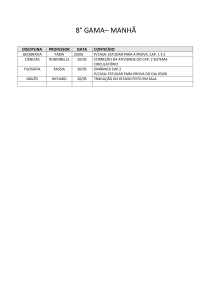
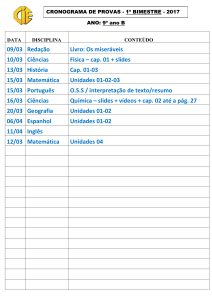
Bruna Del Vechio Koike
Avaliação comportamental e neuroquímica de ratos em
dessincronização forçada: possíveis implicações para um
modelo animal de oscilações no humor
.
Tese
apresentada
à
Universidade Federal do
Rio Grande do Norte para
obtenção do título de
Doutora em Psicobiologia
Natal
2013
1
Bruna Del Vechio Koike
Avaliação comportamental e neuroquímica de ratos em
dessincronização forçada: possíveis implicações para um
modelo animal de oscilações no humor
Tese
apresentada
à
Universidade Federal do Rio
Grande do Norte para
obtenção
do
título
de
Doutora em Psicobiologia
Orientador: John Fontenele Araujo
Co-orientador: Horacio de la Iglesia
Natal
2013
3
Catalogação da Publicação na Fonte. UFRN / Biblioteca Setorial do Centro de Biociências
Koike, Bruna Del Vechio.
Avaliação comportamental e neuroquímica de ratos em dessincronização forçada: possíveis implicações para
um modelo animal de oscilações do humor. / Bruna Del Vechio Koike. – Natal, RN, 2013.
125.: il.
Orientador: Prof. Dr. John Fontenele Araújo.
Coorientador: Prof. Dr. Horácio de La Iglesia.
Tese (Doutorado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Programa de PósGraduação em Psicobiologia.
1. Transtornos afetivos – Tese. 2. Modelo animal – Tese. 3. Dessincronização forçada – Tese. I. Araújo, John
Fontenele. II. La Iglesia, Horácio de. III. Universidade Federal do Rio Grande do Norte. IV. Título.
RN/UF/BSE-CB
CDU 616.895.4
2
Título: Avaliação comportamental e neuroquímica de ratos sob o protocolo de
dessincronização forçada: possíveis implicações para um modelo animal de
oscilações no humor
Autora: Bruna Del Vechio Koike
Data da defesa: 05 de julho de 2013
Banca Examinadora:
_________________________________________________
Prof. Dr. Horacio de la Iglesia
University of Washington
_________________________________________________
Prof. Dr. Mario Pedrazzoli Neto
Universidade de São Paulo
_________________________________________________
Profa. Dra. Tatiana Lima Ferreira
Universidade Federal do ABC
_________________________________________________
Prof. Dra. Elaine Gaviolli
Departamento de Biofísica e Farmacologia – UFRN
_________________________________________________
Prof. Dr. John Fontenele Araujo (orientador)
Departamento de Fisiologia – UFRN
4
RESUMO
Os indivíduos com Transtorno Bipolar apresentam alterações no sistema de
temporização circadiano, mostrando deslocamento de fase em diversas variáveis
fisiológicas. Muitos argumentos demonstram que alterações nos ritmos circadianos
podem ser parte da fisiopatologia do transtorno bipolar. Dada a necessidade de
maior elucidação sobre a fisiopatologia da doença, o objetivo deste trabalho foi a
validação do protocolo de dessincronização forçada como modelo animal para o
transtorno bipolar. Para isso, submetemos ratos Wistar ao protocolo de
dessincronização forçada que consiste num ciclo de claro/escuro simétrico de 22h.
Sob este protocolo, os ratos dissociam o ritmo da atividade locomotora em dois
componentes rítmicos: um sincronizado ao ciclo claro/escuro imposto de 22h; e
outro componente com período maior que 24h que segue o período endógeno do
animal. Como esses ritmos possuem períodos diferentes acabam coincidindo em
alguns momentos, o que nomeamos de CAP (do inglês coincidence active phase) e
a fase oposta, de não coincidência, chamamos de NCAP (do inglês non-concidence
active phase). A hipótese é que em CAP os animais apresentassem um
comportamento semelhante aos indivíduos bipolares em estado de mania e os
animais em NCAP, sintomas semelhantes aos indivíduos na fase depressiva.
Encontramos algumas evidências que estão descritas detalhadamente ao longo
desta tese. Em resumo dos dados comportamentais temos que os animais em
dessincronização forçada mostram-se mais estressados, com a atividade
exploratória reduzida e avaliação de risco prejudicada. Os animais no momento CAP
são menos depressivos e apresentam maior atividade locomotora concentrada na
fase escura. Enquanto os animais em NCAP apresentam maior ansiedade, mas não
apresentam anedonia. Através dos testes farmacológicos vimos que a
dessincronização foi abolida, mas não foi possível mensurar se a atividade dos
animais volta aos níveis basais. Encontramos que os animais dissociados
apresentam maior número de células marcadas para o receptor serotoninérgico
5HT1A na região da amígdala, supostamente indicando que há uma maior inibição
da amígdala nestes animais. Tomando os dados em conjunto, conseguimos validar
parcialmente o protocolo de dessincronização forçada como modelo animal para
oscilações do humor.
5
ABSTRACT
Bipolar disorder has been growing in several countries. It is a disease with
high mortality and has been responsible by the social isolation of the patients. Bipolar
patients have alterations in circadian timing system, showing a phase shift in various
physiological variables. There are several arguments demonstrating alterations in
circadian rhythms may be part of the bipolar disorder pathophysiology. Given the
necessity for further elucidation, the goal of this study was to validate the forced
desynchronization protocol as an animal model for bipolar disorder. To do this,
Wistar rats were submitted to a forced desynchronization protocol which consists in a
symmetrical light dark cycle with 22h. Under this protocol, rats dissociate the
locomotor activity rhythm into two components: one synchronized to the light / dark
cycle with 22h, and another component with period longer than 24 hours following
the animal endogenous period. These rhythms with different periods sometimes
there is coincidence, which we named CAP (Coincidence Active Phase) and the
opposite phase, non-coincidence, called NCAP (Non-Concidence Active Phase). The
hypothesis is that in CAP animals present a mania-like behavior and animals in
NCAP depressive-like behavior. We found some evidence described in detail
throughout this thesis. In sum, the animals under forced desynchronization protocol
were more stressed, showed an increase in stereotypic behaviors such as grooming
and reduction in other behaviors such as risk assessment and vertical exploration
when compared to the control group. The CAP animals showed increased locomotor
activity, especially during the dark phase when compared to controls (rats under T24)
and less depressive behavior in the forced swim test. The animals in NCAP showed
a higher anxiety in elevated plus maze, but they don’t have ahnedonia. The animals
under dissociation have more labeled 5HT1A cells at the amygdala area, which
appoint that they have more amygdala inhibition. Taking these data together, we
could partially validated the forced desynchronization protocol as an animal model for
mood oscillations.
6
SUMÁRIO
1.INTRODUÇÃO ...................................................................................................
1.1.Transtorno Bipolar do Humor...........................................................................
1.2. Alterações dos ritmos biológicos no transtorno bipolar ..................................
1.3. Sincronização do organismo com o meio externo .........................................
1.4. Serotonina e neurotransmissão .....................................................................
1.5. Lítio ................................................................................................................
1.6. Modelos experimentais ..................................................................................
1.5.1. Dessincronização forçada ...........................................................................
2.OBJETIVO GERAL ............................................................................................
2.1. Objetivos específicos .....................................................................................
3. MATERIAIS E MÉTODOS ................................................................................
3.1. Para a validação aparente .............................................................................
3.1.1. Animais .......................................................................................................
3.1.2.Testes comportamentais ..............................................................................
3.1.2.1. Labirinto em cruz elevado ........................................................................
3.1.2.2. Campo aberto ..........................................................................................
3.1.2.3. Teste do nado forçado .............................................................................
3.1.2.4. Preferência claro-escuro .........................................................................
3.1.3. Bateria de testes .........................................................................................
3.1.4. Atividade locomotora....................................................................................
3.1.6. Teste de consumo de glicose .....................................................................
3.2. Teste farmacológico para Validação Preditiva...............................................
3.3. Testes Neuroquímicos para a Validação do Contruto ...................................
3.3.1. Hibridização in situ do receptor de serotonina 5HT2A ...............................
3.3.2. Imunohistoquímica para o receptor 5HT1A ................................................
3.4. Análise estatística ..........................................................................................
4. RESULTADOS...................................................................................................
4.1. Resultados dos testes comportamentais para a validade aparente .............
4.1.1. Atividade locomotora ...................................................................................
4.1.2. Testes comportamentais .............................................................................
4.1.2.1. Bateria de testes .......................................................................................
4.1.2.2. Campo aberto ...........................................................................................
4.1.2.3. Labirinto em cruz elevado ........................................................................
4.1.2.4. Nado forçado ............................................................................................
4.1.3. Teste de consumo de glicose .....................................................................
4.2. Resultados para a Validade Preditiva ............................................................
4.2.1. Teste farmacológico – lítio ..........................................................................
4.3. Testes Neuroquímicos para a Validação do Construto ..................................
4.3.1. Hibridização in situ ......................................................................................
4.3.2. Imunihistoquímica .......................................................................................
5. DISCUSSÃO .....................................................................................................
6. CONCLUSÕES .................................................................................................
7. REFERÊNCIAS BIBLIOGRÁFICAS .................................................................
ANEXOS.................................................................................................................
11
13
16
17
19
22
23
24
30
30
31
32
32
32
33
34
35
36
37
38
39
39
41
41
42
43
44
44
44
46
46
48
51
52
53
54
54
57
57
60
62
80
81
93
7
AGRADECIMENTOS
Agradeço primeiramente à minha mãe, que sempre me apoiou em tudo o que eu quis
fazer, sempre acreditou em mim e me incentivou a seguir os meus objetivos. Sempre foi
meu teto e meu chão, meu tudo! Te amo mais que o mundo!
Em segundo lugar, agradeço ao meu querido orientador John. O John me abriu
muitas portas. Em seu laboratório, pude aprender a mexer com animais desde a
manipulação, bioterismo e a parte dos testes comportamentais; aprendi o quanto é
importante fazer colaborações e principalmente aprendi a socializar! Aprendi como curtir ser
pesquisador, como curtir os orientandos e também a não me intrometer em brigas de
cachorro grande. Aprendi também o que é uma “vidinha marromeno” e estou colocando em
prática os seus ensinamentos! Ganhei um grande e querido amigo.
Agradeço à Mariana Dias Leite, que foi meu braço direito em grande parte deste
trabalho. É uma excelente aluna, muito dedicada e responsável, além de ser uma grande
amiga! Obrigada Mari, você sabe o quanto foi importante nesse trabalho.
Agradeço muito a todos os meus companheiros do laboratório que, com toda
certeza, atrapalhei muito nestes anos ao falar o tempo todo no laboratório, os conselhos
tortos e até um pouco de “revolta/anarquia”: Miguelzinho, Kathiane, Breno, Bruno, Felipe,
Cleanto, Sionaldo, Valciclênio, Rafael, Rafael Pink, Marconi (que sempre me salvou),
Crhistiane, Nathalinha, Everly, Gabi, Manu, Julio. Agradeço também os companheiros de
outros laboratórios que sempre tiveram a maior prontidão para me ajudar como a Andrea de
Sá, o Wall Hatori, Ronaldo, André Pontes, Rovena, Alícia, Aline Belísio, Professora Miriam,
Professor Rodrigo do Ice, Professora Vanessa e Professora Elaine da Farmaco.
Duas que se tornaram minhas irmãs e confidentes por aqui: Carol e Laís, vocês
moram
no
meu
coração!
Acolheram-me
numa
fase
bem
difícil
de
readaptação/frustração/desmotivação que não foi fácil de passar. Aqui também incluo Kathi,
que ouviu muitos desabafos e me deu sábios conselhos. Sempre pude contar com essas
amigas. Obrigada meninas =)
Agradeço todos os professores e funcionários do departamento que sempre foram
solícitos e atenciosos no trato e ajuda.
Agradeço em especial às pessoas “out of lab” que fazem parte da minha vida, que
fazem parte de mim e me fazem lembrar todos os dias o quanto sou humana e o quanto
essa vida é boa! Aqui estão inclusos Clarisbela, Josy, Jana, Luisa prima, Bia e em especial
o meu amor, Igor, que foi toda a parte boa dessa vida out of lab nesses últimos tempos <3.
8
Lista de Figuras
Figura
Descrição
Página
Figura 1
Actogramas sob diferentes regimes de luz
24
Figura 2
Actograma e periodograma de um animal dessincronizado
26
Figura 3
Expressão alternada das subdivisões dos núcleos supraquiasmáticos
27
Figura 4
Ilustração do Labirinto em cruz elevado
32
Figura 5
Ilustração do campo aberto
33
Figura 6
Ilustração do aparato do nado forçado
34
Figura 7
Ilustração da caixa de preferência claro/escuro
35
Figura 8
Foto das condições do teste de consumo de glicose
38
Figura 9
Perfil da atividade locomotora dos diferentes grupos
44
Figura 10
Histogramas da atividade locomotora separados por grupo e por
condição de luz
44
Figura 11
Distância percorrida no CA durante a bateria de testes
46
Figura 12
Resultados do deslocamento no CA dos animais dissociados
48
Figura 13
Resultados da análise do tempo despendido no centro do CA
48
Figura 14
Grooming durante o CA
49
Figura 15
Rearing durante o CA
49
Figura 16
Head down durante o LCE
50
Figura 17
Latência para o comportamento de desespero no NF
51
Figura 18
Resultados do teste de consumo de glicose
52
Figura 19
Porcentagem do consumo de glicose
52
Figura 20
Actogramas e periodogramas representativos dos animais antes e
depois do tratamento com o lítio
53
Figura 21
Atividade locomotora dos animais antes e durante o tratamento com
lítio
54
Figura 22
Imagens da hibridização in situ
56
Figura 23
Expressão de RNAm do receptor 5HT2A
56
Figura 24
Foto representativa das células marcadas para o receptor 5HT1A.
58
9
Lista de Tabelas
Tabela
Descrição
Página
Tabela 1
Grupos experimentais da bateria de testes
36
Tabela 2
Resultados do LCE
50
Tabela 3
Atividade dos animais antes e durante o tratamento com lítio
54
Tabela 4
Resumo dos achados comportamentais dos animais dissociados
em comparação ao grupo controle.
58
Tabela 5
Resultados da hibridização in situ para o receptor 5HT2A no
grupo controle
59
Tabela 6
NODs da hibridização in situ para o receptor 5HT2A nos animais
dissociados
60
Tabela 7
Contagem de células 5HT1A positivas
61
10
Lista de abreviações
ACTH
Hormônio adrenocorticotrófico
CA
Campo Aberto
CAP
Coincidence Activity Phase
CE
Ciclo claro/escuro
CPF
Córtex pré-frontal
Dm
Dorsomedial
EEG
Eletroencefalograma
GSK3
Glycogen Synthase Kinase
HPA
Hipotálamo – Hipófise - Adrenal
LCE
Labirinto em Cruz Elevado
NCAP
Non-Coincidence Activity Phase
NF
Nado Forçado
NOD
Normalized Optical Density
NSQs
Núcleos supraquiasmáticos
PCE
Caixa de Preferência ao Claro ou Escuro
REM
Rapid Eye Movement
T
Período circadiano
TB
Transtorno Bipolar
vl
Ventrolateral
11
1. INTRODUÇÃO
A agitada vida moderna, com mais recursos à disposição e mais
responsabilidades a cumprir, tornam o dia de 24h curto para a realização de todas
as tarefas. A duração do sono vem sendo gradativamente reduzida durante os dias
da semana ao longo dos anos (Vetter e cols, 2012), aumentando a privação de sono
entre os indivíduos, e isso certamente ocorre devido às facilidades tecnológicas e ao
grande número de informações cotidianas no século XXI (Louzada e Menna-Barreto,
2003) aliadas a maior quantidade de compromissos sociais como o horário de
trabalho, de estudos, cursos, prática de exercícios físicos ou mesmo o tempo
dispensado para ficar com a família ou com os amigos. Essa privação de sono
crônica tem consequências nos ritmos biológicos dos indivíduos, tema que
abordaremos ao longo desta tese.
Cada vez mais a exigência em torno do desempenho pessoal torna-se
exaustiva. Como mecanismos de fuga, os indivíduos podem desenvolver diferentes
patologias, causando alterações no humor, o que pode se traduzir no jeito de
pensar, sentir e em outros comportamentos do indivíduo.
Em grandes cidades, como a grande São Paulo, cerca de 30% dos habitantes
apresentam transtornos mentais. Entre estes, a prevalência dos transtornos de
ansiedade aparece em primeiro lugar com cerca de 20%, seguido dos transtornos de
humor que afetam 11% dos indivíduos (Andrade e cols., 2012).
Os transtornos do humor afastam os indivíduos do trabalho, alterando uma
das funções humanas fundamentais para o bem estar social do indivíduo, pois o
trabalho é uma reafirmação de nossa identidade e status social, uma vez que
proporciona satisfação pessoal e estabelece nossa rotina social diária. Além disso, o
trabalho está ligado à demanda financeira individual e reflete o status funcional do
indivíduo. O grau em que a doença interfere com a atividade do mercado laboral e a
produtividade individual constitui um componente importante da carga da doença
(Frank e Koss, 2005).
12
Os transtornos do humor podem ser classificados como unipolares ou
bipolares e afetam atualmente cerca de 340 milhões de pessoas em todo o mundo
(ABRATA, 2009). A diferença básica entre eles é que transtornos unipolares
apresentam predominantemente sintomas depressivos e os bipolares apresentam
sintomas maníacos intercalados com os depressivos (Mitchell e cols., 1992).
O transtorno unipolar, também denominado depressão, é a principal doença
com efeitos sociais e na saúde. Impossibilita os indivíduos de prosseguirem
naturalmente suas atividades laborais e pessoais (WHO, 2004).
O transtorno bipolar (TB) corresponde a um dos mais prevalentes e
potencialmente graves transtornos psiquiátricos. Estima-se que a prevalência
mundial seja de aproximadamente 29,5 milhões de pessoas, cerca de 513 pessoas
afetadas a cada 100.000. É considerada uma das doenças de maior mortalidade e
morbidade (WHO, 2004).
Indivíduos acometidos têm maiores taxas de desemprego e estão mais
sujeitos a utilizarem serviços médicos e serem hospitalizados (Lima e cols., 2003).
Por este motivo, os custos associados ao TB são substanciais. Os pacientes utilizam
os serviços de saúde incluindo hospitalização e medicação, além de apresentarem
comorbidade em 70% dos casos tanto com outras doenças mentais – como
transtornos de ansiedade ou uso de substâncias de abuso – quanto com doenças
físicas – como absentismo, desemprego, comportamento sexual de risco e, em
casos extremos, o suicídio (Merikangas e cols., 2007). Somando-se os custos
diretos e indiretos em consequência desta patologia foi estimado um valor de
US$252.212,00 por pessoa (durante a vida) nos EUA, gerando um custo total para o
país de aproximadamente 24 bilhões de dólares, sendo 44% somente para custos
indiretos (Miret e cols., 2013). Na Inglaterra, uma estimativa recente do custo com
pacientes bipolares foi de 1,6 bilhões de libras. Com a inclusão dos custos
empregatícios, o total aumenta para 5,2 bilhões de libras! (McCrone e cols., 2010).
13
1.1. Transtorno Bipolar do Humor
O transtorno bipolar ocupa o quinto lugar entre as principais causas de
incapacidade em todo o mundo em indivíduos jovens, tanto por morte prematura
quanto por inabilidade, e afeta cerca de 1-6% da população brasileira (ABRATA,
2009). Um indivíduo com transtorno bipolar do humor pode apresentar grandes
oscilações no seu estado de humor o que é impactante na vida do paciente, de sua
família e da sociedade, causando muitas vezes prejuízos irreparáveis em vários
setores da vida do indivíduo, interferindo no trabalho, nas relações afetivas e
familiares. O TB é uma doença que se caracteriza pela alternância de humor: ora
ocorrem episódios de mania (euforia), ora de depressão, com períodos intercalados
de normalidade (Moreno e Moreno, 2005).
A fase de mania é caracterizada por euforia exacerbada, aumento do discurso
e da velocidade da fala (tagarelice), pensamentos desconexos ou muito rápidos,
atividade mental e física acentuada, aumento da energia, redução da necessidade
de sono, irritabilidade, acuidade perceptiva, paranóia, redução do senso crítico,
ideias de grandeza, aumento do interesse sexual e impulsividade. Pacientes nessa
fase apresentam humor efervescente e exaltado, mas com irritabilidade subjacente
(Goodwin e Jamison, 2007).
Por sua vez, a fase de depressão possui características praticamente opostas
à fase de mania, com o abrandamento e redução da maioria dos aspectos
emocionais e comportamentais, como a fala, energia, sexualidade e a habilidade de
experimentar prazeres (anedonia). Entretanto, a severidade dos sintomas pode
variar bastante desde a simples redução da atividade física e mental, com leve
distorção na cognição e na percepção, até estados profundos de depressão,
desilusão, pensamentos suicidas, alucinação e obscurecimento da consciência. O
humor na maioria dos pacientes nessa fase mostra-se pessimista, desolador e
desesperado (Goodwin e Jamison, 2007).
14
1.2. Alterações dos Ritmos Biológicos no Transtorno Bipolar
Muitos pesquisadores sugerem que os episódios de mania e/ou depressão
emergem como consequências da ruptura dos ritmos circadianos nos indivíduos
suscetíveis. Frequentemente são observadas nos pacientes alterações de
comportamento relacionadas aos ritmos biológicos, especialmente a partir de
eventos que perturbam os zeitgebers sociais, ou seja, fatos que marcam época,
como a morte dos pais, separação, demissão ou início de trabalho em período
integral (Ehlers e cols., 1988; Malkoff-Schwartz e cols., 2000; Grandin e cols., 2006).
Geralmente os indivíduos apresentam os sintomas dos transtornos de humor
mais acentuados durante as primeiras horas do dia, o que vai claramente
melhorando ao entardecer, essa flutuação na qualidade do humor faz parte dos
sintomas encontrados nos indivíduos depressivos. Além disso, essas flutuações são
acompanhadas em paralelo por alterações na psicomotricidade, na capacidade de
atenção e na ansiedade que também apresentam a mesma flutuação e ocorre uma
melhora ao longo do dia (Poterfield e cols., 1997; Rusting e Larsen, 1998).
Para a maioria dos pacientes ocorrem mudanças regulares e repetitivas na
intensidade dos sintomas depressivos ao longo do dia, geralmente apresentando
uma piora pela manhã (Wirz-Justice, 2006). Em um estudo realizado na Nova
Zelândia, cerca de 30% dos pacientes de uma instituição apresenta padrões opostos
na flutuação do humor. Uma observação relatada para estes pacientes foi a fraca
resposta aos antidepressivos serotoninérgicos, altas taxas de triptofano comparados
aos indivíduos saudáveis e inclusive uma evidência genética: maior frequência do
alelo curto do polimorfismo de repetição em tandem da região promotora do
transportador de serotonina. Com isso, os índices da razão triptofano/metabólitos de
serotonina aumentam, apontando alterações no sistema sertoninérgico e sugerindo
sua influência na variação diurna do humor (Joyce e cols., 2005).
Além da flutuação do humor, alterações em diversos parâmetros rítmicos têm
sido observadas nos pacientes com transtorno de humor. A desregulação de apenas
um dos ritmos como o ciclo sono/vigília pode induzir episódios de mania ou de
15
depressão e, corroborando com este dado, a remissão das crises geralmente
acompanham a regularização das anormalidades circadianas. A manipulação do
ciclo sono/vigília tem sido usada com eficácia no tratamento da depressão há mais
de 30 anos (Pflug e Tolle, 1971). Uma única noite de privação de sono pode
restaurar a eutimia em aproximadamente 60% dos pacientes depressivos (WirzJustice e van der Hoofdakker, 1999).
Mais de 80% dos pacientes depressivos têm queixas da má qualidade do
sono, sejam elas na dificuldade de iniciar o sono, mantê-lo ou mesmo o despertar
precoce (Maurizi, 2000). Alterações na arquitetura do sono também são observadas
no TB, como a redução do sono de ondas lentas, redução da duração do sono REM,
aumento de sua densidade e o encurtamento de sua latência (Reynolds e Kupler,
1987; Stefos e cols., 1998; Thase, 2000). Essas alterações no sono REM podem ser
interpretadas como resultado do avanço de fase do sistema de temporização
circadiano e gera uma relação de fase anormal entre vários outros componentes
rítmicos (Gruber e cols., 2011; Wehr e cols., 1979). Em outras palavras, sabemos
que deve haver sintonia entre os ritmos biológicos que devem ter um ângulo de fase
constante, estável, ou seja, as fases dos ritmos devem apresentar uma distância
temporal constante. Como um exemplo, o crepúsculo reduz a incidência de luz sobre
os olhos e somente depois que a fase de escuro se inicia é que ocorre a fase de
início da secreção de melatonina, há uma relação entre essas fases. Quando
dizemos que houve um avanço de fase, significa que uma das fases se adiantou em
relação à outra. Quanto mais estável essa relação de fase, melhor os organismos
podem se adaptar, podendo antecipar atividades ou recursos antes mesmo que eles
sejam necessário, poupando energia.
Algumas variáveis metabólicas rítmicas apresentam mudança na relação de
fase após o início da patologia. No caso do o eixo HPA (hipotálamo – hipófise –
adrenal) se encontra hiperativo na depressão, pois a hipercortisolemia está
fortemente associada à depressão e, em especial, aos indivíduos que apresentam
os subtipos melancólicos e psicóticos. Na hipercortisolemia, a secreção do cortisol é
irregular e perde a relação de fase com o ritmo de secreção do ACTH (hormônio
16
adrenocorticotrófico), é observado um desacoplamento entre a temporalidade de
secreção desses hormônios. Essa hipercortisolemia pode ser devido à hipertrofia do
córtex adrenal ou à secreção anormal de ACTH, ou à alta sensibilidade do córtex
adrenal na resposta ao ACTH. No geral, há um aumento na secreção de ACTH nos
pacientes independentemente do status de hipercortisolemia, onde sua meia-vida é
reduzida (Carroll e cols., 2007). Os ritmos do ACTH e do cortisol tendem a ser mais
lentos na depressão e com a acrofase atrasada nos adultos (Souêtre e cols., 1989).
Outras variáveis fisiológicas que se apresentam em fase avançada nos
indivíduos depressivos são a temperatura corporal, secreção de cortisol e de
melatonina, alguns neurotransmissores (como a norepinefrina e seu metabólito 4hidroxifenilglicol), bem como a alteração de amplitude das curvas secretórias.
Entretanto, o ritmo de outras variáveis permanece com a relação de fase estável
como a secreção de prolactina e hormônio do crescimento (Koenigsberg e cols.,
2004; Monteleone e Maj, 2008).
