
e não rizosférica do solo após o primeiro cultivo
de pinus nos tratamentos testemunha, aplicação
de potássio, aplicação de calcário e calcário mais
potássio avaliados para cada classe de solo.....79
Valores de K extraível (Mehlich 1) nas porções
rizosférica e não rizosférica do solo após o
primeiro cultivo de pinus nos tratamentos
testemunha, aplicação de potássio, aplicação de
calcário e calcário mais potássio avaliados para
cada classe de solo...........................................80
Valores de K não trocável (HNO3) (mg kg-1) nas
porções rizosférica e não rizosférica do solo
após o primeiro cultivo de pinus nos tratamentos
testemunha, aplicação de potássio, aplicação de
calcário e calcário mais potássio avaliados para
cada classe de solo...........................................81
Valores de K na solução nas porções rizosférica
e não rizosférica do solo após o segundo cultivo
de pinus nos tratamentos testemunha, aplicação
de potássio, aplicação de calcário e calcário mais
potássio avaliados para cada classe de solo.....82
Valores de K extraível (Mehlich 1) nas porções
rizosférica e não rizosférica do solo após o
segundo cultivo de pinus nos tratamentos
testemunha, aplicação de potássio, aplicação de
Tabela 2.13 -
Tabela 2.14 -
Tabela 2.15-
Tabela 2.16-
Tabela 2.17-
calcário e calcário mais potássio avaliados para
cada classe de solo...........................................83
Valores de K não trocável (HNO3) (mg kg-1) nas
porções rizosférica e não rizosférica do solo
após o segundo cultivo de pinus nos tratamentos
testemunha, aplicação de potássio, aplicação de
calcário e calcário mais potássio avaliados para
cada classe de solo...........................................84
Teores de K, Ca e Mg nas acículas do primeiro
cultivo de pinus nos tratamentos testemunha,
aplicação de potássio, aplicação de calcário e
calcário mais potássio avaliados para cada classe
de solo.............................................................87
Teores de K, Ca e Mg no caule das mudas do
primeiro cultivo de pinus nos tratamentos
testemunha, aplicação de potássio, aplicação de
calcário e calcário mais potássio avaliados para
cada classe de solo...........................................88
Teores de K, Ca e Mg nas acículas do segundo
cultivo de pinus nos tratamentos testemunha,
aplicação de potássio, aplicação de calcário e
calcário mais potássio avaliados para cada classe
de solo.............................................................89
Teores de K, Ca e Mg no caule das mudas do
segundo cultivo de pinus nos tratamentos
testemunha, aplicação de potássio, aplicação de
calcário e calcário mais potássio avaliados para
cada classe de solo...........................................90
LISTA DE FIGURAS
Figura 1.1 –
Figura 1.2 –
Figura 1.3 –
Figura 1.4 –
Figura 1.5 –
Difratogramas da fração argila do Nitossolo
(NT) de amostras tratadas com saturação de K
(A) nas temperaturas de 25ºC (AK25), 100ºC
(AK100), 350ºC (AK350) e 550ºC (AK550), e
de amostras tratadas com saturação de Mg (B)
e Mg + etileno glicol (AMGEG)...................30
Difratogramas da fração argila de amostras do
Latossolo (LT) tratadas com saturação de K
(A) nas temperaturas de 25ºC (AK25), 100ºC
(AK100), 350ºC (AK350) e 550ºC (AK550), e
de amostras tratadas com saturação de Mg (B)
e Mg + etileno glicol (AMGEG)...................31
Difratogramas da fração argila do Cambissolo
(CB) de amostras tratadas com saturação de K
(A) nas temperaturas de 25ºC (AK25), 100ºC
(AK100), 350ºC (AK350) e 550ºC (AK550), e
de amostras tratadas com saturação de Mg (B)
e Mg + etileno glicol (AMGEG)...................34
Difratogramas da fração argila do Cambissolo
sob cultivo de pinus (CP) de amostras tratadas
com saturação de K (A) nas temperaturas de
25ºC (AK25), 100ºC (AK100), 350ºC
(AK350) e 550ºC (AK550), e de amostras
tratadas com saturação de Mg (B) e Mg +
etileno glicol (AMGEG)...............................35
Difratogramas da fração argila do Argissolo
(AG) de amostras tratadas com saturação de K
(A) nas temperaturas de 25ºC (AK25), 100ºC
(AK100), 350ºC (AK350) e 550ºC (AK550), e
de amostras tratadas com saturação de Mg (B)
e Mg + etileno glicol (AMGEG)...................37
Figura 1.6 –
Figura 1.7 –
Figura 1.8 –
Figura 1.9 –
Figura 1.10 –
Figura 1.11 –
Figura 1.12 –
Figura 1.13 –
Valores de pH no Nitossolo (A) e no Latossolo
(B) em função da adição de doses de calcário,
na presença ou não de adição de 1000 mg kg-1
de K..............................................................39
Valores de pH no Cambissolo (A) e no
Cambissolo sob cultivo de pinus (B) em
função do aumento das doses de calcário, na
presença ou não de adição de 1000 mg kg-1 de
K...................................................................40
Valores de pH no Argissolo em função do
aumento das doses de calcário, na presença ou
não de adição de 1000 mg kg-1 de
K...................................................................41
Teor de K na solução no Nitossolo (A) e no
Latossolo (B) em função do aumento das doses
de calcário, na presença ou não de adição de
1000 mg kg-1 de K.........................................43
Teor de K na solução no Cambissolo (A) e no
Cambissolo sob cultivo de pinus (B) em
função do aumento das doses de calcário, na
presença ou não de adição de 1000 mg kg-1 de
K...................................................................44
Teor de K na solução no Argissolo em função
do aumento das doses de calcário, na presença
ou não de adição de 1000 mg kg-1 de K.........45
Teor de K extraível no Nitossolo (A) e no
Latossolo (B) em função do aumento das doses
de calcário, na presença ou não de adição de
1000 mg kg-1 de K.........................................47
Teor de K trocável no Cambissolo (A) e no
Cambissolo sob cultivo de pinus (B) em
função do aumento das doses de calcário, na
presença ou não de adição de 1000 mg kg-1 de
K...................................................................48
Figura 1.14 –
Figura 1.15 –
Figura 1.16 –
Figura 1.17 –
Teor de K extraível no Argissolo em função do
aumento das doses de calcário, na presença ou
não de adição de 1000 mg kg-1 de K..............49
Teor de K não trocável no Nitossolo (A) e no
Latossolo (B) em função do aumento das doses
de calcário, na presença ou não de adição de
1000 mg kg-1 de K.........................................50
Teor de K não trocável no Cambissolo (A) e
Cambissolo sob cultivo de pinus (B) em
função do aumento das doses de calcário, na
presença ou não de adição de 1000 mg kg-1 de
K...................................................................51
Teor de K não trocável no Argissolo em função
do aumento das doses de calcário, na presença
ou não de adição de 1000 mg kg-1 de K.........52
SUMÁRIO
1.1
1.2
1.3
1.3.1
1.3.2
1.3.3
1.4
1.5
2.1
2.2
2.3
2.3.1
2.3.2
2.4
2.5
3
INTRODUÇÃO GERAL..................................
21
CAPÍTULO I – DISTRIBUIÇÃO DAS
FORMAS
DE
POTÁSSIO
EM
DECORRÊNCIA DA APLICAÇÃO DE
DOSES DE CALCÁRIO...................................
23
INTRODUÇÃO.................................................
23
MATERIAL E MÉTODOS..............................
25
RESULTADOS E DISCUSSÃO......................
29
ANÁLISES MINERALÓGICAS.......................
29
pH........................................................................
38
TEORES DE POTÁSSIO...................................
41
CONCLUSÃO...................................................
54
REFERÊNCIAS BIBLIOGRÁFICAS............
55
CAPÍTULO II – FORMAS DE POTÁSSIO
EM SOLOS CATARINENSES E SUA
UTILIZAÇÃO POR PLANTAS DE PINUS...
59
INTRODUÇÃO.................................................. 59
MATERIAL E MÉTODOS............................... 62
RESULTADOS E DISCUSSÃO........................ 65
TEORES NO SOLO............................................. 69
TEORES NA PLANTA........................................ 86
CONCLUSÕES................................................... 93
REFERÊNCIAS BIBLIOGRÁFICAS............
94
CONSIDERAÇÕES FINAIS............................. 104
ANEXOS............................................................ 105
21
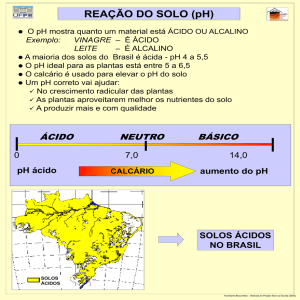
INTRODUÇÃO GERAL
O cultivo de espécies do gênero pinus é bastante
utilizado no país, especialmente na região sul. Em muitos casos,
os povoamentos são estabelecidos em áreas de baixa fertilidade,
que seriam inapropriadas para o cultivo de outras espécies, e
ainda assim apresentam rápido crescimento e boa produtividade,
sem a necessidade do aumento dos teores de nutrientes no solo.
Os estudos de adubação realizados com pinus são
inconclusivos, sendo comum a falta de resposta à fertilização
nos parâmetros de crescimento e nos teores foliares. A ausência
de resposta em experimentos de adubação e os níveis adequados
de crescimento de plantas de pinus em solos de baixa fertilidade
indicam que estas plantas podem usar formas não prontamente
disponíveis dos nutrientes presentes no solo. As formas
estruturais e não-trocáveis podem contribuir para a nutrição das
plantas, justificando o crescimento satisfatório.
Dentre os nutrientes essenciais ao desenvolvimento
vegetal, o K é um dos requeridos em maior quantidade e que está
presente no solo predominantemente na forma estrutural. Além
disso, é um nutriente ao qual muitas vezes não se visualiza
resposta à adubação em diversas culturas, incluindo o pinus,
indicando que este nutriente está sendo suprido a partir de fontes
não prontamente disponíveis. Assim, é importante avaliar a
relação entre a disponibilidade das formas não trocáveis deste
nutriente com o crescimento vegetal.
A liberação destas formas depende da mineralogia dos
solos e também de outros fatores químicos, como pH e
concentração eletrolítica, que alteram a dinâmica dos nutrientes
no solo. Assim, outras práticas de manejo, como a calagem,
podem interferir diretamente na disponibilidade do nutriente no
solo, considerando que as variações no pH podem aumentar a
adsorção de cátions no solo e acelerar a dissolução de minerais,
22
sendo importante avaliar a interação entre esses fatores para
otimizar as práticas de adubação.
Outro fator que altera a velocidade de liberação de
formas não trocáveis é a interação solo-planta. Superfícies
vegetadas apresentam maiores taxas de intemperismo se
comparadas com áreas não vegetadas, devido à liberação de
ácidos e compostos orgânicos das plantas através das raízes.
Assim, a interface de contato das raízes com o solo, a rizosfera,
pode apresentar composição química diferenciada em relação ao
restante do volume de solo, sendo importante o estudo desta
fração do solo para melhor entendimento dos processos de
absorção dos nutrientes.
Conhecendo a dinâmica do nutriente nos solos com
diferenciada composição mineralógica, é possível otimizar o
manejo dos povoamentos florestais e corrigir os níveis de
fertilidade, quando necessário, para obter maior produtividade.
O objetivo deste trabalho foi avaliar a distribuição das
formas de K não trocável, extraível e na solução em cinco solos
influenciada pela adição de potássio e doses de calcário, bem
como a contribuição destas formas na nutrição e crescimento de
mudas de pinus.
23
CAPÍTULO I – DISTRIBUIÇÃO DAS FORMAS DE
POTÁSSIO EM DECORRÊNCIA DA APLICAÇÃO DE
DOSES DE CALCÁRIO
1.1
INTRODUÇÃO
O potássio é um nutriente requerido em grandes
quantidades pelas culturas. No solo, pode estar nas seguintes
formas: estrutural, fazendo parte de minerais primários; não
trocável, onde o K está fixado nas entrecamadas dos
argilominerais
do
tipo
2:1;
trocável,
adsorvido
eletrostaticamente às cargas elétricas negativas; e livre na
solução. A forma trocável é a que normalmente controla a
disponibilidade de K na solução do solo. Todas essas formas,
entretanto, se encontram em equilíbrio químico dinâmico com a
solução do solo, porém com diferentes constantes de equilíbrio,
sendo passível a liberação das reservas quando as formas mais
prontamente disponíveis são removidas da solução do solo
(SPARKS; CARSKI, 1985; KAMINSKI et al., 2007; MELO et
al., 2009; MOUHAMAD; ALSAEDE; IQBAL, 2016).
A reserva de K no solo está diretamente ligada ao
material de origem do solo, à sua composição mineralógica e ao
grau de intemperismo a que foi submetido ao longo do tempo. A
presença de minerais que possuem K em suas estruturas, a
exemplo de micas e feldspatos, resulta em maiores reservas de
K e liberação de fontes não trocáveis em relação a solos que não
possuem esses minerais. Solos mais jovens, com predomínio de
argilominerais do tipo 2:1, também possuem boas reservas de K
pela sua presença nas cargas negativas das entrecamadas
(MELO et al., 2005; MEDEIROS et al., 2014; CHAVES et al.,
2015). O aumento da concentração de K em solos cultivados,
especialmente na rizosfera das plantas, pode favorecer a fixação
em minerais micáceos, como a vermiculita (CALVARUSO et
al., 2014). Entretanto, o uso de fertilizantes potássicos não se
justifica para aumentar a reserva de K no solo, pois com maior
24
disponibilidade pode ocorrer maior absorção pelas plantas,
configurando um consumo de luxo (KAMINSKI et al., 2007).
A liberação de K das formas não trocáveis e estruturais,
assim como o equilíbrio de K no solo, são influenciados por uma
série de fatores. Dentre eles, estão os fatores relativos ao próprio
mineral (composição química, tamanho de partícula,
imperfeições estruturais, grau de depleção de K, alterações nas
cargas das entrecamadas) e ao meio (atividade biológica, ciclos
de umedecimento e secagem, atividade de K na solução,
composição da solução, lixiviação, potencial redox, e
temperatura) (MOUHAMAD; ALSAEDE; IQBAL, 2016). A
presença de outros cátions no solo e o pH do mesmo também
estão entre esses fatores. Dessa forma, a calagem pode interferir
na dinâmica entre as formas de K no solo (DAS; SAHA, 2013).
O objetivo deste trabalho foi avaliar a distribuição das
formas de K não trocável, extraível e na solução em cinco solos
influenciada pela adição de potássio e doses de calcário.
25
1.2
MATERIAL E MÉTODOS
O experimento foi conduzido no Laboratório de Química
e Fertilidade do Solo da Universidade do Estado de Santa
Catarina, em Lages, SC, em 2015. Foram utilizados cinco
classes de solo: um Latossolo Bruno (LT), um Nitossolo
Vermelho (NT), um Argissolo Amarelo (AG) e dois
Cambissolos Húmicos (CB), sendo um deles proveniente de um
plantio de pinus de quarta rotação (CP). As amostras desses
solos foram peneiradas em peneiras com malhas de abertura de
2mm, e posteriormente foram secas para obtenção da fração
terra fina seca ao ar. A caracterização inicial dos solos, antes do
experimento (Tabela 1.1) foi realizada segundo os
procedimentos descritos por Tedesco et al. (1995), à exceção dos
teores não trocáveis e totais de K, que foram determinados
segundo Knudsen et. al. (1986) e Jackson et al., 1986,
respectivamente.
Para fins de caracterização, foram realizadas análise de
textura e mineralogia. A análise mineralógicas foi realizada após
a separação das frações granulométricas. A areia foi separada
por peneiramento; e o silte e a argila por sedimentação, com base
na lei de Stokes (GEE; BAUDER, 1986). A identificação dos
minerais potencialmente fornecedores de K (micas e minerais
2:1) foi feita por difratometria de raios X (DRX), nas três
frações, utilizando um difratômetro de raios-X Philips, modelo
PW 3710, dotado de tubo de cobre, com ângulo de compensação
θ/2θ e monocromador de grafite, com variação angular de 3,2 a
42º2θ. A velocidade angular foi de 0,02º 2θ/s, com tempo de 1
segundo de leitura por passo.
Foram realizados tratamentos adicionais para diferenciar
os minerais secundários do tipo 2:1 da fração argila (WHITTIG;
ALLARDICE, 1986): saturação por Mg, saturação por Mg
seguida por solvatação com etilenoglicol, saturação por K
seguida por secagem ao ar (25ºC) e saturação por K seguida por
secagem a 100ºC, 350ºC e 550 °C, respectivamente. Após esses
26
tratamentos, as amostras foram alocadas em lâminas de vidro e
analisadas por DRX.
Tabela 1.1 - Caracterização química e física dos solos antes da
implantação do experimento.
Areia
Silte Argila
pH
Al3+ H+Al Ca2+ Mg2+
---------g kg-1---------------cmolc dm-3-------NT*
40
221
740
3,7
5,2
8,6
2,1
1,0
LT
46
247
707
3,7
6,2
10,3
3,6
1,3
CB
236
334
430
3,5
10,2
16,8
1,2
0,5
AG
457
287
256
3,8
2,9
4,8
2,9
1,3
CP
171
352
478
3,6
13,4
22,2
0,8
0,2
Ke
Knt
P
C
MO
SB
T
V
---------mg kg-1-----------%----%
NT
98,0
217
5,7
25,9
4,5
3,3
11,9 27,7
LT
80,0
198
8,4
27,5
4,7
5,1
15,3 32,7
CB
76,0
366
10,3
37,7
6,5
1,8
18,6
9,7
AG
63,0
162
7,8
21,1
3,6
4,4
9,1
47,2
CP
126,0
789
11,4
27,9
4,8
1,3
5,2
5,2
*NT: Nitossolo; LT: Latossolo; CB: Cambissolo; AG: Argissolo; CP:
Cambissolo sob cultivo de pinus; Ke: potássio extraível; Knt: potássio não
trocável; MO: matéria orgânica; SB: soma de bases; T: capacidade de troca
de cátions; V: saturação de bases. Fonte: próprio autor, 2016.
Os tratamentos consistiram de um fatorial envolvendo
doses de calcário e doses de potássio. As doses de calcário
corresponderam a 0, 0,25, 0,5, 1,0 e 1,5 vezes a quantidade
indicada pelo índice SMP para elevar o pH até 6,0; as doses de
K corresponderam a 0 e 1000 mg kg-1. Como a concentração de
K na solução do solo é geralmente baixa, utilizou-se uma alta
dose de K aplicada para possibilitar maior viualização dos
efeitos da variação do pH nesta fração do K do solo. Utilizou-se
calcário dolomítico, com poder relativo de neutralização total
(utilizado PRNT) de 88%; o K foi aplicado via líquida, na forma
de cloreto de potássio (KCl). As quantidades de calcário
utilizadas para cada tratamento, em cada solo, são apresentadas
na tabela 1.2.
27
Utilizaram-se amostras de 1,0 kg de solo (base seca).
Após a aplicação do calcário, as amostras de solo foram
armazenadas em sacos plásticos, as quais permaneceram
incubadas por 30 dias, sendo constantemente umedecidas para
manter o solo no teor de água de aproximadamente 80% daquele
retido na capacidade de campo. Após este período, foi aplicada
a solução de KCl nos tratamentos com K, a qual foi
homogeneizada manualmente com o solo, seguindo-se um novo
período de incubação de mais 30 dias. As amostras dos
tratamentos sem K receberam água no mesmo volume líquido
aplicado da solução de KCl. Decorrido este tempo de reação,
foram determinados o pH, os teores de K não trocável (Knt), de
K extraível (Ke) e de K na solução dos solos (Ks).
Tabela 1.2 – Dose equivalente de calcário aplicada em cada
tratamento para cada um dos solos.
Dose equivalente (Mg ha-1)
0,25
0,50
1,00
1,50
NT
4,5
9,0
18,0
27,0
LT
5,5
11,0
22,0
33,0
CB
6,0
12,0
24,0
36,0
AG
2,1
4,2
8,4
12,6
CP
6,0
12,0
24,0
36,0
*NT: Nitossolo; LT: Latossolo; CB: Cambissolo; AG: Argissolo; CP:
Cambissolo sob cultivo de pinus. Fonte: próprio autor, 2016.
Foi realizada a extração das diferentes formas de K
utilizando solução fervente de HNO3 1,0 mol L1 (KNUDSEN
et. al., 1986) e solução de Mehlich-1 (TEDESCO et al., 1995).
A parte do K extraída com Mehlich-1 foi denominada K
extraível, e o K extraído com HNO3, subtraído o teor do K
extraível, foi denominado K não trocável. A extração da solução
do solo para determinação do K foi realizada adicionando água
deionizada nas amostras, na proporção de 1:1, com posterior
filtragem em recipiente de vácuo após período de contato de 12
28
horas (ERNANI; BARBER, 1993). A determinação dos teores
de todas as formas de K foi realizada por leitura em fotômetro
de emissão.
Os dados obtidos foram submetidos à análise de
variância e, quando significativa, submetidos à análise de
regressão (p < 0,05).
29
1.3
RESULTADOS E DISCUSSÃO
1.3.1 ANÁLISES MINERALÓGICAS
Os difratogramas das frações areia e silte apresentaram
picos intensos de quartzo e quantidades mínimas de outros
minerais nas amostras (ANEXOS). Sendo assim, apenas a
composição da fração argila foi considerada na discussão que
segue.
Os difratogramas do Nitossolo (NT) e do Latossolo (LT)
(Figuras 1.1 e 1.2) evidenciam que a composição mineralógica
desses dois solos é semelhante. Em ambos, foi verificada
presença de caulinita, indicada pelos picos em 7,2 e 3,56 Å, que
são destruídos com o aquecimento das amostras a 550ºC. Foi
observado, também, um pico de mineral 2:1 expansível no NT e
no LT, respectivamente em 14,1 e 13,98 Å, sendo esta a posição
do pico atribuída à vermiculita ou à esmectita. Foram ainda
detectados pequenos picos em torno de 4,82, 4,15 e 3,34 Å,
indicando a presença de gibbsita, goethita e quartzo,
respectivamente. Os demais pontos demarcados nos
difratogramas são picos de 2ª ou 3ª ordem dos minerais já
citados.
30
Figura 1.1 – Difratogramas da fração argila do Nitossolo (NT)
de amostras tratadas com saturação de K (A) nas temperaturas
de 25ºC (AK25), 100ºC (AK100), 350ºC (AK350) e 550ºC
(AK550), e de amostras tratadas com saturação de Mg (B) e Mg
+ etileno glicol (AMGEG).
Fonte: próprio autor, 2016.
31
Figura 1.2 – Difratogramas da fração argila de amostras do
Latossolo (LT) tratadas com saturação de K (A) nas
temperaturas de 25ºC (AK25), 100ºC (AK100), 350ºC (AK350)
e 550ºC (AK550), e de amostras tratadas com saturação de Mg
(B) e Mg + etileno glicol (AMGEG).
Fonte: próprio autor, 2016.
32
Os picos da caulinita nesses solos foram intensos, porém
não bem definidos, apresentando assimetria, o que indica que o
mineral não é puro, tem presença de interestratificação do tipo
caulinita-2:1. Considerando que não foi observado pico de mica
nos mesmos (posição de 10 Å), é provável que o mineral 2:1
identificado nas amostras seja a esmectita, já que a vermiculita
é derivada da mica (MELO et al., 2004).
Com o aquecimento das amostras tratadas com K a
350ºC e 550ºC, ocorre o colapso da estrutura dos minerais 2:1,
intensificando os picos em torno de 10 Å, atribuídos aos
minerais 2:1 não expansíveis. Neste caso, houve pouca mudança
do pico inicial, com diluição do pico em direção a 10 Å,
indicando forte intercalação com hidróxidos de Al nas
entrecamadas (MELO et al., 2004). As amostras apresentaram
ainda comportamento anômalo em resposta ao aquecimento a
550ºC, com picos largos em 11,34 Å, não sendo verificados os
picos de segunda ordem desses minerais, que deveriam ocorrer
em aproximadamente 5,5 Å. Este efeito pode ser causado por
forte contribuição de minerais interestratificados, como a
caulinita estratificada com esmectita, e parte da esmectita
interestratificada com algum outro argilomineral do tipo 2:1.
Nos tratamentos com Mg e Mg + etileno glicol, não houve
expansão dos minerais 2:1, indicando novamente a presença de
polímeros, que não permitem a expansão das camadas.
Os Cambissolos CB e CP, também apresentaram
constituição mineralógica semelhante entre si (Figuras 1.3 e
1.4). Os difratogramas indicaram presença de caulinita (picos
em 7,2 e 3,56 Å), minerais 2:1 expansíveis (pico em 14 Å),
pequena quantidade de gibbsita (4,85 Å), quartzo (4,26 e 3,34
Å) e goethita (4,25 Å). No CP, foi ainda observada a presença
de mineral 2:1 não expansível (10 Å), provavelmente mica ou
ilita.
Da mesma forma que nos solos anteriores, o pico da
caulinita foi largo e assimétrico, em função da presença de
caulinita de pequeno tamanho ou, mais provavelmente, devido à
33
presença de interestratificação do tipo caulinita-2:1 (MELO et
al., 2004). O pico dos argilominerais 2:1 expansíveis, em torno
de 14 Å na amostra tratada com K a 25ºC, sofre contração com
o aquecimento em direção a 10 Å. Porém, até a temperatura de
550ºC ainda se mantém uma assimetria causada por intercalação
com polímeros de Al, que não foram completamente destruídos
pela elevação da temperatura, ou pela interestratificação com
minerais 1:1. Os tratamentos com Mg e Mg+etilenoglicol não
alteraram a posição dos picos de minerais 2:1, não sendo
possível identificar qual mineral estava presente na amostra.
Esmectitas puras poderiam expandir até 17 Å, enquanto a
vemiculita iria até 15,5, mas a presença de polímeros impediu a
expansão e a identificação precisa do mineral.
34
Figura 1.3 – Difratogramas da fração argila do Cambissolo (CB)
de amostras tratadas com saturação de K (A) nas temperaturas
de 25ºC (AK25), 100ºC (AK100), 350ºC (AK350) e 550ºC
(AK550), e de amostras tratadas com saturação de Mg (B) e Mg
+ etileno glicol (AMGEG).
Fonte: próprio autor, 2016.
35
Figura 1.4 – Difratogramas da fração argila do Cambissolo sob
cultivo de pinus (CP) de amostras tratadas com saturação de K
(A) nas temperaturas de 25ºC (AK25), 100ºC (AK100), 350ºC
(AK350) e 550ºC (AK550), e de amostras tratadas com
saturação de Mg (B) e Mg + etileno glicol (AMGEG).
Fonte: próprio autor, 2016.
36
No Argissolo (AG), diferentemente do observado nos
outros solos, a caulinita apresentou um pico bem definido, sem
indícios de estratificação (Figura 1.5). Pode ser observado,
também, o pico intenso dos argilominerais 2:1 não expansíveis
(10 Å), mica e ilita. Os argilominerais 2:1 expansíveis aparecem
em pequena quantidade com pico em 13,5 Å, que fica mais
evidenciado no tratamento com Mg expandido a 14,3 Å. O pico
em 10 Å contrai com o aquecimento, mas mantém uma
assimetria leve na base, indicando a presença de polímeros de
Al nas entrecamadas. Foram observados indícios da presença de
gibbsita, quartzo e goethita.
Com base na composição mineralógica, supõe-se que o
Cambissolo sob cultivo de pinus e o Argissolo teriam maior
possibilidade de fornecer K oriundo da reserva mineral,
considerando que esse solo apresentou predomínio de
argilominerais do tipo 2:1 e presença de mica. A cinética de
liberação de K na caulinita e na esmectita é geralmente rápida,
enquanto que na vermiculita e na mica é mais lenta (SPARKS;
CARSKI, 1985). Assim, o Cambissolo seria capaz de fornecer
K tanto a curto quanto a médio e longo prazos.
Ao comparar quatro classes de solos (Neossolo,
Cambissolo, Argissolo e Latossolo), no Ceará, Diniz et al.
(2007) encontraram maiores teores de K no Cambissolo em
relação aos demais, e os menores no Latossolo, compatível com
o alto grau de intemperismo e quantidade de caulinita neste solo.
O Argissolo apresentou altos valores de K total, predominando
na fração areia (84,3%). Resultado semelhante foi obtido por
Medeiros et al. (2014), em solos da Paraíba, onde os solos menos
desenvolvidos apresentaram maiores reservas de K, tanto
trocável quanto não trocável, pela maior quantidade de mica e
minerais 2:1 presentes nessas condições.
37
Figura 1.5 – Difratogramas da fração argila do Argissolo (AG)
de amostras tratadas com saturação de K (A) nas temperaturas
de 25ºC (AK25), 100ºC (AK100), 350ºC (AK350) e 550ºC
(AK550), e de amostras tratadas com saturação de Mg (B) e Mg
+ etileno glicol (AMGEG).
Fonte: próprio autor, 2016.
38
1.3.2 pH
Para todos os solos, houve aumento do pH com o
aumento das doses de calcário aplicadas (Figuras 1.6, 1.7 e 1.8).
Em todos eles, o pH inicial era muito baixo e foi elevado para
valores acima de 6,0, para o qual a correção foi calculada.
Embora o método SMP seja considerado adequado para
determinar a necessidade de calcário em uma ampla gama de
solos (ERNANI; ALMEIDA, 1986), neste caso resultou em
valores superestimados. Mais estudos são necessários para
avaliar as causas deste resultado.
Em todos os solos avaliados, a adição de 1000 mg kg-1
de K ocasionou pequeno decréscimo nos valores de pH em
relação aos tratamentos sem a adição de K (Figuras 1.6, 1.7 e
1.8). Isso ocorre em função do efeito indireto do K sobre a
atividade dos íons H+, por causa da troca iônica entre esses
elementos no complexo de troca (SOUSA; MIRANDA;
OLIVEIRA, 2007) e pelo aumento da concentração eletrolítica
do solo, que resulta em queda no pH em solos com predomínio
de cargas elétricas negativas (ALMEIDA; ERNANI, 1996).
O aumento proporcional semelhante entre os
solos, mesmo com quantidades diferentes de calcário, se dá pelo
maior tamponamento de alguns deles. Nos dois Cambissolos, a
quantidade aplicada foi maior que nos demais solos,
possivelmente pelo tamponamento causado pelo alto teor de
matéria orgânica, de Al3+ e de H+Al encontrado nesses dois
solos em relação aos demais (Tabela 1.1).
39
Figura 1.6 – Valores de pH no Nitossolo (A) e no Latossolo (B)
em função da adição de doses de calcário, na presença ou não de
adição de 1000 mg kg-1 de K.
8
A
pH
6
4
2
2
Sem K
y = 2,2x + 4,4 R = 93,50
Com K
y = 2,13x + 4,34 R = 94,04
2
0
0,00
0,25
0,50
1,00
1,50
1,00
1,50
8
B
pH
6
4
2
Sem K
y = 2,02x + 4,8 R2 = 93,4
Com K
y = 2,1x + 4,2 R2 = 90,5
0
0,00
0,25
0,50
Fração da dose de calcário
Fonte: próprio autor, 2016.
40
Figura 1.7 – Valores de pH no Cambissolo (A) e no Cambissolo
sob cultivo de pinus (B) em função do aumento das doses de
calcário, na presença ou não de adição de 1000 mg kg-1 de K.
8
A
pH
6
4
2
2
Sem K
y = 1,9x + 4,6 R = 92,7
Com K
y = 2,1x + 4,0 R = 94,4
2
0
0,00
0,25
0,50
1,00
1,50
1,00
1,50
8
B
pH
6
4
2
2
Sem K
y = 1,6x + 5,2 R = 87,8
Com K
y = 1,6x + 4,7 R2 = 95,0
0
0,00
0,25
0,50
Fração da dose de calcário
Fonte: próprio autor, 2016.
41
Figura 1.8. Valores de pH no Argissolo em função do aumento
das doses de calcário, na presença ou não de adição de 1000 mg
kg-1 de K.
8
pH
6
4
2
Sem K
y = 2,046x* + 4,4044 R2 = 93,91
Com K
y = 2,239x** + 3,6527 R2 = 96,32
0
0,00
0,25
0,50
1,00
1,50
Fração da dose de calcário
Fonte: próprio autor, 2016.
1.3.3 TEORES DE POTÁSSIO
Os teores de K na solução (Ks) dos solos foram os que
demonstraram mais expressivamente os efeitos da adição de
doses de calcário, sendo significativos em todos os solos,
independentemente da adição ou não de K (Figuras 1.9, 1.10 e
1.11).
No Nitossolo, os teores de Ks diminuíram com e sem a
aplicação de K com o aumento da dose de calcário aplicada
(Figura 1.9A). Na ausência da adição de K, a magnitude da
redução proporcionada pelo aumento da adição de calcário foi
de 85%, enquanto que com adição de K, foi de apenas 24%. Esta
maior redução sem a adição de K é relativa, por um efeito de
escala. Como os valores sem a adição de K são menores,
42
pequenas variações resultam em maior porcentagem de variação
do que quando considerados altos valores.
Apesar das diferenças na composição mineralógica, o
Latossolo e os Cambissolos apresentaram comportamento
bastante semelhante na variação dos teores de K na solução
devido à adição de calcário (Figuras 1.9B e 1.10). Nos
tratamentos com adição de K, houve redução do Ks de cerca de
45% com o aumento das doses de calcário, nos três solos. Sem
adição de K, a redução foi menor no Latossolo e no Cambissolo,
33 e 39%, respectivamente, e maior no Cambissolo sob cultivo
de pinus, que apresentou redução de aproximadamente 60%.
O Argissolo apresentou diminuição no Ks pela aplicação
de calcário de aproximadamente 20% da menor para a maior
dose, sem a adição de K. Com a adição de K, os valores variaram
entre as doses de calcário, porém sem uma tendência clara de
aumento ou diminuição.