Mesmo com o avanço de fase em grande parte dos ritmos, indivíduos
bipolares eutímicos podem apresentar atraso de fase na curva de secreção de
melatonina (Nunrberger e cols., 2000), talvez seja por este motivo que os indivíduos
bipolares apresentam uma preferência diurna mais vespertina (Wood e cols., 2009).
A pesquisa sobre alterações nos ritmos circadianos no transtorno de humor
surgiu nos anos 60, com as primeiras evidências de alterações de ritmo na secreção
de noradrenalina e metabólitos coletados em fluidos corporais de pacientes
depressivos (Schildkraut, 1965). Outra evidência bastante marcante da relação entre
os transtornos de humor e os ritmos biológicos é que a ação das drogas
antidepressivas e estabilizadoras do humor afeta a ritmicidade do relógio biológico.
Além de causar melhoria nos sintomas, restauram o ritmo circadiano em humanos e
animais (Monteleone e Maj, 2008). O que ainda não se sabe é se as alterações
observadas nos ritmos biológicos desses pacientes são resultantes do rompimento
dos ritmos sociais (zeitgebers externos) ou se têm origem no componente interno
dos indivíduos, como por exemplo, vulnerabilidade cognitiva, características da
personalidade e anormalidades no marcapasso circadiano (Grandin e cols., 2006).
17
Diversos estudos genéticos tem sugerido que determinados genes relógio –
genes que fazem parte da circuitaria molecular do marcapasso circadiano – são
ajustados conforme a manifestação dos transtornos do humor. Temos alguns
exemplos na literatura, como o polimorfismo T3111C do gene Clock cujo alelo C em
homozigose está relacionado com a alta recorrência de episódios nos bipolares
(Benedetti e cols., 2003); ao mesmo tempo em que camundongos mutantes para o
gene Clock estão sendo considerados animais modelo para mania (Roybal e cols.,
2007). Outros genes relógio como Bmal1, TIM e PER3 também foram associados
com o transtorno bipolar (Kato, 2007).
Uma grande dúvida que permanece é como o rompimento da ritmicidade
circadiana está associado aos transtornos de humor. Sabe-se que os pacientes
bipolares apresentam maior desorganização dos ritmos circadianos mesmo quando
eutímicos, com ou sem relação com distúrbios de sono, assim demonstrando que
esses pacientes têm ciclo circadiano diferente independentemente de distúrbios
puros de sono (Jones e cols., 2005). Essa desorganização no sistema de
temporização circadiano não está associada com o atual estado de humor, por outro
lado, a terapia de regularização dos ritmos sociais faz melhorar o humor dos
pacientes bipolares (Frank e cols., 2000).
1.3. Sincronização do organismo com o meio externo
Para o sistema de temporização dos mamíferos, uma pista ambiental crucial
para a sincronização com o ambiente é o ciclo claro/escuro (CE) que é capaz de
sincronizar praticamente todos seus ritmos circadianos (Marques e Menna-Barreto,
2005). Sendo a retina a única estrutura sensível ao sinal luminoso em mamíferos, a
informação do ciclo CE ambiental é captada pelas células ganglionares da retina.
Essas informações sobre o nível de luminosidade do ambiente são transmitidas
através do trato retino-hipotalâmico diretamente aos núcleos supraquiasmáticos do
hipotálamo (NSQs). Considerados osciladores centrais, os NSQs são as principais
estruturas envolvidas na sincronização dos ritmos biológicos dos mamíferos com o
18
ciclo claro/escuro (Schibler e cols., 2005). Atuam como osciladores centrais do
organismo, que por sua vez sincronizam os osciladores periféricos. Portanto,
alterações
na
ritmicidade
circadiana
podem
ser
consequências
do
mau
funcionamento (ou não sincronização) dos NSQs, gerando o posicionamento
anormal das fases em vários dos ritmos biológicos (Pandi-Perumal e cols. 2009).
A dessincronização dos núcleos supraquiasmáticos pode explicar muitos dos
aspectos clínicos do transtorno de humor recorrentes (incluindo sazonalidade e tipo
de
episódio),
por
meio
de
efeitos
secundários
sobre
sistemas
de
neurotransmissores. A condição de ritmo em livre-curso e ritmos não sincronizados
ao ciclo claro/escuro de 24h pode dessincronizar os outros ritmos circadianos,
provocando uma desordem temporal interna e afetando de forma adversa o humor
(Wehr e cols., 1983).
Os NSQs apresentam diversas projeções diretas e indiretas. Uma das
projeções indiretas dos NSQs acaba por estimular a glândula pineal que produz um
importante hormônio do sistema de temporização circadiano que é a melatonina. A
melatonina é um hormônio sinalizador da noite para o organismo, uma vez que é
produzida somente nesta fase. Dessa forma, regula as atividades rítmicas
circadianas nos mamíferos. A redução da amplitude de sua curva de secreção, bem
como o avanço ou atraso em sua fase, pode ser parcialmente responsável pela má
qualidade do sono observada nos pacientes com TB (Pandi-Perumal e cols., 2009).
A melatonina é o neurormônio com maior influência na atividade elétrica nos
neurônios do NSQ, pois é capaz de inibir o padrão de disparos desses neurônios,
provavelmente através da ativação de mecanismos GABAérgicos (Golombek e cols.,
1996) e da ativação de receptores MT1 e MT2 presentes em larga escala nos NSQs
(Dubocovich e Markowska, 2005).
Além da melatonina, a serotonina também tem influência nas alterações de
sono. O sistema serotoninérgico inibe neurônios colinérgicos da ponte, diminuindo a
latência do sono REM e modulando os neurônios do tálamo e do córtex pré-frontal,
induzindo a fase de sono de ondas lentas (Thase, 2000). Além do seu papel no
sono, há uma densa projeção serotoninérgica dos núcleos da rafe que inerva os
19
NSQs, bem como um grande número de receptores serotoninérgicos na região
ventrolateral dos NSQs. A estimulação elétrica da rafe aumenta a liberação de
serotonina no NSQ e provoca avanços e atrasos de fase, sugerindo que a
serotonina deve ter algum papel na sincronização não fótica, além de modular a
atividade no trato retino hipotalâmico já que agonistas serotoninérgicos podem
atenuar ou aumentar a resposta a estímulos fóticos (Prosser, 2003).
Amplamente distribuída pelo sistema nervoso central, a serotonina parece
estar associada a muitos comportamentos como a inibição de comportamentos
agressivos, a regulação de ritmos circadianos e do humor, ao sono, à atividade
sexual, ao apetite, às funções neuroendócrinas, a regulação da temperatura
corporal, sensibilidade à dor, atividade motora, funções cognitivas e à facilitação de
comportamentos motores apetitivos (fome e sexo) (Thase, 2000; Ballone, 2005).
1.4. Serotonina e neurotransmissão
No transtorno bipolar, a serotonina parece ter um papel fundamental na
fisiopatologia da doença, uma vez que os níveis deste neurotransmissor, bem como
seus metabólitos (5-HIAA ou ácido 5-hidroxindolacético), aparecem alterados no
cérebro (estudos pós-morte) e no líquor dos pacientes com TB, principalmente na
fase depressiva, onde a concentração está reduzida. Na mania, os resultados são
menos consistentes, pois há divergência entre os estudos (para revisão, ver
Mahmood e Silverstone, 2001). Como a serotonina é liberada difusamente em quase
todo o encéfalo, o conhecimento dos tipos de receptores e sua exata localização são
fundamentais para o entendimento da neurotransmissão serotoninérgica. Como
exemplo, podemos citar o receptor 5HT1A que, frente a uma grande quantidade de
serotonina na fenda sináptica, pode ter diferentes consequências: quando localizado
na pré-sinapse de neurônios serotoninérgicos ocorre redução na liberação de
serotonina como um sinal de feedback negativo; quando na pós-sinapse de
neurônios corticais ocorre supra regulação (up regulation) dos receptores
20
serotoninérgicos (Akimova e cols., 2009), além de promover ação inibitória visto que
este receptor é ligado à proteína Gi.
Estudos que avaliaram a influência do sistema serotoninérgico na função
neuroendócrina,
principalmente
em
relação
ao
hormônio
do
crescimento,
observaram uma tendência a menor atividade pré-sináptica e maior atividade póssinápica serotoninérgica nos pacientes em mania, sugerindo uma hipoatividade
serotoninérgica no hipotálamo (Newman e cols., 1998; Shiah e Yatham, 2000). A
hipoatividade serotoninérgica foi relatada por Mahmood e Silverstone (2001)
também para pacientes com depressão bipolar e eutímicos. Além disso, os
medicamentos estabilizadores de humor possuem a capacidade de aumentar a
atividade serotoninérgica na mania (Shiah e Yatham, 2000).
Outras evidências são os tratamentos farmacológicos da fase depressiva que,
geralmente, envolvem drogas que aumentam significativamente a quantidade de
serotonina na fenda sináptica (os inibidores seletivos de recaptação de serotonina),
bem como os antipsicóticos atípicos, que possuem a capacidade de inibir a
recaptação de serotonina e de bloquear receptores 5HT2 A e C, enquanto ativam
receptor 5HT1A (Goodwin e Jamison, 2007). Esse fato corrobora com a teoria
monoaminérgica da depressão, que foi a primeira teoria de importância sobre a
etiologia da doença, a qual postulou que esta era devida à deficiência de
neurotransmissores
monoaminérgicos,
principalmente
noradrenalina
(NA)
e
serotonina (5-HT). As evidências baseiam-se na ação das drogas que depletam
esses neurotransmissores e podem induzir depressão, além da sugestão de supra
regulação (up regulation) anormal dos receptores serotoninérgicos pós-sinápticos.
Por outro lado, os antidepressivos com ações farmacológicas que potencializam
esses neurotransmissores, aumentam sua disponibilidade na fenda sináptica, seja
por infra regulação (down regulation) nos receptores pós-sinápticos ou pelo bloqueio
das bombas de recaptação, ou até pela inibição da enzima degradadora de
monoaminas (MAO – monoamina oxidase) e, como consequência, o tratamento
resulta na melhoria geral dos sintomas (Stahl, 2006). Porém, essa teoria sofreu
diversas críticas, pois não explicava vários fatos, como a ausência de resposta aos
21
antidepressivos em alguns pacientes ou a ação de drogas que também aumentam a
atividade monoaminérgica (como anfetamina e cocaína) não serem clinicamente
efetivas, além da demora na resposta terapêutica com a administração dos
antidepressivos, já que a mudança nos níveis de monoaminas na sinapse ocorre em
algumas horas e a resposta terapêutica requer a administração contínua por
semanas (Baldessarini, 1989).
Uma outra hipótese relevante à ciclicidade da doença é o paradigma de
kindling. Postulada por Post e cols. (1992), essa teoria desenvolve-se sobre o
achado
fisiológico
de
que
estímulos
elétricos
ou
químicos
intermitentes,
subliminares, produzem despolarização cada vez mais forte no sistema límbico, a
qual pode levar a um foco convulsivo permanente independente. Embora uma
ligação direta entre fenômeno de kindling e a recorrência clínica não possa ser
estabelecida com facilidade, a hipótese possui a vantagem de explicar inúmeros
achados clínicos em uma teoria como: os episódios iniciais de TB tendem a ser
precipitados por estressores ambientais, enquanto episódios posteriores tendem a
ser desencadeados, com menos frequência, por fatores psicossociais; a gravidade
dos episódios não tratados tende a piorar com o tempo; o intervalo entre os
episódios de alteração do humor diminuem com o tempo; e eventos estressantes na
infância podem predispor o indivíduo a transtornos do humor na idade adulta
(Kendler e cols., 2000).
Para o transtorno bipolar, o tratamento com lítio é o preferido e o mais
frequente entre os pacientes bipolares. O lítio não exerce ação direta sobre a
neurotransmissão serotoninérgica, nem aumenta sua concentração, mas é capaz de
aumentar a captação de serotonina (Carli e cols., 1997). Quando o tratamento é
interrompido, os sintomas da doença retornam rapidamente, podendo sugerir o
importante papel da alteração na captação de serotonina na fisiopatologia e sua
normalização no tratamento do transtorno bipolar (Carli e cols., 1997).
22
1.5. Lítio
O lítio sabidamente age na GSK3 (Glycogen Synthase Kinase-3), que é uma
proteína quinase altamente conservada e possui duas formas de apresentação
GSK3α e GSK3β. Além de inibir diretamente a ação da GSK3, o lítio aumenta a
fosforilação inibitória da serina da GSK3 (De Sarno e cols, 2002), assim como outros
neuromoduladores como antidepressivos, antipsicóticos e estabilizadores de humor
que também aumentam fosforilação da GSK3 e a inibem. A GSK3 encontra-se
reduzida tanto durante a hiperatividade quanto no estado semelhante à depressão
em camundongos, bem como em indivíduos bipolares sintomáticos. Ou seja, em
roedores a fosforilação insuficiente da serina da GSK3 pode desempenhar um papel
crítico em distúrbios comportamentais, como ocorre no transtorno bipolar (Polter e
cols, 2010).
A GSK3 no sistema nervoso central é regulada pela serotonina, via receptores
5HT1 e 5HT2, e age como um modulador intermediário no sistema da
neurotransmissão serotoninérgica. A presença da serotonina no cérebro resulta na
rápida fosforilação da serina N-terminal da GSK3 e a inativação da sua atividade
constitutiva (Polter e Li, 2011).
Em somatória, podemos acrescentar que o período endógeno de humanos
(Johnsson e cols., 1983) e ratos (Kripke e Wyborney, 1980) mostram-se alongados
quando estão em tratamento com o lítio, o que corrobora com os achados
encontrados no ritmo de seus neurotransmissores que apresentam atraso de fase na
secreção (Kafka e cols., 1982; McEachron e cols., 1982). Alterações rítmicas
também são observadas quanto à fase, amplitude e a responsividade à luz (para
revisão ver Klemfuss, 1992). Essas alterações podem ser devido a ação da GSK3
na regulação e/ou estabilização das proteínas do relógio molecular circadiano como
PER2, CRY2, CLOCK, REV-ERBα e BMAL1 (Harada e cols, 2005; Yin e cols, 2006;
Sahar e cols, 2010; Spengler e cols, 2009).
23
1.6. Modelos Experimentais
A neurociência se depara com uma grande dificuldade em estudar o cérebro
humano in vivo. Mesmo com o expoente avanço das técnicas não invasivas, ainda
restam inúmeras questões em aberto quanto à fisiologia e à biologia molecular do
cérebro humano. Um meio útil pelo qual a ciência vem se desenvolvendo é através
de modelos animais.
Atualmente há muitos modelos e protocolos que tentam mimetizar os
transtornos mentais em animais de experimentação. Entretanto, apresentam certas
limitações. A começar pela forma através da qual é feita o diagnóstico. O diagnóstico
é fenomenológico, ou seja, baseado em sinais, sintomas e no curso da doença
descritos no manual DSM-V (Diagnostic and Statistical Manual of Mental Disorders –
fifth edition (APA, 2013)). Muitos dos sintomas utilizados para o diagnóstico da
doença em humanos não são capazes de serem avaliados em modelos animais
(como culpa, tristeza, alucinações, ilusões), enquanto outros podem ser somente
inferidos.
Apesar da existência de controvérsia entre os dados, alguns modelos foram
bem sucedidos em descobrir um pouco mais sobre a doença, como os modelos de
depressão (estresse moderado crônico, modelo de desespero aprendido, modelos
de separação parental) e os modelos que tentam mimetizar o estado de mania
(hiperatividade induzida por psicoestimulantes, privação de sono, alteração do ritmo
circadiano, kindling). Entretanto, esses modelos apresentam apenas um dos estados
de humor. A característica de alternar períodos de depressão, mania e eutimia de
forma recorrente e cíclica, torna bastante difícil o desenvolvimento de um modelo
ideal (Gavioli e cols., 2009).
Para a validação de um modelo animal usado para estudar patologias
humanas é necessário que sejam cumpridos ao menos três critérios: validade
aparente, validade do construto e validade preditiva. A validade aparente, ou de
face, baseia-se nas semelhanças entre os sinais e sintomas observados em
humanos que sofrem da patologia em questão com o fenótipo apresentado pelos
24
animais submetidos ao modelo sugerido. A validade do construto é baseada na
etiologia, nas bases biológicas e fatores desencadeadores da doença. Por fim, a
validade preditiva, ou farmacológica, é contemplada quando os fármacos utilizados
em humanos são capazes de reverter os sinais e sintomas da doença observados
nos animais experimentais (Gavioli e cols., 2009). No presente trabalho
pretendemos contemplar os três critérios de validação.
1.6.1. Dessincronização forçada
O período circadiano (T) tanto dos humanos (Czeisler e cols.,1999), quanto
dos ratos é em média pouco maior de 24h e todos os dias seus ritmos biológicos são
arrastados através de pistas ambientais (zeitgebers). Nos mamíferos, a principal
pista ambiental é o ciclo CE.
O sistema responsável pela geração da ritmicidade circadiana (sistema de
temporização circadiano) consiste numa gama de osciladores com diferentes
frequências que devem estar em sintonia para um funcionamento ótimo. O grau de
acoplamento estre os osciladores pode induzir diferentes padrões rítmicos: quando o
grau de acoplamento é alto, os osciladores apresentam um ângulo de fase estável e
geram um único ritmo; se os osciladores estiverem desacoplados, outros padrões
podem ser gerados (Vilaplana e cols., 1997).
O acoplamento dos osciladores pode ser um processo gradual, como foi
demonstrado por Campuzano e colaboradores (1998): quando o ritmo do CE é
próximo ao período endógeno (por exemplo, de 23,5h), o sistema já é capaz de
sincronizar completamente com o ciclo externo e apresenta um único ritmo de
atividade locomotora. Isso significa que a região ventrolateral (vl) do NSQ, que
recebe o estímulo luminoso, é capaz de arrastar o ritmo da região dorsomedial (dm)
do NSQ e ajustá-lo ao CE externo. Entretanto, há o limite mínimo de sincronização à
luz, onde os animais não conseguem mais sincronizar-se completamente ao ciclo de
claro/escuro imposto. A região vl segue o ritmo imposto pelo CE ambiental, já que
recebe projeções diretas da retina, enquanto a região dm dos NSQs não consegue
25
sincronizar-se a ciclos menores do que 23,5h e como consequência, os osciladores
ficam parcialmente desacoplados, o que significa que cada região do oscilador
apresenta-se em uma fase diferente, mandando sinais diferentes para o resto do
organismo no lugar de sinalizar uma única fase. Com isso, a atividade locomotora
dos animais acaba seguindo tanto a sinalização proveniente da região vl do NSQ
como também responde a sinalização proveniente da região dm do NSQ. O
comportamento final é a dissociação do ritmo de atividade locomotora, na qual um
único animal apresenta dois componentes rítmicos: um deles sinalizado pela região
vl e sincronizado ao ciclo CE externo; e outro componente sinalizado pela região dm,
não sincronizado ao CE, com período maior que 24h semelhante ao livre curso
(Figura 1).
Figura 1. Actogramas mostrando o ritmo de atividade dos animais sobre diferentes regimes
de luz com períodos simétricos de 21h; 21,5h; 22h; 23h e 23,5h (Campuzano e cols., 1998).
Cambras e colaboradores (2004) realizaram um trabalho no qual os animais
foram expostos a diferentes fotofases variando o período entre 22 ou 23h. Sob estes
protocolos, os ratos apresentaram uma gama de padrões de atividade locomotora,
desde o arrastamento total do ritmo (ajustado ao ciclo claro/escuro) até o ritmo em
livre curso (expressão do componente endógeno). Os animais submetidos ao
26
período de 23h e longas fotofases ficaram sincronizados ao ciclo claro/escuro
externo, enquanto os animais submetidos ao período de 22h e fotofases curtas
apresentaram o ritmo da atividade locomotora em livre curso (maior que 24h). Os
autores sugeriram que o processo de sincronização do sistema de temporização
circadiano pode ser gradualmente distribuído entre o componente circadiano
dependente de luz e o componente não sincronizado à luz que apresenta o período
semelhante ao ritmo endógeno do animal, indicando que populações distintas de
osciladores podem estar em diferentes níveis de (des)acoplamento.
A fim de investigar o desacoplamento dos osciladores centrais (NSQs) o
mesmo grupo (de la Iglesia e cols., 2004) buscou um marcador evidente da
ritmicidade circadiana e investigou a expressão de RNAm dos genes relógio Per1 e
Bmal1 que se expressam alternadamente de dia e de noite, respectivamente. Como
dito anteriormente, sob o protocolo de dessincronização forçada (T22, 11h claro e
11h escuro) dois componentes do ritmo de atividade locomotora são expressos
simultaneamente no animal (T22 e T>24). Entretanto, como possuem períodos
diferentes, esses ritmos acabam coincidindo em certos momentos e com isso
podemos separar o comportamento de um mesmo animal em quatro momentos
distintos (Figura 2):
A – quando há coincidência da fase de baixa atividade locomotora dos dois
componentes rítmicos; ambos se encontram na fase dia, coincidindo o dia subjetivo
com o dia objetivo (claro);
B – quando há coincidência da fase de alta atividade locomotora em ambos
os componentes rítmicos, coincidindo então a noite subjetiva com a noite objetiva
(escuro). Neste trabalho chamaremos esta fase de CAP (do inglês: coincidence
active phase);
C – representa a fase em que os dois componentes rítmicos estão em não
coincidência, o animal apresenta atividade locomotora correspondente à noite
subjetiva, mas encontra-se na fase clara, ou seja, no dia objetivo;
27
C
A
B
D
Figura 2. Actograma de um rato mantido em um ciclo claro/escuro de 22h (11h claro, 11h
escuro) e plotado duplamente, onde cada linha representa dois dias: o dia n seguido do dia
n+1 horizontalmente, na segunda linha estão representados os dias n+1 e n+2, e assim
sucessivamente.
Dois componentes rítmicos da atividade locomotora estão sendo
expressos simultaneamente com diferentes períodos: um deles está sincronizado com o
ciclo claro/escuro (T22, em cinza abaixo da barra preta) e outro em livre curso, não
sincronizado com o ciclo CE, que apresenta um período maior que 24h (T>24) apontado
pela linha em vermelho. A, B, C e D indicam as diferentes fases as quais os animais
estavam no momento (ver texto).
28
D – o inverso de C, o animal apresenta atividade locomotora correspondente
ao dia subjetivo (repouso), mas está na noite objetiva, ou seja, na fase escura. Neste
trabalho chamaremos esta fase de NCAP (do inglês: non-coincidence active phase).
Ao avaliar a expressão dos genes da circuitaria molecular circadiana nos
NSQs nessas diferentes fases, os autores observaram que as regiões dm e vl dos
NSQs realmente se desacoplam, expressando-se alternadamente. O RNAm do gene
Per1, que é encontrado em altas concentrações durante a fase clara (dia objetivo),
quando sincronizado com a pista externa, é expresso em todo o corpo dos NSQs
(Figura 3A) e, quando a fase escura (noite objetiva) está sincronizada com a
atividade do animal (noite subjetiva), não há expressão de RNAm nos NSQs (Figura
3B). Portanto, em momentos de coincidência (Figura 2, momentos A e B), os NSQs
comportam-se igualmente aos dos animais em T24: as duas regiões dos NSQs
estão na mesma fase e expressam Per1 durante o dia e não o expressam durante a
noite. Todavia, quando não há coincidência entre os dois componentes (Figura 2,
momentos C e D), a expressão de RNAm do gene Per1 na região vl segue conforme
a sinalização do ciclo CE, ou seja, quando o animal apresenta atividade
correspondente a noite subjetiva, o que é contraditório, uma vez que o animal está
no claro, no dia objetivo (Figura 3C) e mais expresso na região dorsomedial quando
expressa o dia subjetivo na noite objetiva (Figura 3D). O contrário foi observado para
RNAm do gene Bmal1, que é expresso em antifase ao gene Per1. Assim, sugere-se
que as regiões dos NSQs se desacoplam quando não há coincidência das fases de
dia subjetivo e objetivo, deixando claro que a região ventrolateral segue o padrão
rítmico sincronizada às pistas ambientais e a expressão dos clock genes na região
dorsomedial reflete o ritmo endógeno do animal (de la Iglesia e cols., 2004).
Esse fato sugere que o desacoplamento de subpopulações anatomicamente
identificáveis de neurônios oscilantes nos NSQs por si só podem levar a
dessincronização de diferentes processos fisiológicos e circadianos, semelhante ao
observado em humanos. Portanto, os ritmos circadianos podem ser desacoplados
de maneira previsível e estável nesse modelo de dessincronização forçada em ratos.
Nessas circunstâncias, a regulação da temperatura corporal circadiana e do sono
29
REM demonstra as mesmas propriedades em humanos em dessincronização
forçada (Cambras e cols., 2007).
Figura 3. Secção coronal de cérebro mostrando a expressão dos genes Per1 e Bmal1 nos
NSQs de ratos sacrificados nos momentos A, B, C e D descritos na figura 1 (de la Iglesia e
cols., 2004).
Estudos do nosso laboratório (Neto, 2006) mostraram que ratos submetidos à
dessincronização forçada apresentam um comportamento semelhante à mania
quando testados durante a fase de coincidência da noite subjetiva com a noite
objetiva. Uma avaliação comportamental durante a fase de não coincidência sugeriu
um padrão comportamental semelhante à depressão (Araujo, dados não publicados,
Gonzalez e de la Iglesia 2008). Adicionalmente, uma análise do EEG mostrou que
ratos na fase de mania/coincidência apresentam um padrão de hipossonia em
oposição à fase de depressão/não coincidência onde é observado um padrão de
hipersonia (Gonzalez e de la Iglesia 2008).
Dessa forma, listo a seguir os objetivos do meu doutorado.
30
2. OBJETIVO GERAL
Avaliar se o protocolo de dessincronização forçada atende os critérios de
validação para um modelo animal do transtorno bipolar.
2.1. Objetivos específicos
Validação aparente (ou de face) através dos testes comportamentais: campo
aberto; labirinto em cruz elevado; nado forçado e teste de consumo de glicose
dos animais dessincronizados durante a fase de coincidência e não
coincidência;
Validação preditiva (ou farmacológica) com administração de lítio a fim de
verificar se os parâmetros comportamentais alterados são revertidos;
Validação do construto (ou etiológica) com a avaliação da transmissão
serotoninérgica em áreas límbicas, como amígdala e córtex pré-frontal,
através das técnicas de imunohistoquímica e hibridização in situ para os
receptores 5HT1A e 5HT2A.
31
3. MATERIAIS E MÉTODOS
Conforme os objetivos deste trabalho foi necessário submeter os animais ao
protocolo de dessincronização forçada descrito por Cambras e cols. (2004). Com
esta finalidade, os ratos foram submetidos a um ciclo claro/escuro com período de
22h, sendo 11h claro e 11h escuro. Ao fim de um mês a maioria dos animais
apresentava claramente os dois componentes rítmicos da atividade locomotora
dissociados (Figura 2).