Considerando o predomínio de caulinita na maioria dos
solos (Tabela 1.3) e assim a presença predominante de cargas
elétricas variáveis dependentes de pH, a diminuição dos teores
de K na solução dos solos está relacionada com a maior adsorção
causada pelo aumento das cargas elétricas negativas livres.
Além disso, a complexação de Al3+ e a dissociação de H+ da
superfície dos componentes sólidos resulta em maior adsorção
dos cátions presentes na solução devido ao aumento do número
de cargas elétricas negativas livres na superfície dos colóides
(ERNANI, 2016).
A redução expressivamente maior do Ks quando não
houve adição de K em alguns solos em relação a outros pode ser
justificada pela menor disponibilidade inicial de K na solução.
Dessa forma, uma maior quantidade do nutriente tem que ser
movida para o complexo de troca a fim de manter o equilíbrio
das formas no solo.
43
Figura 1.9 - Teor de K na solução no Nitossolo (A) e no
Latossolo (B) em função do aumento das doses de calcário, na
presença ou não de adição de 1000 mg kg-1 de K.
350
2
Sem K y = 11,80x *** - 27,94x*** + 19,24
300
Com K
y = 9,99x2 - 50,60x* + 201,47
R2 = 95,32
A
R2 = 93,90
-1
K solução (mg kg )
250
200
150
100
50
0
0,00
0,25
0,50
1,00
1,50
350
Sem K
300
Com K
y = -0,65x2** + 0,32x + 1,94
R2 = 64,02
y = 56,69x2** - 134,53x*** + 188,85
B
R2 = 89,95
-1
K solução (mg kg )
250
200
150
100
50
0
0,00
0,25
0,50
1,00
Fração da dose de calcário
Fonte: próprio autor, 2016.
1,50
44
Figura 1.10 – Teor de K na solução no Cambissolo (A) e no
Cambissolo sob cultivo de pinus (B) em função do aumento das
doses de calcário, na presença ou não de adição de 1000 mg kg1
de K.
350
300
Sem K
y = 0,59x2** - 1,12x** + 2,26
R2 = 36,62
Com K
y = 81,94x2*** - 174,01x*** + 199,12
A
R2 = 87,08
-1
K solução (mg kg )
250
200
150
100
50
0
0,00
0,25
0,50
1,00
1,50
350
300
Sem K
y = 2,58x2** - 5,64x** +5,02
R2 = 81,02
Com K
y = 98,90x2*** - 214,71x*** + 201,09
B
R2 = 98,29
-1
K solução (mg kg )
250
200
150
100
50
0
0,00
0,25
0,50
1,00
Fração da dose de calcário
Fonte: próprio autor, 2016.
1,50
45
Figura 1.11 – Teor de K na solução no Argissolo em função do
aumento das doses de calcário, na presença ou não de adição de
1000 mg kg-1 de K.
350
300
-1
K solução (mg kg )
250
200
Sem K
150
y = 1,49x2* - 2,92x** + 4,95
2
Com K y = 12,20x - 8,08x + 305,56
R2 = 96,37
R2 = 7,82
100
50
0
0,00
0,25
0,50
1,00
1,50
Fração da dose de Calcário
Fonte: próprio autor, 2016.
Os teores de K extraível (Ke) apresentaram menor
variação em relação aos tratamentos que os teores de Ks (Figuras
1.12, 1.13 e 1.14). O Nitossolo e o Latossolo apresentaram
tendências semelhantes, com diminuição dos teores de Ke com
o aumento da dose de calcário tanto com a adição de K quanto
sem (Figura 1.12). A magnitude da diminuição foi maior sem a
adição de K, sendo de 60% para o Nitossolo e de 30% para o
Latossolo, enquanto que com a adição de K esses valores caíram
para 4% e 12%, respectivamente. Da mesma forma que
observado para o Ks, essas diferenças são relativas à escala de
valores.
46
Os dois Cambissolos, sendo de composição muito
semelhante, apresentaram o mesmo comportamento, também
com diminuição do Ke com o aumento da dose de calcário,
porém somente quando foi adicionado K. Essa diminuição foi
pouco expressiva, sendo de apenas 10% e 6% para o Cambissolo
e o Cambissolo sob cultivo de pinus, respectivamente. No
Argissolo não foi observado efeito dos tratamentos no Ke.
Considerando que o teor de Ks diminuiu com o aumento
das doses de calcário que passaram para a forma adsorvida, era
esperado aumento do Ke, porém como o aumento é pequeno,
pode não ter sido mensurado pelos métodos utilizados. Além
disso, é possível que as formas que já se encontravam adsorvidas
tenham formado ligações de maior energia em função do
aumento da sua atividade na solução, quando K foi adicionado,
fazendo com que não fossem mais quantificados pelo método de
extração empregado. No caso do Argissolo, onde não houve
diferença, a diminuição do Ks deve ter ocorrido na mesma
magnitude desta passagem para formas mais indisponíveis,
mantendo os teores no complexo de troca inalterados.
47
Figura 1.12 – Teor de K extraível no Nitossolo (A) e no
Latossolo (B) em função do aumento das doses de calcário, na
presença ou não de adição de 1000 mg kg-1 de K.
A
1000
-1
K extraível (mg kg )
800
600
400
2
Sem K
y = 114,51x ** - 244,45x** + 200,00
Com K
y = -39,48x** + 938,66
2
R = 93,57
2
R = 43,64
200
0
0,00
0,25
0,50
1,00
1000
1,50
B
-1
K extraível (mg kg )
800
600
400
Sem K
y = -11,45x** + 52,24
R2 = 86,26
Com K
y = -42,41x* + 854,57
R = 38,76
2
200
0
0,00
0,25
0,50
1,00
Fração da dose de Calcário
Fonte: próprio autor, 2016.
1,50
48
Figura 1.13 – Teor de K extraível no Cambissolo (A) e no
Cambissolo sob cultivo de pinus (B) em função do aumento das
doses de calcário, na presença ou não de adição de 1000 mg kg1
de K.
1000
A
-1
K extraível (mg kg )
800
600
Sem K
Com K
y = -65,00x*** + 851,25
R2 = 86,65
400
200
0
0,00
0,25
0,50
1,00
1,50
1000
B
-1
K extraível (mg kg )
800
600
Sem K
Com K y = -45,93x2* + 16,41x + 785,06
400
R2 = 60,14
200
0
0,00
0,25
0,50
1,00
Fração da dose de calcário
Fonte: próprio autor, 2016.
1,50
49
Figura 1.14 – Teor de K extraível no Argissolo em função do
aumento das doses de calcário, na presença ou não de adição de
1000 mg kg-1 de K.
1000
-1
K extraível (mg kg )
800
600
Sem K
Com K
400
200
0
0,00
0,25
0,50
1,00
1,50
Fração da dose de calcário
Fonte: próprio autor, 2016.
A calagem não afetou os teores de Knt em quatro dos
cinco solos avaliados (Figuras 1.15, 1.16 e 1.17). No Nitossolo,
ocorreu diminuição do Knt com o aumento da dose de calcário,
porém somente quando não foi adicionado K (Figura 1.15A). A
magnitude da redução foi de aproximadamente 50%.
Neste caso, a diminuição pode ter ocorrido devido ao
aumento da energia de ligação do K que já estava na forma não
trocável. Estudos de cinética de liberação de K apontam mais de
uma taxa ou velocidade, indicando a liberação de formas de K
com diferentes energias de interação (MEURER; CASTILHOS,
2001; CHAVES et al., 2015). Além disso, em comparação com
outros métodos de extração, com ácido oxálico e NaHSO4, o
ácido nítrico extrai as menores quantidades de nutrientes
50
(CHAVES et al., 2015), podendo não ter sido eficiente para
extrair as formas de K mais fortemente retidas pelos solos.
Figura 1.15 – Teor de K não trocável no Nitossolo (A) e no
Latossolo (B) em função do aumento das doses de calcário, na
presença ou não de adição de 1000 mg kg-1 de K.
1000
A
800
-1
K HNO3 (mg kg )
Sem K
Com K
600
400
200
0
0,00
0,25
0,50
1,00
1000
1,50
B
800
-1
K HNO3 (mg kg )
Sem K
600
Com K
400
200
0
0,00
0,25
0,50
1,00
Fração da dose de calcário
Fonte: próprio autor, 2016.
1,50
51
Figura 1.16 – Teor de K não trocável no Cambissolo (A) e
Cambissolo sob cultivo de pinus (B) em função do aumento das
doses de calcário, na presença ou não de adição de 1000 mg kg1
de K.
1000
A
800
-1
K HNO3 (mg kg )
Sem K
Com K
600
400
200
0
0,00
0,25
0,50
1,00
1,50
B
1000
-1
K HNO3 (mg kg )
800
600
400
200
Sem K
Com K
0
0,00
0,25
0,50
1,00
Fração da dose de Calcário
Fonte: próprio autor, 2016.
1,50
52
Já no Argissolo, houve variação no Knt somente sem a
adição de K (Figura 1.17), ocorrendo aumento do Knt com o
aumento da quantidade de calcário aplicada. Este aumento
observado pode ser reflexo da precipitação de hidróxidos de Fe
e Al, que dificultavam a liberação do K da posição não trocável
(DAS; SAHA, 2013), bem como permitindo a entrada e fixação
do Ke nestes mesmos espaços.
Figura 1.17 – Teor de K não trocável no Argissolo em função do
aumento das doses de calcário, na presença ou não de adição de
1000 mg kg-1 de K.
1000
800
-1
KHNO3 (mg kg )
Sem K
y = -12,6x2 + 27,8x + 37,4
R2 = 83,2
Com K
600
400
200
0
0,00
0,25
0,50
1,00
1,50
Fração da dose de calcário
Fonte: próprio autor, 2016.
Os resultados obtidos diferem dos observados por Das e
Saha (2013), que ao avaliarem o efeito de diferentes doses de
calcário obtiveram os maiores teores de Knt no solo
completamente corrigido. Essa diferença pode ser devida à
composição mineralógica do solo avaliado por estes autores em
53
relação aos solos desse estudo. Os teores de Knt obtidos por
esses autores foram bem superiores aos observados no presente
estudo, variando de 1650 mg kg-1, na ausência de calagem, a
2270 mg kg-1 com calagem completa.
Entre os solos avaliados, os maiores teores foram
encontrados no Cambissolo sob cultivo de pinus, com teor
médio de 533 mg kg-1 sem adição de K. Este valor se justifica,
uma vez que solos que tenham maior teor de argila ou
predomínio de esmectita ou ilita na composição mineralógica
apresentam altos teores de Knt (BRITZKE et al., 2012), e este
foi o único solo que apresentou predomínio de argilominerais do
tipo 2:1 na fração argila (Tabela 1.3).
Assim como para o Ke, os menores teores de Knt são
encontrados em solos mais intemperizados, devido a menor
presença de minerais primários, fontes de K (MELO et al.,
2005).
Embora a aplicação de K tenha resultado em teores muito
maiores de Knt comparado com a ausência de aplicação desse
nutriente, em condições de campo a adubação não deve ser
considerada como uma maneira de aumentar a reserva dos solos,
uma vez que existem perdas e consumo pelas plantas das formas
mais prontamente disponíveis, sendo a reserva dependente das
características intrínsecas de cada solo (KAMINSKI et al.,
2007).
54
1.4
CONCLUSÕES
O aumento da dose de calcário eleva o pH em todos os
solos, e a adição combinada com a adubação potássica resulta
em valores de pH inferiores aos atingidos somente com a
calagem.
O teor de K na solução do solo diminui com o aumento
da dose de calcário, em todos os solos, sendo que no Latossolo
e nos Cambissolos essa redução é menor quando a calagem é
combinada com a adubação potássica.
O teor de K extraível diminui com o aumento das doses
de calcário, em todos os solos exceto o Argissolo, com a adição
combinada de K.
O teor de K não trocável aumentou com o aumento da
calagem somente no Argissolo, quando não houve adição de K.
A calagem diminui o teor de K na solução em solos com
variadas texturas e composições mineralógicas, podendo
resultar em menores perdas por lixiviação. Entretando, também
resulta em menor disponibilidade para as plantas, por diminuir o
teor das formas prontamente disponíveis.
55
1.5
REFERÊNCIAS BIBLIOGRÁFICAS
ALMEIDA, J.A.; ERNANI, P.R. Influência do solvente, da
relação solo/solvente, e da incubação das amostras úmidas na
variação do pH de solos catarinenses. Ciência Rural, v. 26, n.
1, p. 81-85, 1996.
BRITZKE, D.; SILVA, L.S.; MOTERLE, D.F.;
RHEINHEIMER, D.S.; BORTOLUZZI, E.C. A study of
potassium dynamics and mineralogy in soils from subtropical
Brazilian lowlands. J. Soils Sediments, v. 12, p. 185-197,
2012.
CALVARUSO, C.; COLLIGNON, C.; KIES, A.;
TURPAULT, M. Seasonal evolution of the rhizosphere effect
on major and trace elements in soil solutions of Norway Spruce
(Picea abies Karst) and Beech (Fagus sylvatica) in an acidic
forest soil. Open Journal of Soil Science, v. 4, p. 323-336,
2014.
CHAVES, E.; PEDRON, F.A.; MELO, V.F.; DALMOLIN,
R.S.D. Reserva mineral de K por diferentes métodos em
Argissolos subtropicais. Revista Brasileira de Ciência do
Solo, Viçosa, v. 39, p. 1088-1099, 2015.
DAS, R.; SAHA, D. Changes in non-exchangeable K in an acid
and the corresponding limed soil as affected by addition of
nitrogenous and potassic fertilizers. Journal of Crop and
Wood, v. 9, n. 1, p. 99-102, 2013.
ERNANI, P.R. & BARBER, S.A. Composição da solução do
solo e lixiviação de cátions afetadas pela aplicação de cloreto e
sulfato de cálcio em um solo ácido. Revista Brasileira de
Ciência do Solo, 10:41-46, 1993.
56
ERNANI, P.R., ALMEIDA, J.A. Comparação de métodos
analíticos para avaliar a necessidade de calcário dos solos do
Estado de Santa Catarina. Revista Brasileira de Ciência do
Solo, Campinas, v. 10, n. 2, p. 143-150, 1986.
ERNANI, P.R. Química do solo e disponibilidade de
nutrientes. 2.ed. Lages: O autor. 2016.
GEE, G.W. & BAUDER, J.W. Particle-size analysis. In:
KLUTE,A., ed. Methods of soil analysis. 2.ed. Madison,
American Society of Agronomy. p.383-412. 1986.
JACKSON, M.L.; LIM, C. H. & ZELAZNY, L.W. Oxides,
hydroxides, and aluminosilicates. In: KLUTE, A. (Ed.)
Methods of soil analysis. Parte 1: Physical and mineralogical
methods. Madson: American Society of Agronomy, 1986. p.
101-150.
KAMINSKI, J.; BRUNETTO, G.; MOTERLE, D.F.;
RHEINHEIMER, D.S. Depleção de formas de potássio do solo
afetada por cultivos sucessivos. Revista Brasileira de Ciência
do Solo, v. 31, p. 1003-1010, 2007.
KNUDSEN, D.; PETERSON, G.A. & PRATT, P.F. Lithium,
sodium and potassium. In: PAGE, A. L. (Ed.), Methods of soil
analysis. Part 2. Chemical and microbiological properties,
American Society of Agronomy. Madison, p. 225-246, 1986.
MEDEIRO, J.S.; OLIVEIRA, F.H.T.; SANTOS, H.C.;
ARRUDA, J.A.; VIEIRA, M.S. Formas de potássio em solos
representativos do Estado da Paraíba. Revista Ciência
Agronômica, Fortaleza, v. 45, n. 2, p. 417-426, 2014.
MELO, G.W.; MEURER, E.J.; PINTO, L.F.S. Fontes de
potássio em solos distroférricos cauliníticos originados de
57
basalto no Rio Grande do Sul. Revista Brasileira de Ciência
do Solo, Viçosa, v. 28, p. 597-603, 2004.
MELO, V.F.; CORRÊA, G.F.; RIBEIRO, A.N.; MASCHIO,
P.A. Cinética de liberação de potássio e magnésio pelos
minerais da fração argila de solos do triângulo mineiro.
Revista Brasileira de Ciência do Solo, v. 29, p. 533-545,
2005.
MELO, V.F.; CASTILHOS, R.M.V. & PINTO, L.F.S. Reserva
mineral dos solo. In: MELO, V.F. & ALLEONI, L.R.F. (eds).
Química e Mineralogia do Solo, Parte I, SBCS. p. 251-332,
2009.
MEURER, E.J.; CASTILHOS, R.M.V. Liberação de potássio
de frações de solos e sua cinética. Revista Brasileira de
Ciência do Solo, Viçosa, v. 25, p. 823-829, 2001.
MOUHAMAD, R.; ALSAEDE, A.; IQBAL, M. Behavior of
potassium in soil: a mini review. Chemistry International, v.
2, n. 1, p. 58-69, 2016.
SPARKS, D.L.; CARSKI, T.H. Kinetics of potassium
exchange in heterogeneous systems. Applied Clay Science,
Amsterdam, v. 1, p. 89-101, 1985.
TEDESCO, M.J.; GIANELLO, C.; BISSANI, C.A.;
BOHNEN, H. & VOLKWEISS, S. Análise de solo, plantas e
outros materiais. Porto Alegre, Universidade Federal do Rio
Grande do Sul, 1995. 174p.
WHITTIG, L.D. & ALLARDICE, W.R. X-ray diffraction
techniques. In: KLUTE, A., ed. Methods of soil analysis.
Madison, American Society of Agronomy, p. 331-362. 1986.
58
59
CAPÍTULO II – FORMAS DE POTÁSSIO EM SOLOS
CATARINENSES E SUA UTILIZAÇÃO POR MUDAS
DE PINUS
2.1
INTRODUÇÃO
As árvores do gênero Pinus foram introduzidas no Brasil
há muitos anos e sempre apresentaram bom desenvolvimento,
mesmo em solos considerados pouco férteis, gerando um
conceito de que essas plantas poderiam dar origem a florestas
altamente produtivas sem a necessidade de manejo com
fertilizantes (SIMÕES et al., 1970). Ainda assim, diversos
estudos foram conduzidos no sentido de buscar o aumento da
sua produtividade por meio da fertilização do solo, porém os
resultados obtidos foram inconclusivos (LOPES et al., 1983).
No cultivo florestal a dinâmica dos nutrientes ocorre de
maneira diferente do observado em solos cultivados com
espécies anuais devido a vários fatores, entre eles a duração do
ciclo de cultivo, a profundidade explorada pelo sistema radicular
e a ciclagem interna de nutrientes, que faz com que a planta não
dependa exclusivamente do solo para suprir as suas necessidades
(SMETHURST, 2010). Além disso, em alguns casos, não se
obtém resposta à adubação pois os nutrientes estão sendo
providos por outras fontes não prontamente disponíveis, como
por exemplo, por meio do intemperismo dos minerais do solo.
Assim, a determinação dos minerais presentes nas diferentes
frações do solo possibilita quantificar a reserva de nutrientes e
estimar a sua contribuição para a nutrição vegetal, auxiliando no
manejo das culturas, especialmente as de ciclo longo ou perenes
(MELO et al., 1995). Desta forma, o estudo da mineralogia dos
solos é de grande importância para determinar o potencial de
fertilidade dos solos (REATTO et al., 1998).
O potássio se destaca dentre todos os elementos que são
passíveis de liberação a partir das reservas minerais. O K é um
elemento requerido em grandes quantidades pelas culturas,
60
sendo mais de 170 kg ha-1 do nutriente exportado na colheita de
árvores inteiras (SCHUMACHER et al., 2013). No entanto,
como na prática grande parte da biomassa permanece na área, a
exportação real seria de cerca de 90 kg ha-1, considerando
somente o tronco.
Geralmente, os solos possuem baixa concentração de K
na solução e predomínio da forma estrutural, ocorrendo intensa
liberação das reservas em função do equilíbrio existente entre as
formas do nutriente no solo, constituindo um importante fator de
produtividade das culturas (MELO et al., 2009). Como exemplo,
no trabalho de Alves et al. (2013), foram observadas correlações
positivas e significativas entre os teores de K nas acículas de
Pinus taeda e os teores não trocáveis de K extraídos com HNO3
fervente, evidenciando a participação dessa forma na nutrição
das plantas. A absorção resulta em um processo contínuo de
depleção das formas não trocáveis, especialmente quando os
teores disponíveis no solo são baixos (KAMINSKI et al., 2007).
Os feldspatos potássicos e as micas são, normalmente, os
principais minerais potencialmente fornecedores de K, sendo
abundantes numa grande variedade de rochas (BORTOLUZZI
et al., 2005; DINIZ et al., 2007). Devido à facilidade de
intemperização, esses minerais contribuem para a reserva de K
da fração argila de diferentes solos (MELO et al., 2003). A
planta pode obter nutrientes mesmo das frações mais grosseiras,
já que a capacidade de liberação do nutriente é maior em solos
com elevados teores de K na fração areia, devido à presença de
minerais primários (MELO et al., 2004; KOELE et al., 2010),
porém, em solos altamente intemperizados e com material de
origem pobre, ressalta-se a importância das frações mais finas
na reserva do mineral (MELO et al., 2003; BRITZKE et al.,
2012).
As plantas vasculares, principalmente as árvores com
sistema radicular profundo, atuam mais intensamente no
processo de intemperismo dos minerais em comparação com
plantas não-vasculares ou superfícies não-vegetadas
61
(ANDREWS et al., 2008), especialmente quando o mineral
possui na sua composição algum nutriente em falta nas formas
disponíveis (BAKKER et al., 2004; BORTOLUZZI et al.,
2005). O mesmo se aplica ao potássio, uma vez que diversos
trabalhos demonstram o aumento do intemperismo de minerais
micáceos quando sob influência radicular de espécies florestais
(LEYVAL;BERTHELIN, 1991; CALVARUSO et al., 2006;
WALLANDER, 2000).
De maneira geral, as alterações causadas no solo e nos
minerais ocorrem na região da rizosfera, que pode ser definida
como o volume de solo influenciado pela atividade radicular
(GOBRAN et al., 1998; HINSINGER, 1998). A rizosfera difere
do restante do volume do solo em vários aspectos devido à
interação de processos bióticos e abióticos que podem
influenciar a biogeoquímica dos solos florestais, dentre eles a
atividade fúngica e microbiana, e a indução das plantas através
da absorção de água e nutrientes e da exsudação de compostos
orgânicos (BAKKER et al., 1999; GOBRAN et al. 1998;
TURPAULT et al., 2005). Os exsudatos radiculares, como os
ácidos orgânicos, podem alterar o pH da rizosfera através da
absorção de cátions e ânions (MARSCHNER, 1995). Esta
alteração tem influência nos componentes minerais presentes no
solo, resultando em menor quantidade de minerais
intemperizáveis e transformação acelerada da passagem dos
minerais de uma forma para outra (LEYVAL; BERTHELIN,
1991; GOBRAN et al., 1998).
O objetivo do presente trabalho foi avaliar a distribuição
das formas de potássio em diferentes solos, e sua contribuição
na nutrição e crescimento de mudas de pinus, em casa-devegetação.
62
2.2
MATERIAL E MÉTODOS
Foram coletadas cinco classes de solo, de forma a
representar diversas composições mineralógicas, sendo um
Latossolo Bruno (LT), um Nitossolo Vermelho (NT), um
Argissolo Amarelo (AG) e dois Cambissolos Húmicos (CB),
sendo um deles proveniente de um plantio de pinus de quarta
rotação (CP). As amostras foram coletadas na profundidade de
0 a 30 cm, através da abertura de trincheiras.
Após a coleta dos solos, foram realizadas análises
químicas, físicas e mineralógicas para a caracterização inicial
(Tabela 2.1). Para a realização da análise química, as amostras
foram passadas em peneira de 2 mm para obtenção da fração
denominada terra fina seca ao ar (TFSA). Na sequência, elas
foram secas em estufa à temperatura de 60°C, sendo
determinados os valores de pH, C, e os teores trocáveis de K,
Ca, Mg, Al e P seguindo os procedimentos descritos por Tedesco
et al. (1995).
Para as análises físicas e mineralógicas foi realizada a
separação da fração areia, por peneiramento; as frações argila e
silte foram separadas por sedimentação, com base na lei de
Stokes (GEE; BAUDER, 1986). A identificação dos minerais
potencialmente fornecedores de K (micas e minerais 2:1) foi
feita por difratometria de raios X (DRX), nas três frações.
Foram realizados tratamentos adicionais para diferenciar
os minerais secundários do tipo 2:1 da fração argila (WHITTIG;
ALLARDICE, 1986): saturação por Mg, saturação por Mg e
solvatação com etilenoglicol, saturação por K e secagem ao ar
(25ºC), saturação por K e secagem a 100ºC, 350ºC e 550 °C.
Após os tratamentos, as amostras foram novamente analisadas
por DRX.
As amostras de solo coletadas foram secas ao ar,
peneiradas (4 mm) e homogeneizadas para compor a amostra
representativa de cada solo. As amostras de solo
homogeneizadas foram acondicionadas em vasos de 4 kg de solo
63
onde foi plantada uma muda de Pinus taeda, formando a unidade
experimental. O trabalho foi conduzido em casa de vegetação,
em Lages, nos anos de 2012 e 2013.
Tabela 2.1 – Caracterização química e física dos solos antes da
implantação de experimento.
Areia
Silte Argila
pH
Al3+ H+Al Ca2+ Mg2+
-1
---------g kg ---------------cmolc dm-3-------NT*
40
221
740
3,7
5,2
8,6
2,1
1,0
LT
46
247
707
3,7
6,2
10,3
3,6
1,3
CB
236
334
430
3,5
10,2
16,8
1,2
0,5
AG
457
287
256
3,8
2,9
4,8
2,9
1,3
CP
171
352
478
3,6
13,4
22,2
0,8
0,2
Ke
Knt
P
C
MO
SB
T
V
---------mg kg-1-----------%----%
NT
98,0
217
5,7
25,9
4,5
3,3
11,9 27,7
LT
80,0
198
8,4
27,5
4,7
5,1
15,3 32,7
CB
76,0
366
10,3
37,7
6,5
1,8
18,6
9,7
AG
63,0
162
7,8
21,1
3,6
4,4
9,1
47,2
CP
126,0
789
11,4
27,9
4,8
1,3
5,2
5,2
*NT: Nitossolo; LT: Latossolo; CB: Cambissolo; AG: Argissolo; CP:
Cambissolo sob cultivo de pinus; Ke: potássio extraível; Knt: potássio não
trocável; MO: matéria orgânica; SB: soma de bases; T: capacidade de troca
de cátions; V: saturação de bases. Fonte: próprio autor, 2016.
O experimento foi composto por quatro tratamentos:
adubação potássica (TK), aplicação de calcário (TC), aplicação
de calcário + adubação potássica (TCK) e uma testemunha (TT).
O potássio foi aplicado na forma de KCl, na concentração de 50
mg kg-1; o calcário foi aplicado na quantidade recomendada para
elevar o pH a 6,0 de acordo com o método SMP, misturado em
todo o solo do vaso. Em todos os vasos, foram adicionados 30
mg kg-1 de N, na forma de ureia, e 90 mg kg-1 de P, na forma de
fosfato de cálcio (PA).
Utilizou-se o delineamento estatístico inteiramente
casualizado, com quatro repetições para cada solo, totalizando
64
80 unidades experimentais. Foram conduzidos dois cultivos
consecutivos, o primeiro com duração de seis meses, e o
segundo, utilizando o mesmo solo do primeiro, com duração de
nove meses.
Após cada cultivo, foi realizada a medição da altura e o
diâmetro do colo de cada planta. Em seguida, a planta coletada
foi separada em acículas, caule e raízes, e esse material foi seco
em estufa à 60ºC, até massa constante, para determinação da
massa seca de raiz e da parte aérea. Posteriormente, o material
da parte aérea foi moído para a determinação dos teores de K,
Ca e Mg, digerido pelo uso da mistura de ácido nítrico e ácido
perclórico (TEDESCO et al., 1995), sendo a determinação do K
feita por fotometria de emissão e a de Ca e Mg por
espectrofotometria de absorção atômica.
A análise química do solo foi realizada novamente após o
término de cada um dos cultivos. Também foram realizadas
análises químicas no solo rizosférico, que foi separado da massa
do solo através de leve agitação do sistema radicular, e
posteriormente separado das raízes por uma agitação mais
vigorosa (BAZIRAMAKENGA et al., 1995). Foram
determinados os teores trocáveis de Ca, Mg e K (TEDESCO et
al., 1995). Além do teor trocável de K (Mehlich-1), aqui
denominado de extraível, o K extraído com solução fervente de
HNO3 1,0 mol L1 (KNUDSEN et. al., 1986), subtraído o teor
do K extraível, foi denominado K não trocável; A extração da
solução do solo para determinação do K foi realizada
adicionando água deionizada na amostra, na proporção de 1:1,
com posterior filtragem em recipiente de vácuo, após período de
contato de 12 horas (ERNANI; BARBER, 1993). A
determinação dos teores de todas as formas de K foi realizada
por fotometria de emissão.
Os dados obtidos foram submetidos à análise de variância;
quando houve significância, as médias foram comparadas pelo
teste de Tukey a 5% de probabilidade de erro.
65
2.3
RESULTADOS E DISCUSSÃO
No primeiro cultivo, foram observados efeitos dos
tratamentos no desenvolvimento das mudas apenas em dois dos
cinco solo (no LT e no AG), sendo que em ambos a aplicação de
calcário diminuiu o crescimento das plantas (Tabela 2.2). Os
efeitos sobre o crescimento das plantas nestes tratamentos foi
visível, sendo que além do crescimento diminuído as plantas
também apresentaram amarelecimento de acículas.
Todos os parâmetros avaliados no Argissolo (AG) foram
inferiores nos tratamentos com aplicação de calcário em relação
aos demais tratamentos. Para o Latossolo (LT), a massa seca da
parte aérea das mudas foi inferior nos tratamentos com calcário
em relação aos demais; o diâmetro e a massa seca de raízes no
tratamento com K + calcário foram inferiores aos da testemunha,
e a altura das plantas no tratamento com calcário foi inferior à
altura no tratamento com K.
De maneira geral, o pinus não foi afetado pelo baixo
valor de pH, pois não houve melhoria do crescimento das plantas
pela adição de calcário, nem mesmo no CB e no CP, que
apresentaram altos teores de Al trocável (Tabela 2.1). O mesmo
é válido para Ca e Mg, já que não houve resposta à adição de
calcário mesmo quando os teores eram muito baixos, a exemplo
do CP. A adição isolada de K não incrementou o
desenvolvimento das plantas nos solos, mesmo quando os teores
eram muito baixos, como no caso do AG.
66
Tabela 2.2 – Altura, diâmetro e massa seca de raiz e parte aérea
de mudas de pinus, no primeiro cultivo, em função da adição de
K, calcário e da combinação de ambos, em cinco solos
catarinenses.
NT*
LT
CB
AG
CP
Testemunha
Potássio
Calcário
Calcário + K
57,5
60,8
64,0
59,8
a**
a
a
a
Altura de mudas (cm)
66,3 a
58,0 a 60,3
68,0 a
59,0 a 60,3
51,8 b
59,5 a 46,3
62,5 ab 56,3 a 52,5
a
a
b
ab
67,0
59,8
62,5
68,0
a
a
a
a
Testemunha
Potássio
Calcário
Calcário + K
8,1
8,8
8,2
8,6
a
a
a
a
Diâmetro de mudas (cm)
9,3 a
9,0 a 9,0
8,5 ab 8,9 a 9,0
8,2 ab 8,5 a 6,3
7,1 b
7,9 a 6,1
a
a
b
b
8,8
8,7
9,0
8,4
a
a
a
a
Massa seca de raiz (g vaso-1)
9,2 a
8,9 a 8,6 a
7,6 ab 9,1 a 8,3 a
6,8 ab 7,5 a 4,0 b
5,2 b
7,2 a 4,3 b
9,1
10,1
9,9
7,6
a
a
a
a
Testemunha
Potássio
Calcário
Calcário + K
7,2
8,1
7,1
7,6
a
a
a
a
Massa seca de parte aérea (g vaso-1)
Testemunha
20,6 a
25,2 a
21,3 a 22,9 a
25,9 a
Potássio
20,6 a
25,3 a
24,5 a 23,7 a
25,0 a
Calcário
19,9 a
16,4 b
21,9 a 9,0 b
24,7 a
Calcário + K
18,6 a
18,0 b
19,3 a 9,8 b
22,2 a
*Testemunha: sem adição de K e calcário; NT (Nitossolo); LT (Latossolo);
CB (Cambissolo); AG (Argissolo); CP (Cambissolo sob cultivo de pinus).
**Números seguidos de letras iguais nas colunas para cada solo não diferem
estatisticamente pelo teste de Tukey (p < 0.05). Fonte: próprio autor, 2016.
Os efeitos prejudiciais da calagem observados no
primeiro cultivo se mantiveram no segundo cultivo, e foram
observadas diferenças em todos os solos (Tabela 2.3).
A calagem prejudicou a altura das plantas apenas no LT;
já no CP o efeito da calagem foi o inverso, tendo as mudas no
67
tratamento com K + calcário sido superiores às mudas da
testemunha.
A massa seca das raízes foi menor nos tratamentos com
calcário, em relação aos demais, apenas no LT e no AG.
O diâmetro das mudas no tratamento com K foi superior
a um dos tratamentos com calcário em todos os solos. Para os
solos NT e LT, o tratamento com K + calcário apresentou os
menores valores; os solos CP e AG tiveram os menores valores
no tratamento só com calcário, e no CB os dois tratamentos com
calcário proporcionaram diâmetro das mudas inferiores aos sem
calcário.
Os valores de massa seca da parte aérea foram superiores
na testemunha em relação aos do tratamento com K+calcário no
NT, LT e CB, e aos do tratamento só com calcário no AG. O
solo CP não apresentou diferença nesta variável.