Como cada animal passa por momentos de CAP e NCAP alternadamente, os
grupos foram separados conforme o momento no qual o animal estava no primeiro
dia de teste comportamental. Por exemplo, se um animal estava no momento de
CAP no dia do teste do campo aberto (CA) – que foi o primeiro aplicado – então,
este animal ficou incluso no grupo CAP e todos os testes seguintes foram aplicados
quando o animal estava no momento CAP. A mesma lógica foi utilizada para montar
o grupo NCAP. Cabe salientar que todos os animais experimentais foram
submetidos ao mesmo protocolo e posteriormente separados nos grupos CAP e
NCAP conforme o momento no qual estavam durante o primeiro teste.
Os dias para a realização dos testes comportamentais foram selecionados
com o auxílio de uma rotina no MatLab (descrita no Anexo I). A rotina foi
desenvolvida para acompanharmos a transição entre as fases individualmente com
a finalidade de visualizar qual a melhor data para a execução dos testes. Além disso,
a confirmação do melhor momento e dia para a realização dos testes foi confirmada
através das análises dos actogramas por dois pesquisadores. Todos os
experimentos comportamentais com os animais dissociados foram executados no
meio da fase escura (ZT16-17) e comparados com os resultados dos testes dos
animais controles que também foram realizados no meio da fase escura (ZT18).
Os animais controle e os animais da bateria de testes foram expostos ao ciclo
claro/escuro com período de 24h: 12h claro e 12h escuro durante todo o
experimento.
32
Este projeto foi aprovado pelo Comitê de ética para o uso de animais da
UFRN sob protocolo nº 021/2010.
3. 1. Para a Validação Aparente
3.1.1. Animais
Foram utilizados ratos machos Wistar, provenientes do biotério do Laboratório
de Neurobiologia e Ritmicidade Biológica do Departamento de Fisiologia da UFRN,
com aproximadamente 60 dias de vida no início do experimento. Antes dos
experimentos, os animais estavam submetidos às condições de CE do biotério que é
de 12:12 (as luzes acendem às 6h e apagam às 18h). Durante todo o experimento,
os animais tiveram livre acesso a alimentação e água, e foram mantidos em gaiolas
individuais, em ambiente com ciclo claro/escuro controlados conforme o grupo em
que estavam (controles 12:12 ou dissociados 11:11), temperatura entre 21 e 22°C e
som atenuado.
3.1.2. Testes comportamentais
Os horários e dia dos testes comportamentais foram selecionados conforme
cada grupo:
- Grupo CAP (T22): no meio da fase escura (entre ZT 16 e 17) quando ocorre
coincidência da noite objetiva com a noite subjetiva do animal, ou seja, fase escura
coincidindo com a fase de maior atividade do animal;
- Grupo NCAP (T22): no meio da fase escura (entre ZT 16 e 17) quando o animal
apresenta atividade reduzida na noite objetiva.
O número de animais utilizados para os testes comportamentais foi: n=13
para os animais na condição de CAP; n=14 para os animais na condição de NCAP e
n=8 para os animais do grupo controle.
33
Para a bateria de testes foram utilizados 5 ratos para cada grupo (n=5) que
permaneceram sob o regime de luz T24: 12/12, como os controles.
Foram selecionados quatro dos testes comportamentais mais utilizados na
literatura para testar ansiedade, depressão e anedonia. Segue a descrição de cada
um deles.
3.1.2.1. Labirinto em cruz elevado
O labirinto em cruz elevado (LCE) é uma ferramenta utilizada para avaliar
principalmente a ansiedade e a avaliação de risco a qual o animal é exposto
(Carobez e Bertoglio, 2005).
O aparato é composto por quatro braços de madeira medindo, cada um, 50
cm de comprimento por 12 cm de largura. Dois dos braços são circundados
lateralmente por paredes opacas de 40 cm e são dispostos perpendicularmente aos
outros dois braços, que permanecem abertos, sem paredes laterais. Todo o conjunto
permanece elevado a 50 cm do solo (Figura 4).
Os animais foram colocados no centro do aparato, com a face voltada para o
braço aberto, e permaneceram por 5 minutos no teste. Ao ser colocado no aparato,
é comum o animal preferir os braços fechados, pois ratos apresentam tigmotaxia –
preferem estar em contato com paredes verticais – e neste local sentem-se mais
seguros. O tempo que o animal passa nos braços abertos e a porcentagem do
número de entradas nesses braços são as medidas utilizadas para a determinação
do índice de ansiedade. Conjuntamente, o número de entradas nos braços fechados
é utilizado para avaliar a atividade locomotora do animal (Carobez e Bertoglio,
2005). A avaliação de risco também foi mensurada neste teste através da
observação do comportamento de stretching, movimento no qual o animal localizado
no centro do aparato se alonga próximo à entrada do braço aberto para avaliar o
quão seguro é o local antes de entrar.
34
Figura 4. Ilustração do aparato para o teste do labirinto em cruz elevado (LCE).
3.1.2.2. Campo aberto (Insight® modelo EP 154)
O teste de campo aberto (CA) foi realizado numa arena circular com 60 cm de
diâmetro dividida em quadrantes centrais e periféricos, cercada de paredes acrílicas
de 50 cm (altura), base em acrílico de 100 cm x 80 cm (Figura 5). Os animais foram
inicialmente colocados no centro do aparato e observados por 5 minutos.
Foram aferidos: o deslocamento total, o deslocamento no centro e na periferia
do aparato, o tempo gasto em cada uma das áreas e a taxa de defecação. Também
avaliamos o comportamento de grooming e rearing. O grooming é caracterizado por
movimentos repetitivos de autolimpeza que, quando em grande quantidade, são
indicativos de stress (Kalueff e Tuohimaa, 2005). Já o rearing é quando o animal fica
sobre as duas patas traseiras realizando a exploração vertical do aparato.
O teste do campo aberto é um clássico para avaliar comportamento locomotor
e emocionalidade. Os animais que exploram mais o centro do campo aberto são
considerados menos ansiosos do que os animais que evitam cruzar ou permanecem
menos tempo nos quadrantes centrais (Prut e Belzung, 2003).
35
Figura 5. Ilustração do aparato campo aberto utilizado.
3.1.2.3. Teste do nado forçado (Insight® modelo EP 180)
O teste do nado forçado (NF) foi realizado num aparato cilíndrico de acrílico
de 50cm de altura com diâmetro de 30cm (Insight® – Figura 6), cheio de água até
15cm antes da borda, abaixo dos sensores, para que o animal não consiga fugir
nem tocar o fundo do aparato. Este teste tem duração de 5 minutos.
Neste teste o animal é exposto ao estímulo aversivo e estressante – é
colocado dentro da água sem possibilidades de fuga – e então é mensurado o
“comportamento de desespero”. Chamamos de comportamento de desespero
movimentos intensos e vigorosos das quatro patas, numa tentativa de escapar do
cilindro. É mensurada a latência para cessação deste comportamento, o que implica
que quanto maior for a latência, mais o animal lutou pela vida; e o contrário, latência
reduzida, significa que o animal apresenta comportamentos tipo depressivos, já que
desiste mais facilmente (Castagné e cols, 2009).
36
Figura 6. Ilustração do aparato utilizado para o teste do nado forçado (Insight® modelo EP
180).
3.1.2.4. Preferência Claro - Escuro (Insight® modelo EP 157)
O equipamento é composto por dois compartimentos separados por uma
divisória com uma pequena passagem entre o lado claro (revestido com chapa de
acrílico branco - A x L x P (cm) 21 x 45 x 41) e o lado escuro (revestido com chapa
de acrílico preto - A x L x P (cm) 21 x 35 x 41), com tampa de acrílico transparente
(Figura 7). Os animais iniciam o teste no lado claro do aparato e foram observados
por 5 minutos.
Para avaliar o nível de ansiedade e o comportamento exploratório foram
aferidos: o tempo de permanência e o deslocamento em cada um dos lados, bem
como o número de episódios de comportamentos de rearing e grooming, tanto no
lado claro quanto no lado escuro.
37
Figura 7. Foto ilustrativa do aparato de preferência ao claro ou escuro.
Todos os testes descritos até aqui foram inteiramente filmados, para qualquer
possível reavaliação posterior.
3.1.3. Bateria de testes
Consideramos prudente avaliar previamente a melhor sequência para a
realização dos testes comportamentais de modo que a execução de um teste não
interfira nos resultados do seguinte. Como a literatura sobre este assunto está mais
voltada para experimentos comportamentais em camundongos (caracterização de
cepas e animais geneticamente modificados) (van Galeen e Steckler, 2000; McIlwain
e cols., 2001; Lad e cols., 2010) e, além disso, não abrangem alguns dos testes que
foram por nós utilizados, elaboramos um experimento piloto no qual os animais
realizaram baterias de testes de avaliação comportamental em diferentes
sequências, como mostra a Tabela 1.
Tabela 1. Grupos experimentais e respectiva sequência de testes.
Grupos
1º Teste
2º Teste
3º Teste
4º Teste
G1
CA
LCE
NF
PCE
G2
LCE
CA
NF
PCE
G3
NF
CA
LCE
PCE
G4
NF
LCE
CA
PCE
CA – teste de Campo Aberto; CE – teste de preferência Claro – Escuro; NF – teste de Nado Forçado; LCE –
teste de Labirinto em Cruz Elevado
38
Foram utilizados 20 ratos Wistar machos com 3 meses de idade no início do
experimento, divididos em 4 grupos com 5 animais cada. Os testes tiveram inicio às
18:30h (ZT12,5), correspondente às primeiras horas da fase escura, momento no
qual os ratos tem maior atividade exploratória. Cada grupo experimental realizou um
único teste por dia, durante quatro dias consecutivos. O intervalo entre os testes foi
de 24h. Todos os testes foram realizados no escuro total.
Os resultados deste experimento foram válidos para estabelecer em qual
sequência ocorre o menor efeito de ordem.
3.1.4. Atividade locomotora
Os animais permaneceram alojados em gaiolas individuais com sensores
infravermelhos 5cm acima de cada uma, a fim de registrar a atividade locomotora
individualmente. Os dados de registro da atividade locomotora e os ciclos de luz
foram controlados continuamente pelo sistema SAP – Sistema de Aquisição
Periódica (desenvolvido pelo LNRB, Dr. Marconi Câmara) durante todo o
experimento.
Periodicamente foram construídos gráficos para visualização do ritmo de
atividade locomotora – actogramas e periodogramas – no programa El Temps (Dr.
Antony Dièz-Nogueira, Universidade de Barcelona, 1999), a fim de acompanhar a
dissociação dos componentes rítmicos, tanto do componente sincronizado ao ciclo
CE imposto (T22) quanto do componente não sincronizado ao CE (T>24). O
acompanhamento e a análise da atividade locomotora durante o experimento
farmacológico também foi realizada dessa forma.
39
3.1.6. Teste de Consumo de Glicose
O teste de consumo de glicose consiste num paradigma de escolha entre
duas garrafas: uma delas contendo água e a outra contendo uma solução de 0,5%
de glicose diluída em água. As duas garrafas são igualmente oferecidas para os
animais, como mostra a figura 8, por um período 24h. Ao fim do teste, é aferido o
consumo individual de cada uma das garrafas. Este teste tem sido utilizado
extensivamente há mais de 20 anos em modelos animais de depressão para
mensurar a anedonia (Willner e cols., 1987), um sinal crucial da depressão e
indicador comportamental do estado depressivo em roedores. O animal anedônico,
depressivo, perde a habilidade de sentir prazeres, portanto teoricamente não terá
preferência pelo sabor doce.
Figura 8. Foto ilustrativa das condições do teste de consumo de glicose.
3.2. Teste farmacológico para a Validação Preditiva
Como o lítio é o tratamento mais comum para o transtorno bipolar, o
elegemos para ser utilizado neste experimento a fim de verificar se essa condição de
40
ciclicidade era capaz de ser revertida. O lítio foi administrado na forma de carbonato
de lítio (Li2CO3), através do medicamento Carbolitium® (Eurofarma).
A primeira tentativa de administração do lítio foi feita através da diluição do
medicamento na água. Para tanto, mensuramos a quantidade de água ingerida
pelos animais (~20ml/dia) e fizemos uma média do consumo para a correta diluição
do medicamento. Baseando-se na média do consumo de água, diluímos inicialmente
1mg de lítio/4ml água para que fosse administrada a dose diária correta do
medicamento. Entretanto, os animais passaram a consumir menos líquido. Por esse
motivo, reajustamos a concentração do lítio na água para 6mg/4ml. Dessa forma, os
animais passaram a consumir ainda menos água, alguns chegaram a ficar
completamente privados por dias. Com isso, a única alternativa foi modificar a via de
administração.
Como o protocolo de dessincronização forçada é baseado na ritmicidade
circadiana, a administração do fármaco não poderia interferir na ritmicidade dos
animais. Então, a via de administração eleita foi através da ração. Quebramos os
pellets de ração até que virassem pó e umedecemos a massa para ser misturada ao
medicamento macerado na concentração de 3,6g/kg de ração. Após estarem bem
misturados, refizemos os pellets, assamos no forno para secar e esperamos esfriar
antes de disponibilizá-las ad libitum por 21 dias para os ratos. Essa dose foi
determinada através de estimativas do consumo de ração. Iniciamos com 2mg de
lítio/kg de ração e, mensurando a litemia dos animais duas vezes na semana,
conseguimos atingir a litemia desejada de >1,0 mEq/ml na maioria dos animais com
a dose de 3,6mg/kg. A litemia foi aferida no Centro de Patologia Clínica de Natal,
através do aparelho AVL 9180 (Roche) que utiliza a metodologia de eletrodo íon
seletivo.
Neste experimento foi avaliada a atividade locomotora dos animais tratados
em suas homecages, bem como o ritmo desta, a fim de avaliar o efeito do lítio nos
animais em dissociação forçada.
41
3.3. Testes neuroquímicos para a Validação do Construto
3.3.1. Hibridização in situ do receptor de serotonina 5HT2A
Este procedimento foi realizado no LabCrono do Professor Horacio de la
Iglesia na Universidade de Washington durante o doutorado sanduíche.
Submetemos 30 ratos Wistar machos com 2 meses no início do experimento
ao protocolo de dessincronização forçada. Após 45 dias, 24 deles apresentavam
claramente os dois componentes rítmicos da atividade locomotora bem dissociados.
Esses animais foram sacrificados por decaptação em momentos diferentes
compondo 4 grupos, com n=6 cada:
1 – CAP fase escura (ZT16);
2 – CAP fase clara (ZT5);
3 – NCAP fase escura (ZT16);
4 – NCAP fase clara (ZT5).
O grupo controle deste experimento foi composto por 10 animais submetidos
ao ciclo claro/escuro T24h. Metade (n=5) foi sacrificada na fase clara (ZT6) e metade
na fase escura (ZT18).
Os encéfalos foram retirados e imediatamente imersos em metilbutano a
-30ºC para congelamento. Para este experimento foi utilizado somente o hemisfério
direito dos animais, pois o outro hemisfério foi destinado a outro experimento do
laboratório. Os cérebros congelados foram mantidos a -80ºC até serem fatiados em
slices de 16µm no criostato e dispostos em lâminas de vidro para o processamento.
O mesmo material foi utilizado para a imunohistoquímica descrita no item a seguir.
Foi utilizado o protocolo de hibridização in situ descrito em de la Iglesia (2007)
com algumas modificações. O protocolo do procedimento completo para a obtenção
das imagens encontra-se no Anexo II.
42
As probes foram gentilmente cedidas pelo professor Juli Hsiao-Huei Wu da
Keck School of Medicine da University of Southern California e correspondem às
sequências descritas no Anexo III.
Os filmes autorradiográficos (Kodak®) foram expostos por 5 dias em escuro
total. Cada densidade atribuída é o resultado da média de dois slices de cada
animal. Os filmes foram escaneados e analisados através do programa ImageJ para
a obtenção da densidade óptica das áreas selecionadas: amígdala e córtex préfrontal nas regiões pré-límbica e pré-motora.
3.3.2. Imunohistoquímica para o receptor 5HT1A
Como a intenção era fazer as duas técnicas (hibridização in situ e
imunohistoquímica), foi realizado o mesmo tratamento descrito no item anterior.
Portanto,
os
animais
não
foram
perfundidos
para
a
realização
das
imunohistoquímicas. É sabido que sem a perfusão os tecidos ficariam um pouco
prejudicados para a imunohistoquímica, mas optamos por realizar esta técnica assim
mesmo para aproveitarmos o tecido oriundo deste experimento, a fim de investigar a
quantidade de receptores expressos e não só a expressão de RNAm dos mesmos.
Para a detecção do receptor 5HT1A, o anticorpo utilizado foi SR-1A (H-119)
da Santa Cruz Biotechnologies (SC-10801) na concentração de 2ug/ml. O anticorpo
é uma IgG policlonal feita em coelho, cujo epitopo corresponde aos aminoácidos 218
– 336 do receptor de serotonina 1A de seres humanos recomendado para
identificação deste mesmo receptor em camundongos, ratos e humanos.
Para a detecção do receptor 5HT2A, o anticorpo utilizado foi o SR-2A (A-15)
da Santa Cruz Biotechnologies (SC-15073) que é uma IgG policlonal feita em cabra,
cujo epitopo correspondente localiza-se próximo a região N-terminal do receptor
5HT2A de humanos. Entretanto, não consegui realizar a padronização desta reação
enquanto estive no laboratório do professor Horácio de la Iglesia, ficando este
experimento a ser realizado a posteriori.
43
Para revelação foi utilizado o kit ABC da Vector Laboratories® e DAB (3, 3'diaminobenzidine). O protocolo completo desta técnica está descrito no anexo IV.
As fotos foram tiradas no LabNeuro da UFRN, através do microscópio óptico
Olympus BX41 de campo claro. As imagens foram digitalizadas pela câmera Nikon
DXM1200 acoplada ao microscópio.
A contagem de células foi feita através do programa ImageJ, onde todas as
estruturas circulares ou ovoides que superavam a densidade óptica média somada a
três vezes o desvio padrão (M+(3xSD)) do background em 16 bits. As regiões
escolhidas para a análise foram amígdala basolateral e córtex pré-frontal região prélímbica. Foi tirada uma única foto em aumento de 40x da área desejada e contadas
todas as células marcadas presentes no campo.
Ficaram alocadas 6 a 9 slices do hemisfério direito de um animal em cada
lâmina. Destas, foram eleitas as duas melhores e feita a soma da contagem de
células para que cada animal tivesse um único número para as análises estatísticas.
Optamos pela soma, pois como os cortes possuem mais de 100µm de distância
entre eles (devido a confecção de 6 séries), o mesmo neurônio não seria contado
mais de uma vez.
3.4. Análise estatística
Inicialmente foram realizadas estatísticas descritivas para a análise visual dos
dados determinando os valores de média, erro padrão e os gráficos.
Para avaliar os parâmetros da atividade locomotora, comportamentais e o
consumo de glicose, primeiramente foi aplicado o teste de Kolmogorov-Smirnov para
avaliar a distribuição dos dados. Nos casos em que a distribuição obedece a uma
curva normal foi aplicado o teste ANOVA de uma via, seguido do teste pós-hoc de
Tukey. Para a análise molecular foi aplicada a ANOVA de duas vias, dadas as
diferentes condições de iluminação dos grupos, com posterior teste de pós-hoc de
Tukey. O nível de significância adotado foi de p<0,05.
44
4. RESULTADOS
4.1. Resultados dos testes comportamentais para Validade Aparente
4.1.1. Atividade locomotora
Entre os animais dissociados, não há diferença entre as médias do total de
atividade por hora quando analisadas as fases de claro e escuro conjuntamente, ou
seja, a quantidade de atividade locomotora diária dos animais em CAP não difere
dos animais em NCAP num período de 22h (610,7±66,2ua/h x 517,4±39,1ua/h;
p=0,08). Entretanto, os animais em CAP apresentam atividade aumentada em suas
gaiolas comparada a dos animais controle (610,7±66,2ua/h x 458,84±57,8ua/h;
p=0,001), mostrando que os animais dissociados em fase de coincidência (CAP)
apresentam hiperlocomoção, diferentemente dos animais em NCAP que apresentam
a média diária de atividade locomotora semelhante a média diária dos animais
controle (517,4±39,1ua/h x 458,84±57,8ua/h; p=0,34). Todavia, quando analisamos
separadamente cada uma das fases do ciclo de iluminação podemos observar
claramente a diferença no perfil diário de atividade locomotora entre os grupos CAP
e NCAP. Os animais, quando estão no momento de CAP, apresentam mais
atividade na fase escura e menos atividade na fase clara, característica de animais
noturnos (ratos), assim como os controles. Porém, quando estão no momento de
NCAP esse quadro se modifica. Os animais apresentam atividade semelhante no
claro e no escuro (Figura 9).
Como o perfil de atividade entre os animais em CAP e os controles é
semelhante (ver Figura 10), durante a fase clara não foi observada diferença na
quantidade de atividade (320,0±24,6ua/h x 211,8±34,9ua/h; p=0,45). Entretanto, há
um grande aumento da atividade nos animais em CAP durante a fase escura
quando comparados ao grupo controle (901,4±30,3ua/h x 705,9±40,5ua/h; p=0,02).
Isso mostra que o aumento de atividade diária dos animais em CAP está
concentrado na fase escura, mostrando que a hiperlocomoção do grupo CAP ocorre
nesta fase. Entre os grupos NCAP e controle as diferenças também foram
marcantes
(claro:
576,8±43,7ua/h
x
234,7±42,0ua/h;
p=0,0001;
escuro:
45
446,1±47,9ua/h x 723,7±45,7ua/h; p=0,001), devido a diferente distribuição da
atividade nessas duas fases (Figura 9).
Figura 9. Perfil da atividade locomotora em média por hora nos grupos CAP (vermelho),
NCAP (azul) e Controle (preto). O eixo Y traz o tempo em segundos/ h da movimentação do
animal, No eixo x a escala temporal em horas em zeitgeber time.
*
*
*
Figura 10. Atividade locomotora expressa em segundos/h. À esquerda estão apresentadas
as médias do período completo (22 ou 24h) dos grupos, onde podemos observar a diferença
entre o grupo controle e o grupo CAP. No centro, a atividade locomotora na fase clara é
maior no grupo NCAP quando comparada aos grupos CAP e controle. À direita, na fase
escura, a atividade locomotora é diferente quando comparamos os grupos aos pares (Ctl ≠
CAP; Ctl ≠ NCAP; CAP ≠ NCAP). *p<0,05.
46
Quando realizamos a análise intragrupo, comparando a atividade da fase
clara com a da fase escura entre os grupos, o esperado seria encontrarmos
diferenças em todos os grupos já que ratos possuem atividade noturna e a fase de
repouso alocada na fase clara. Esse fato ocorre com os animais do grupo CAP
(claro 320,0±24,6ua/h x escuro 901,4±30,3ua/h; p=0,0001) e com os animais do
grupo controle (claro 211,8±34,9ua/h x escuro 705,9±40,5ua/h; p=0,0001), porém
não ocorre nos animais do grupo NCAP (claro 546,3±42,9ua/h x escuro
488,6±66,4ua/h; p=0,92) que possuem quantidade semelhante de atividade
locomotora na fase clara e na fase escura, mostrando um deslocamento da fase
ativa do animal.
As análises da atividade locomotora foram realizadas através de ANOVA de
uma via para a comparação entre a atividade diária e ANOVA de duas vias para a
comparação entre a atividade no claro e no escuro.
Através da rotina de MATLAB descrita por Gonçalves e colaboradores (2010)
pudemos verificar a variabilidade intradiária (IV), ou seja, a fragmentação da
atividade locomotora dentro de um dia. A IV é semelhante para os indivíduos NCAP
e controle (1,08±0,03 x 1,07±0,03; p=0,97). Entretanto, os animais em CAP mostram
fragmentação da atividade bastante reduzida comparada com os outros grupos
(0,88±0,03 x NCAP p=0,0008; CAP x Ctl p=0,0007).
4.1.2. Testes comportamentais
4.1.2.1. Bateria de testes
Os animais foram testados conforme a sequência especificada na Tabela 1.
Ao final do experimento, o que observamos foi um aumento da ansiedade nos
animais que iniciam a bateria pelo LCE (G2) quando comparados aos animais que
iniciam o teste pelo CA (G1). Os animais que iniciam a bateria de testes pelo LCE,
quando o segundo teste é o CA, apresentam menor exploração no centro do CA,
mostrando que depois de realizarem o teste do LCE os animais ficam mais ansiosos
do que os animais cujo primeiro teste é o CA (G1: 24,2±3,2 quadrantes x
47
G2:13,2±2,2; p=0,027) (Figura 11). Entretanto, o deslocamento na periferia do CA,
bem como o deslocamento total durante o teste, foi semelhante entre esses grupos.
Esse aumento da ansiedade no G2 também pode ser observado pela análise do
teste de PCE, onde estes permaneceram menos tempo no compartimento claro do
que os animais do G1 (G1: 158,6±10,6s x G2:145,2±11,8s, p=0,008) mesmo que
ambos tenham realizado o teste do NF 24h antes, mostrando que o LCE não é
sugestivo para iniciar a bateria de testes.
*
Figura 11. Distância percorrida no teste do campo aberto. O gráfico mostra a distância em
número de quadrantes percorridos em média por grupo em separado: à esquerda, no centro
e à direita na periferia do aparato no experimento bateria de testes (*p<0,05).
*
Os outros parâmetros avaliados nos testes LCE e NF não apresentaram
diferença entre os grupos na sequência, revelando que não sofrem interferência
significativa do teste antecedente.
As análises entre os grupos foram feitas através de ANOVA de uma via.
48
Com base nestes resultados, concluímos que a ordem mais adequada para
serem realizados os experimentos é do teste menos ansiogênico, nesse caso o CA,
para os mais ansiogênicos, o LCE seguido do NF.
O teste de preferência ao claro ou escuro não foi utilizado na bateria de testes
para os animais dissociados, pois realizamos um experimento piloto para verificar se
havia alguma predileção pelo lado no qual o animal iniciava o teste, o que foi
devidamente comprovado: os animais que iniciam o teste no compartimento claro
exploram mais esse lado do aparato quando comparados aos animais que iniciam o
teste no compartimento escuro (28,0±6,9 x 9,2±3,9 ; p=0,04). Optamos por não
incluir este teste na bateria comportamental dos dissociados.
Os resultados deste experimento geraram o artigo que compõe o Anexo V.
4.1.2.2. Campo aberto
Os dados obtidos no campo aberto com os animais dissociados e controle
estão exibidos nos gráficos a seguir (Figuras 12 e 13). O deslocamento foi aferido
contando-se o número de quadrantes percorridos pelo animal. Vale lembrar que os
testes foram realizados no meio da fase escura, tanto para os animais dissociados
(no ZT 16-17) quanto para os animais controle (no ZT 18).