De maneira geral, o desenvolvimento das plantas no
segundo cultivo foi inferior ao observado no primeiro cultivo,
mesmo tendo havido um tempo de condução três meses menor.
A calagem não é uma prática de uso generalizado nos
povoamentos florestais no Brasil, sendo praticada mais no
sentido de corrigir deficiências de Ca e Mg do que para elevar o
pH, já que o pinus é tolerante à acidez do solo (SMETHURST,
2010). Quando realizada, especialmente combinada com
adubação completa, pode resultar em incrementos no
crescimento inicial (LOPES et al., 1983) e também de plantas
adultas de pinus (ROSBERG et al., 2006). Em trabalhos
realizados em povoamentos de pinus, a calagem diminuiu os
teores de Cu, Zn, Ni e Mn na solução do solo, sendo que o teor
do último também decresceu nas acículas (DEROME;
SAARSALMI, 1999; GRONFLATEN et al., 2005). O P. taeda
parece ser uma espécie sensível a baixos teores de
micronutrientes, podendo resultar em redução no crescimento
em altura e volume do povoamento (VOGEL; JOKELA, 2011).
No presente estudo, os sintomas foram possivelmente causados
por deficiência de Mn.
68
Em cultivos no campo, a diminuição da disponibilidade
de micronutrientes pela calagem pode não resultar em
deficiência como observado no presente estudo. Como somente
a camada superficial do solo é corrigida, com aplicação de
calcário em superfície ou incorporada nos primeiros 20 cm, as
raízes crescem além desta camada e compensam a diminuição
da disponibilidade dos nutrientes. No cultivo a deficiência fica
mais evidente uma vez que todo o volume de solo explorado
pelas raízes foi corrigido, não havendo compensação da menor
disponibilidade.
Embora o K seja um nutriente entre os mais limitantes
(REISSMANN; WISNIEWSKI, 2004) e especialmente
importante para espécies florestais, já que a formação da
madeira depende do transporte de nutrientes através dos canais
de K (SMETHRUST et al., 2010), a adubação potássica ou a
omissão do nutriente normalmente não afeta o crescimento de
plantas do gênero Pinus (VOGEL et al., 2005). Segundo Crous
et al. (2008), a aplicação de K pode resultar em aumento de
volume apenas após o teor foliar atingir o nível crítico. O fósforo
é citado como o nutriente com maior efeito e resposta à
fertilização (SIMÕES, 1970; FLOR, 1977; VOGEL et al.,
2005).
69
Tabela 2.3 – Altura, diâmetro e massa seca de raiz e parte aérea
de mudas de pinus, no segundo cultivo, em função da adição de
K, calcário e da combinação de ambos, em cinco solos
catarinenses.
NT*
LT
CB
AG
Altura de mudas (cm)
a
56,5 a
55,7
ab 54,7 a
61,0
bc 53,5 a
54,0
c
49,2 a
58,2
Testemunha
Potássio
Calcário
Calcário + K
59,7
53,7
60,0
57,7
a**
a
a
a
62,7
58,7
50,5
46,2
Testemunha
Potássio
Calcário
Calcário + K
7,1
7,6
6,7
6,1
ab
a
ab
b
Diâmetro de mudas (cm)
7,1 ab 7,7 a
7,4
7,6 a
7,4 a
8,3
6,3 bc 5,9 b
5,7
5,1 c
5,6 b
6,6
Testemunha
Potássio
Calcário
Calcário + K
5,7
6,7
5,2
4,2
a
a
a
a
CP
a
a
a
a
55,3
57,5
59,7
65,5
b
ab
ab
a
ab
a
c
bc
7,8
8,1
6,8
8,0
ab
a
b
ab
Massa seca de raiz (g vaso-1)
7,7 a
7,2 a
7,5 ab
6,7 a
5,7 a
9,7 a
5,0 ab 4,7 a
4,7 b
3,2 b
4,5 a
6,2 b
6,7
7,5
6,5
7,8
a
a
a
a
Massa seca de parte aérea (g vaso-1)
Testemunha
20,9 a
20,1 a
19,4 a
19,4 a
16,6 a
Potássio
16,1 ab
16,9 ab 17,1 ab 19,9 a
19,9 a
Calcário
14,6 ab
11,9 bc 13,6 ab 10,1 b
17,9 a
Calcário + K
11,9 b
6,9 c
10,9 b
14,9 ab 22,9 a
*Testemunha: sem adição de K e calcário; NT (Nitossolo); LT (Latossolo);
CB (Cambissolo); AG (Argissolo); CP (Cambissolo sob cultivo de pinus).
**Números seguidos de letras iguais nas colunas para cada solo não diferem
estatisticamente pelo teste de Tukey (p < 0.05). Fonte: próprio autor, 2016.
2.3.1
pH E TEORES DE NUTRIENTES NO SOLO
O pH no solo após o primeiro cultivo, tanto na porção
rizosférica quanto não rizosférica, foi maior nos tratamentos que
receberam a calagem, resultando em diminuição dos teores de
Al trocável (Tabela 2.4). Em alguns solos, o pH alcançou valores
acima de 6,0, o que diminuiu a disponibilidade dos
70
micronutrientes, como o Mn, que resultou nos prejuízos no
crescimento observados nas tabelas 2.2 e 2.3. Nos tratamentos
sem adição de calcário, o pH da rizosfera foi superior ao não
rizosférico apenas no NT, LT e AG, mas não diferiu nos
Cambissolos. Já nos tratamentos com calagem, o pH da rizosfera
foi superior ao não rizosférico em todos os solos. Este resultado
não era esperado, pois o pinus é considerada uma cultura que
acidifica o solo, e cuja rizosfera normalmente é mais ácida que
o restante do solo (LOPES et al., 1984). Porém, em outras
culturas, foi constatado que o pH da rizosfera pode aumentar
próximo das raízes quando o pH do solo é baixo (YOUSSEF;
CHINO, 1988).
71
Tabela 2.4 - Valores de pH e teores de Al (cmolc dm-3) nas
porções rizosférica (R) e não rizosférica (NR) do solo após o
primeiro cultivo de pinus, em função da adição de K, calcário e
da combinação de ambos, em cinco solos catarinenses.
Testemunha
K
Calcário
Calc + K
pH
NT*
NR
4,8
b B**
4,7 b B
5,7
aB
5,9
aB
R
5,4
bA
5,5 b A 6,4
aA
6,7 a A
LT
NR
5,4
bB
4,9
cB
6,1
aB
6,1
aB
R
5,8
cA
5,1 d A 6,4 b A 6,9 a A
CB
NR
5,1
cA
4,8 d A 5,6 b B 5,9
aB
R
5,0
bA
4,7 c A
6,1 a A
6,2 a A
AG
NR
5,8
bB
5,5 b B
6,4
aB
6,7
aB
R
6,0
bA
6,2 b A 7,3
aA
7,3 a A
CP
NR
5,2
bA
4,9 b A 6,1
aB
6,1
aB
R
5,3
bA
5,1 b A 6,8
aA
6,5 a A
Al (cmolc dm-3)
NT
NR
1,1
aB
1,6
aB
0,0 b A 0,0 b A
R
1,9
aA
2,3 a A
0,4 b A 0,5 b A
LT
NR
1,9
aB
1,9
aB
0,0 b B
0,0 b A
R
2,5
aA
3,1 a A
0,6 b A 0,5 b A
CB
NR
5,6
aB
5,5
aB
0,0 b A 0,0 b A
R
8,2
aA
8,2 a A
0,1 b A 0,1 b A
AG
NR
0,2
aB
0,3 a A
0,0 b A 0,0 b B
R
0,4
aA
0,3 a A
0,0 b A 0,1 b A
CP
NR
7,4
aB
7,0
aB
0,0 b A 0,0 b A
R
9,9
aA
9,4 a A
0,1 b A 0,2 b A
* Testemunha: sem adição de K e calcário; NT (Nitossolo); LT (Latossolo);
CB (Cambissolo); AG (Argissolo); CP (Cambissolo sob cultivo de pinus).
**Números seguidos de letras iguais não diferem estatisticamente pelo teste
de Tukey (p <0,05). Letras minúsculas comparam tratamentos e letras
maiúsculas comparam solo rizosférico e não rizosférico, dentro de cada solo.
Fonte: próprio autor, 2016.
Após o segundo cultivo, o pH permaneceu maior nos
dois tratamentos que receberam calcário em relação aos demais,
tanto na porção rizosférica quanto não rizosférica do solo, para
todos os solos, com exceção do CP (Tabela 2.5). Os valores
72
obtidos na rizosfera foram superiores aos do solo não rizosférico
em todos os tratamentos no AG; em todos os tratamentos, exceto
na testemunha, no NT e LT, nos tratamentos que receberam
calcário no CB, mas no CP não houve diferença entre as porções.
Em relação ao primeiro cultivo, o pH foi maior nos tratamentos
que receberam calcário, indicando que este continuou reagindo
mesmo após um ano e meio da aplicação.
Os teores de Al diminuíram seguindo o aumento do pH,
com os menores valores observados nos tratamentos com adição
de calcário. No AG não houve diferença entre os tratamentos,
pois o pH foi elevado em todos os tratamentos. Apesar da
diferença observada no pH em todos os solos, apenas no LT e
no AG os valores de Al foram menores na rizosfera do que no
solo não rizosférico.
As alterações no pH são as modificações químicas mais
documentadas na rizosfera. Em solos florestais, o pH na
rizosfera pode ser significativamente menor em função da
liberação de ácidos orgânicos e respiração de raízes e da
microflora, exsudação e liberação de prótons (CALVARUSO et
al., 2014). O efeito mais proeminente se dá pela absorção
diferenciada de nitrato ou amônio, que representam 70% de
todos os ânions e cátions absorvidos pelas plantas. Uma maior
taxa de absorção de cátions comparada com a de ânions leva a
perda de prótons e à acidificação da rizosfera, enquanto que a
maior taxa de absorção de ânions, comparada com a de cátions,
leva ao consumo de H+, além da exsudação de OH- e HCO3-, e
isso resulta no aumento do pH da rizosfera (MARSCHNER,
1995).
73
Tabela 2.5 - Valores de pH e teores de Al (cmolc dm-3) nas
porções rizosférica (R) e não rizosférica (NR) do solo após o
segundo cultivo de pinus em função da adição de K, calcário e
da combinação de ambos, em cinco solos catarinenses.
Testemunha
K
Calcário
Calc + K
pH
NT* NR
5,7
b A** 5,5 b B
6,5
aB
6,5
aB
R
5,9
bA
5,8 b A
7,5
aA
7,4
aA
LT
NR
5,8
bA
5,6 b B
6,5
aB
6,6
aB
R
5,9
cA
5,9 c A
7,6
aA
7,2
bA
CB
NR
5,3
bA
5,3 b A
5,9
aB
6,1
aB
R
5,5
bA
5,5 b A
6,7
aA
6,7
aA
AG
NR
6,1
bB
6,2 b B
7,0
aB
7,0
aB
R
6,5
bA
6,6 b A
7,9
aA
7,8
aA
CP
NR
5,2
aA
5,2 a A
6,6
aA
6,5
aA
R
5,4
aA
5,5 a A
7,0
aA
5,2
aA
Al (cmolc dm-3)
NT
NR
0,4
ab A
0,4 a A
0,1
b A 0,2
ab A
R
0,3
aA
0,3 a A
0,0
b A 0,0
bA
LT
NR
0,7
aA
0,8 a A
0,2
b A 0,2
bA
R
0,4
aB
0,4
aB
0,0
bB
0,0
bB
CB
NR
3,3
aA
3,7 a A
0,3
b A 0,3
bA
R
3,3
aA
3,8 a A
0,0
b A 0,0
bA
AG
NR
0,3
aA
0,2 a A
0,2
aA
0,2
aA
R
0,0
aB
0,0
aB
0,0
aB
0,0
aB
CP
NR
5,4
aA
5,2 a A
0,3
b A 0,3
bA
R
4,8
aB
4,8 a A
0,0
b A 0,0
bA
* Testemunha: sem adição de K e calcário; NT (Nitossolo); LT (Latossolo);
CB (Cambissolo); AG (Argissolo); CP (Cambissolo sob cultivo de pinus).
**Números seguidos de letras iguais não diferem estatisticamente pelo teste
de Tukey (p <0,05). Letras minúsculas comparam tratamentos e letras
maiúsculas comparam solo rizosférico e não rizosférico, dentro de cada solo.
Fonte: próprio autor, 2016.
Os teores de Ca e Mg, após o primeiro cultivo, também
foram afetados pelos tratamentos (Tabela 2.6), sendo superiores
nos tratamentos que receberam calagem, já que o calcário
fornece esses dois nutrientes. Em povoamentos de pinus já
estabelecidos, a aplicação de calcário aumentou os teores de Ca
74
e Mg nas camadas superficiais do solo, mesmo com aplicação a
lanço, sem incorporação (BATISTA et al., 2015).
Os teores de Ca na porção não rizosférica foram maiores
do que os da rizosfera nos tratamentos com adição de calcário,
mas não diferiram entre as frações nos demais tratamentos.
Apenas no CB os teores no solo não rizosférico foram superiores
em todos os tratamentos. Para os teores de Mg, também foram
observadas diferenças entre as frações apenas nos tratamentos
com calcário. Diferentemente do observado para o Ca, os
maiores teores foram observados na rizosfera. Considerando que
Ca e Mg são transportados no solo por fluxo de massa, era
esperado que os teores na rizosfera fossem superiores para
ambos, já que este tipo de transporte leva até as raízes
quantidades maiores do que a planta absorve (BARBER et al.,
1963). É possível que o efeito tenha sido observado apenas para
o Mg em função da maior mobilidade deste no solo em
comparação ao Ca.
75
Tabela 2.6 - Teores de Ca e Mg (cmolc dm-3) nas porções
rizosférica (R) e não rizosférica (NR) do solo após o primeiro
cultivo de pinus em função da adição de K, calcário e da
combinação de ambos, em cinco solos catarinenses.
Testemunha
K
Calcário
Calc + K
Ca (cmolc dm-3)
NT* NR 1,38 b A** 1,28 b A 6,54 a A 6,21 a A
R
1,25
bA
1,22 b A 4,34 a B 4,42 a B
LT
NR 2,09
cA
2,09 c A 7,77 b A 8,38 a A
R
2,32
bA
2,21 b A 5,09 a B 5,09 a B
CB
NR 1,11
cA
1,05 c A 7,31 a A 7,07 b A
R
0,81
bB
0,78 b B 4,77 a B 4,77 a B
AG NR 1,87
bA
1,85 b A 5,29 a A 5,42 a A
R
1,80
bA
1,79 b A 3,79 a B 3,88 a B
CP
NR 0,97
bA
1,07 b A 8,19 a A 7,54 a A
R
0,97
bA
1,03 b A 5,10 a B 5,11 a B
Mg (cmolc dm-3)
NT
NR 1,04
bA
0,84 b A 6,03 a A 5,51 a B
R
0,87
bA
0,82 b A 6,37 a A 6,11 a A
LT
NR 1,36
bA
1,40 b A 6,47 a B 7,28 a B
R
1,56
bA
1,63 b A 8,47 a A 8,33 a A
CB
NR 0,54
bA
0,48 b A 6,43 a B 6,15 a B
R
0,39
bA
0,37 b A 7,51 a A 7,46 a A
AG NR 1,28
bA
1,23 b A 4,33 a A 4,04 a B
R
1,38
bA
1,44 b A 4,29 a A 4,60 a A
CP
NR 0,31
bA
0,37 b A 6,81 a B 7,27 a B
R
0,41
bA
0,39 b A 8,84 a A 8,93 a A
* Testemunha: sem adição de K e calcário; NT (Nitossolo); LT (Latossolo);
CB (Cambissolo); AG (Argissolo); CP (Cambissolo sob cultivo de pinus).
**Números seguidos de letras iguais não diferem estatisticamente pelo teste
de Tukey (p <0,05). Letras minúsculas comparam tratamentos e letras
maiúsculas comparam solo rizosférico e não rizosférico, dentro de cada solo.
Fonte: próprio autor, 2016.
Os teores de Ca e Mg, após o segundo cultivo,
permaneceram maiores nos dois tratamentos que receberam
aplicação de calcário em comparação com a testemunha e com
o tratamento com aplicação de potássio, tanto na porção
rizosférica como na não rizosférica do solo (Tabela 2.7). Os
76
teores de Ca diferiram entre rizosfera e solo não rizosférico
apenas nos tratamentos com aplicação de calcário, devido à sua
maior disponibilidade no solo. Os teores de Mg na rizosfera
foram superiores nos tratamentos com aplicação de calcário no
NT, CB e CP, em todos os tratamentos no LT, mas não diferiram
no AG.
Comparando visualmente os teores após o primeiro e o
segundo cultivos, verifica-se pequena variação, sendo mais
evidente a diminuição nos tratamentos com calcário, o que era
esperado, considerando que houve absorção pelas plantas e não
foi feita aplicação destes nutrientes entre um cultivo e outro.
Em condições de campo, os teores de Ca e Mg tendem a
ser maiores na rizosfera do que no restante do solo, porém essa
condição varia de acordo com a estação do ano. Durante a
primavera, apesar de haver maior absorção da planta, por estar
em período de crescimento, a maior atividade biológica nesse
período possibilita o maior acúmulo de nutrientes na rizosfera,
por favorecer a decomposição da matéria orgânica e o
intemperismo de minerais (CALVARUSO et al., 2014).
77
Tabela 2.7 - Teores de Ca e Mg (cmolc dm-3) nas porções
rizosférica e não rizosférica do solo após o segundo cultivo de
pinus, em função da adição de K, calcário e da combinação de
ambos, em cinco solos catarinenses.
Testemunha
K
Calcário
Calc + K
Ca (cmolc dm-3)
NT* NR 1,35 b A**
1,27 b A 5,12 a A 5,15 a A
R
1,27
bA
1,15 b A 4,45 a B 4,67 a B
LT
NR 2,00
bA
1,95 b B 6,92 a A 6,77 a A
R
2,12
bA
2,42 b A 5,92 a B 6,27 a B
CB
NR 1,15
bA
1,07 b A 5,85 a A 6,00 a A
R
1,12
bA
1,07 b A 5,22 a B 5,27 a B
AG NR 1,47
bA
1,42 b A 3,80 a A 4,02 a A
R
1,37
bA
1,27 b A 3,52 a B 3,47 a B
CP
NR 1,02
bA
1,05 b A 6,75 a A 7,02 a A
R
0,92
bA
0,90 b A 5,97 a B 5,97 a B
Mg (cmolc dm-3)
NT
NR 1,30
bA
1,22 b A 4,42 a B 4,47 a B
R
1,47
bA
1,37 b A 4,92 a A 5,02 a A
LT
NR 1,62
aB
1,60 b B 5,00 a B 4,80 a B
R
2,05
bA
2,12 b A 5,65 a A 5,97 a A
CB
NR 1,07
bA
1,02 b A 5,00 a B 4,92 a B
R
1,10
bA
1,07 b A 5,65 a A 5,72 a A
AG NR 1,45
bA
1,45 b A 3,55 a A 3,65 a A
R
1,57
bA
1,42 b A 3,67 a A 3,77 a A
CP
NR 0,67
bB
0,95 b A 4,65 a B 4,75 a B
R
0,95
bA
0,90 b A 5,80 a A 5,77 a A
* Testemunha: sem adição de K e calcário; NT (Nitossolo); LT (Latossolo);
CB (Cambissolo); AG (Argissolo); CP (Cambissolo sob cultivo de pinus).
**Números seguidos de letras iguais não diferem estatisticamente pelo teste
de Tukey (p <0,05). Letras minúsculas comparam tratamentos e letras
maiúsculas comparam solo rizosférico e não rizosférico, dentro de cada solo.
Fonte: próprio autor, 2016.
Para todos os solos estudados, os teores de K na solução
do solo, após o primeiro cultivo, foram maiores nos tratamento
com adição de K combinado com calcário em comparação com
um ou mais tratamentos (Tabela 2.8). Na porção não rizosférica
do solo, no Latossolo e nos Cambissolos, o tratamento com
78
K+calcário foi superior a todos os tratamentos, enquanto no
Nitossolo e no Argissolo, ele foi apenas superior à testemunha.
Essa diferença entre os solos pode ter ocorrido em função dos
maiores teores atingidos no Nitossolo e no Argissolo em relação
aos demais, em todos os tratamentos. No solo rizosférico, apenas
no Latossolo houve efeito de tratamento, onde o tratamento com
K+calcário permaneceu com maiores valores em relação aos
demais.
Como a calagem promove o aumento do número de
cargas negativas livres no complexo de troca, era esperado que
o teor de K na solução diminuísse, aumentando a quantidade
adsorvida (K extraível), uma vez que as formas de K no solo
estão em equilíbrio. Como os teores aumentaram, é possível que
a competição do Ca e Mg adicionado com o calcário pelas cargas
tenha mantido o K na solução, embora esse efeito seja
considerado menor do que o do aumento das cargas.
Em todos os solos avaliados, os teores observados na
rizosfera foram menores do que na porção não rizosférica em
pelo menos um tratamento. No Latossolo e nos Cambissolos, a
diferença foi observada somente no tratamento com K+calcário,
que apresentou os maiores valores. No Nitossolo e no Argissolo,
com exceção da testemunha, em todos os tratamentos foram
encontrados menores teores na rizosfera.
Para nutrientes transportados majoritariamente por
difusão até as raízes, como o K, a superfície radicular e a
rizosfera tendem a apresentar menor concentração em relação ao
restante do solo, em função da formação de uma zona de
depleção no entorno das raízes causada pela alta taxa de
absorção em relação ao transporte até as raízes (BARBER et al.,
1963). Entretanto, Calvaruso et al. (2014) encontraram maiores
teores de K na solução do solo na rizosfera de diferentes espécies
florestais, atribuindo o fato a um balanço positivo entre entradas
(intemperismo de minerais, fluxo de massa) e saídas (absorção).
79
Tabela 2.8 – Valores de K na solução nas porções rizosférica e
não rizosférica do solo após o primeiro cultivo de pinus, em
função da adição de K, calcário e da combinação de ambos, em
cinco solos catarinenses.
Testemunha
K
Calcário
Calc + K
K na solução (mg kg-1)
NT* NR
1,7
b A** 3,3 ab A 2,7 ab A 4,4 a A
R
0,8
aA
1,1
aB
0,9
aB
1,8 a B
LT
NR
1,1
bA
2,1
bA
1,9
bA
4,6 a A
R
0,2
bA
1,1
bA
1,1
bA
2,7 a B
CB
NR
1,5
bA
1,1
bA
1,6
bA
3,6 a A
R
1,1
aA
1,5
aA
1,2
aA
1,7 a B
AG
NR
1,8
cA
4,0
bA
5,2
bA
7,5 a A
R
1,1
aA
1,5
aB
1,7
aB
2,8 a B
CP
NR
1,3
bA
1,9
bA
2,3
bA
3,8 a A
R
1,1
aA
1,2
aA
1,5
aA
1,6 a B
* Testemunha: sem adição de K e calcário; NT (Nitossolo); LT (Latossolo);
CB (Cambissolo); AG (Argissolo); CP (Cambissolo sob cultivo de pinus).
**Números seguidos de letras iguais não diferem estatisticamente pelo teste
de Tukey (p <0,05). Letras minúsculas comparam tratamentos e letras
maiúsculas comparam solo rizosférico e não rizosférico, dentro de cada solo.
Fonte: próprio autor, 2016.
A adição de potássio, combinada ou não com a aplicação
de calcário, aumentou os teores de K extraível no solo não
rizosférico em 4 dos 5 solos: NT, LT, CB e AG (Tabela 2.9). No
CB, o aumento ocorreu apenas quando da aplicação de
K+calcário. O efeito na rizosfera não foi tão evidente, sendo o
tratamento com adição de potássio superior à testemunha e ao
tratamento com calagem no NT, e superior no tratamento com
K+calcário em relação à testemunha e o tratamento com
calagem no CB e AG.
Comparando os teores encontrados no solo não
rizosférico e na rizosfera, no NT os teores foram menores na
rizosfera; no CB e AG os teores foram maiores na rizosfera; e
no LT e CP não houve diferença entre as duas porções. Os
resultados em sentidos opostos observados nos diferentes solos
80
podem ser efeito da diferença da concentração do nutriente nos
solos, da passagem do K extraível para outras formas, ou da
liberação de formas estruturais à medida que ocorreu a absorção.
Tabela 2.9 - Valores de K extraível (Mehlich 1) nas porções
rizosférica e não rizosférica do solo após o primeiro cultivo de
pinus, em função da adição de K, calcário e da combinação de
ambos, em cinco solos catarinenses.
Testemunha
K
Calcário
Calc + K
K Mehlich (mg kg-1)
NT* NR 49,3 b A** 70,5 a A 38,7 b A 70,5 a A
R
49,0
bA
69,3 a A 45,4 b A 55,7 ab B
LT
NR 33,7
bA
54,2 a A 28,7 b A 56,3 a A
R
42,5
aA
58,0 a A 42,8 a A 60,2 a A
CB
NR 36,7
bB
53,0 a A 34,5 b B 58,5 a A
R
42,5
cA
48,9 b A 46,5 bc A 56,9 a A
AG NR 30,5
bB
45,0 a B 32,0 b B 48,7 a B
R
46,4
cA
54,9 ab A 52,4 bc A 60,2 a A
CP
NR 54,5
cA
70,0 ab A 59,0 bc A 76,5 a A
R
61,2
aA
67,8 a A 60,8 a A 68,8 a A
* Testemunha: sem adição de K e calcário; NT (Nitossolo); LT (Latossolo);
CB (Cambissolo); AG (Argissolo); CP (Cambissolo sob cultivo de pinus).
**Números seguidos de letras iguais não diferem estatisticamente pelo teste
de Tukey (p <0,05). Letras minúsculas comparam tratamentos e letras
maiúsculas comparam solo rizosférico e não rizosférico, dentro de cada solo.
Fonte: próprio autor, 2016.
A adubação potássica influenciou os teores de K não
trocáveis, de forma bastante variável entre os solos (Tabela
2.10). Foram observados efeitos de tratamento no solo não
rizosférico somente no Argissolo e no Latossolo, com os
maiores valores no tratamento K+calcário e testemunha,
respectivamente. Nestes solos e também no Nitossolo houve
diferenças na rizosfera, com valores maiores nos tratamentos
com aplicação de K e K+calcário em comparação com a
testemunha. Quando comparados os teores no solo rizosférico e
não rizosférico, somente no Cambissolo sob cultivo de pinus os
81
teores da rizosfera foram menores; não houve diferença no
Nitossolo; e nos demais os teores da rizosfera foram maiores em
pelo menos um tratamento.
Tabela 2.10 - Valores de K não trocável (HNO3) (mg kg-1) nas
porções rizosférica e não rizosférica do solo após o primeiro
cultivo de pinus, em função da adição de K, calcário e da
combinação de ambos, em cinco solos catarinenses.
Testemunha
K
Calcário
Calc + K
K HNO3 (mg kg-1)
NT* NR 72
a A**
94
aA
76
aA
87
aA
R
65
bA
104
aA
57
bA
69 ab A
LT
NR 108
aB
85
ab A
77
bA
86 ab B
R
84
bA
101 ab A
95
ab A 123 a A
CB
NR 145
aB
154
aB
140
aB
156 a A
R
173
aA
180
aA
169
aA
178 a A
AG
NR 72
bA
78
bB
77
bA
99
aA
R
75
bA
96
aA
81
ab A 101 a A
CP
NR 402
aA
399
aA
418
aA
419 a A
R
151
aB
139
aB
148
aB
144 a B
* Testemunha: sem adição de K e calcário; NT (Nitossolo); LT (Latossolo);
CB (Cambissolo); AG (Argissolo); CP (Cambissolo sob cultivo de pinus).
**Números seguidos de letras iguais não diferem estatisticamente pelo teste
de Tukey (p <0,05). Letras minúsculas comparam tratamentos e letras
maiúsculas comparam solo rizosférico e não rizosférico, dentro de cada solo.
Fonte: próprio autor, 2016.
Após o segundo cultivo, os teores de K na solução
continuaram apresentando comportamento semelhante ao
observado no primeiro cultivo, com o tratamento contendo K +
calcário apresentando os maiores valores em relação a um ou
mais tratamentos, no solo não rizosférico (Tabela 2.11). Já na
rizosfera, foram observadas diferenças somente no solo AG,
onde o tratamento com K + calcário foi maior ao tratamento
testemunha. O solo não rizosférico apresentou teores superiores
à rizosfera em todos os solos.
82
Tabela 2.11– Valores de K na solução nas porções rizosférica e
não rizosférica do solo após o segundo cultivo de pinus em
função da adição de K, calcário e da combinação de ambos, em
cinco solos catarinenses.
Testemunha
K
Calcário
Calc + K
K na solução (mg kg-1)
NT* NR
1,1
c A** 2,9
bA
2,3 bc A 8,2 a A
R
0,7
aA
1,3
aB
1,3
aA
2,1 a B
LT
NR
1,1
bA
1,8
bA
2,1
bA
6,1 a A
R
0,8
aA
1,3
aA
1,1
aB
1,5 a B
CB
NR
1,7
bA
2,8
bA
2,2
bA
4,7 a A
R
0,7
aB
0,9
aB
1,2
aB
1,4 a B
AG
NR
5,3
aA
3,3 ab A 2,6
bA
5,4 a A
R
1,1
bB
1,6 ab B 1,8 ab A 3,5 a B
CP
NR
2,4
cA
4,9
bA
4,2 bc A 7,0 a A
R
1,6
aA
1,5
aB
1,7
aB
2,0 a B
* Testemunha: sem adição de K e calcário; NT (Nitossolo); LT (Latossolo);
CB (Cambissolo); AG (Argissolo); CP (Cambissolo sob cultivo de pinus).
**Números seguidos de letras iguais não diferem estatisticamente pelo teste
de Tukey (p <0,05). Letras minúsculas comparam tratamentos e letras
maiúsculas comparam solo rizosférico e não rizosférico, dentro de cada solo.
Fonte: próprio autor, 2016.
Os teores de K extraível, após o segundo cultivo, também
apresentaram comportamento semelhante ao obtido após o
primeiro cultivo, sendo que a adição de K combinada com
calcário apresentou os maiores valores em todos os solos
(Tabela 2.12). No NT, LT e CP, a diferença ainda se manteve no
tratamento com K. Houve diferença entre a rizosfera e o solo não
rizosférico somente no LT e no CP.
83
Tabela 2.12 - Valores de K extraível (Mehlich 1) nas porções
rizosférica e não rizosférica do solo após o segundo cultivo de
pinus em função da adição de K, calcário e da combinação de
ambos, em cinco solos catarinenses.
Testemunha
K
Calcário
Calc + K
K Mehlich (mg kg-1)
NT* NR 34,2 b A** 76,5 a A 35,5 b A 70,2 a A
R
39,6
bA
72,6 a A 44,3 b A 76,7 a A
LT
NR 27,5
bB
54,2 a B 32,0 b A 73,5 a A
R
44,5
bA
81,6 a A 41,1 b A 81,0 a A
CB
NR 31,0
cA
49,7 b A 32,7 c A 65,0 a A
R
36,7
bA
46,1 b A 38,7 b A 61,5 a A
AG
NR 22,2
cA
39,5 b A 29,2 c A 48,7 a A
R
23,8
cA
40,8 b A 31,5 c A 49,4 a A
CP
NR 52,5
bA
81,0 a A 58,0 b A 85,5 a A
R
53,8
bA
76,9 a A 53,8 b A 76,0 a B
* Testemunha: sem adição de K e calcário; NT (Nitossolo); LT (Latossolo);
CB (Cambissolo); AG (Argissolo); CP (Cambissolo sob cultivo de pinus).
**Números seguidos de letras iguais não diferem estatisticamente pelo teste
de Tukey (p <0,05). Letras minúsculas comparam tratamentos e letras
maiúsculas comparam solo rizosférico e não rizosférico, dentro de cada solo.
Fonte: próprio autor, 2016.
Para o Knt, os tratamento com aplicação de K,
combinada ou não com a calagem, resultaram nos maiores teores
em todos os solos exceto o Cambissolo sob cultivo de pinus,
onde não houve diferença (Tabela 2.13). As diferenças entre solo
rizosférico e não rizosférico foram bastante variadas, diferindo
entre solos e entre tratamentos em um mesmo solo.
O aumento dos teores de Knt no tratamento com
K+calcário pode ser devido à aplicação de K, e também devido
ao efeito do aumento do pH na precipitação de hidróxidos de Fe
e Al que estavam bloqueando as entrecamadas dos
argilominerais, permitindo a liberação do K na posição não
trocável (DAS; SAHA, 2013).
84
Tabela 2.13 - Valores de K não trocável (HNO3) (mg kg-1) nas
porções rizosférica (R) e não rizosférica (NR) do solo após o
segundo cultivo de pinus em função da adição de K, calcário e
da combinação de ambos, em cinco solos catarinenses.
Testemunha
K
Calcário
Calc + K
K HNO3 (mg kg-1)
NT* NR 50
b A**
83
aA
49
bA
63 ab B
R
51
cA
83
ab A
67
bc A 100 a A
LT
NR 61
bB
106
aA
98
aA
116 a A
R
99
ab A
122
aA
79
bB
107 a A
CB
NR 151
bc A
187
aA
130
cB
164 ab A
R
147
bA
162 ab B 156 ab A 181 a A
AG NR 70
bA
87
ab A
85
ab A 104 a A
R
73
bA
105
aA
99
ab A 124 a A
CP
NR 402
aA
401
aA
444
aA
403 a A
R
384
aA
384
aA
388
aA
369 a A
* Testemunha: sem adição de K e calcário; NT (Nitossolo); LT (Latossolo);
CB (Cambissolo); AG (Argissolo); CP (Cambissolo sob cultivo de pinus).