Os animais dissociados, tanto em momentos de CAP quanto em momentos
de NCAP, apresentam menor deslocamento do que os ratos do grupo controle nos 5
minutos de teste no CA (F(2,30)=11,741; p<0,001; Posthoc Tukey CAP: p=0,008;
NCAP: p<0,0001). Quando analisamos o deslocamento dos animais na periferia do
aparato, podemos perceber que essa redução no deslocamento dos animais
dissociados ocorre principalmente na exploração dessa região quando comparados
ao grupo controle (F(2,30)=10,216; p<0,0001; Posthoc Tukey CAP: p=0,025; NCAP:
p<0,0001). No centro do CA, os animais dissociados também deslocam-se menos
do que os animais do grupo controle (F(2,30)=3,913, p=0,031; Posthoc Tukey CAP:
p=0,049, NCAP: p=0,045) e consequentemente ficam menos tempo no centro do
aparato (F(2,30)=5,065, p=0,013; Posthoc Tukey CAP: p=0,025, NCAP: p=0,018).
49
*
*
*
Figura 12. Médias do deslocamento total, no centro e na periferia do aparato. As barras em
preto representam os animais controle T24, em branco CAP e as barras em cinza os
animais em NCAP. p<0,05 entre o controle e os grupos dissociados em todos os parâmetros
avaliados.
*
Figura 13. Porcentagem do tempo despendido no centro no aparato do CA pelos grupos
Controle, CAP e NCAP. p<0,05 entre o grupo controle e os animais dissociados, ambos
CAP e NCAP.
Realizamos a análise de outros comportamentos como grooming e rearing. O
que observamos foi que os animais em dissociação, tanto CAP como NCAP,
50
apresentaram comportamentos estereotipados. Aumento na quantidade de grooming
foi observado nos animais dissociados quando comparados ao grupo controle
(F(2,33)=6,673, p=0,004; CAP: p=0,003 e NCAP: p=0,009) e redução na exploração
vertical, chamado de rearing (F(2,33)=8,236, p=0,001; CAP: p=0,001 e NCAP:
p=0,016) (Figuras 14 e 15).
Figura 14. Quantidade média de grooming realizado durante o teste do campo aberto pelos
grupos controle, CAP e NCAP respectivamente. Os animais dissociados, tanto em CAP
quanto em NCAP, apresentam mais grooming do que os animais do grupo controle. *p<0,05.
Figura 15. Quantidade média de rearing realizado durante o teste do campo aberto pelos
grupos controle, CAP e NCAP respectivamente. Os animais dissociados, tanto em CAP
quanto em NCAP, apresentam menos rearing do que os animis do grupo controle, *p<0,05.
51
4.1.2.3. Labirinto em Cruz Elevado
Foi avaliado o tempo de permanência dos animais separadamente em cada
uma das partes do aparato, bem como o número de entradas em cada local. Os
animais de todos os grupos apresentaram a atividade exploratória semelhante neste
teste. Entretanto, os animais do grupo NCAP permanecem menos tempo nos braços
abertos do labirinto quando comparado ao grupo controle (Posthoc Tukey p=0,003),
indicando maior ansiedade, como mostra a Tabela 2.
Tabela 2. Parâmetros avaliados no teste do labirinto em cruz elevado dos animais em
dessincronização forçada.* p<0,05 entre o grupo NCAP e controle.
Controle
CAP
NCAP
(Média±EP)
(Média±EP)
(Média±EP)
Tempo no braço aberto
90,2±13,4
61,2±11,6
Tempo no braço fechado
135,2±5,2
Tempo no centro
Parâmetro
ANOVA
Significância
40,3±7,0
F(2,30)= 4,35
p=0,01*
180,3±15,2
201,6±9,6
F(2,30)= 4,85
p=0,01*
74,6±7,8
58,5±4,8
58,1±4,6
F(2,30)= 1,83
p=0,17
Nº entradas no braço aberto
6,7±1,3
5,5±0,8
3,8±0,8
F(2,30)= 2,05
p=0,14
Nº entradas no braço fechado
8,7±1,4
9,4±0,9
8,7±0,8
F(2,30)= 0,18
p=0,83
Nº entradas no centro
14,8±2,5
15,1±1,5
12,9±1,5
F(2,30)= 0,58
P=0,56
Os comportamentos estereotipados também foram avaliados durante este
teste. Não houve diferença entre os grupos quanto a quantidade de grooming
(F(2,39)=1,432, p=0,252) ou rearing (F(2,29)=2,250, p=0,12). Entretanto pudemos
observar que os animais dissociados apresentaram menor avaliação do risco
quando estavam nos braços abertos do que os animais do grupo controle, um
comportamento chamado head down (F(2,39)=3,011, p=0,034), como mostra a
figura 16.
52
*
Figura 16. Quantidade média do comportamento chamado head down, para a avaliação de
risco, realizado durante o teste do labirinto em cruz elevado. O grupo controle realiza mais
vezes este comportamento do que os grupos CAP e NCAP, *p<0,05.
4.1.2.4. Nado forçado
No teste do nado forçado foi avaliada a latência em segundos para o
comportamento de desespero. Os animais do grupo CAP apresentaram a latência
significativamente aumentada quando comparada a latência dos outros grupos
(F(2,31)=6,339, p=0,005; Potshoc Tukey CAP x NCAP: p=0,014; CAP x Ctl:
p=0,011), mostrando que os animais em CAP apresentam um comportamento do
tipo antidepressivo, já que lutam mais pela vida (Figura 17).
Figura 17. Latência em segundos para o comportamento de desespero no teste do nado
forçado. A curta latência para esse comportamento reflete o estado depressivo no rato. Os
animais em CAP apresentaram latência aumentada quando comparado aos grupos controle
e NCAP. *p<0,05.
53
4.1.3. Teste de consumo de glicose
Os dados provenientes do teste de consumo de glicose estão apresentados
na figura 18 e 19. Todos os animais consumiram volume semelhante de glicose
diluída em água (F(2,36)=0,890, p=0,420). Os animais em CAP consumiram mais
água durante o teste do que os animais em NCAP (F(2,36)=5,195, p=0,011; Posthoc
Tukey p=0,009).
*
Figura 18. Consumo de consumo de água e glicose em ml. As barras em preto representam
o consumo médio dos animais do grupo controle, em branco os animais do grupo CAP e as
barras em cinza apontam o consumo dos animais do grupo NCAP. À esquerda o consumo
de água e à direita o consumo de glicose a 0,5%.
Quando comparamos o consumo intragrupo pela porcentagem de glicose no
consumo total, percebemos que os animais em NCAP têm uma tendência a um
maior consumo da glicose como mostra a figura 19, mas não é estatisticamente
significante (F(2,36)=2,597, p=0,089).
54
Figura 19. Gráfico ilustrando a porcentagem de consumo da solução contendo 0,5% de
glicose em relação ao consumo total. Não há diferença entre os grupos.
4.2. Resultados para a Validade Preditiva
4.2.1. Teste farmacológico - Lítio
Todos os animais atingiram a litemia na dosagem terapêutica descrita na
literatura (entre 1,0 e 2,0 mEq/ml) (Gould e cols., 2003; Treiser e cols., 1980; Yin e
cols., 2006). Os animais receberam a ração com lítio por 21 dias.
O lítio foi capaz de alterar a ritmicidade circadiana dos animais como
podemos observar na figura 20. Os animais previamente dissociados tiveram o ritmo
T22 completamente abolido e o ritmo >24h teve seu período prolongado de 1474±20
min para 1605±97 min em média (de 24,56h para 26,75h).
55
A
B
C
Figura 20. (A) Actogramas representativos de cinco diferentes animais sob o protocolo de
dessincronização forçada. Após 35 dias de dessincronização, iniciamos a administração do
lítio na ração 3,6g/kg. Nota-se que a administração do lítio modificou o ritmo dos animais. (B
e C) Periodogramas representativos de dois animais antes (B) e depois (C) da
administração do lítio.
Tabela 3. Quantidade de atividade expressa pelos animais experimentais em suas
homecages, antes e durante o tratamento com lítio na dosagem de 3,6mg/kg ração.
Atividade em ua/h
Dias do tratamento
(média ± erro)
Antes
219,7±16,2
5 – 10
210,1±18,9
11 – 15
205,4±19,4
16 – 20
188,8±20,1
56
Realizamos a análise da atividade locomotora a cada 5 dias de tratamento, o
que encontramos foi que paulatinamente a atividade dos animais foi reduzida, como
mostra a tabela a seguir (Tabela 3). Nos últimos 5 dias de tratamento a atividade
locomotora encontrava-se reduzida em 15% quando comparada à atividade antes do
tratamento, durante a dissociação destes mesmos animais (Figura 21).
*
Figura 21. Os pontos representam a média ±erro padrão do registro da atividade locomotora
dos animais em suas homecages antes e durante o tratamento com o lítio. A análise foi
realizada separadamente a cada 5 dias de tratamento. Nos últimos 5 dias de tratamento (de
16 a 20) a atividade foi bruscamente reduzida quando comparada ao ritmo antes do início do
tratamento (*p=0,02).
Essa redução na atividade locomotora é significante (F(3,44)=5,060,
p=0,004); os resultados dos testes de posthoc de Tukey vão sendo gradativamente
significantes comparados ao controle (5 dias p=0,054; 10 dias p=0,050; 15 dias
p=0,037) e torna-se mais evidente nos últimos 5 dias do tratamento
Entretanto, nos últimos dias de administração do lítio os animais
apresentavam-se bastante debilitados, mesmo com a administração concomitante
de cloreto de sódio 0,9% para correção do equilíbrio eletrolítico. Alguns animais
57
foram a óbito antes do final do experimento. No 21º dia de administração do
fármaco, o experimento foi interrompido para evitar o sofrimento dos animais.
Com a finalidade de facilitar o entendimento a respeito dos testes
comportamentais, a tabela 4 traz um resumo dos resultados comportamentais.
Tabela 4. Resumo dos achados comportamentais dos animais dissociados em comparação
ao grupo controle.
4.3. Testes neuroquímicos para a Validação do Construto
4.3.1. Hibridização in situ
Avaliando os resultados da hibridização in situ para o receptor 5HT2A,
verificamos que nenhuma das áreas analisadas apresenta variação circadiana na
expressão de RNAm no grupo controle, como mostra a tabela abaixo (Tabela 4).
58
Tabela 5. Expressão de RNAm do receptor 5HT2A nas fases clara e escura do grupo
controle (T24) no córtex pré-frontal pré-límbico, córtex pré-frontal pré-motor e na amígdala.
Na coluna da direita a significância do teste t.
Região
Claro
Escuro
Significância (p)
(Densidade óptica normalizada)
CPF pré-límbico
1,60±0,13
1,44±0,02
0,255
CPF pré-motor
1,70±0,06
1,60±0,05
0,276
Amígdala
1,20±0,04
1,29±0,07
0,344
As imagens da hibridização in situ estão representadas na figura a seguir
(Figura 22).
1mm
1mm
Figura 22. Hibridização in situ do córtex pré-frontal e da amígdala nos ratos dissociados,
marcando a expressão de RNAm do receptor 5HT2A. As linhas em preto representam as
áreas que foram utilizadas para a leitura da densidade óptica das áreas do córtex pré-frontal
(áreas pré-límbica e pré-motora, respectivamente) e da amígdala.
Não encontramos diferença na expressão de RNAm dos receptores 5HT2A
entre os grupos. As áreas avaliadas foram a amígdala e o córtex pré-frontal, tanto a
porção pré-motora quanto a porção pré-límbica como podemos ver nos gráficos a
seguir.
59
Figura 23. Expressão de RNAm de receptores serotoninérgicos do tipo 5HT2A na amígdala,
córtex pré-frontal pré-límbico e córtex pré-frontal pré-motor, respectivamente. As três barras
a esquerda de cada gráfico mostram a expressão durante o claro e as três barras da direita
a expressão durante o escuro. Não houve diferença estatística nos níveis de expressão de
RNAm entre os grupos.
A quantificação da densidade de receptor 5HT2A presente nas áreas
desejadas foi feita através da mensuração e posterior normalização das densidades
ópticas. Foi escolhida uma área de referência, onde a densidade óptica era mínima
aparentemente e este valor serviu como denominador para o valor mensurado da
área desejada. A fórmula a seguir resume como foi calculada a NOD (Normalized
Optical Density):
As NODs dos animais dissociados podem ser encontradas na tabela 6.
60
Tabela 6. Resultados da análise das densidades ópticas normalizadas resultantes da
hibridização in situ para o receptor 5HT2A no córtex pré-frontal pré-límbico, córtex préfrontal pré-motor e na amígdala.
Região
CAP
NCAP
Claro
Escuro
Claro
Escuro
CPF pré-límbico
1,17±0,03
1,24±0,02
1,49±0,04
1,45±0,06
CPF pré-motor
1,66±0,03
1,59±0,06
1,69±0,05
1,53±0,07
Amígdala
1,24±0,02
1,17±0,03
1,21±0,01
1,26±0,04
4.3.2. Imunohistoquímica
Primeiramente verificamos se havia variação circadiana do receptor 5HT1A
no grupo controle. Vimos que tanto na região pré-límbica do córtex pré-frontal
(F(2,31)= 0,359; p=0,702) quanto na região da amígdala (claro: 178,8± 17,3 x
escuro: 151,0±13,9; p=0,2) não há diferença entre a fase clara e a escura, quanto ao
número de células que expressam o receptor 5HT1A. Em outras palavras, não há
variação circadiana no número de células que expressam este receptor nas áreas
analisadas no grupo controle.
Ao comparar a marcação entre os grupos encontramos maior número de
células marcadas nos animais dissociados do que nos animais controle quando
considerada a região da amígdala (F(2,33)=8,481; p=0,001; Posthoc Tukey CAP:
p=0,006; NCAP: p=0,002).
Quando analisamos nas diferentes condições de luz em separado, os animais
sacrificados em NCAP na fase escura apresentaram maior número de receptores
5HT1A na região da amígdala quando comparados aos animais controle
(F(5,33)=4,379; p=0,005; Posthoc Tukey p=0,02), como mostrado na tabela 7. Os
animais em CAP não apresentaram diferença estatística dos animais controle
quando analisados nas diferentes condições de luz.
61
Tabela 7. Contagem de células 5HT1A positivas no córtex pré-frontal e na amígdala.
Córtex pré-frontal
Amígdala
Escuro
Claro
Escuro
Claro
Controle
177,4±21,8
186,8±12,9
151,0±13,9
178,8±17,3
CAP
204,5±26,6
188,5±9,5
195,6±11,9
218,0±9,3
NCAP
185,5±14,0
193,2±12,1
210,3±5,9
216,6±13,7
A contagem de células marcadas para o receptor 5HT1A na região do córtex
pré-frontal
não
foi
estatisticamente
significante
nas
diferentes
condições
experimentais e de luz.
A figura 24 representa a marcação do receptor 5HT1A respectivamente em
cortes do córtex pré-frontal e amígdala.
50µm
50µm
Figura 24. Foto representativa das células marcadas para o receptor 5HT1A em um corte
do córtex pré-frontal à esquerda e um corte da amígdala à direita.
62
5. DISCUSSÃO
No presente estudo tivemos por objetivo promover alterações na ritmicidade
circadiana em animais saudáveis a fim de verificar se estas poderiam causar
alterações comportamentais semelhantes às apresentadas no transtorno bipolar. O
método eleito foi o protocolo de dessincronização forçada o qual promove a
dissociação do ritmo de atividade locomotora em ratos através de alterações no
período do ciclo de CE. Conseguimos observar a dissociação em cerca de 90% dos
animais submetidos ao protocolo. A hipótese do trabalho foi de que os animais
submetidos a este protocolo apresentassem alterações cíclicas de comportamento
alternando as fases de coincidência (CAP) – a qual supostamente os animais
estariam num estado semelhante à mania – com as fases de não coincidência
(NCAP), fase na qual o animal supostamente estaria num estado semelhante à
depressão, pois mesmo estando na fase escura não apresenta a atividade
locomotora correspondente. Alternando estes diferentes momentos, um mesmo
animal poderia mimetizar a ciclagem que é espontânea entre a mania e depressão
observada nos indivíduos com transtorno bipolar.
Em trabalhos realizados anteriormente com o protocolo de dessincronização
forçada, é facilmente observável que na fase de CAP os animais apresentam a
atividade locomotora aumentada e mais concentrada (Campuzano e cols., 1998;
Cambras e cols., 2004; de la Iglesia e cols., 2004; Cambras e cols., 2007). Somado
aos achados encontrados por Neto e cols. (2006), vimos que os animais se
comportam de modo diferente dependendo da fase em que se encontram neste
protocolo. Estas alterações se repetem a cada 7 dias aproximadamente
(dependendo do período endógeno do animal). As fases de CAP e NCAP se
revezam alternadamente no mesmo animal (por isso é considerado um fenômeno
cíclico) e podem representar oscilações no humor, levando-nos a sugerir este como
modelo animal para o transtorno bipolar. No entanto, para a validação de uma
doença humana em um modelo animal devem ser contemplados três critérios: a
validação de face, validação preditiva e validação do construto, como explicado
anteriormente na introdução desta tese.
63
Começando pela validação de face (ou aparente), esta consiste na
semelhança entre os sintomas observados em humanos e o fenótipo dos animais
tidos como modelo. No nosso caso, um modelo animal ideal para estudo do
transtorno bipolar incluiria comportamentos espontâneos e progressivos que
oscilariam entre a manifestação de aumento e redução de um determinado
comportamento, que poderiam similar o comportamento humano apresentado nas
fases de mania ou depressão, contemplando então a validade aparente (Gavioli e
cols., 2009).
A primeira evidência comportamental que encontramos no modelo foi na
análise da atividade locomotora dos animais, a qual revelou que os animais
dissociados apresentam padrões de atividade locomotora diferente dos ratos
mantidos em ciclos de claro/escuro simétricos de 24h (aqui chamados de grupo
controle) e esses padrões se repetem de forma cíclica. Além disso, os animais em
CAP apresentaram atividade locomotora muito mais elevada do que os animais
controle com o mesmo perfil de distribuição ao longo do período (Figuras 9 e 10).
Esse dado mostra a hiperlocomoção dos animais em CAP. Essa é uma das
características mais observadas nos modelos de mania: a hiperatividade locomotora,
tanto nos modelos induzidos por psicoestimulantes (Jornada e cols., 2009; Herman e
cols., 2007; Morris e cols., 2006; Frey e cols., 2006; Scotti e cols., 2011) quanto nos
modelos genéticos (Young e cols., 2010; Roybal e cols., 2007).
Nos protocolos de dessincronização forçada em humanos (Czeisler e cols.,
1999; Waterhouse e cols., 2004), não há nada referente à quantificação da atividade
locomotora, há somente a caracterização dos ritmos e aferição do período
endógeno. O mesmo ocorre com os estudos de dessincronização forçada em ratos,
há a caracterização dos ritmos, em diferentes situações, mas nenhum quantifica a
atividade para podermos comparar (Campuzano e cols., 1998; Cambras e cols.,
2004; Cambras e cols., 2007; Schwartz e cols., 2009).
Quando analisamos a atividade locomotora das diferentes fases do ciclo
claro/escuro em separado, encontramos semelhança entre os animais durante os
dias em CAP e os controles na fase clara: baixa atividade, como era de se esperar
64
destes animais noturnos. Contudo, podemos observar que o aumento da atividade
observada em CAP está toda concentrada na fase escura, a fase ativa do animal.
Ou seja, há hiperatividade locomotora quando os animais estão na fase de CAP. Na
fase escura, os animais em CAP ficam mais ativos do que os animais controle.
Alternadamente, quando os ratos estão em NCAP apresentam um valor
intermediário de locomoção considerando a média diária dos grupos. Todavia, a
atividade dos animais em NCAP é igualmente distribuída ao longo das 22h e quando
deveria apresentar alta atividade (na fase escura) a atividade mostra-se reduzida e
bastante fragmentada, revelando que a fase ativa não está mais tão bem definida e
uma aparente mudança de fase deste ritmo. É importante ressaltar que os testes
comportamentais foram realizados durante a fase escura. Com isso, comparamos o
desempenho dos animais nas tarefas durante a sua fase mais ativa. Tanto os
controles quanto os animais em CAP, certamente estavam no meio de sua noite
subjetiva durante os testes. Entretanto, os animais em NCAP no meio da fase
escura estão no meio da noite objetiva, mas seu relógio endógeno provavelmente
não aponta para a mesma fase, não estão necessariamente na noite subjetiva, e é
por esse motivo que este modelo serve para demonstrar nos animais o que pode
acontecer com os humanos que acabam por dessincronizar seus ritmos biológicos e
apresentar maior propensão para desenvolver transtornos.
A avaliação da atividade motora e o comportamento exploratório nos roedores
é um dos métodos mais largamente utilizados para determinar os efeitos
comportamentais das manipulações genéticas, farmacológicas e fisiológicas. Entre
os estudos sobre as características do transtorno do humor, muitos autores
concordam com a hiperlocomoção nos pacientes em fase de mania (Angst e cols.,
2003; Benazzi e cols., 2007; Teicher, 1995; Young e cols., 2010). A hiperatividade
locomotora dos animais em CAP foi observada apenas nos dados de registro da
atividade em suas homecages, pois quando testados no campo aberto estes animais
apresentaram atividade exploratória reduzida em relação aos animais controle.
Assim como os animais em NCAP que também apresentaram redução na atividade
exploratória durante o teste do campo aberto. Essa redução é referente tanto à
exploração no centro do aparato quanto na periferia do mesmo. Não podemos
65
afirmar que os animais dissociados, tanto em CAP quanto em NCAP, apresentaram
aumento da ansiedade neste teste, por mais que tenham explorado menos o centro
do CA, pois a atividade exploratória total é reduzida.
Alguns estudos em camundongos relatam que é bastante frequente os
animais apresentarem pouca atividade ou até pequenos momentos de freezing
imediatamente quando expostos a um novo ambiente, como numa situação de teste
(Tang e Sanford, 2005). Nem sempre a atividade exploratória ou espontânea
apresentada no campo aberto reflete a atividade locomotora observada na
homecage, talvez pela novidade ou talvez pela manipulação prévia dos animais.
Tang e colaboradores (2002) verificaram que camundongos comportam-se
diferentemente quando testados pela primeira vez no campo aberto ou num
ambiente onde o animal já fora previamente habituado. No final, sugerem aos
pesquisadores
cautela
em
analisar
a
atividade
locomotora
em
testes
comportamentais, pois os resultados destes nem sempre correspondem à atividade
do animal e sim ao desempenho no teste. Ao manipular os animais dissociados,
pude perceber que eles ficam bastante ariscos e assustados, muitos responsivos ao
toque.
Resultados prévios do nosso laboratório (Neto, 2006) com os animais
dissociados mostraram que estes exploram mais o campo aberto do que os animais
controle, dado este oposto ao encontrado neste estudo, porém além de serem
animais de outra procedência (vindos do Biotério do Centro de Ciências Biológicas
da Universidade Federal de Pernambuco – Recife), estes possuíam idade mais
avançada do que os utilizados neste estudo (4-7 meses x 2-3 meses,
respectivamente). A atividade exploratória dos animais controle no nosso
experimento teve em média 77 quadrantes de deslocamento num campo aberto
circular, enquanto o deslocamento médio dos animais controle do estudo anterior foi
de 28±21 quadrantes num aparato quadrado. Portanto, fica difícil a comparação dos
resultados.
Já no teste do LCE, como todos os grupos exploraram igualmente o labirinto,
podemos afirmar que no momento de NCAP os animais apresentam ansiedade
66
aumentada, mensurada pelo tempo reduzido em que permaneceram no braço
aversivo do aparato (braço aberto). Ratos possuem uma propriedade chamada
tigmotaxia, a qual faz com que estes animais prefiram estar em cantos ou ambientes
fechados e escuros, pois nestes ambientes o animal sente-se mais seguro devido ao
menor risco de ser predado. Essa propriedade nos permite inferir que quanto maior
a exposição em ambientes aversivos, menor a ansiedade do animal, e por outro
lado, quanto menor exposição em situações aversivas, maior a ansiedade do animal.
Neste caso o menor tempo de exposição do grupo NCAP no braço aversivo nos
permite afirmar que estes animais apresentam aumento na ansiedade.
Observando o comportamento dos animais durante os testes, também
observamos que os animais submetidos ao protocolo de dessincronização forçada
apresentam
comportamentos
estereotipados.
Os
animais
em
dissociação
apresentaram maior quantidade de grooming, em ambos os momentos CAP e
NCAP, sugerindo certo grau de ansiedade durante o teste do CA para ambos
(Figura 15). O aumento do grooming também foi observado em outro modelo de
mania, os animais knockout para o gene GluR6 (Shatiel e cols., 2008). Os animais
dissociados também apresentaram menor quantidade de rearing durante este
mesmo teste (Figura 16), o que revela menor exploração vertical do aparato pelos
animais dissociados.
No teste do LCE, os ratos dissociados (tanto em CAP quanto em NCAP)
apresentam menor avaliação de risco. Este dado foi aferido pelo comportamento de
head down, o qual o animal permanece no centro do labirinto e estica o corpo a fim
de avaliar se o braço a ser explorado é seguro. Os indivíduos bipolares,
principalmente em fase de mania, apresentam avaliação do risco reduzida, realizam
ações impulsivas, sem julgamento prévio, pois nesta fase o julgamento mostra-se
prejudicado (para revisão, ver Einat, 2007).
Os indivíduos depressivos apresentam uma característica chamada anedonia,
que reflete a perda da capacidade de sentir prazer. Os testes comportamentais mais
utilizados para avaliar essa característica em animais são: o nado forçado e o teste
de consumo de glicose. Não conseguimos caracterizar anedonia nos ratos na
67
condição de NCAP, nem no nado forçado nem no teste de consumo de glicose.
Entretanto, a partir do teste do nado obtivemos a informação de que os animais em
CAP apresentam um perfil do tipo antidepressivo menos depressivos, pois
apresentam a latência para o comportamento desesperado bastante aumentada, o
que reflete a luta deles pela sobrevivência, característica oposta à encontrada nos
indivíduos depressivos.
Quanto ao teste de consumo de glicose, o resultado encontrado nos animais
em NCAP pode ser explicado pelo fenômeno de binge, no qual indivíduos com
transtornos ou privados de sono aumentam significativamente o consumo alimentar
em certo período de tempo, principalmente alimentos ricos em gordura ou altamente
palatáveis (Avena e Bocarsly, 2012). Visto que os animais em NCAP possuem o
padrão do ciclo de atividade/repouso bastante conturbado, o binge poderia ser uma
explicação plausível para o aumento no consumo de glicose. Outra explicação
poderia advir do sistema de recompensa. Há um aumento no uso de substâncias
ilícitas nos pacientes com transtorno bipolar, principalmente quando o diagnóstico é
tardio, numa tentativa de aliviar os sintomas (Kolodziej e cols, 2005; Levander e cols,
2007; Mitchell e cols, 2007; Gao e cols, 2010).