**Números seguidos de letras iguais não diferem estatisticamente pelo teste
de Tukey (p <0,05). Letras minúsculas comparam tratamentos e letras
maiúsculas comparam solo rizosférico e não rizosférico, dentro de cada solo.
Fonte: próprio autor, 2016.
Apesar da diferença de mineralogia e condição química
inicial dos solos ser bastante variada (Tabela 1.3), em muitos
casos o comportamento dos atributos em função dos tratamentos
foi semelhante.
Comparando a condição do solo após o primeiro e o
segundo cultivo, houve pouca variação nos teores de um cultivo
para o outro, mesmo nos tratamentos onde não foi feita a
aplicação de K, com exceção do Ks no CP, que apresentou
redução considerável de um cultivo para o outro.
As formas prontamente disponíveis de K em plantios de
pinus tendem a manter um teor relativamente constante ao longo
do tempo, pela combinação da liberação do nutriente pelo
intemperismo de minerais e pela ciclagem rápida ocasionada por
lixiviação do dossel da planta e da serapilheira (ABRÃO et al.,
2015). Porém, se não houver reposição do K removido pelas
85
plantas, as formas não trocáveis tendem a reduzir com os
cultivos sucessivos (KAMINSKI et al., 2007).
De maneira geral, o tratamento que resultou nos maiores
teores das diferentes formas de K foi o tratamento com
K+calcário, sendo sempre superior a um ou mais tratamentos,
em todos os solos.
O comparativo entre a rizosfera e o solo não rizosférico
foi variável entre os tratamentos e entre os solos. No NT e LT,
os teores de Ks e Knt se correlacionaram negativamente na
rizosfera, pois, nos casos em que houve diferença, quando o Ks
na rizosfera era maior que no solo não rizosférico, o Knt era
menor (Tabelas 2.8 a 2.13). O mesmo comportamento foi
observado no AG, porém o Ks foi menor e o Knt maior na
rizosfera em comparação ao solo não rizosférico. Esse resultado
pode indicar que ocorre a liberação das formas não trocáveis
para restabelecer o equilíbrio químico na rizosfera.
Já no CB, os teores, tanto de Ks quanto de Knt, foram
maiores na rizosfera em comparação ao solo não rizosférico, no
primeiro cultivo, e menores, no segundo. No CP, nos casos onde
houve diferença, os teores de K na rizosfera foram sempre
menores.
Segundo Marschner (1995b) e Hinsinger (1998), a
superfície radicular e a rizosfera apresentam menor
concentração de K em relação ao restante do solo, em função do
seu transporte por difusão que gera uma zona de depleção no
entorno das raízes. Entretanto Bath & Nye (1973) afirmam que
para o P, elemento igualmente transportado por difusão, a
depleção ocorre na região do rizocilindro, enquanto que a
concentração ao redor das raízes primárias tende a ser maior.
Além do transporte, a liberação das formas não trocáveis
também tem grande influência nos teores da rizosfera. Argila em
contato com solução de ácido acético (0,1 mol L-1) libera
quantidades significativas de K, mesmo com pouco tempo de
contato (MELO et al., 2005), indicando que os ácidos orgânicos
86
liberados pelas raízes podem aumentar a disponibilidade de K
pós cultivo.
A diferença no comportamento entre solos pode estar
relacionada com a maior quantidade de argilominerais 2:1
expansíveis nos Cambissolos do que nos outros solos avaliados.
A liberação das formas não trocáveis de K está diretamente
relacionada com a presença de minerais 2:1 (BRITZKE et al.,
2012), dessa forma alterando a dinâmica do equilíbrio no solo.
2.3.2 TEORES NA PLANTA
De maneira geral, os teores de K, Ca e Mg nas acículas
foram alterados pelos tratamentos (Tabela 2.14). Os teores de Ca
e Mg foram superiores nos tratamentos com calagem em todos
os solos. Os teores de K foram maiores nos tratamentos que
receberam adubação potássica nos solos CB, AG e CP; no LT
apenas o tratamento com K+calcário foi superior; e no NT
apenas o tratamento com calagem foi inferior à testemunha e ao
tratamento com K+calcário.
Os resultados diferem dos obtidos por Batista et al.
(2015), que observaram maiores teores de K nas acículas quando
não foi feita a aplicação de calcário, justificando que o Ca e o
Mg teriam efeito antagônico na absorção do K.
87
Tabela 2.14 - Teores de K, Ca e Mg nas acículas do primeiro
cultivo de pinus em função da adição de K, calcário e da
combinação de ambos, em cinco solos catarinenses.
NT*
Testemunha
Potássio
Calcário
Calcário + K
Testemunha
Potássio
Calcário
Calcário + K
12,89
11,55
7,92
13,34
0,29
0,46
1,20
1,62
a**
a
b
a
b
b
a
a
LT
CB
AG
CP
6,46
11,33
8,71
15,11
K (mg kg-1)
c 6,15 b 5,72
ab 9,42 a 9,05
bc 5,77 b 5,69
a 8,01 a 10,70
b 6,31
a 9,45
b 6,10
a 10,89
b
a
b
a
0,26
0,52
2,77
2,38
Ca (mg kg-1)
b 0,33 a
b 0,41 a
a 0,63 a
a 0,64 a
b
b
a
a
b
b
a
a
0,32
0,20
1,54
1,56
0,10
0,36
1,73
1,92
Mg (mg kg-1)
Testemunha
1,04 b
1,24 b 1,05 b 1,34 b 0,58 b
Potássio
0,96 b
1,24 b 0,97 b 0,99 b 0,69 b
Calcário
2,65 a
3,53 a 2,12 a 2,51 a 1,86 a
Calcário + K
2,43 a
3,48 a 1,98 a 2,64 a 1,80 a
*Testemunha: sem adição de K e calcário; NT (Nitossolo); LT (Latossolo);
CB (Cambissolo); AG (Argissolo); CP (Cambissolo sob cultivo de pinus).
**Números seguidos de letras iguais nas colunas para cada solo não diferem
estatisticamente pelo teste de Tukey (p < 0.05). Fonte: próprio autor, 2016.
Avaliando os teores no caule, os efeitos dos tratamentos
não foram tão evidentes quanto nas acículas (Tabela 2.15). Os
maiores teores de Mg nos tratamentos com calagem foram ainda
observados, com exceção para o NT onde não houve diferença
com relação à testemunha. O LT apresentou valores altos
enquanto o CP manteve os baixos valores, como observado nas
acículas.
Os teores de Ca foram maiores nos tratamentos com
calagem apenas no LT, não apresentando diferenças no CB, AG
e CP. Esse resultado está possivelmente relacionado com os
maiores teores iniciais de Ca no LT, que juntamente com o
nutriente disponibilizado no calcário resultou em maior
88
disponibilidade e consequentemente aumento nos teores na
planta.
O efeito dos tratamentos nos teores de K no caule pode
ser observado em todos os solos com exceção do NT, que não
apresentou diferenças. Nos demais solos, a aplicação de K
combinada com a calagem ou isolada elevou os teores no tecido.
Tabela 2.15- Teores de K, Ca e Mg no caule das mudas do
primeiro cultivo de pinus em função da adição de K, calcário e
da combinação de ambos, em cinco solos.
NT*
Testemunha
Potássio
Calcário
Calcário + K
Testemunha
Potássio
Calcário
Calcário + K
6,62
8,56
6,07
8,62
0,36
0,23
0,66
0,78
b**
a
b
a
ab
b
ab
a
LT
CB
AG
CP
K (g kg-1)
6,09 bc 4,70 bc
8,53 a
6,46 ab
5,23 c
4,36 c
7,55 ab 6,68 a
3,98
6,50
3,71
5,55
bc
a
c
ab
4,63
6,04
3,97
5,36
ab
Ca (g kg-1)
0,29 a
0,24 a
0,39 a
0,41 a
0,32
0,36
0,67
0,51
a
a
a
a
0,17
0,43
0,51
0,59
a
a
a
a
0,42
0,40
1,55
0,95
c
c
a
b
a
b
ab
Mg (g kg-1)
Testemunha
1,00 b
1,03 b
0,83 b
0,92 b
0,61 b
Potássio
0,88 b
0,95 b
0,78 b
1,04 b
0,67 b
Calcário
1,47 a
2,31 a
1,63 a
1,86 a
1,61 a
Calcário + K
1,42 a
2,05 a
1,64 a
1,64 a
1,44 a
*Testemunha: sem adição de K e calcário; NT (Nitossolo); LT (Latossolo);
CB (Cambissolo); AG (Argissolo); CP (Cambissolo sob cultivo de pinus).
**Números seguidos de letras iguais nas colunas para cada solo não diferem
estatisticamente pelo teste de Tukey (p < 0.05). Fonte: próprio autor, 2016.
Após o segundo cultivo, os teores de K, Ca e Mg nas
acículas continuaram mostrando efeito dos tratamentos, como
observado após o primeiro cultivo (Tabela 2.16). Os teores de K
foram maiores nos tratamentos com adição de K em todos os
solos, exceto no CP onde não houve diferença.
89
Os teores de Ca foram superiores nos tratamentos com
calagem no NT, LT e AG, mas nos Cambissolos não houve
diferença. Para o Mg, os teores foram maiores nos tratamentos
com calagem, em todos os solos.
Tabela 2.16- Teores de K, Ca e Mg nas acículas do segundo
cultivo de pinus nos em função da adição de K, calcário e da
combinação de ambos, em cinco solos.
NT*
Testemunha
Potássio
Calcário
Calcário + K
Testemunha
Potássio
Calcário
Calcário + K
6,29
8,13
5,69
9,46
1,09
1,06
2,03
2,03
bc**
ab
c
a
b
b
a
a
LT
CB
AG
CP
4,69
7,63
4,36
9,16
K (g kg-1)
b 4,31 b
a 6,48 a
b 4,21 b
a 6,69 a
3,66
5,94
4,24
6,59
c
ab
bc
a
5,86
6,04
4,60
5,79
a
a
a
a
1,08
1,01
2,02
2,89
Ca (g kg-1)
c 1,19 a
c 1,15 a
b 1,63 a
a 1,75 a
1,00
1,13
1,68
1,48
b
ab
a
ab
1,24
1,28
1,78
1,29
a
a
a
a
Mg (g kg-1)
Testemunha
1,58 b
1,41 b 1,28 b 1,27 b
1,01 c
Potássio
1,36 b
1,35 b 1,34 b 1,20 b
0,86 c
Calcário
2,71 a
2,23 a 2,35 a 1,86 a
2,06 a
Calcário + K
2,78 a
2,47 a 2,29 a 1,52 ab 1,63 b
*Testemunha: sem adição de K e calcário; NT (Nitossolo); LT (Latossolo);
CB (Cambissolo); AG (Argissolo); CP (Cambissolo sob cultivo de pinus).
**Números seguidos de letras iguais nas colunas para cada solo não diferem
estatisticamente pelo teste de Tukey (p < 0.05). Fonte: próprio autor, 2016.
Assim como observado no primeiro cultivo, após o
segundo cultivo os teores de K, Ca e Mg no caule seguiram o
mesmo comportamento do que foi observado nas acículas, com
maiores teores de K nos tratamentos com adubação potássica, e
maiores teores de Ca e Mg nos tratamentos que receberam
calagem (Tabela 2.17).
90
Tabela 2.17- Teores de K, Ca e Mg no caule das mudas do
segundo cultivo de pinus nos em função da adição de K, calcário
e da combinação de ambos, em cinco solos.
NT*
Testemunha
Potássio
Calcário
Calcário + K
Testemunha
Potássio
Calcário
Calcário + K
3,92
6,17
4,04
5,21
0,80
0,75
1,05
1,09
b**
a
b
ab
bc
c
ab
a
LT
3,61
5,39
3,49
4,79
0,68
0,63
1,14
1,43
CB
AG
CP
b
a
b
ab
K (g kg-1)
3,74 b
6,69 a
3,50 b
5,29 a
3,41
4,97
2,88
5,27
b
a
b
a
3,73
5,64
3,93
5,34
c
a
bc
ab
c
c
b
a
Ca (g kg-1)
0,67 b
0,72 ab
0,87 ab
0,94 a
0,65
0,69
0,90
0,72
a
a
a
a
0,74
0,69
0,87
0,73
a
a
a
a
Mg (g kg-1)
Testemunha
1,25 bc
1,08 c
1,07 b
1,23 a 0,89 b
Potássio
1,13 c
1,17 bc 1,10 b
1,18 a 0,91 b
Calcário
1,72 ab
1,71 b
1,87 a
1,60 a 1,67 a
Calcário + K
1,86 a
2,27 a
1,79 a
1,45 a 1,75 a
*Testemunha: sem adição de K e calcário; NT (Nitossolo); LT (Latossolo);
CB (Cambissolo); AG (Argissolo); CP (Cambissolo sob cultivo de pinus).
**Números seguidos de letras iguais nas colunas para cada solo não diferem
estatisticamente pelo teste de Tukey (p < 0.05). Fonte: próprio autor, 2016.
Em todos os solos, os teores de K na planta aumentaram
em função da adubação potássica, entretanto esses teores não
resultaram em incremento no crescimento das plantas, mesmo
naquelas do tratamento com K, que não sofreram os efeitos
negativos da calagem (Tabelas 2.2 e 2.3).
No CP, os efeitos da adubação não foram tão claros
quanto nos outros solos, pois houve diferença entre os
tratamentos com e sem a aplicação de K somente nas acículas
do primeiro cultivo, e no caule somente quando a adubação não
foi combinada com a calagem. No segundo cultivo, não houve
efeito. Esse comportamento diferenciado pode estar relacionado
com o predomínio de argilominerais 2:1 nesse solo (Capítulo 1),
91
que resultou em altos teores iniciais de K, especialmente os não
trocáveis. Dessa forma, a condição natural do solo se mostrou
eficaz em fornecer elevadas quantidades de K para as plantas,
não ocorrendo resposta à adubação.
Apesar da pequena variação nos teores de K no solo
entre os cultivos (Tabelas 2.8 a 2.13), os teores de K na planta,
tanto nas acículas quanto no caule, diminuíram do primeiro para
o segundo cultivo. Isso pode estar relacionado com a deficiência
nutricional causada pela elevação do pH que prejudicou o
desenvolvimento da planta como um todo. Além dos menores
teores, as plantas no segundo cultivo foram menores (Tabela
2.3), indicando que a absorção de K foi menor do que no
primeiro cultivo.
O teor foliar de K considerado ideal para o bom
desenvolvimento de plantas adultas de pinus é de 6,0 g kg-1
(SWITZER; NELSON, 1972; REISSMANN; WISNIEWSKI,
2000). Assim, mesmo nos tratamentos sem adição de K, as
plantas mantiveram um teor adequado no primeiro cultivo
(Tabela 2.14), estando acima ou próximo dessa faixa em todos
os solos. Já no segundo cultivo (Tabela 2.16), a queda nos teores
deixa as plantas em desequilíbrio nutricional, embora seja
comum o relato de árvores com teores semelhantes, 4,5 g kg-1
em média, sem sintomas de deficiência (ALBAUGH et al.,
2010; LUCIO et al., 2010). No trabalho de Batista et al. (2015),
a adubação potássica elevou os teores de K tanto no solo quanto
na planta, porém estes permaneceram abaixo do limite crítico
mesmo com alto teor do nutriente no solo.
Para o Ca, o ideal é que o teor seja de pelo menos 1,6 g
kg-1, sendo adequada a faixa entre 1,0 e 5,0 g kg-1 (SWITZER;
NELSON, 1972; ZÖTTL, 1973). O teor médio de Ca encontrado
em árvores de diferentes idades no Brasil é de 3,1 g kg-1 (LUCIO
et al., 2010), enquanto que nos Estados Unidos é de 1,8 g kg-1
(ALBAUGH et al., 2010), diferença justificada pela menor taxa
de crescimento das plantas na América do Norte. No presente
estudo, da mesma forma que foi observada para o K, os teores
92
de Ca foliares no primeiro cultivo estavam adequados, embora
próximos do limite inferior, enquanto que no segundo cultivo
não atingiram o mínimo necessário, mesmo nos tratamentos com
calagem em alguns solos.
Para o Mg, Chaves & Corrêa (2005) encontraram, nas
acículas de plantas de P. caribeae com adequado
desenvolvimento, teores médios de 0,4 g kg-1, sendo detectados
sinais de deficiência apenas quando o teor atingiu 0,1 g kg-1.
Entretanto, valores nesta faixa não são comuns, sendo
normalmente encontrados para P. taeda teores variando de 1,0 g
kg-1 a 1,5 g kg-1 (RUBILAR et al., 2005; VIERA;
SCHUMACHER, 2009; ALBAUGH et al., 2010; LUCIO et al.,
2010; LONDERO et al., 2011).
93
2.4
CONCLUSÕES
A calagem resulta em aumento do pH e dos teores de Ca
e Mg, e a adubação potássica aumenta os teores de K em todos
os solos, com efeito em todas as formas avaliadas.
Nas condições de solo avaliadas, as plantas de pinus não
apresentam incremento no crescimento inicial em resposta à
adubação potássica. Com exceção dos Cambissolos, nas demais
classes de solo avaliadas a calagem prejudica o crescimento das
mudas.