Cerca de 70% dos indivíduos bipolares tiveram uso/abuso de drogas no
histórico (Gao e cols, 2008), sendo álcool e maconha os mais frequentes, seguidos
de cocaína e opióides (Cerullo e cols, 2007). A história pregressa recente ou distante
do uso de substâncias nos pacientes com TB de ciclagem rápida aumenta
significantemente o risco para o desenvolvimento de outras comorbidades, como
transtornos de ansiedade e pânico (Gao e cols, 2008). Os efeitos ansiolíticos do
álcool em curto prazo (possivelmente mediados por mecanismos GABAérgicos)
podem agir conjuntamente com os efeitos ansiogênicos do álcool no sentido de
gerar um ciclo vicioso oscilando entre o uso do álcool e os sintomas de ansiedade.
Esse ciclo vicioso aumenta o risco de desenvolver comorbidades, já que é um
processo interativo, independente de qual foi a comorbidade que se iniciou primeiro
(Kushner e cols, 1999).
68
O teste de consumo de glicose deste trabalho foi baseado no protocolo
descrito por Martynhak e colaboradores (2011). Talvez não tenha sido a melhor
metodologia escolhida, pois em outros protocolos, por exemplo, no descrito por
Dzirasa e colaboradores (2010), os animais primeiramente aprendem a consumir a
solução com sacarose (e não glicose), ficam privados de água por 18 horas antes do
teste e então é disponibilizado para o animal somente a solução de sacarose – sem
a garrafa contendo água – para não causar competição pelo consumo. Por outro
lado, disponibilizando em conjunto a garrafa contendo água damos opção de
escolha para o animal e conseguimos avaliar a preferência e não somente a
quantidade de consumo.
Em resumo dos dados comportamentais temos que os animais em
dessincronização forçada mostram-se mais estressados, dados pelas observações
do grooming, além da atividade exploratória reduzida e avaliação de risco
prejudicada. Os animais no momento CAP apresentam perfil do tipo antidepressivo
pelo teste NF e apresentam maior atividade locomotora concentrada na fase escura.
Enquanto os animais em NCAP apresentam maior ansiedade, avaliado no LCE.
Tomando em conjunto os dados descritos até agora, não podemos afirmar
que estes ratos apresentam comportamento semelhante aos animais controle não
dessincronizados. Parecem estar numa condição de estado afetivo alterado,
sugerindo que o humor destes animais oscila quando comparado nestas diferentes
fases ao grupo controle. Como conseguimos caracterizar somente a mania de forma
cíclica, há a possibilidade de estarmos fazendo terapia de avanço de fase, já que
avançamos a luz todos os dias em duas horas. Em outras palavras, é possível que
não demos chance da depressão se estabelecer, já que avançando o ritmo da luz,
arrastamos a fase dos animais e impedimos que eles expressem o comportamento
depressivo. Além do que a privação de sono aguda tem efeito antidepressivo (WirzJustice e van der Hoofdakker, 1999), o que corrobora com nossos achados.
E
ainda, talvez não tenhamos utilizado um teste sensível o suficiente para a detecção
do comportamento depressivo. Dessa forma, contemplamos a validação aparente de
69
forma parcial já que caracterizamos o comportamento semelhante à mania, mas não
o comportamento depressivo nestes animais.
Um modelo de mania que vem sendo bastante discutido na literatura é o
camundongo mutante ou knockdown ClockΔ19 que apresenta os três critérios de
validação requeridos. Sabe-se que o gene Clock é um dos principais genes
envolvidos na circuitaria molecular circadiana e está envolvido no sistema de
recompensa do sistema límbico. Os animais mutantes ClockΔ19 apresentam o
comportamento
menos
depressivo,
redução
na
ansiedade,
aumento
no
comportamento de risco, hiperatividade, aumento da preferência por sacarose e
cocaína e aumento na taxa de autoestimulação intracraniana. Com isso, contempla
a validação de face para tornar-se um modelo de mania (McClung e cols., 2005;
Roybal e cols., 2007).
A vantagem do nosso modelo sobre o knockout ClockΔ19 é que o modelo de
dessincronização forçada produz uma ciclagem previsível no comportamento dos
animais diferentemente do knockout ClockΔ19, cujo modelo não apresenta nenhuma
evidência de que estes ciclem entre a mania e a depressão ou mesmo haja
oscilação no humor (Roybal e cols., 2007). Por outro lado, os animais mutantes
ClockΔ19 contemplam os outros critérios de validação, como a validação preditiva, a
qual estabelece que os fármacos utilizados para reverter os sintomas em humanos
devem ter o mesmo efeito sobre os modelos animais.
Tem sido demonstrado que o lítio – principal fármaco adotado para o
tratamento dos pacientes com o transtorno – aumenta o período circadiano em
diversas espécies, incluindo cultura celular, plantas, invertebrados e vertebrados
(para revisão ver Li e cols., 2012). O lítio tem ação inibitória conhecida sobre a
enzima GSK3β (Gould e cols., 2004), cuja estabilidade da proteína é dependente de
outras moléculas do relógio molecular circadiano. Para dar maior detalhamento, a
estabilidade da GSK3β depende da molécula rev-erb que por sua vez mantêm a
proteína BMAL1 (que dimeriza com o Clock) num estado reprimido. A inabilidade de
degradação da rev-erb é suficiente para prevenir o início da oscilação dos genes
relógio. Um (ou mais componente) do soro que mimetiza um zeitgeber induz a
70
fosforilação e inativação da GSK3β, resultando na degradação da rev-erb e iniciando
o ciclo de expressão gênica dos genes relógio (Yin e cols, 2006). Além disso, o
tratamento com lítio aumenta a expressão de neurotrofinas, como BDNF (brain
derived neurotrophic fator) e o fator de crescimento neural entre outros, além de ser
parte da via de ativação da enzima AKT, que exerce um importante papel nos
comportamentos relacionados ao humor e à recompensa (Krishnan e cols., 2008).
O tratamento com lítio é capaz de reverter quase todas as anormalidades
observadas nos animais ClockΔ19, incluindo os comportamentos relacionados à
ansiedade e depressão (Coque e cols., 2011).
Ocorreram alguns contratempos no experimento do tratamento com o lítio
deste trabalho. Os animais ficaram muito debilitados ao final do experimento,
portanto temos que levar em consideração as alterações comportamentais
observadas. Em conversa informal com alguns pesquisadores que utilizaram o lítio,
foi observada a mesma reação em ratos e camundongos. Alguns deles modificaram
o fármaco e passaram a administrar valproato, outros reduziram a concentração de
dose terapêutica para uma dose subterapêutica, na qual a litemia gira em torno de
0,1-0,5 mEq/ml e os animais permanecem saudáveis até o final do tratamento
crônico de 21 a 28 dias (Coque e cols., 2011; relatos pessoais).
Diferentes resultados podem ser encontrados variando a concentração do
lítio. Tratamentos que produzem níveis séricos por volta de 1,3 -1,4 mmol/L reduzem
o tempo de imobilidade de animais no nado forçado (Bersudsky e cols., 2007)
diferentemente de quando a concentração sérica é menor: à concentração de
0,8mmol/L não há efeito no nado forçado. Com níveis séricos de 0,4 mmol/L os
animais apresentam redução na atividade locomotora, mas nenhuma alteração nos
testes comportamentais foi relatada nos testes com os animais ClockΔ19 (Roybal e
cols., 2007). Nestes animais, o tratamento crônico com lítio por 10 dias fez com que
os animais apresentassem novamente os níveis basais de ansiedade, além de
apresentarem o mesmo tempo de imobilidade no nado forçado. Entretanto a mesma
redução na atividade locomotora foi observada tanto para os animais mutantes
quanto para os animais controle, sugerindo que o lítio reduz a atividade locomotora
dos animais mesmo em doses subterapêuticas (Roybal e cols., 2007).
71
No nosso experimento, pudemos observar o efeito do lítio na ritmicidade dos
animais logo nos primeiros dias de tratamento. Os animais perdem o componente
rítmico sincronizado ao ciclo claro/escuro externo de 22h e passam a apresentar
somente um componente rítmico de atividade locomotora com o período alongado.
Portanto, sabemos que o lítio muda o período do ritmo da atividade locomotora dos
animais mesmo sob o ciclo claro/escuro do protocolo de dessincronização forçada
(T22) continuamente, mas através do nosso experimento não foi possível afirmar se
a atividade locomotora volta aos níveis basais do controle pelo estado de saúde dos
animais. Talvez se tentássemos utilizar uma dose subterapêutica ou outro
medicamento, a quantidade de atividade dos animais experimentais não ficaria tão
reduzida e fosse mais fiel à comparação.
Por fim, não conseguimos contemplar a validação preditiva de forma
satisfatória. Seriam necessários novos experimentos. O que podemos afirmar deste
experimento farmacológico é que o lítio efetivamente alterou a ritmicidade circadiana
dos animais dissociados, fazendo com que eles realmente abolissem a dissociação
e alongassem o período de atividade locomotora.
Seguindo para o próximo critério de validação, discutiremos a seguir sobre a
validação do construto, a qual busca semelhanças nas bases biológicas
desencadeadoras da doença, ou seja, na etiologia do transtorno.
A fim de avaliar como a neurotransmissão poderia estar alterada nestes
animais, buscamos alterações no sistema serotoninérgico. Um dos mecanismos
pelos quais a luz pode influenciar o humor é através da habilidade de induzir
rapidamente o aumento da liberação de serotonina. A taxa de produção da
serotonina é maior nos dias longos do que nos dias curtos e aumenta conforme a
intensidade luminosa (Lambert e cols, 2002). Isso pode explicar a depressão
sazonal. Além disso, a serotonina parece estar envolvida em mudanças de fase
fóticas e não fóticas (Mitlsberger e cols, 2000), pois os NSQs têm projeções diretas
para os núcleos da rafe (Kawano e cols, 1996) e, ao mesmo tempo, estimulações
elétricas na rafe acabam aumentando a liberação de serotonina nos NSQs,
causando avanço de fase nos ritmos circadianos (Yamaka e cols, 2010).
72
Em modelos animais de ansiedade e depressão, a serotonina é um
neurotransmissor
chave
que
contribui
nas
alterações
de
comportamento
relacionadas ao estresse (Graeff e cols., 1996; Lucki, 1998; Maier and Watkins,
2005). Em adição, o sistema sertoninérgico é um importante alvo farmacológico no
tratamento de doenças afetivas (Blier and de Montigny, 1999). Entretanto, a relação
entre este sistema e o comportamento ansioso e estressado ainda não é clara
(Cooper e cols, 2008).
A investigação dos receptores serotoninérgicos presentes neste trabalho fora
realizada em doutorado sanduíche no laboratório de ritmos circadianos do professor
Horacio de la Iglesia na Universidade de Washington. Foram realizados os
procedimentos de imunohistoquímica para o receptor 5HT1A e a hibridização in situ
para o receptor 5HT2A.
As análises da hibridização in situ para o receptor 5HT2A não são
animadoras, uma vez que não foi possível identificar nenhuma diferença entre os
grupos na expressão de RNAm dos receptores nas áreas escolhidas. Talvez
possamos ter perdido informação pela marcação ter ficado fraca e alguma diferença
sutil entre a densidade desses receptores infelizmente não tenha sido detectada.
Todavia, obtivemos resultados bastante interessantes para o receptor
serotoninérgico 5HT1A. Através da técnica de imunohistoquímica para marcação
desse receptor verificamos que não há variação circadiana, ou seja, há o mesmo
número de células marcadas para este receptor tanto na fase clara quanto na fase
escura nos animais controle.
Ao comparar os animais dissociados com os animais controle, verificamos
que os animais dissociados apresentam maior marcação na região da amígdala.
Sabemos que há uma maior ativação do metabolismo da amígdala nos indivíduos
bipolares (Drevets e cols, 2002). Uma das vias para ativação da amígdala é pelo
córtex pré-frontal (CPF), pois é sabido que pacientes com alterações na interação
CPF-amígdala geralmente apresentam uma inabilidade de regular respostas afetivas
73
(Rozenkranz e cols, 2003). Entretanto, não encontramos diferença no CPF entre os
grupos, sugerindo que não há alterações nessa via.
Por outro lado, a condição imposta pelo protocolo de dessincronização
forçada pode estar servindo de estímulo aversivo por causar o rompimento da
ritmicidade circadiana, e, dessa forma, estar ativando os núcleos dorsais da rafe
que, através de suas projeções para a amígdala, causam uma exacerbação
serotoninérgica favorecendo a liberação de serotonina na amígdala (Deakin e
Graeff, 1991). Esse aumento de serotonina na amígdala pode estar causando o
aumento da ansiedade encontrada nos animais em NCAP. O maior número de
células marcadas com receptores 5HT1A na amígdala dos animais em NCAP pode
ser resultante de uma tentativa de restaurar a inibição da amígdala e reduzir esse
potencial efeito ansiogênico, pois a ativação dos receptores 5HT1A na amígdala tem
sabidamente um efeito ansiolítico (de Andrade Strauss e cols, 2013). Nessa situação
de hiperativação serotoninérgica temos a hiperativação dos receptores 5HT1A, que
são inibitórios (pois estão acoplados a proteína Gi) e causam redução na ativação da
amígdala (Drevets e cols, 2007).
Ao mesmo tempo, projeções da rafe para outras áreas como a substância
cinzenta periarquedutal causa inibição de comportamentos relacionados ao pânico.
Em situações de perigo potencial, onde não se tem um perigo real como na
ansiedade, teria a ativação dessas duas vias e o animal apresentaria o
comportamento de esquiva e não de fuga. O que teria até uma relevância evolutiva,
pois numa situação de estresse seria mais vantajoso para o animal se esconder do
que ter um “ataque de pânico” e optar pela fuga, opção na qual se tornaria uma
presa mais fácil. A serotonina teria um papel modulatório na amígdala, promovendo
esse efeito ansiogênico (Paul e cols, 2013).
No modelo de depressão induzida através da separação materna também foi
encontrado aumento da expressão de 5HT1A na amígdala, assim como do
transportador de serotonina, o que corrobora com nossos dados (Vicentic e cols,
2006). Essas alterações na neurotransmissão serotoninérgica permanecem na vida
74
adulta destes animais, o que pode implicar numa maior suscetibilidade a transtornos
afetivos no futuro (Vicentic e cols, 2006).
Em humanos com transtorno bipolar há redução de cerca de 41% do receptor
5HT1A na rafe (Drevets e cols, 1997). Essa redução pode refletir numa menor
inibição da rafe em produzir serotonina, fazendo com que estes indivíduos tenham
uma maior transmissão serotoninérgica influenciando tanto na quantidade quanto na
temporalidade da secreção de serotonina em outras áreas do cérebro, o que pode
justificar o nosso achado de maior ativação do receptor 5HT1A na amígdala numa
tentativa de reduzir a ativação na amígdala nos animais dissociados.
Infelizmente não foi possível realizar a contagem das células nos núcleos da
rafe, pois como tínhamos somente um hemisfério cerebral quando chegávamos na
altura da rafe já não tínhamos mais estruturas neuroanatômicas o suficiente para
verificarmos a correta posição da rafe e realizarmos os cortes do mesmo núcleo
consistentemente. Por este motivo os cortes ficaram em alturas diferentes e não foi
possível a identificação dos núcleos da rafe, muito menos a contagem das células
neste local, apesar de as imunohistoquímicas terem sido feitas nesses cortes.
Os resultados da transmissão serotoninérgica também podem estar
relacionados com a dissociação pelo fato de que alterações nesse sistema podem
causar não somente mudanças no período circadiano, mas também na alocação da
atividade locomotora ao longo do dia. Dessa forma, disfunções nesse sistema
podem resultar em mudanças no padrão de atividade comportamental (Paulus e col,
2012).
Neste
caso,
as
alterações
rítmicas
causadas
pelo
protocolo
de
dessincronização que estariam gerando essas alterações serotoninérgicas nos
animais.
Os receptores serotoninérgicos 5HT1A, 5HT2A e 5HT2C parecem ter um
papel proeminente na modulação dopaminérgica no córtex pré-frontal, com o 5HT1A
e 5HT2A estimulando a liberação de dopamina, enquanto o 5HT2C a inibe, talvez
por ações na VTA ou interneurônios GABAérgicos (Alex e cols, 2007). A ativação
dos receptores 5HT1A densamente encontrados nos neurônios do córtex pré-frontal
75
aumenta a atividade dos neurônios dopaminérgicos da VTA e, consequentemente, a
liberação mesocortical de dopamina (Diaz-Mataix e cols, 2005).
Os estudos a partir do mutante ClockΔ19 mostram que a regulação do
sistema dopaminérgico da área tegmental ventral (VTA) pela CLOCK pode ser
devido a essa proteína funcionar como um fator de transcrição. Nos animais
mutantes muitos dos genes que são expressos na VTA para o controle da atividade
dopaminérgica estão com a expressão alterada, incluindo a expressão da enzima
tirosina hidroxilase – passo limitante para a síntese da dopamina – que tem
expressão circadiana na VTA e pode estar sob controle dos genes relógio (Weber e
cols., 2004). Essa atividade dopaminérgica alterada pode ser vista nos trabalhos de
Dzirasa e cols. (2011), onde foi realizado o registro eletrofisiológico de áreas
límbicas como o núcleo accumbens, a VTA e amígdala. O animal ClockΔ19
apresenta um déficit na sincronização dos disparos neuronais com a atividade
oscilatória do núcleo accumbens. Essa “disfunção” no núcleo accumbens pode estar
contribuindo para os animais apresentarem sintomas semelhantes à mania como
resultado dos mecanismos neurofisiológicos gerados pelo funcionamento anormal
do mecanismo molecular circadiano (Dzirasa e cols., 2010; Dzirasa e cols, 2011),
reforçando a hipótese apresentada nesta discussão. Em conjunto, não é somente o
aumento ou decréscimo da taxa de disparos neuronais entre as estruturas, mas os
padrões de disparo, de oscilações e a coerência entre as estruturas que ditam os
comportamentos e o humor resultantes (Coque e cols., 2011). Esses resultados
fazem o link entre os comportamentos de ansiedade dos animais mutantes com a
disfunção oscilatória do núcleo accumbens, e sugerem que essas alterações
oscilatórias induzidas pelo polimorfismo do gene Clock podem contribuir com as
manifestações relacionadas à ansiedade observadas na mania. Conjuntamente,
esses achados corroboram com a hipótese de que o sistema dopaminérgico e o
sistema de temporização circadiano estão intrinsicamente interligados (McClung e
cols, 2005).
O funcionamento normal do cérebro depende crucialmente do ajuste e
coordenação temporal apropriado. Essa fina regulação parece envolver as
76
frequências oscilatórias assim como o delicado balanço entre a sincronização e a
dessincronização da interação dos grupamentos neuronais (Uhlhaas e Singer,
2006).
Olhando por este ponto de vista, podemos considerar que no nosso modelo
ocorre o desacoplamento entre as regiões ventrolateral e dorsomedial dos núcleos
supraquiasmáticos (de la Iglesia e cols., 2004). Causado pela dessincronização
forçada, esse desacoplamento gera diferentes outputs: a região ventrolateral recebe
projeções diretas da retina e dita o ritmo conforme o ciclo claro/escuro ambiental
imposto de 22h; ao mesmo tempo, a região dorsomedial, que está sob coordenação
relativa em relação a região ventrolateral, gera o ritmo semelhante ao livre curso (de
la Iglesia e cols., 2004). Esse fenômeno é possível devido a anatomia dos NSQs,
uma vez que as gap junctions desse núcleo parecem acoplar os neurônios somente
dentro de cada subdivisão, mas não entre elas (Colwell, 2000).
Aparentemente o funcionamento ótimo do sistema circadiano é resultante da
integração da informação entre os diferentes osciladores do sistema. A integração
entre ritmos com diferentes períodos deve acontecer pela interação entre os
osciladores, ou seja, deve haver acoplamento entre os osciladores para que seja
gerado um ritmo. Chamamos de acoplamento a capacidade de um oscilador com
período e fase próprios de ajustar-se aos outros osciladores com os quais interage.
O sucesso e/ou estabilidade desse acoplamento depende do período característico
de cada um dos osciladores e da força do acoplamento. Quando a força de
acoplamento é relativamente fraca, os deslocamentos na relação de fase entre
ambos os osciladores manifestam-se como mudanças no período do sistema
acoplado, o qual é conhecido como coordenação relativa (Golombek e AguilarRoblero, 2003). A diferença entre acoplamento e sincronização consiste em que no
último caso um ciclo ambiental age de forma unidirecional com um oscilador
biológico, enquanto no acoplamento, um oscilador biológico age de forma recíproca
com outro oscilador do mesmo organismo. Apesar de uma possível semelhança
entre os mecanismos, os substratos morfológicos podem ser completamente
diferentes nos dois processos.
77
É válido considerar o fenômeno de batimento descrito por Granada e cols.
(2011), o qual sugere que um único oscilador pode gerar mais de um ritmo, ou seja,
um oscilador pode ter diferentes outputs. O fenômeno do batimento se dá pelo
aparecimento de uma modulação mais lenta na amplitude da saída do oscilador. Os
autores realizaram uma simulação computacional em conjunto com a análise de
dados experimentais sobre a dessincronização forçada em T22 e sugeriram uma
superposição linear de mais de um sinal de saída de um único oscilador, que não
possuem uma relação de fase e sim um padrão, um período determinado, que
acabam por se sobrepor e gerar os componentes rítmicos observados na
dessincronização forçada. Na simulação feita por Granada e cols. (2011), além dos
dois componentes encontrados por nós, foi descrito um componente extra com
período próximo de 28h.
Refinetti (2006) em seu livro “Circadian Phisiology” critica a existência de mais
de um oscilador central. Segundo o autor, os mamíferos apresentam um único
oscilador central, pois há vários anos tentam explicar a dessincronização interna dos
ritmos circadianos e só conseguem mostrar ou a dessincronização do ritmo do ciclo
sono/vigília ou a dessincronização de outras variáveis fisiológicas (como o ritmo da
temperatura corporal), mas nunca todos os fenômenos em conjunto. Além de dizer
que estes estudos nunca são replicáveis.
Realizamos um experimento em paralelo avaliando a temperatura central dos
animais em dessincronização forçada, o qual gerou um artigo ainda não publicado,
mas coloquei nesta tese como anexo V para quem tiver curiosidade em lê-lo. Neste
trabalho fizemos a simulação computacional do modelo levando em conta a variável
temperatura central. Nosso modelo reproduziu a mesma variação aumentada da
amplitude de saída da região dorsomedial dos NSQs, levando-nos a supor que esta
alta variabilidade da amplitude na saída deve gerar instabilidade na fase dos ritmos
circadianos.
Como diz McClung (2011) “it’s all about timing”! Enquanto o sistema de
temporização circadiano é internamente controlado, ele é fortemente influenciado
pelo ambiente (ou por pistas ambientais). Portanto, alterações no ciclo claro/escuro,
78
disponibilidade de nutrientes e exposição ao estresse certamente vão ter impacto na
ritmicidade biológica. Os circuitos de recompensa e do humor no cérebro tem seu
próprio ritmo e são sincronizados pelo NSQ, mas podem ser dessincronizados do
NSQ em certas condições. Clock genes têm um importante papel no circuito
relacionado ao humor, não somente em regular a atividade circadiana, mas também
na sincronização ultradiana dos disparos neuronais dentro e entre estruturas
cerebrais. Essa sincronização entre várias estruturas cerebrais, peptídeos e
hormônios no organismo, juntamente com o ambiente são cruciais para manutenção
do humor saudável.
Adicionalmente às explicações fisiológicas, sob um olhar mais antroposófico
essa “dessincronização” nos humanos muitas vezes é causada porque a atividade
humana tende ao futuro, planejamos o futuro esperando que ele se torne o presente
perdendo a orientação de futuro normal (“vou fazer isso depois de: me formar, de
ganhar dinheiro, de casar..."), e criando uma “tensão apetitiva”, uma perspectiva
iminente.
A temporalidade é um reino vazio que precisa ser constantemente
preenchido, o que é ignorado na medida em que nossas necessidades não vão
sendo atendidas, porque nunca está satisfeita com o momento seguinte como cada
momento por sua vez gera a potencialidade do outro, ainda por vir. Pacientes com
depressão severa tem pensamentos e desejos, mas esses pensamentos produzem
uma reação reversa e se sentem totalmente desencorajados de seguir adiante,
produzindo
uma
sensação
de
desilusão.
O
futuro
é
caracterizado
fenomenologicamente como abertura à mudança e movimento, sem essa abertura, o
futuro parece estático e determinístico, e o resultado pode muito bem ser a
desesperança, o desespero e até um sofrimento eterno por estar vivo. Por outro
lado, em fase de mania, os indivíduos tem a percepção da “eternidade” do momento
completamente inversa como achar que esse momento de grandiosidade sentida
durante a mania durará para sempre (Willie, 2005).
Para finalizar, um pensamento filosófico que resume o nosso modelo: “a
melancolia envolve um sofrimento derivado da incompatibilidade entre o tempo
79
subjetivo e o tempo objetivo, o qual é estranho a temporalidade do seu “eu”, e é visto
como um passo para trás, ou uma paralisação da temporalidade subjetiva, causando
uma dessincronização entre a percepção do tempo interno e externo originando
grande estresse psicológico e, quando em excesso, até transtornos psiquiátricos”
(Kupke, 2000).
80
6. CONCLUSÕES
O objetivo desta tese foi parcialmente cumprido, pois embora não haja uma
relação causal direta entre o modelo aqui apresentado e o transtorno bipolar,
encontramos um número considerável de evidências de oscilações previsíveis no
humor.
Os animais em dessincronização forçada apresentam características como
estresse, avaliação de risco prejudicada e redução na exploração vertical
independentemente
da
fase
em
que
estão.
Encontramos
alternância
de
comportamentos dos animais em CAP e NCAP, mostrando que a ansiedade
apresentada pelos animais em NCAP se alterna ciclicamente com a alta atividade
locomotora e comportamento antidepressivo encontradas nos animais em CAP.
Portanto, a validade aparente foi contemplada para o comportamento semelhante à
mania e não contemplada para o comportamento depressivo
Constatamos o efeito do lítio no alongamento do período apresentado pelos
animais dissociados, contudo não foi possível constatar se a atividade locomotora
dos animais pós-tratamento volta aos níveis basais. Não sendo possível contemplar
a validade preditiva.
A neuroquímica dos animais dissociados mostra-se alterada, apresentando
um maior número de receptores 5HT1A na região da amígdala, mostrando uma
possível redução na ativação da amígdala nestes animais. Dessa forma, podemos
afirmar que contemplamos uma parte da validade do construto.