Em todos os solos avaliados, o pH na rizosfera é maior
do que no solo não rizosférico, em todos os tratamentos. Já os
teores de Ca e Mg diferem somente quando aplicado calcário,
sendo menor e maior na rizosfera, respectivamente.
O aumento dos teores dos nutrientes no solo aumenta os
teores de Ca, Mg e K nas acículas, mas esse aumento não se
reflete no crescimento das mudas.
A calagem e adubação potássica de mudas de pinus não
é indicada, pois não resulta em incrementos no crescimento.
94
2.5
REFERÊNCIAS BIBLIOGRÁFICAS
ABRÃO, S.F.; ROSA, S.F.; REINERT, D.J.; REICHERT,
J.M.; SECCO, D.; EBLING, A.A. Alterações químicas de um
Cambissolo Húmico causadas por reflorestamento com Pinus
taeda em área de campo natural. Floresta, Curitiba, v. 45, n. 3,
p. 455-464, 2015.
ALBAUGH, J.M.; BLEVINS, L.; ALLEN, H.L.; ALBAUGH,
T.J.; FOX, T.R.; STAPE, J.L.; RUBILAR, R.A.
Characterization of foliar macro and micronutrient
concentrations and ratios in loblolly pine plantations in the
Southeastern United States. South. J. Appl. For., v. 34, n. 2,
2010.
ALVES, M.J.F.; MELO, V.F.; REISSMANN, C.B. &
KASEKER, J.F. Reserva mineral de potássio em Latossolo
cultivado com Pinus taeda L. Revista Brasileira de Ciência
do Solo, 37: 1599- 1610, 2013.
ANDREWS, M.Y.; AGUE, J.J. & BERNER, R.A. Weathering
of soil minerals by angiosperm and gymnosperm trees.
Mineral Mag, 72:11-14, 2008.
BAKKER, M.R.; KERISIT, R.; VERBIST, K. & NYS, C.
Effects of liming on rhizosphere chemistry and growth of the
fine roots and of shoots of sessile oak (Quercus petraea). Plant
and Soil, 217: 243-255, 1999.
BAKKER, M.R.; GEORGE, E.; TURPAULT, M.-P.; ZHANG,
J.J. & ZELLER, B. impact of Douglas-fir and Scots pine
seedlings on plagioclase weathering under acidic conditions.
Plant and Soil, 266: 247-259, 2004.
95
BARBER, S.A.; WALKER, J.M.; VASEY, E.H. Mechanisms
for movement of plant nutrients from soil and fertilizer to plant
root. J. Agric. Food Chem., v. 11, n. 3, p. 204-207, 1963.
BATH, K.K.S. & NYE, P.H. Diffusion of phosphate to plant
roots in soil. I. Quantitative autoradiography of the depletion
zone. Plant and Soil, 38:161-175, 1973.
BATISTA, A.H.; MOTTA, A.C.V.; REISSMANN, C.B.;
SCHNEIDER, T.; MARTINS, I.L.; HASHIMOTO, M. Liming
and fertilization in Pinus taeda plantations with severe nutrient
deficiency in savanna soils. Acta Scientiarum Agronomy,
Maringá, v. 37, n. 1, p. 117-125, 2015.
BAZIMARAKENGA, B.;SIMARD, R. R. & LEUROX, G.D.
Determination of organic acids in soil extracts by ion
chromatography. Soil Biology and Biochemistry, 27:349-356,
1995.
BORTOLUZZI, E.C.; SANTOS, D.R. dos; KAMINSKI, J.;
GATIBONI, L.C. & TESSIER, D. Alterações na mineralogia
de um Argissolo do Rio Grande do Sul submetido à fertilização
potássica. R. Bras. Ci. Solo, 29:327-335, 2005.
BRITZKE, D.; SILVA, L.S.; MOTERLE, D.F.;
RHEINHEIMER, D.S.; BORTOLUZZI, E.C. A study of
potassium dynamics and mineralogy in soils from subtropical
Brazilian lowlands. J. Soils Sediments, v. 12, p. 185-197,
2012.
CALVARUSO, C.; TURPAULT, M.P. & FREY-KLETT,P.
Root-Associated Bacteria Contribute to Mineral Weathering
and to Mineral Nutrition in Trees: a Budgeting Analysis.
Applied Environmental Microbiology, 72: 1258–1266,
2006.
96
CALVARUSO, C.; COLLIGNON, C.; KIES, A.;
TURPAULT, M. Seasonal evolution of the rhizosphere effect
on major and trace elements in soil solutions of Norway Spruce
(Picea abies Karst) and Beech (Fagus sylvatica) in an acidic
forest soil. Open Journal of Soil Science, v. 4, p. 323-336,
2014.
CHAVES, R.Q.; CORRÊA, G.F. Macronutrientes no sistema
solo-Pinus caribaea em plantios apresentando amarelecimento
das acículas e morte de plantas. Revista Árvore, 29:691-700,
2005.
DAS, R.; SAHA, D. Changes in non-exchangeable K in an acid
and the corresponding limed soil as affected by addition of
nitrogenous and potassic fertilizers. Journal of Crop and
Wood, v. 9, n. 1, p. 99-102, 2013.
DEROME, J. & SAARSALMI, A. The effect of liming and
correction fertilisation on heavy metal and macronutrient
concentrations in soil solution in heavy-metal polluted scots
pine stands. Environmental Pollution, 104:249-259, 1999.
DINIZ, S.F.; BASTOS, F.O.M.; LIMA, R.H.C.; JIMENEZRUEDA, J.R. Fontes de potássio não trocável e potássio total
em quatro solos do Estado do Ceará. Geociências, São Paulo,
v.26, n. 4, p. 379-386, 2007.
ERNANI, P.R. & BARBER, S.A. Composição da solução do
solo e lixiviação de cátions afetadas pela aplicação de cloreto e
sulfato de cálcio em um solo ácido. Revista Brasileira de
Ciência do Solo, 10:41-46, 1993.
FLOR, H.M. Estudos sobre produção e forma do fuste de Pinus
taeda L. numa área de ensaio de adubação na Fazenda
Canguiri. Revista Floresta, 8(1): 69-70, 1977.
97
GEE, G.W. & BAUDER, J.W. Particle-size analysis. In:
KLUTE,A., ed. Methods of soil analysis. 2.ed. Madison,
American Society of Agronomy. p.383-412. 1986.
GOBRAN, G.R.; CLEGG, S. & COURCHESNE, F.
Rhizospheric processes influencing the biogeochemistry of
forest ecosystems. Biogeochemistry, 42: 107-120, 1998.
GRONFLATEN, L.K.; AMUNDSEN, L.; FRANK, J. &
STEINNES, E. Influence of liming and vitality fertilization on
trace element concentrations in Scots pine Forest soil and
plants. Forest Ecology and Management, 213: 261-271,
2005.
HINSINGER, P. How do plant roots acquire mineral nutrients?
Chemical processes involved in the rhizosphere. Advances in
Agronomy, 64: 225-265, 1998.
KAMINSKI, J.; BRUNETTO, G.; MOTERLE, D.F.;
RHEINHEIMER, D.S. Depleção de formas de potássio do solo
afetada por cultivos sucessivos. Revista Brasileira de Ciência
do Solo, v. 31, p. 1003-1010, 2007.
KNUDSEN, D.; PETERSON, G.A. & PRATT, P.F. Lithium,
sodium and potassium. In: PAGE, A. L. (Ed.), Methods of soil
analysis. Part 2. Chemical and microbiological properties,
American Society of Agronomy. Madison, p. 225-246, 1986.
KOELE, N.; HILDEBRAND, E.E. & SCHACK-KIRCHNER,
H. Effects of weathering state of coarse-soil fragments on treeseedling nutrient uptake. Journal of Plant Nutrition and Soil
Science, 173: 245-251, 2010.
98
LEYVAL, C. & BERTHELIN, J. Weathering of a mica by
roots and rhizospheric microorganisnms of pine. Soil Science
Society of America Journal, 55: 100-106, 1991.
LONDERO, E.K.; SCHUMACHER, M.V.; SZYMCZAK,
D.A. & VIERA, M. Exportação e reposição nutricional no
primeiro desbaste de um povoamento de Pinus taeda em área
de segunda rotação. Ciência Florestal, 21:487-497, 2011.
LOPES, M.I.M.S.; GURGEL, M.A. de O.; GARRIDO, L.M.
do A.G. & MELLO, F. de A.F. de. Adubação Mineral de Pinus
oocarpa Schiede. Anais da E.S.A. Luiz de Queiroz, 40:585601, 1983.
LOPES, M.I.M.S.; GARRIDO, M.A.O.; MELLO, F.A.F.
Influência do cultivo de Pinus sobre a manta orgânica e
propriedades químicas de um Latossolo Vermelho escuro
primitivamente sob vegetação de Cerrado. Anais da E.S.A.
Luiz de Queiroz, v. 41, p. 155-170, 1984.
LUCIO, A.D.C.; ROSSATO, R.A.R.; SCHUMACHER, M.V.;
FORTES, F.O.; STORCK, L.; WITSCHORECK, R. Limites de
precaução e de controle em análises nutricionais de espécies
florestais. Revista Árvore, Viçosa, v. 34, n. 3, p. 529-537,
2010.
MARSCHNER, H. Rizosphere pH effects on phosphorus
nutrition. In: Genetic manipulation of crop plants to
enhance integrated nutrient management in cropping
systems. p. 107-115. 1995.
MELO, V.F.; BARROS, N.F.; COSTA, L.M.; NOVAIS, R.F.
& FONTES, M.P.F. Formas de potássio e de magnésio em
solos do Rio Grande do Sul, e sua relação com o conteúdo na
planta e com a produção em plantio de eucalipto. R. Bras. Ci.
Solo, 19:165-167, 1995.
99
MELO, V.F. Potássio e magnésio em minerais de solos e
relação entre propriedades da caulinita com formas não
trocáveis destes nutrientes. Viçosa, MG, 1998. 205 f. Tese
(Doutorado em Solos e Nutrição de Plantas) – Setor de
Ciências Agrárias, Universidade Federal de Viçosa.
MELO, V.F.; CORRÊA, G.F.; MASCHIO, P.A.; RIBEIRO,
A.N. & LIMA, V.C. Importância das espécies minerais no
potássio total da fração argila de solos do triângulo mineiro. R.
Bras. Ci. Solo, 27:807-819, 2003.
MELO, V.F.; RIBEIRO, A.N.; MASCHIO, P.A.; CORRÊA,
G.F. & LIMA, V.C. Mineralogia e formas de potássio e
magnésio em diferentes classes de pesos e tamanhos da fração
areia de solos do triângulo mineiro. R. Bras. Ci. Solo, 28:219231, 2004.
MELO, V.F.; CORRÊA, G.F.; RIBEIRO, A.N.; MASCHIO,
P.A. Cinética de liberação de potássio e magnésio pelos
minerais da fração argila de solos do triângulo mineiro.
Revista Brasileira de Ciência do Solo, v. 29, p. 533-545,
2005.
MELO, V.F.; CASTILHOS, R.M.V. & PINTO, L.F.S. Reserva
mineral dos solo. In: MELO, V.F. & ALLEONI, L.R.F. (eds).
Química e Mineralogia do Solo, Parte I, SBCS. p. 251-332,
2009.
REATTO, A.; FONTES, M.P.F.; ALVAREZ V., V.H.;
RESENDE, M.; KER, J.C. & COSTA, L.M. Caracterização
Mineralógica, potencial de reserva e sustentabilidade agrícola
de alguns sítios florestais de eucalipto da região do Vale do Rio
Doce (MG). R. Bras. Ci. Solo, 22:255-266, 1998.
100
REISSMANN, C.B. & WISNIEWSKI, C. Aspectos
nutricionais de plantios de Pinus. In: GONÇALVES, J.L.M. &
BENEDETTI, V (eds.). Nutrição e Fertilização Florestal.
427 p, 2000.
REISSMANN, C. B.; WISNIEWSKI, C. Nutritional aspects of
pine plantation. In: GONÇALVES, J. L. M.; BENEDETTI, V.
(Org). Forest nutrition and fertilization. Piracicaba: IPEF,
2004. p. 141-170.
ROSBERG, I.; FRANK, J. & STUANES, A.O. Effects of
liming and fertilization on tree growth and nutrient cycling in a
Scots pine ecosystem in Norway. Forest Ecology and
Management, 237:191-207, 2006.
RUBILAR, R.A.; ALLEN, H.L. & KELTING, D.L.
Comparison of biomass and nutrient content equations for
successive rotations of loblolly pine plantations on an Upper
Coastal Plain Site. Biomas and Bioenergy, 28: 548-564, 2005.
SCHUMACHER, M.V.; WITSCHORECK, R.; CALIL, F.N.;
LOPES, V.G. Biomassa e nutrientes no corte raso de um
povoamento de Pinus taeda L. de 27 anos de idade em
Cambará do Sul – RS. Ciência Florestal, Santa Maria, v. 23,
n. 2, p. 321-332, 2013.
SIMÕES, J.W.; MELLO, H.A.; MASCARENHAS
SOBRINHO, J. & COUTO, H.T.Z. A adubação acelera o
desenvolvimento inicial de plantações de Pinus caribaea var.
bahamensis. IPEF, 1:59-80, 1970.
SMETHURST, P.J. Forest fertilization: Trend in knowledge
and practice compared to agriculture. Plant Soil, v. 335, p. 83100, 2010.
101
SWITZER, G.L. & NELSON, L.E. Nutrient Accumulation and
Cycling in Loblolly Pine (Pinus taeda L.) Plantation
Ecosystems: The First Twenty Years. Soil Science Society of
America – Proceedings, 36: 143-147, 1972.
TEDESCO, M.J.; GIANELLO, C.; BISSANI, C.A.;
BOHNEN, H. & VOLKWEISS, S. Análise de solo, plantas e
outros materiais. Porto Alegre, Universidade Federal do Rio
Grande do Sul, 1995. 174p.
TURPAULT, M.-P.; UTÉRANO, C.; BOUDOT, J.-P. &
RANGER, J. Influence of mature Douglas fir roots on the solid
soil phase of the rhizosphere and its solution chemistry. Plant
and Soil, 275: 327-336, 2005.
VIERA, M. & SCHUMACHER, M.V. Concentração e
retranslocação de nutrientes em acículas de Pinus taeda L.
Ciência Florestal, 19:375-382, 2009.
VOGEL, H.L.M.; SCHUMACHER, M.V.; STORCK, L. &
WITSCHORECK, R. Crescimento inicial de Pinus taeda L.
relacionado a doses de N, P e K. Ciência florestal, 15(2):199206, 2005.
VOGEL, J.G.; JOKELA, E.J. Micronutrient limitations in two
managed Southern pine stands planted on Florida Spodosols.
Soil Sci. Soc. Am. J., v. 75, n. 3, 2011.
WALLANDER, H. Use of strontium isotopes and foliar K
content to estimate weathering of biotite induced by pine
seedling colonized by ectomycorrhizal fungi from two different
soils. Plant and Soil, 222: 215-229, 2000.
102
WHITTIG, L.D. & ALLARDICE, W.R. X-ray diffraction
techniques. In: KLUTE,A., ed. Methods of soil analysis.
Madison, American Society of Agronomy, p. 331-362. 1986.
YOUSSEF, R.A.; CHINO, M. Root-induced changes in the
rhizosphere of plants. Soil Science and Plant Nutrition, v. 35,
n. 3, p. 461-468, 1989.
ZÖTTL, H.W. Diagnosis of nutritional disturbances in forest
stands. In: FAO-IUFRO. International Symposium on Forest
Fertilization. p 75-95, 1973.
103
3 CONSIDERAÇÕES FINAIS
Os resultados obtidos demonstram que o pH tem grande
influência na distribuição das formas de potássio no solo, mesmo
em solos com diferentes composições químicas e mineralógicas.
Porém, quando esta interação envolve a muda de pinus, a
dinâmica de cada solo apresenta grande variabilidade. Mais
estudos são necessários para afirmar quais atributos estão
influenciando esta relação.
Não foi possível isolar a contribuição das formas não
trocáveis na nutrição das mudas de pinus. Os solos utilizados
apresentavam teores iniciais de potássio extraível médios a altos,
o que mascarou as possíveis contribuições das formas menos
disponíveis e os efeitos da adubação. Experimentos futuros
deveriam ser realizados em solos com teores iniciais muito
baixos, e com maior número de cultivos sucessivos, para
garantir o esgotamento das fontes prontamente disponíveis.
104
105
ANEXOS
ANEXO 1 – Difratograma das frações areia (A) e silte (B) do
Nitossolo (NT)
106
ANEXO 2 – Difratograma das frações areia (A) e silte (B) do
Latossolo (LT)
107
ANEXO 3 – Difratograma das frações areia (A) e silte (B) do
Cambissolo (CB)
108
ANEXO 4 – Difratograma das frações areia (A) e silte (B) do
Cambissolo sob cultivo de pinus (CP)
109
ANEXO 5 – Difratograma das frações areia (A) e silte (B) do
Argissolo (AG)