Em suma, podemos concluir que o modelo de dessincronização forçada
contempla parcialmente os critérios de validação para um modelo de oscilações no
humor e traz novas informações para o estudo dos modelos animais deste
transtorno. Estudos mais aprofundados deste modelo podem levar a uma direção de
transtorno bipolar de ciclagem rápida.
81
7. REFERÊNCIAS BIBLIOGRÁFICAS
ABRATA – Associação Brasileira de Familiares, Amigos e Portadores de
Transtornos Afetivos. Acessível por www.abrata.org.br link: transtornos afetivos.
Acessado em setembro de 2009.
Akimova E, Lanzenberger R, Kasper S. The Serotonin-1A Receptor in Anxiety
Disorders. Biol Psychiatry. 2009; 66:627–635.
Alex KD e Pehek EA. Pharmacologic mechanisms of serotonergic regulation of
dopamine neurotransmission. Pharmacol Ther. 2007 February ; 113(2): 296–320.
Andrade LH, Wang Y-P, Andreoni S, Silveira CM, Alexandrino-Silva C, et al. (2012)
Mental Disorders in Megacities: Findings from the São Paulo Megacity.
Angst J, Gamma A, Benazzi F, Ajdacic V, Eich D, Rossler W. Diagnostic issues in
bipolar disorder. Eur Neuropsychopharmacol. 2003;13(suppl 2):S43-S50.
APA – American Psychiatric Association Press. Diagnostic and Statistical Manual of
Mental Disorders. 5th ed, Washington, DC, 2013.
Avena NM e Bocarsly ME. Dysregulation of brain reward systems in eating disorders:
Neurochemical information from animal models of binge eating, bulimia nervosa, and
anorexia nervosa. Neuropharmacology 63 (2012):87-96.
Baldessarini RJ. Current status of antidepressants: clinical pharmacology and
therapy. J Clin Psychiatry, 1989; 50(4): 117-126.
Ballone GJ. Serotonina. in PsiqWeb, Internet, disponível em www.psiqweb.med.br,
revisto em 2005.
Benazzi F. Testing new diagnostic criteria for hypomania. Ann Clin Psychiatry.
2007;19(2):99-104.
Benedetti, F., Serretti, A., Colombo, C., Barbini, B., Lorenzi, C., Campori, E.,
Smeraldi, E., 2003. Influence of CLOCK gene polymorphism on circadian mood
fluctuation and illness recurrence in bipolar depression. Am. J. Med. Genet. B
Neuropsychiatr. Genet. 123B, 23–26.
Bersudsky, Yuly; Shaldubina, Alona; Belmaker, R.H.Lithium's effect in forced-swim
test is blood level dependent but not dependent on weight loss. Behavioural
Pharmacology: February 2007 - Volume 18 - Issue 1 - pp 77-80.
Blier P e de Montigny C. Serotonin and Drug-Induced Therapeutic Responses in
Major
Depression,
Obsessive–Compulsive
and
Panic
Disorders.
Neuropsychopharmacology, Volume 21, Issue 2, Supplement 1, August 1999, Pages
91S-98S.
82
Cambras T, Chiesa J, Araujo J, Díez-Noguera A. Effects of photoperiod on rat motor
activity rhythm at the lower limit of entrainment. J Biol Rhythms. 2004 Jun; 19(3):21625.
Cambras T, Weller JR, Anglès-Pujoràs M, Lee ML, Christopher A, Díez-Noguera A,
Krueger JM, de la Iglesia HO. Circadian desynchronization of core body temperature
and sleep stages in the rat. Proc Natl Acad Sci U S A. 2007 May; 104(18):7634-9.
Campuzano A, Vilaplana J, Cambras T, Diez-Nogueira A. Dissociation of the Rat
Motor Activity Rhythm Under T Cycles Shorter Than 24 Hours. Physiology &
Behavior, Vol. 63, No. 2, pp. 171–176, 1998.
Carli M, Afkhami-Dastjerdian S and Reader TA. Effects of a Chronic Lithium
Treatment on Cortical Serotonin Uptake Sites and 5-HT1A Receptors.
Neurochemical Research, 1997; 22(4): 427-435.
Carobrez AP, Bertoglio LJ. Ethological and temporal analyses of anxiety-like
behavior: The elevated plus-maze model 20 years on. Neuroscience and
Biobehavioral Reviews 29 (2005) 1193–1205.
Carroll BJ, Cassidy F, Naftolowitz D, Tatham NE, Wilson WH, Iranmanesh A, Liu PY,
Veldhuis JD. (2007), Pathophysiology of hypercortisolism in depression. Acta
Psychiatrica Scandinavica, 115: 90–103.
Castagné V, Porsolt RD, Moser P. Use of latency to immobility improves detection of
antidepressant-like activity in the behavioral despair test in the mouse. European
Journal of Pharmacology 616 (2009) 128–133.
Cerullo MA e Strakowski MS. The prevalence and significance of substance use
disorders in bipolar type I and II disorder. Substance Abuse Treatment, Prevention,
and Policy 2007, 2:29.
Colwell, C.S. (2000). Rhythmic coupling among cells in the suprachiasmatic nucleus.
J. Neurobiol. 43, 379–388.
Cooper MA, McIntyre KE, Huhman KL. Activation of 5-HT1A autoreceptors in the
dorsal raphe nucleus reduces the behavioral consequences of social
defeat. Psychoneuroendocrinology, Volume 33, Issue 9, October 2008, Pages 12361247.
Coque L, Mukherjee S, Cao JL, Spencer S, Marvin M, Falcon E, Sidor MM, Birnbaum
SG, Graham A, Neve RL, Gordon E, Ozburn AR, Goldberg MS, Han MH, Cooper
DC, McClung CM. Specific Role of VTA Dopamine Neuronal Firing Rates and
Morphology in the Reversal of Anxiety-Related, but not Depression-Related Behavior
in the ClockD19 Mouse Model of Mania. Neuropsychopharmacology (2011) 36,
1478–1488.
Czeisler CA, Duffy JF, Shanahan TL, Brown EN, Mitchell JF, Rimmer DW, Ronda
JM, Silva EJ, Allan JS, Emens JS, Dijk DJ, Kronauer RE. Stability, precision, and
83
near-24-hour period of the human circadian pacemaker. Science. 1999 Jun
25;284(5423):2177-81.
de Andrade Strauss CV, Vicente MA, Zangrossi H. Activation of 5-HT1A receptors in
the rat basolateral amygdala induces both anxiolytic and antipanic-like effects.
Behavioural Brain Research 246 (2013) 103– 110.
de Andrade Strauss CV, Vicente MA, Zangrossi H. Activation of 5-HT1A receptors in
the rat basolateral amygdala induces both anxiolytic and antipanic-like effects.
Behavioural Brain Research 246 (2013) 103– 110.
de la Iglesia HO, Cambras T, Schwartz WJ, Díez-Noguera A. Forced
desynchronization of dual circadian oscillators within the rat suprachiasmatic
nucleus. Curr Biol. 2004 May 4;14(9):796-800.
de la Iglesia.(2007). In situ hybridization of Suprachiasmatic nucleus slices. Chapter
41. From: Methods in Molecular Biology, vol 362: Circadian Rhythms: Methods and
Protocols. Edited by: E. Rosato. Humana Press Inc., Totowa, NJ. Pp513-531.
De Sarno P, Li X, Jope RS (2002). Regulation of Akt and glycogen synthase kinase-3
beta phosphorylation by sodium valproate and lithium. Neuropharmacology 43:
1158–1164.
Deakin JFW, Graeff FG. 5-HT and mechanisms of defense. Journal of
Psychopharmacology 1991;5:305–15.
Díaz-Mataix L, Scorza MC, Bortolozzi A, Toth M, Celada P, Artigas F. Involvement of
5-HT1A Receptors in Prefrontal Cortex in the Modulation of Dopaminergic Activity:
Role in Atypical Antipsychotic Action. The Journal of Neuroscience, November 23,
2005. 25(47):10831–10843 • 10831.
Drevets WC, Price JL, Bardgett ME et al.Glucose metabolism in the amygdala in
depression: Relationship to diagnostic subtype and plasma cortisol levels.
Pharmacology, Biochemistry and Behavior 71 (2002) 431– 447.
Drevets WC, Price JL, Simpson JR et al. Subgenual prefrontal cortex abnormalities
in mood disorders. Letter to nature.(1997). Nature Vol 386(24): 826-7.
Drevets WC, Thase M, Moses E, Price J et al. Serotonin-1A Receptor Imaging in
Recurrent Depression: Replication and Literature Review. Nucl Med Biol. 2007
October; 34(7): 865–877.
Dubocovich ML, Markowska M. Functional MT1 and MT2 melatonin receptors in
mammals. Endocrine. 2005 Jul;27(2):101-10.
Dzirasa K, Coque L, Sidor MM, Kumar S, Dancy EA, Takahashi JS e cols.. (2010).
Lithium ameliorates nucleus accumbens phasesignaling dysfunction in a genetic
mouse model of mania. J Neurosci 30: 16314–16323.
84
Dzirasa K, McGarity DL, Bhattacharya A, Kumar S, Takahashi JS, Dunson D,
McClung CA, Nicolelis MAL. Impaired Limbic Gamma Oscillatory Synchrony during
Anxiety-Related Behavior in a Genetic Mouse Model of Bipolar Mania. The Journal of
Neuroscience, April 27, 2011 • 31(17):6449–6456 • 6449.
Ehlers CL, Frank E, Kupfer DJ. Social zeitgebers and biological rhythms. Archives of
General Psychiatry. 1988; 45, 948-952.
Einat H, 2007. Different behaviors and different strains: potential new ways to model
bipolar disorder. Neuroscience and Biobehavioral Reviews 31 (2007) 850–857.
Frank E, Swartz HA, Kupfer DJ. Interpersonal and social rhythm therapy: managing
the chaos of bipolar disorder. Biol Psychiatry. 2000 Sep 15; 48(6):593-604.
Frank RG e Koss C. Mental Health and Labor Markets Productivity Loss and
Restoration. Working Paper No. 38. February 2005. The Disease Control Priorities
Project is a joint effort of The World Bank, the Fogarty International Center of the
National Institutes of Health, the Bill & Melinda Gates Foundation, and the World
Health Organization
Frey BN, Andreazza AC, Rosa AR, Martins MR, Valvassori SS, Réus GZ, e cols..
Lithium increases nerve growth factor levels in the rat hippocampus in an animal
model of mania. Behavioural Pharmacology 2006;17:311–8.
Gao K, Chan PK, Verduin ML et al. Independent Predictors for Lifetime and Recent
Substance Use Disorders in Patients with Rapid Cycling Bipolar Disorder: Focus on
Anxiety Disorders. Am J Addict. 2010 ; 19(5): 440–449.
Gao K, Verduin ML, Kemp DE, et al. Clinical correlates of patients with rapid cycling
bipolar disorder and a recent history of substance use disorder: a subtype
comparison from baseline data of 2 randomized, placebo-controlled trials. J Clin
Psychiatry. 2008; 69:1057–1063.
Gavioli EC, Streck EL, Kapczinski F, Quevedo J. Modelos animais do transtono
Bipolar. In: Kapczinski F, Quevedo J. Transtorno Bipolar Teoria e Clínica. Porto
Alegre; Artmed, 2009; 44-60.
Golombek DA e Aguilar-Roblero R. Mecanismos de temporização em vertebrados.
In: Marques N e Menna-Barreto L. Cronobiologia: Princípios e Aplicações. 3 ed.
Editora da Universidade de São Paulo, 2003.
Golombek DA, Pévet P, Cardinali DP. Melatonin effects on behavior: possible
mediation by the central GABAergic system. Neurosci Biobehav Rev. 1996 Autumn;
20(3):403-12.
Gonçalves BSB, Carneiro BTS, Silva CA, Fernandes DAC, Fortes FS, Ribeiro JMG,
Cerqueira RC, Rolim SAM, Araújo JF. Mathematical model of the interaction between
the dorsal and ventral regions of the suprachiasmatic nucleus of rats. Sleep Sci.
2010;3(1): 40–44.
85
González MMC, de la Iglesia HO. 2008. X Taller Argentino de Neurociencias P51.
Goodwin FK e Jamison KR. Manic-Depressive Illness Bipolar Disorders and
Recurrent Depression. Second edition. New York: Oxford University Press. 2007. pp
463-653.
Gould TD, Chen D; Manji HK. In Vivo Evidence in the Brain for Lithium Inhibition of
Glycogen
Synthase
Kinase-3.
Neuropsychopharmacology (2004), 29, 32–38,
advance online publication, 27 August 2003; doi:10.1038/sj.npp.1300283.
Graeff FG, Guimarães FS, De Andrade TGCS, Deakin J. Role of 5-HT in stress,
anxiety, and depression. Pharmacology Biochemistry and Behavior, Volume 54,
Issue
1, May
1996, Pages
129-141
Granada AE, Cambras T, Dı´ez-Noguera A, Herzel H. Circadian desynchronization.
Interface Focus (2011) 1, 153–166.
Grandin LD, Alloy LB, Abramson LY. The social zeitgeber theory, circadian rhythms
and mood disorders: review and evaluation. Clin Psychol Rev. 2006 Oct; 26(6):67994.
Gruber J, Harvey AG, Wang PW, Brooks JO, Thase ME, Sachs GS, Ketter GA.
Sleep functioning in relation to mood, function, and quality of life at entry to the
Systematic Treatment Enhancement Program for Bipolar Disorder (STEP-BD).
Journal of Affective Disorders 114 (2009) 41–49.
Harada Y, Sakai M, Kurabayashi N, Hirota T, Fukada Y (2005) Ser-557phosphorylated mCRY2 is degraded upon synergistic phosphorylation by glycogen
synthase kinase-3 beta. J Biol Chem 280: 31714–31721.
Herman L, Hougland T, El-Mallakh RS. Mimicking human bipolar ion dysregulation
models mania in rats. Neuroscience & Biobehavioral Reviews 2007;31: 874–81
Johnsson A, Engelman W, Pflug B, Klemke W. Period lengthening of human
circadian rhythms by lithium carbonate, a prophylatic for depressive disorders. Int J
Chronobiol. 1983; 8(3):129-47.
Jones SH, Hare DJ, Evershed K. Actigraphic assessment of circadian activity and
sleep patterns in bipolar disorder. Bipolar Disord. 2005 Apr; 7(2):176-86.
Jornada LK e cols.. Effects of mood stabilizers on hippocampus and amygdala BDNF
levels in an animal model of mania induced by ouabain. Journal of Psychiatric
Research (2009),
Joyce, P.R., Porter, R.J., Mulder, R.T., Luty, S.E., McKenzie, J.M., Miller, A.L.,
Kennedy, M.A., 2005. Reversed diurnal variation in depression: associations with a
differential antidepressant response, tryptophan: large neutral amino acid ratio and
serotonin transporter polymorphisms. Psychol. Med. 35, 511 – 517.
86
Kafka MS, Wirz-Justice A, Naber D, Marangos PJ, O’Donohue TL, Wehr TA. Effect of
Lithium on Circadian Neurotransmitter Receptor Rhythms. Neuropsychobiology
1982;8:41-50.
Kalueff AV e Tuohimaa P. The grooming analysis algorithm discriminates between
different levels of anxiety in rats: potential utility for neurobehavioural stress research.
Journal of Neuroscience Methods 143 (2005) 169–177.
Kato T. Molecular genetics of bipolar disorder and depression. Psychiatry Clin
Neurosci. 2007 Feb;61(1):3-19.
Kawano, H., Decker, K. & Reuss, S. (1996) Is there a direct retinaraphesuprachiasmatic nucleus pathway in the rat? Neurosci. Lett., 212, 143–146.
Kendler KS, Thornton LM, Gardner CO. Stressful life events and previous episodes
in the etiology of major depression in women: an evaluation of the "kindling"
hypothesis. Am J Psychiatry. 2000 Aug;157(8):1243-51.
Klemfuss H (1992) Rhythms and the pharmacology of lithium. Pharmacol Ther 56:
53–78. doi: 10.1016/0163-7258(92)90037-Z
Koenigsberg HW, Teicher MH, Mitropoulou V, Navalta C, New AS, Trestman R, e
cols.. 24-h Monitoring of plasma norepinephrine, MHPG, cortisol, growth hormone
and prolactin in depression. J Psychiatr Res 2004;38(5):503-11.
Kolodziej ME, Griffin ML, Najavits LM, Otto MW, Greenfield SF, Weiss RD. Anxiety
disorders among patients with co-occurring bipolar disorder and substance use
disorder. Drug Alcohol Depend. 2005; 80:251–257.
Kripke DF, Wyborney VG. Lithium slows rat circadian activity rhythms, Life Sciences,
Volume 26, Issue 16, 21 April 1980, Pages 1319-1321.
Krishnan V, Han MH, Mazei-Robison M, Iniguez SD, Ables JL, Vialou V e cols.
(2008). AKT signaling within the ventral tegmental area regulates cellular and
behavioral responses to stressful stimuli. Biol Psychiatry 64: 691–700.
Kupke C. Melancholy as time-structuring-disorder: a transdisciplinary approach.
Paper presented at: Fourth International Conference on Philosophy and Psychiatry:
Madness, Science and Society; August 26-29, 2000; Florence, Italy.
Kushner MG, Sher KJ, Erickson DJ. Prospective analysis of the relation between
DSM-III anxiety disorders and alcohol use disorders. Am J Psychiatry. 1999;
156:723–732.
Lad HV, Liu L, Paya-Cano JL, e cols.. Behavioural battery testing: evaluation and
behavioural outcomes in 8 inbred mouse strains. Physiology & behavior.
2010;99(3):301-16.
Lambert GW, Reid C, Kaye DM, Jennings GL, Esler MD. Effect of sunlight and
season on serotonin turnover in the brain. Lancet 2002 Dec 7;360(9348):1840e2
87
Levander E, Frye MA, McElroy S, et al. Alcoholism and anxiety in bipolar illness:
differential lifetime anxiety comorbidity in bipolar I women with and without
alcoholism. J Affect Disord. 2007; 101:211–217.
Li J, Lu W-Q, Beesley S, Loudon ASI, Meng Q-J (2012) Lithium Impacts on the
Amplitude and Period of the Molecular Circadian Clockwork. PLoS ONE 7(3):e33292.
doi:10.1371/journal.pone.0033292
Lima MS, Tassi JL, Novo IP, Mari JJ. Epidemiologia do transtorno bipolar. 2003 http://www.hcnet.usp.br/ipq/revista/vol32/s1/15.html
Louzada F e Menna-Barreto L. Sleep-wake cycle expression. In adolescence:
influences of social contexto. Biological Rhythm Research. 2003, 34(2): 129-136.
Lucki I. The spectrum of behavior influenced by serotonin.
Psychiatry, Volume 44, Issue 3, 1 August 1998, Pages 151-162.
Biological
Mahmood T e Silverstone T. Serotonin and bipolar disorder. Review. Journal of
Affective Disorders, 2001; 66:1–11.
Maier SF e Walkins LR. Stressor controllability and learned helplessness: The roles
of
the
dorsal
raphe
nucleus,
serotonin,
and
corticotropin-releasing
factor. Neuroscience & Biobehavioral Reviews, Volume 29, Issues 4–5, 2005, Pages
829-841
Malkoff-Schwartz S, Frank E, Anderson BP, Hlastala SA, Luther JF, Sherrill JT,
Houck PR, Kupfer DJ. Social rhythm disruption and stressful life events in the onset
of bipolar and unipolar episodes. Psychol Med. 2000 Sep; 30(5):1005-16.
Marques N e Menna-Barreto LS. 2005. Cronobiologia: Princípios e Aplicações. 3ª
edição. Coleção Acadêmica. 448p.
Martynhak BJ, Correia D, Morais LH, Araujo P, Andersen ML, Lima MMS, Louzada
FM, Andreatini R. Neonatal exposure to constant light prevents anhedonia-like
behavior induced by constant light exposure in adulthood. Behavioural Brain
Research 222 (2011) 10–14.
Maurizi CP. A preliminary understanding of mania: roles for melatonin, vasotocin and
rapid-eye-movement sleep. Med Hypotheses. 2000 Jan; 54(1): 26-9.
Maurizi CP. A preliminary understanding of mania: roles for melatonin, vasotocin and
rapid-eye-movement sleep. Med Hypotheses. 2000 Jan; 54(1): 26-9.
McClung CA, Sidiropoulou K, Vitaterna M, Takahashi JS, Whiten FJ, Cooper DC,
Nestler EJ. Regulation of dopaminergic transmission and cocaine reward by the
Clock gene. PNAS, June 28, 2005, 102 (26): 9378-9381.
McClung CA. Circadian rhythms and mood regulation: Insights from pre-clinical
models. European Neuropsychopharmacology (2011) 21, S683–S693.
88
McCrone, P ; Sabes-Figueira, R. ; Razzouk, D. Economic of bipolar disorders. In:
Lakshni N; Maj M. (Org.). In Bipolar Disorders: Clinical and Neurobiological
Foundations. 1 ed. Londres: John Wiley &Sons Ltd, 2010, v. 1, pp. 90-95.
McEachron DL, Kripke DF, Hawkins R, Haus E, Pavlinac D, Deftos L. Lithium Delays
Biochemical Circadian Rhythms in Rats. Neuropsychobiology 1982;8:12-29
McIlwain KL, Merriweather MY, Yuva-Paylor L a, Paylor R. The use of behavioral test
batteries: effects of training history. Physiology & behavior. 2001;73(5):705-17.
Mental Health Survey, Brazil. PLoS ONE 7(2): e31879.
Merikangas KR, Akiskal HS, Angst J, Greenberg PE, Hirschfeld RMA, et al. (2007)
Lifetime and 12-month prevalence of bipolar spectrum disorder in the National
Comorbidity Survey replication. Arch Gen Psychiatry 64: 543–552.
Miret M, Ayuso-Mateos JL, Sanchez-Moreno J, Vieta E. Depressive disorders and
suicide: Epidemiology, risk factors, and burden. Neuroscience & Behavior reviews.
2013.
Mistlberger RE, Holmes MM. Behavioral feedback regulation of circadian rhythm
phase angle in light-dark entrained mice. Am J Physiol Regul Integr Comp
Physiol. 2000 Sep;279(3):R813-21.
Mitchell JD, Brown ES, Rush AJ. Comorbid disorders in patients with bipolar disorder
and concomitant substance dependence. J Affect Disord. 2007; 102:281–287.
Mitchell P, Parker G, Jamieson K, Wilhelm K, Hickie I, Brodaty H, Boyce P, HadziPavlovic D, Roy K. Are there any differences between bipolar and unipolar
melancholia? J Affect Disord. 1992 Jun; 25(2):97-105.
Moga, M. M. and Moore, R. Y. (1997), Organization of neural inputs to the
suprachiasmatic nucleus in the rat. J. Comp. Neurol., 389: 508–534.
Monteleone P e Maj M. The circadian basis of mood disorders: recent developments
and treatment implications.Eur Neuropsychopharmacol. 2008 Oct; 18(10):701-11.
Moore RY, Speh JC. Serotonin innervation of the primate suprachiasmatic nucleus.
Brain Res. 2004 Jun 4; 1010(1-2):169-73.
Moreno RA e Moreno DH. Da psicose maníaco-depressiva ao espectro bipolar.
Segmento Pharma, São Paulo, 2005. 448p.
Morris HV, Dawson GR, Reynolds DS, Atack JR, Stephens DN. 2006. Both alpha2
and alpha3 GABAA receptor subtypes mediate the anxiolytic properties of
benzodiazepine site ligands in the conditioned emotional response paradigm. Eur J
Neurosci 23(9):2495–2504.
89
Neto, SPD. Efeito da Dissociação do Ritmo Circadiano da Atividade Motora sobre a
Atividade Exploratória, Memória, Ansiedade e Depressão em Ratos. Dissertação de
Mestrado em Psicobiologia, UFRN, 2006.
Newman ME, Shapira B, Lerer B. Evaluation of central serotonergic function in
affective and related disorders by the fenfluramine challenge test: a critical review. Int
J Neuropsychopharmacol. 1998 Jul;1(1):49-69.
Nurnberger JI, Adkins S, Lahiri DK, e cols.. Melatonin suppression by light in
euthymic bipolar and unipolar patients. Archives of general psychiatry.
2000;57(6):572-9.
Pandi-Perumal SR, Moscovitch A, Srinivasan V, Spence DW, Cardinali DP, Brown
GM. Bidirectional communication between sleep and circadian rhythms and its
implications for depression: lessons from agomelatine. Prog Neurobiol. 2009 Aug;
88(4):264-71.
Paul ED e Lowry CA. Functional topography of serotonergic systems supports the
Deakin/Graeff hypothesis of anxiety and affective disorders. Journal of
Psychopharmacology 0(0) 1–17; 2013.
Paul JR, Johnson RL Jope RS, Gamble KL. Diruption of circardian rhythmicity and
suprachiasmatic action potential frequency in a maouse model with constitutive
activation of glycogen synthase kinase 3. Neuroscience 226 (2012) 1–9.
Paulus EV e Mintz EM. Developmental disruption of the serotonin system alters
circadian rhythms. Physiology & Behavior 105 (2012) 257–263.
Pflug B, Tolle R (1971): Therapie endogener Depressionen durch Schlafentzug.
Nervenarzt 42:117–124 (translated).
Polter AM e Li X. Glycogen synthase kinase-3 is an intermediate modulator of
serotonin neurotransmission. Frontiers in Molecular Neuroscience 4(31): 1-14, 2012.
Polter AM, Beurel E, Yang S, Garner R, Song L, Miller CA, Sweatt, JD, McMahon L,
Bartolucci A, Li X and Jope RS (2010). Deficiency in the inhibitor serinephosphorylation of glycogen synthase kinase – 3 increases sensitivity to mood
disturbances. Neuropsy- chopharmacology 35, 1761–1774.
Porterfield T, Cook M, Deary IJ, Ebmeier KP. Neuropsychological function and
diurnal variation in depression. J Clin Exp Neuropsychol 1997;19(6):906-13.
Post RM, Susan R, Weiss B. Sensitization, kindling, and carbamazepine: an update
on their implications for the course of affective illness. Pharmacopsychiatry. 1992
Jan; 25(1):41-3.
Prosser RA. Serotonin phase-shifts the mouse suprachiasmatic circadian clock in
vitro. Brain Research. 2003. 966; 110-115.
90
Prut L, Belzung C. The open field as a paradigm to measure the effects of drugs on
anxiety-like behaviors: a review. Eur J Pharmacol 2003; 463: 3-33.
Refinetti R. Endogenous Mechanisms. In: Circadian Physiology. Second Edition.
2006. P. 220.
Reynolds CF e Kupfer DJ. Sleep research in affective illness. Sleep. 1987 Jun;
10(3):199-215.
Rosenkranz JA, Moore H, Grace AA. The Prefrontal Cortex Regulates Lateral
Amygdala Neuronal Plasticity and Responses to Previously Conditioned Stimuli. The
Journal of Neuroscience, December 3, 2003. 23(35):11054 –11064.
Rosenkranz JA, Moore H, Grace AA. The Prefrontal Cortex Regulates Lateral
Amygdala Neuronal Plasticity and Responses to Previously Conditioned Stimuli. The
Journal of Neuroscience, December 3, 2003 • 23(35):11054 –11064.
Roybal K, Theobold D, Graham A, DiNieri JA, Russo SJ, Krishnan V e cols. (2007).
Mania-like behavior induced by disruption of CLOCK. Proc Natl Acad Sci USA 104:
6406–6411.
Rusting CL, Larsen RJ. Diurnal patterns of unpleasant mood: associations with
neuroticism, depression, and anxiety. J Pers 1998;66(1):85-103.
Sahar S, Zocchi L, Kinoshita C, Borrelli E, Sassone-Corsi P (2010) Regulation of
BMAL1 protein stability and circadian function by GSK3beta-mediated
phosphorylation. PLoS One 5: e8561.
Schibler U. The daily rhythms of genes, cells and organs. Biological clocks and
circadian timing in cells. EMBO Rep. 6 Spec No:S9-13, 2005.
Schildkraut JJ. The catecholamine hypothesis of affective disorders: a review of
supporting evidence. Am J Psychiat 1965;122:509–22.
Schwartz MD, Wotus C, Liu T, Friesen WO, Borjigin J, Oda GA, de la Iglesia HO.
Dissociation of circadian and light inhibition of melatonin release through forced
desynchronization in the rat. 17540–17545 _ PNAS _ October 13, 2009 _ vol. 106 _
no. 41
Scotti MAL, Lee G, Stevenson GA et al. Behavioral and pharmacological assessment
of a potential new mouse model for mania. Physiology & Behavior 103 (2011) 376–
383.
Shaltiel G, MaengS, Malkesman O, Pearson B, Schloesser RJ, Tragon T,Rogawski
M, Gasior M, Luckenbaugh D, Chen D, and Manji HK. Evidence for the involvement
of the kainate receptor subunit GluR6 (GRIK2) in mediating behavioral displays
related to behavioral symptoms of mania Mol Psychiatry. 2008 September ; 13(9):
858–872.
91
Shiah IS, Yatham LN. Serotonin in mania and in the mechanism of action of mood
stabilizers: a review of clinical studies. Bipolar Disord. 2000 Jun;2(2):77-92.
Souetre E, Salvati E, Belugou JL, e cols. Circadian rhythms in depression and
recovery: evidence for blunted amplitude as the main chronobiological abnormality.
Psychiatry Res. 1989;28:263-278.
Spengler ML, Kuropatwinski KK, Schumer M, Antoch MP (2009) A serine cluster
mediates BMAL1-dependent CLOCK phosphorylation and degradation. Cell Cycle 8:
4138–4146.
Stahl SM. Psicofarmacologia – Base Neurocientífica e Aplicações Práticas. 2ª
edição. Editora Guanabara. 2006. 617pp.
Stefos G, Staner L, Kerkhofs M, Hubain P, Mendlewicz J, Linkowski P. Shortened
REM latency as a psychobiological marker for psychotic depression? An age-,
gender-, and polarity-controlled study. Biol Psychiatry. 1998 Dec 15; 44(12):1314-20.
Tang X e Sanford LD. Home cage activity and activity-based measures of anxiety in
129P3/J, 129X1/SvJ and C57BL/6J mice. Physiology & Behavior 84 (2005) 105– 115
Tang X, Orchard SM, Sanford LD. Home cage activity and behavioral performance in
inbred and hybrid mice. Behav Brain Res 2002; 136:555–69.
Teicher MH. Actigraphy and motion analysis: new tools for psychiatry. Harv Ver
sychiatry. 1995;3(1):18-35.
Thase ME. Treatment issues related to sleep and depression. J Clin Psychiatry.
2000; 61 Suppl 11:46-50.
Treiser S e Kellar KJ. Lithium: effects on serotonina receptors in rat brain. European
Journal of Pharmacology, 64 (1980): 183-185.
Uhlhaas PJ e Singer W. Neural Synchrony in Brain Disorders: Relevance for
CognitiveDysfunctions and Pathophysiology Neuron 52, 155–168, October 5, 2006
van Gaalen MM, Steckler T. Behavioural analysis of four mouse strains in an anxiety
test Battery. Behavioural Brain Research 115 (2000) 95–106.
Vetter C, Juda M, Roenneberg T. The influence of internal time, time awake, and
sleep duration on cognitive performance in shiftworkers.Chronobiol Int. 2012
Oct;29(8):1127-38.
Vicentic A.Francis D, Moffett M, Lakatos A et al. Maternal Separation alters
serotonergic transporter densities and serotonergic 1A receptors in rat brain.
Neuroscience 140 (2006) 355–365.
Vilaplana J; Cambras T; Campuzano A; Diez-Noguera A. Simultaneous
manifestation of free-running and entrained rhythms in the rat motor activity
explained by a multioscillatory system. Chronobiol. Int. 14:9 –18; 1997.
92
Waterhouse J, Jones K, Edwards B, Harrison Y, Nevill A, Reilly T. Lack of evidence
for a marked endogenous component determining food intake in humans
during forced desynchrony. Chronobiol Int. 2004 May;21(3):445-68.
Wehr TA, Sack D, Rosenthal N, Duncan W, Gillin JC. Circadian rhythm disturbances
in manic-depressive illness. Fed Proc. 1983 Aug;42(11):2809-14.
Wehr TA, Wirz-Justice A, Goodwin FK, Duncan W, Gillin JC. Phase advance of the
circadian sleep-wake cycle as an antidepressant. Science 1979;206(4419):710-3.
WHO – World Health Organization. WHO Library Cataloguing-in-Publication Data.
The
global
burden
of
disease:
2004
update.
Acessível
em:
http://www.who.int/healthinfo/global_burden_disease/GBD_report_2004update_full.p
df
Willie M. Lived Time and Psychopathology. 2005 (12) 3:173-281. The John Hopkins
University Press Journals Division.
Willner P, Towell A, Sampson D, Sophokleous S, Muscat R (1987): Reduction of
sucrose preference by chronic unpredictable mild stress, and its restoration by a
tricyclic antidepressant. Psychopharmacology (Berl) 93:358–364.
Wirz-Justice A, van der Hoofdakker RH. Sleep deprivation in depression: What do we
know, where do we go? Biol Psychiatry, 1999. 46:455-453.
Wirz-Justice A. Biological rhythm disturbances in mood disorders.
Psychopharmacol. 2006 Feb;21 Suppl 1:S11-5. Review.
Int Clin
Wood J, Birmaher B, Axelson D, e cols.. Replicable differences in preferred circadian
phase between bipolar disorder patients and control individuals. Psychiatry:
Interpersonal and Biological Processes. 2009;166:201-209.
Yamakawa GR e Antle MC. Phenotype and function of raphe projections to the
suprachiasmatic nucleus. European Journal of Neuroscience, Vol. 31, pp. 1974–
1983, 2010.
Yin L, Wang J, Klein PS, Lazar MA (2006) Nuclear receptor Rev-erb alpha is a
critical lithium-sensitive component of the circadian clock. Science 311:1002–1005.
Young JW, Goey AKL, Minassian A, Perry W, Paulus MP, Geyer MA. The mania-like
exploratory profile in genetic dopamine transporter mouse models is diminished in a
familiar environment and reinstated by subthreshold psychostimulant administration.
Pharmacology, Biochemistry and Behavior 96 (2010) 7–15.
93
ANEXOS
ANEXO I – Rotina do MatLab para auxiliar a determinação das fases
clear all
dado=xlsread('T22Bruna.xls');
dado=dado-300;
dado=-dado;
%%
tam=fix(length(dado)/264);
dado=dado(1:tam*264,:);
act=reshape(dado,264,tam,26);
%%
clear a
for j=1:26
for i=1:tam
a(i,j)=sum(act(1:132,i,j))-sum(act(133:end,i,j));
end
end
%%
close all
for x=1:26
plot(a(:,x+2))
hold on
plot(a(:,x+2),'o')
figure
hold off
end
94
ANEXO II – Hibridização in situ
In Situ Hybridization with Radioactive Probes
Wear clean gloves throughout the procedure to protect yourself and to prevent RNAse
contamination.
Day 1 - Prehybridization
Buffers
0.1M Phosphate buffered paraformaldehyde (Prepare fresh or a day before):
700 ml of DEPC-treated water
40g of PARA (EMS 19210)
10.931g dibasic sodium phosphate (Na2HPO4, MW 142, Sigma S-3264)
3 beads of NaOH (Sigma S-0889)
Heat on a stirring plate to no more than 55oC. Once clear add:
2.76g Monobasic sodium phosphate (Na2H2PO4, MW 120g, Sigma S-3139)
Cool, adjust pH to 7.3 at 4ºC and bring volume to 1 liter with DEPC-treated water
Filter using sterilizing filters with vacuum or Wattman paper and keep at 4ºC until used
20X SSC:
175.3g NaCl
88.2g sodium citrate (tribasic dihydrate, MW 294.1)
800ml DEPC-treated water
Adjust to pH7.0 and volume to1 liter with DEPC-treated water
Make dilutions to desired strength
TEA HCl/Acetic Anhydride (this solution, without the acetic anhidride, can be stored for up to 2
weeks at room temperature)
18.56g TEA HCl
9g NaCl
20 pellets of NaOH
95
800ml DEPC-treated water
pH to 8.0 and adjust volume to 1 liter with DEPC-treated water
Add 125 µl of acetic anhydride per 50 ml TEA HCl (just before use)
Procedure
1. Remove slides from –80 freezer and lay them out on slide racks or Copeland jars
2. Fix by dipping in 0.1M Phosphate buffered 4% paraformaldehyde. 5 min.
3. Rinse in 2XSSC. 2 min.
4. Acetylate in 0.25% Triethanolamine HCl/Acetic Anhydride with 125µl acetic anhydride per 50 ml
TEA HCl added just before using. 10 min.
5. Quick rinse in 2X SSC.
6. Dehydrate/delipidate as follows:
1 min 70% EtOH (prepared in DEPC-treated water)
1 min 80% " "
2 min 95%
"
1 min 100%
5 min CHCl3
1 min 100% EtOH
1 min 95%"
7. Air dry
Slides may be returned to freezer for storage until hybridization at a later time. How long they can be
stored there, or whether freezing is necessary, needs to be empirically determined. We always
continue the procedure on the same day.
Hybridization
Buffers
Hybridization buffer
Ingredients:
20X SSC
Deionized formamide (Sigma # F-9037)
96
100X Denhardt's:
10g Ficoll (Type 400, Pharmacia)
10g polyvinylpyrollidone
10g BSA (fraction V)
H2O to 500ml
filter; store at -20C
50X Denhardt’s can be also purchased from Sigma (# D-2532)
Yeast tRNA (Roche catalog # 0109495) resuspended at 25mg/ml in DEPC
Sheared single stranded DNA (Sigma catalog # D-7656), diluted to 10mg/ml in DEPC-treated
water.
To shear DNA, draw up and down12 times with 17-22 gauge syringe and store at –20ºC. Just
before added to the hybridization buffer, heat 5 minutes in boiling water and chill quickly in
wet ice.
Dextran sulfate (500,000MW, Sigma)
5M DTT in 0.01 M sodium acetate (pH5.2)
Mix 77.25mg DTT in 100µl sodium acetate immediately before use.
Do not use frozen aliquots of DTT. (Note: this makes enough for only 10ml of hybridization buffer
after the 1:100 dilution you will do below). 0.01 M sodium acetate can be prepared and stored at
room temperature for several months.
Preparation:
Add 2 g dextran sulfate to 4.5 ml DEPC water in 50 ml Falcon tube
Vortex, let stand 10 min vortexing every now and then
Add 10 ml deionized formamide, vortex, heat to 37ºC until in solution, vortexing now and then
(use relatively fresh formamide)
Add 2 ml of 20X SSC.
Add 200 µl 100X Denhardt's (if using 50X add 400 µl)
97
Add 400 µl tRNA (25mg/ml)
Add 800µl sheared single stranded DNA
Adjust volume to 20 ml with DEPC-treated water, vortex, store at -20C (can be stored for up to 3
months)
Immediately before using add to the needed volume of 5M DTT for a dilution of 1:100
Procedure
8.
Prehybridize with a solution containing 50% 4XSSC/50% deionized formamide for 15 to 60
minutes at 37ºC. This step is done by adding approximately 50 µl of solution to the slide and
coversliping it.
From this step on use extreme care when handling radioactive solutions.
9.
Remove the coverslip, tap the slide gently to remove the solution from step 8 and replace
with 25-35µl of hybridization buffer containing 108 cpm of probe per ml. Hybridize overnight
at 55 ºC under cover slips.
Mix 77.25mg DTT in 100µl sodium acetate immediately before use.
Do not use frozen aliquots of DTT. (Note: this must account for 1% of the final volume of
hybridization buffer). 0.01 M sodium acetate can be prepared and stored at room
temperature for several months.
Adjust volume per slide to the size of the coverslip (30 µl enough for a 22X22 mm coverslip.) Use
humidified chamber (Tupperware is OK), putting a couple of blue screwcaps from 50-ml Falcon
tubes containing water so that tissue does not dry out. Depending on container it may be necessary
to further seal with parafilm.
Day 2 - Posthybridization
Buffers
Solution containing 50% formamide (NOT deionized, Sigma # F-7503) and 50% 2X SSC.
Prepare as much as needed for the slide racks or Copeland jars and place in bath at 52ºC.
Prepare the solutions below in the RNAse room, and with miliQ water not treated with DEPC.
RNAse buffer:
100 ml 5M NaCl
1 ml 1M Tris, pH 8.0
98
2 ml 0.5M EDTA, pH 8.0
Adjust volume to 1 liter. It does not need to be pHed.
RNAse buffer can be stored at room temperature for several months.
Just before use, add 100 mg RNase A (Sigma # R-4642) per liter of buffer. Prepare only the
volume needed
20X SSC
Prepared as above
Procedure
Steps 10 to 15 are done with RNAse-free solutions (prepared with DEPC-treated water) in the RNAse
free area
Before starting prepare enough 50% formamide/50% 2XSSC for steps 14 and 15 and place them in
water bath at 52ºC
10.
Set up 3 beakers of 1X SSC solution
11.
Cool and remove coverslips on top of a piece of bench paper (for radioactive waste). If
coverslips are stuck, dip slides in 1XSSC and revise your procedure to seal containers.
12.
Soak and swirl slides briefly in first, second and third beaker and put back on rack or
Copeland jar, containing 1XSSC. 10 min.
13.
Do a second wash in 1XSSC. 10 min.
Depending on how many slides you have these 10 minute washes will be of variable times for each
slides but this does not affect the results according to our experience. Up to this step, slides
hybridized with different probes cannot not be placed in the same container.
14.
Wash in 50% formamide/50% 2X SSC @ 52ºC. 5 min.
15.
Wash in 50% formamide/50% 2X SSC @ 52ºC. 20 min.
As soon as you start this step, set RNAse water bath (in RNAse room) to 37ºC and put in a container
with enough RNAse buffer
16.
Rinse 2 times in 2X SSC at room temperature for 1 min each. These steps can be done in the
RNAse room if more convenient.
99
All rinses up to this step should be treated as radioactive and should be drained in
radioactive sink letting tab water running. From this step on it is not necessary to use
RNAse-free water
17.
Add 3-30mg/L RNAse (less w/ newer RNAse) to the buffer and incubate in RNAse solution at
37ºC. 30 min.
This and all steps below are carried out in the RNAse room. Use RNAse pipettes only.
18.
Wash 2 times in 2XSSC at room temperature. 5 min each.
As soon as you take slide racks out of the bath, set bath to 52ºC. The 5-minute washes can be
extended if time is needed for bath to reach 52ºC
19.
Wash in 50% formamide/50% 2X SSC @ 52ºC. 5 min.
20.
Wash in ethanol series as follows:
70% ethanol in 0.1XSSC. 3 min.
(70 ml 100%ETOH, 30 ml 0.1X SSC)
80% ethanol in 0.1XSSC. 3 min.
95% ethanol in 0.1X SSC. 3 min.
Quick wash in ddH2O
70% ethanol in water. 3 min.
21.
Air dry
100
Anexo III – Sequências das probes utilizadas para a Hibridização in situ
Receptor 5HT1A
CAGANGCGTTACGTATCGGATCCAGAATTCGTGATTTTGAGTGAACAGGAAGGGTCCTGTAAGCCCTACACTCTTCCTCCA
CTTCTTCCTTCTCCATCACCACCACTATTACCACCACTACCACCACCATCATCATCATCAGCGGCAGAACTTGCACTTGATG
ATCTTCTTAAAAGCGTTTTGAAAGTCTTTGTTGAAATAAGCATAAATAACTGGGTTGAGCAGGGAGTTGGAGTAGCCTAGCC
AGTTAATTATGGCACCCAACAACTCAGGCATGTGGCAACTGCTCTCACAGAAAGGTAGGACCAGGGCCACAATGAAAAAGG
GCAGCCAGCAGAGGATGAAGGTGCCCATGATGATGCCCAGTGTCTTCACTGTCTTCCTCTCACGGGCCAAGGCCATCTTG
CGCTTTGCCTCAGCAGTGCGCTCATTTTTTCTCTCCAAGCAGGCGGGGACATAGGAGGTAGCTCCTGATTCGCTGGGCAG
AGGAAGGTGCCCTTTGGAGTTGCCCACTCGATGCACCTCGATCACCTCCAGGGTGGCGTCGTCCTCACCCTGTCTCACCG
CCCCATTAGCGCACGGAGTCCCCACCGCCCTGTTCTCAGCACTGCGCCTGCAGTCCCCACTACCTGGCTGACCATTCAGG
CTCTTCTTGGGGGGCGGGGCCGACGATGTTCCGAAGCTGGTGCCCGCTCCCTTCTTTTCCACCTTCTTGACCGTCTTGCG
GATTCGGAAGCGCGCGGCTCTGAAGATGCGCCCATAGAGGACCAGCATGAGCAGCAGCGGAATATAGAAATCTGAATTCG
TCGACAAGCTTCTCGAGCCTAGGCTAGCTCTAGACCACACGTGTGGGGGCCCGAGCTCGCGGCCGCTGTATTCTATAGTG
TCACCTAAATGGCCGCACAATTCACTGGCCGTCGTTTTACAACGTCGTGACTGGGAAAACCCTNNNTTACCCAACTTAATCG
CCTTGCAGCACATCCCCCTTTCGCCAGCTGGCGTAATAGCGAANANGCCCGCNNCGATCNCCCTTCCCAACAGTNGCGCA
GCCTG
Receptor 5HT2A
CACGTGTGGTCTAGAGCTAGCCTAGGCTCGAGAAGCTTGTCGACGAATTCAGATTAGGAAACCCAGCAGCATATCAGCTAT
GGCAAGTGACATCAGGAAATAGTTGGTGGCATTCTGCAGCTTTTTCTCTAGGGACACTGCCATGATGACCAGTATGTTTCCC
CCAATGGTGAGAATAATCACGACAGTTGTCAATAAAGCAGACCAGTTTTTTTCCTGGAGATGAAGAATGGAGAGGCATGTC
GGTGGGAGGTACCCTTCGCAGGAGAGGTTGGTTCTGTTTTCAGCATCAATTGTCCAGTTCGAGGCTTCGGAAGTGTTAGCA
TCCCTGGAGTTGAAGTCATTAGGGTAGAGCCTCGAGTCGTCACCTAATTGCATTAAGGAGTTTGGAATTGAGCTCAGGGAG
ATATTGTCTTCACAGAGAATTTCCATGTCTAAGCCGGAAGTTGTAGCAGATGAAGTGTGGAGAGACTAGCGAGTCATAGCC
GGGGCTCACCATTCATCTTTACGTCCTGGAGCTATCACGAATTCTGGATCCGATACGTAACGCGTCTGCAGCATGCGTGGT
ACCGAGCTTTCCCTATAGTGAGTCGTATTAGAGCTTGGCGTAATCATGGTCATAGCTGTTTCCTGTGTGAAATTGTTATCCG
CTCACAATTCCACACAACATACGAGCCGGAAGCATAAAGTGTAAAGCCTGGGGTGCCTAATGAGTGAGCTAACTCACATTA
ATTGCGTTGCGCTCACTGCCCGCTTTCCAGTCGGGAAACCTGTCGTGCCAGCTGCATTAATGAATCGGCCAACGCGCGGG
GAGAGGCGGTTTGCGTATTGGGCGCTCTTCCGCTTCCTCGCTCACTGACTCGCTGCGCTCGGTCGTTCGGCTGCGGCGAG
CGGTATCAGCTCACTCAAAGGCGGTAATANGGTTATCCACAGAATCAGGGGATAACGCNNAAAGAACATGTGAGCAAANNN
CAGCAAAGGCCAGGAACCGTAAAAAGCCGCGTTGCTGGCGTTTTTCCATANNTCNCCCCCNGACGAGCATCACAAAATCG
ACGCTCAAGTCNGAGNNGCGAACCCGACAGGAC
101
Anexo IV – Protocolo da Imunohistoquímica para 5HT1A + DAB
Day 1
-
post-fixed with ice-cold 4% paraformaldehyde in 0.1 M phosphate buffer pH7.3
for 5 min
-
washed in PBS (3x5 min)
-
wash in PBS containing 0.4% Triton X-100 (3x5 min)
-
Blocking: incubated in PBS containing 0.4% Triton X-100 and 6% normal goat
serum (NGS) and 0.5% H2O2 for 1h at RT.
-
sections were incubated overnight at 4 °C with the rabbit polyclonal anti-5HT2A antibody (Santa Cruz, USA) at a concentration of 2 ug/ml (1:200) diluted
in PBS containing 0.4% Triton X-100 and 4% NGS.
Day 2
-
Following primary antibody incubation, slides were washed in PBS (3x5 min)
-
incubated for 30 min at RT with a biotinylated goat anti-rabbit (Vector) at a
concentration of 5g/ml (1:400) diluted in PBS containing 1.5% NGS.
-
Following three rinses in PBS (3x5 min)
-
slides were exposed to the ABC reagent (DakoCytomation) for 30 min at RT.
-
Slides were then washed in PBS (3x5 min)
-
stained with 3,3=-diaminobenzidine (DAB) for 3-4 min at RT.
-
Finally, they were washed, dry,coverslipped with Permount and analyzed
microscopically under a bright-field microscope.
102
ANEXO V – EFEITO DA SEQUÊNCIA EM TESTES COMPORTAMENTAIS
Koike, BDV; Leite, MD; Ribeiro, JM; Gonçalves, BSB; Araujo JF
Departamento de Fisiologia, Universidade Federal do Rio Grande do Norte, Brasil
Resumo
Vários estudos tem sido realizados a fim de acessar parâmetros comportamentais
indicativos do estado de humor dos animais como ansiedade, anedonia, depressão
e atividade locomotora. Cada um dos testes avalia certas características e nos traz
determinadas informações, sendo muitas vezes complementares às obtidas em um
outro teste. O mais comumente utilizado é uma bateria de testes para avaliar as
respostas comportamentais dos animais em experimentação, mas como saber que
um teste não interfere no resultado do outro? Para responder essa questão,
realizamos o presente trabalho, onde avaliamos o efeito de ordem nos testes
comportamentais. Além disso, verificamos as diferenças comportamentais com
intervalos entre os testes de 24h ou de 7 dias. Os testes utilizados em nossa bateria
foram: Labirinto em Cruz Elevado (LCE), Campo Aberto (CA), Nado Forçado (NF) e
Preferência Claro-Escuro (PCE), aplicados em diferentes ordens em ratos Wistar
machos, que foram acomodados em grupos de 5 indivíduos, tendo acesso ad libitum
a água e ração. Com os resultados obtidos, podemos sugerir que é importante
determinar a ordem na qual serão realizados os testes para evitar a interferência nos
resultados. Os nossos dados sugerem que os testes com características mais
aversivas e estressantes sejam os últimos. No caso da bateria de testes utilizada em
nosso estudo, sugerimos como ideal a sequência: CA – LCE – PCE – NF.
103
1. Introdução
O estudo das ciências comportamentais nos permite acessar variados
aspectos biológicos do animal, sendo possível observar como essas variáveis se
integram e interagem. Estudos mais invasivos não são capazes de ser realizados
em humanos, como por exemplo estudos de neuroquímica, e por este motivo
modelos animais são utilizados a fim de responder tais perguntas. Segundo van der
Staay (2006), modelo animal é um organismo vivo usado para estudar as relações
cérebro-comportamento sob condições controladas, com a finalidade de ter um
insight, para permitir previsões sobre essas relações em humanos.
Os testes comportamentais nos trazem informações sobre o fenótipo dos
animais e, conforme seu desempenho, podemos inferir qual estado de ânimo está
mais caracterizado pelo comportamento apresentado. Devemos assumir que os
sintomas apresentados pelos animais são comparáveis aos sintomas dos humanos.
Entretanto não pode ser provado que os animais experimentam, por exemplo, a
ansiedade como os humanos, mas é aceitável que alguns padrões comportamentais
dos roedores indiquem ansiedade (Ohl, 2003).
Cada um dos testes nos traz informações sobre determinado comportamento.
Porém, um único teste pode não nos trazer informações suficientes para que
possamos classificar um comportamento. Entretanto um conjunto de testes,
combinados, pode caracterizar melhor o estado de ânimo quando agrupamos os
resultados de uma bateria de testes comportamentais para que se tenha uma
coleção de características que indiquem o real estado afetivo do animal.
Há estudos que utilizam poucas tarefas para verificar as respostas
comportamentais, o que pode acarretar em resultados falso positivos ou falso
negativos (van Gaalen e Steckler, 2000). Assim, o uso de uma bateria de testes
comportamentais se torna vantajoso, uma vez que várias respostas serão testadas
em um mesmo animal, o que permite que tenhamos uma visão mais abrangente do
que está correlacionado a determinado fenótipo. Por este motivo a utilização de uma
bateria de testes é a forma mais comum para avaliar respostas comportamentais
dos animais em experimentação (Crawley e Paylor, 1997; Cryan e Holmes, 2005;
104
Chourbaji et al. 2008; Pandey et al. 2009; Lad et al. 2010). Além disso, a utilização
do mesmo animal em vários testes reduz o número total de animais necessários
para completar um projeto (McIlwain et al., 2001).
Geralmente é eleita uma determinada sequência para que os testes sejam
realizados em uma ordem de tal modo que os testes menos invasivos sejam
executados antes dos mais invasivos. Pois a desvantagem de estudar várias
respostas comportamentais no mesmo animal é a possibilidade de haver efeitos de
um teste que influencie as respostas do teste subsequente (McIlwain et al., 2001).
Alguns estudos de caracterização fenotípica em camundongos preocuparam-se em
avaliar o efeito de ordem dos testes comportamentais (Crawley e Paylor, 1997; Lad
et al. 2010; McIlwain et al., 2001; Paylor et al. 2006). Em ratos, somente um estudo
foi publicado até o momento (Blokland et al., 2012).
A fim de avaliar o efeito de ordem dos testes comportamentais em ratos,
realizamos o presente trabalho, onde verificamos qual a melhor ordem para ser
realizada uma bateria de testes, com intervalos de 24h ou de 7 dias entre os testes.
Os testes utilizados em nossa bateria foram: Campo Aberto (CA), Nado
Forçado (NF), Labirinto em Cruz Elevado (LCE) e Preferência Claro-Escuro (PCE).
2. Materiais e métodos
2.1.Animais experimentais
Utilizamos 40 ratos machos da linhagem Wistar, com idade de 3 meses no
início do experimento, nascidos em cativeiro, oriundos do Laboratório de
Neurobiologia e Ritmicidade Biológica do Departamento de Fisiologia, Universidade
Federal do Rio Grande do Norte, Natal RN, Brasil. Este trabalho foi aprovado pelo
Comitê de Ética no Uso de Animais da UFRN.
Durante o período de experimentação os animais foram mantidos em grupos
de 5 animais por gaiola em ambiente com temperatura e humidade controladas, sob
um ciclo claro/escuro de 12/12h (início às 6h).
105
Os animais foram subdivididos em 8 grupos, com 5 animais cada. Cada grupo
realizou a bateria de testes de avaliação comportamental segundo a ordem descrita
na Tabela 1.
Tabela 1. Grupos experimentais e respectiva sequência de testes.
Grupos
1º Teste
2º Teste
3º Teste 4º Teste
24h 7dias
G1
G5
CA
LCE
NF
PCE
G2
G6
LCE
CA
NF
PCE
G3
G7
NF
CA
LCE
PCE
G4
G8
NF
LCE
CA
PCE
CA – teste de Campo Aberto; PCE – teste de Preferência Claro – Escuro; NF – teste
de Nado Forçado; LCE – teste de Labirinto em Cruz Elevado
O intervalo estabelecido entre os testes dos grupos 1 a 4 foram de 24h. Já
nos grupos de 5 a 8 o intervalo entre os testes foi de 7 dias.
2.2.Handling
Uma vez que os animais têm de ser manipulados para serem colocados nos
aparatos de teste, situação que aumenta o seu nível de estresse, o procedimento de
handling permite a habituação dos animais ao contato e a manipulação pelo
experimentador, diminuindo o estresse causado pela manipulação e diminuindo,
consequentemente, a sua interferência no resultado dos testes de avaliação
comportamental. O procedimento de handling consiste no manuseio dos animais
por curtos períodos de tempo, e foi realizado em duas seções de 5 minutos por
animal, com 3 dias de intervalo, uma semana antes do início dos testes de avaliação
comportamental.
106
2.3. Testes de Avaliação Comportamental
Os testes foram iniciados às 18:30h, correspondente ao início da fase de
escuro, momento no qual os ratos apresentam maior atividade. Cada grupo
experimental realizou um único teste por dia, durante quatro dias consecutivos para
os grupos 1 a 4 ou durante 4 semanas para os grupos 5 a 8, sendo realizado um
teste por semana.
Todos os testes foram realizados no escuro total (<1 lux). Os testes foram
filmados, com câmara de infravermelho, para posterior análise dos resultados.
2.3.1. Campo aberto (Insight® modelo EP 154)
O teste de campo aberto foi realizado numa arena circular, cercada de
paredes acrílicas opacas, base em acrílico de 100 cm x 80 cm dividida em
quadrantes centrais e periféricos, com dimensões: 50 cm (altura) x 60 cm (diâmetro).
Os animais foram colocados inicialmente no centro do aparato e observados por 5
minutos.
Foram aferidos: o deslocamento total (nº de quadrantes), o tempo gasto
(segundos) e o deslocamento no centro e na periferia do aparato.
2.3.2. Labirinto em Cruz Elevado
O labirinto em cruz elevado é composto por quatro braços de madeira medindo,
cada um, 50 cm de comprimento por 12 cm de largura. Dois dos braços são
circundados lateralmente por paredes opacas de 40 cm de altura e dispostos
perpendicularmente aos outros dois braços que permanecem abertos, sem paredes
laterais. Todo o conjunto permanece elevado a 50 cm do solo.
Os animais iniciam o teste no centro do aparato de frente para o braço aberto.
O teste tem a duração de 5 minutos. Para avaliar o nível de ansiedade foram
aferidos: o tempo que o animal passa nos braços abertos, nos braços fechados e no
centro do aparato, bem como número de entradas em cada um destes locais para
107
avaliar a locomoção.
2.3.3. Nado Forçado (Insight® modelo EP 180)
O teste do nado forçado foi realizado num aparato cilíndrico de acrílico (Altura
x Frente x Lado: 61,5 cm x 44 cm x 47 cm), com capacidade para 39 litros de água,
cheio até 15 cm antes da borda, para que o animal não consiga fugir e nem tocar o
fundo do aparato. A temperatura da água foi aferida (~26°C) e em seguida os
animais foram colocados no aparato e observados por 5 minutos. A água do cilindro
foi trocada a cada animal.
Neste teste, o animal é exposto a estímulos aversivos e estressantes e é
mensurada a latência para o “comportamento de desespero”.
2.3.4. Preferência Claro - Escuro (Insight® modelo EP 157)
O equipamento é composto por dois compartimentos separados por uma
divisória com passagem entre o lado claro (revestido com chapa de acrílico branco A x L x P (cm) 21 x 45 x 41) e o lado escuro (revestido com chapa de acrílico preto A x L x P (cm) 21 x 35 x 41), e tampa de acrílico transparente. Os animais iniciaram
o teste no lado claro do aparato e foram observados por 5 minutos.
Para avaliar o nível de ansiedade e o comportamento exploratório foram
aferidos: o tempo de permanência e o deslocamento em cada lado, bem como o
número de transições entre os dois lados.
2.3.5. Registro da temperatura corporal
Para analisarmos a temperatura central do animal durante o teste do nado
forçado, foi realizado um experimento paralelo, utilizando 10 ratos adicionais (que
não foram utilizados na bateria de testes). A temperatura foi registrada utilizando um
termistor (iButton®) que foi implantado na região peritoneal do animal. Os dados
108
foram adquiridos a cada minuto durante 24h antes do teste, durante o nado forçado
e durante 10h depois do teste.
2.4. Processamento e análise de dados
As medidas aferidas descritas acima foram analisadas por meio do teste
ANOVA de uma via, comparando-se o desempenho entre os grupos, seguida do
post-hoc de Tukey. Observadas as diferenças no desempenho, foi avaliada qual a
melhor ordem para serem realizados os testes, onde um tenha a menor interferência
possível sobre o outro. Os resultados mostrados correspondem a média e erro
padrão.
3. Resultados
No presente estudo pudemos observar que os testes realizados em
sequência causam certa interferência no resultado do teste seguinte, tanto com
intervalo de 24h quanto com intervalos maiores, de 7 dias entre eles.
No teste do campo aberto (CA) não houve diferença entre os grupos quanto a
atividade locomotora. Todos os grupos exploraram igualmente o aparato
independente do teste antecedente e do intervalo entre eles. O deslocamento na
periferia do CA também foi semelhante entre os grupos. Entretanto, quando
avaliamos o deslocamento no centro do aparato, os animais do G2 apresentaram
menor exploração do centro durante o teste quando comparados com G1
(13.20±2.21 x 24.20±3.22 quadrantes, p=0.027), como podemos ver na Figura 1A.
O intervalo de 24h entre os testes não mostrou prejuízo no desempenho dos
animais no teste do LCE, tanto a exploração do labirinto quanto o tempo de
permanência em cada um dos braços apresentaram resultados semelhantes em
todos os grupos que realizaram o teste com este intervalo. Mesmo com o intervalo
de 7 dias entre os testes, a atividade exploratória não é diferente entre os grupos, o
que significa que todos os grupos apresentaram semelhanças quanto o número de
entradas em cada um dos braços do aparato. Todavia, nos grupos que fizeram 7
109
dias de intervalo entre os testes foi observada redução no tempo em que os animais
do G8 permaneciam no centro do labirinto quando comparados ao G5 (64.40±8.31s
x 109.00±10.73s, p=0.046), como mostra a Figura 1B. A fins comparativos, fizemos
uma análise do desempenho entre todos os grupos, mesmo com intervalos
diferentes. O que encontramos foi que o G8 permanece menos tempo no centro do
LCE do que o grupo G4 (64.4±8.31 x 105.8±8.02, p=0.02) (Figura 3A), mesmo tendo
realizado a bateria de testes na mesma sequência, o intervalo entre eles também se
mostrou relevante para a realização dos testes.
Foi mensurado o tempo de latência para imobilidade no teste do nado forçado
(NF). Não foram encontradas diferenças entre nenhum dos grupos.
Já no teste de PCE, o G1 teve maior deslocamento no compartimento claro
do que os animais do G2 (67.80±3.8 x 49.60±3.9; p=0.02). E com uma semana de
intervalo, os grupos não apresentaram diferenças quanto ao deslocamento, mas sim
no tempo em que permaneceram em cada um dos compartimentos. O G5
permaneceu mais tempo no compartimento claro do que os grupos G6 e G8,
respectivamente (G5:184.64±9.4 x G6:138.80±7.3, p=0.008; G5 x G8:148.80±7.2,
p=0.04).
A temperatura registrada durante o NF chegou ao valor mínimo de
79.78+1.16% daquela em que o animal se encontrava antes do teste. Isso
representou uma queda de 7.41+0.48ºC. Esse mínimo foi alcançado 12.57+0.52
minutos após o fim do teste. Após 52.7+3.82 minutos do início do teste a
temperatura alcançou 97% do nível basal.
4. Discussão
A
ideia
inicial
deste
trabalho
surgiu
da
hipótese
de
que
testes
comportamentais realizados em sequência podem interferir nos resultados
apresentados, e dessa forma buscamos a melhor ordem na qual poderíamos realizar
a bateria proposta. Encontramos respostas comportamentais diversas de acordo
com a ordem executada.
110
O melhor teste para se iniciar a bateria de testes comportamentais é o campo
aberto, pois não foi observada nenhuma alteração nos resultados dos testes
realizados a posteriori deste. Já o teste do LCE mostrou-se ansiogênico, pois os
animais que primeiramente realizaram o LCE apresentaram ansiedade aumentada
quando o teste seguinte era o CA. A ansiedade neste grupo pode ser aferida pela
redução da exploração dos quadrantes centrais do aparato.
O teste do CA é baseado na propriedade inerente aos ratos chamada
tigmotaxia, que consiste na aversão a lugares abertos e expostos. Preferencialmente
os ratos ficam em cantos ou lugares entocados, pois como são presas de uma gama
de predadores, essa propriedade aumenta suas chances de sobrevivência. Quando
o animal tem sua ansiedade reduzida, expõe-se mais ao “perigo”, por outro lado,
quando a ansiedade está aumentada, como no caso do grupo 2, os animais
apresentam menor exploração da região central do CA. Esse resultado sugere que o
LCE não é uma boa escolha para iniciar-se a sequência de testes.
O resultado do teste PCE corrobora com esta afirmação uma vez que os
animais que iniciaram a bateria de testes pelo LCE apresentaram ansiedade
aumentada quando comparados aos animais do G1. O compartimento claro deste
aparato é aversivo para os ratos que são animais noturnos, o que significa que
quando a ansiedade está aumentada a tendência é a redução na exploração da
região mais aversiva do PCE.
Vimos que o teste do nado forçado causa efeito subsequente no teste LCE.
Os animais apresentam avaliação do risco reduzida no LCE quando precedido pelo
NF, pois permanecem menos tempo no centro do aparato avaliando o novo
compartimento a ser explorado. Portanto, este estudo sugere que teste do NF deve
ser executado após o teste do LCE.
Ao mesmo tempo o NF não deve ser o primeiro teste realizado na bateria
comportamental, pois mesmo após uma semana os animais apresentam redução no
tempo em que permanecem no centro do LCE avaliando o risco quando comparados
ao G5. Outra evidência para excluir o NF como primeiro teste da bateria é o
resultado encontrado no teste PCE, mesmo após 3 semanas os animais do G8
111
apresentam ansiedade aumentada neste teste quando comparados aos resultados
do G5, mensurada pelo tempo de permanência no compartimento claro deste
aparato.
Há disponível na literatura alguns estudos relatando o efeito de ordem em
camundongos, principalmente em testes de animais transgênicos (p.ex. Paylor et al.,
2006; McIlwain et al., 2001; Voikar et al., 2004). Em ratos, o primeiro estudo
realizado para testar o efeito de ordem numa bateria de testes comportamentais foi
recentemente publicado e apresentou um resultado bastante diferente do
encontrado por nosso grupo (Blokland et al., 2012). A mesma linhagem de ratos
(Wistar) foi utilizada, porém a metodologia foi bastante diferente. Blokland e
colaboradores (2012) encontraram efeito de ordem somente quando os animais
foram repetidamente expostos ao mesmo teste, o que sugere que o efeito de ordem
encontrado pode ter sido influenciado pela repetição no mesmo teste.
Neste estudo não realizamos treinamento prévio nos animais a fim de acessar
o desempenho dos animais na sua primeira exposição ao teste. Outra observação é
que nem todas as combinações possíveis foram testadas, portanto diferentes efeitos
de ordem podem ser encontrados.
O que ficou claro neste estudo é que o teste do NF é o mais estressor entre
eles, o que foi confirmado com o registro da temperatura. Porsolt e colaboradores
(1978) sugerem que a hipotermia registrada durante o nado forçado seja devido ao
stress causado pelo teste. Cinco minutos no teste é suficiente para elevar o nível de
corticosterona sérica, um marcador de situação estressante (Abel, 1993). Com isso
desaconselhamos a utilização desse teste antes de qualquer outro.
A execução dos testes comportamentais em forma de uma bateria de testes
permite a redução no tempo necessário de meses (dependendo do número de
testes) para semanas, poupando tempo e dinheiro, sem afetar na qualidade dos
resultados, além de possibilitar a execução de uma bateria de testes completa.
Reduzindo o intervalo entre os testes, também é reduzido o potencial para
problemas associados com o aumento na idade dos animais (Paylor et al., 2006).
112
Outra vantagem no uso da bateria de testes é a redução significativa no número de
animais utilizados.
Com os resultados obtidos no presente trabalho, podemos sugerir que é
importante determinar a ordem na qual serão realizados os testes para evitar o viés
nos resultados. Os nossos dados sugerem que os testes com características mais
aversivas e estressantes sejam os últimos. No caso da bateria de testes utilizada em
nosso estudo, sugerimos como ideal a sequência: CA – LCE – PCE – NF.
5. Referências
Abel EL. (1993). Physiological Correlates of the forced swim test in rats. Physiology &
Behavior 54:309-317.
Blokland A, Oever S, Gorp D, Draanen M, Schmidt T, Nguyen E, Krugliak A, Napoletano A,
Keuter S, Klinkenberg I (2012). The use of a test battery assessing affective behavior in
rats: Order effects. Behavioural Brain Research 228:16–21.
Chourbaji S, Brandwein C, Vogt MA, Dormann C, Mueller R, Drescher KU, Gross G, Gass P
(2008). Dopamine receptor 3 (D3) knockout mice show regular emotional behavior.
Pharmacological Research 58: 302–307.
Crawley JN, Paylor R (1997). A proposed test battery and constellations of specific
behavioral paradigms to investigate the behavioral phenotypes of transgenic and knockout
mice. Hormones Behavior 31:197–211.
Cryan JF, Holmes A (2005). The ascent of mouse: advances in modeling human depression
and anxiety. Nat Rev Drug Discov 4:775-790
Lad HV, Liu L, Paya-Cano JL, Parsons MJ, Kember R, Fernandes C, Schalkwyk LC (2010).
Behavioural battery testing: Evaluation and behavioural outcomes in 8 inbred mouse strains.
Physiology & Behavior 99:301–316.
McIlwain KL, Merriweather MY, Yuva-Paylor LA, Paylor R (2001). The use of behavioral test
batteries: effects of training history. Physiology & Behavior 73:705–717.
Ohl, F (2003). Testing for anxiety. Clin. Neurosci. Res. 3, 233-238.
Pandey DK, Yadav SK, Mahesh R, Rajkumar R (2009). Depression-like and anxiety-like
behavioural aftermaths of impact accelerated traumatic brain injury in rats: A model of
comorbid depression and anxiety? Behavioural Brain Research 205:436–442.
Paylor R, Spencer CM, Yuva-Paylor LA, Pieke-Dahl S (2006). The use of behavioral test
batteries, II: Effect of test interval. Physiology & Behavior 87: 95–102.
Porsolt RD, Anton G, Blavet N, Jalfre M (1978). Behavioral despair in rats: a new model
sensitive to antidepressant treatments. Eur J Pharmacol 47:379-391.
van der Staay FJ (2006). Animal models of behavioral dysfunctions: basic concepts and
classifications, and an evaluation strategy. Brain Res Rev. Aug 30;52(1):131-159. Review.
van Gaalen MM, Steckler T (2000). Behavioural analysis of four mouse strains in an anxiety
test battery. Behavioural Brain Research 115:95–106.
Voikar V, Vasar E, Rauvala H (2004). Behavioral alterations induced by repeated testing in
C57BL/6J and 129S2/Sv mice: implications for phenotyping screens. Genes Brain Behav
;3:27–38.
113
Anexo VI – Efeito da interação do ciclo claro-escuro de 22 horas e o
período endógeno na relação de fase entre os ritmos circadianos
de atividade/repouso e temperatura central
Autores: Bruno da Silva Brandão Gonçalves1,2, Breno Tercio Santos Carneiro1,
Bruna Del Vechio Koike1, João Miguel Gonçalves Ribeiro1,John Fontenele Araújo1
1 – Departamento de Fisiologia, Centro de Biociências, Universidade Federal do Rio
Grande do Norte –UFRN, Natal (RN), Brasil.
2 – Instituto Federal Sudeste de Minas Gerais – Campus Barbacena, Barbacena
(MG), Brasil
Periódico no qual será submetido: Chronobiology International
RESUMO
Ratos submetidos a ciclos claro-escuro com o período de 22h apresentam dois
componentes rítmicos com períodos diferentes (1320 e maior do que 1440 minutos)
na atividade locomotora. Sugere-se que cada um desses componentes seja gerado
por
uma
região
diferente
dos
núcleos
supraquiasmáticos.
Simulações
computacionais em conjunto com a análise de dados da atividade locomotora
mostraram que esses componentes podem ser gerados por um único oscilador
através de dois fenômenos: a modulação ou a superposição linear. Na superposição
linear ocorre um fenômeno conhecido como batimento, que é o aparecimento de
uma modulação lenta na amplitude da saída do oscilador. Um comportamento
semelhante a esta modulação foi encontrado no ritmo da atividade locomotora e de
secreção de melatonina. Aqui nós mostramos essa modulação através da análise da
114
variação na amplitude da temperatura central. Para isso comparamos o ritmo da
temperatura central de animais submetidos ao ciclo CE de 22 horas com o grupo
controle (expostos ao ciclo CE 24h). Além da variabilidade na amplitude da
temperatura decorrente da superposição linear, nós encontramos uma maior
instabilidade no ângulo de fase entre a atividade e a temperatura. Esses resultados
ajudam a discutir os efeitos da dessincronização interna que ocorre nos seres vivos,
permitindo interpretar que ultrapassar o limite da sincronização pode acarretar
diversas consequências para o organismo, provocando a quebra da ordem temporal
interna levando ao surgimento de doenças.
1. INTRODUÇÃO
Os ritmos circadianos são oscilações de aproximadamente 24 horas,
observadas desde o nível bioquímico até o cognitivo (Kondo et al., 1993; Campos et
al., 2001; Wright et al., 2012). Esses ritmos são controlados por um sistema de
temporização interno, e sincronizados por pistas ambientais (para revisão ver
Schibler et al., 2003). Entende-se que o sistema de sincronização circadiana em
mamíferos é composto de múltiplos osciladores arranjados de forma hierárquica,
sendo o núcleo supraquiasmático (NSQ) do hipotálamo seu oscilador central (Ralph
et. al., 1990, Schwartz, 2002).
Estudos recentes mostram que os NSQs apresentam pelo menos duas
regiões morfologicamente e funcionalmente distintas: região ventrolateral (vl-NSQ) e
região dorsomedial (dm-NSQ) (4). A região vl-NSQ recebe diretamente a informação
luminosa através do trato retino-hipotalâmico (RHT) (Moore e Lenn, 1972). Entre
essas regiões do NSQ, há uma densa projeção de vl para dm com pouca inervação
recíproca (Leak et. al., 1999), o que sugere que a região vl é sincronizada pelo ciclo
claro-escuro e ajusta a fase de dm (Miyake et. al.,2000).
Sob um regime de ciclo CE próximo ao limite inferior de sincronização, ratos
apresentam
dissociação
do
ritmo
de
atividade
locomotora
resultando
na
apresentação de dois componentes rítmicos: um deles sincronizado ao ciclo CE
imposto e outro não sincronizado que segue num período maior que 24h
115
(Campuzano e cols., 1998). A exposição dos animais a diferentes fotofases variando
o período entre 22 ou 23h gera uma gama de padrões de atividade locomotora
diferentes, desde o arrastamento total do ritmo (ajustado ao ciclo claro/escuro
imposto) até o ritmo em livre curso (expressão do componente endógeno), o que
indica que o processo de sincronização do sistema de temporização circadiano pode
ser gradualmente distribuído entre o componente circadiano dependente de luz e o
componente em livre curso, sugerindo que populações distintas de osciladores
podem estar em diferentes níveis de (des) acoplamento (Cambras et al, 2004). Este
mesmo efeito pode ser observado em outros ritmos circadianos, tais como o de
temperatura central e o de sono-vigília (Campuzano et. al., 1998; Vilaplana et. al.,
1997; Cambras et. al., 2007).
Em um trabalho elegante analisando expressão gênica de Per1 e Bmal1, de
la Iglesia et al. (2004) mostraram a relação entre a expressão dos dois componentes
rítmicos da atividade locomotora e as subdivisões do NSQ – vl e dm. O componente
sincronizado ao CE mostrou-se associado à expressão gênica da região vl-NSQ e o
componente com período maior que 24 h está associado à dm-NSQ. Utilizando um
modelo matemático de dois osciladores acoplados, Schwartz et al. (2009) mostraram
que o oscilador na região dm-NSQ apresenta uma coordenação relativa em ratos
mantidos em CE de 22 h. Além disso, mostraram que a liberação de melatonina
apresenta uma variação na sua amplitude que pode estar associada ao sinal de
saída da região dm-NSQ. Em outras palavras, a atividade ocorre quando o animal
deveria estar em repouso (misaligned phase). A estabilidade da relação de fase
entre diferentes ritmos circadianos está reduzida.
No presente trabalho, investigamos o comportamento da relação de fase
entre dois ritmos dependentes do NSQ, o ritmo de atividade-repouso e o ritmo da
temperatura central. Para isso utilizamos simulações computacionais e registros
experimentais de ratos mantidos em ciclo CE simétricos de 24 e 22 horas.
116
2. MATERIAL E MÉTODOS
2.1. Modelo computacional
O modelo utilizado nesse trabalho foi o descrito por Gonçalves et al. (2010)
que simula dois osciladores, cada um representando duas regiões do NSQ. O
oscilador vl-NSQ recebe a informação luminosa e sua saída é projetada para o
oscilador dm-NSQ. A saída utilizada para nossas análises será do oscilador dmNSQ que simula a região dorsomedial. O sistema foi submetido ao ciclo claro-escuro
(CE) de 24 horas (T24) e de 22 horas (T22) de forma semelhante aos experimentos.
Cada oscilador foi descrito pelo modelo de Goodwin (1965) composto por três
variáveis (X, Y e Z). Nesses osciladores a variável X representou a concentração de
um gene relógio fictício, Y a proteína produzida por X e Z um inibidor da produção de
X. No modelo adaptado por Gonçalves et al. (2010) a luz tinha o efeito de aumentar
a taxa de produção do gene relógio X. Os parâmetros foram ajustados para que os
períodos endógenos dos osciladores vl-NSQ e dm-NSQ fossem iguais a 24.25 e
24.4 horas respectivamente.
2.2. Procedimentos experimentais
Condições gerais
Foram utilizados 18 ratos Wistar (2-4 meses de idade) no estudo. Os animais
foram mantidos individualmente em gaiolas de polipropileno dentro de módulos de
isolamento com controle do ciclo CE e da temperatura (23 ± 1°C). Água e ração
(Labina, Purina®) foram disponibilizadas à vontade durante todos os experimentos.
A atividade motora foi registrada através de sensores infravermelhos e a
temperatura corporal através de sensores ibutton (Maxim Integrated Products, Inc.).
Todos os protocolos experimentais foram aprovados pelo Comitê de Ética para o
Uso de Animais da Universidade Federal do Rio Grande do Norte (projetos nºs.
026/2009, 021/2010, 038/2010). Este estudo foi realizado de acordo com os
princípios éticos da Sociedade Brasileira de Ciência em Animais de Laboratório
(SBCAL).
117
Procedimentos cirúrgicos
Os animais foram anestesiados através de injeções intraperitoneais de
cetamina (100 mg/kg) e xilazina (10 mg/kg). Os sensores de temperatura foram
inseridos na cavidade peritoneal através de uma incisão de aproximadamente um
cm. Após a cirurgia, foram administradas gentamicina (0,1 ml) e dexametasona (0,1
ml) e os animais permaneceram em recuperação por pelo menos 10 dias até o início
dos experimentos. Os mesmos procedimentos foram utilizados para retirar os
sensores após o fim dos experimentos.
Experimentos
Um grupo de 10 ratos foi sincronizado em CE 12:12 por 18 dias (grupo T24)
enquanto outro grupo de oito ratos foi submetido ao protocolo de dessincronização
forçada (T22), que consiste num ciclo CE 11:11 por 30 dias até apresentarem
dissociação do ritmo de atividade locomotora. Este grupo foi registrado por mais 100
dias consecutivos para registro e avaliação do ritmo em dessincronização forçada.
Análise de dados
Os dados de temperatura foram registrados durante 28 dias e para a análise
foram filtrados utilizando uma média móvel cuja janela foi de 10 horas. A amplitude
foi calculada como o valor máximo da temperatura subtraído pelo valor pelo mínimo.
Para cada animal foi calculado o coeficiente de variação (CV), definido como o
desvio padrão das amplitudes diárias da temperatura por sua média.
O CV é
calculado para cada animal a partir das N amplitudes. Sendo N o número de dias
registrados.
O registro completo da atividade locomotora em T22 foi utilizado para
calcularmos as frequências e períodos presentes no sinal. Para isso foi utilizado o
periodograma Sokolove-Bushel (Sokolove e Bushel, 1978). O cálculo da frequência
de batimento (fb) foi feito através da fórmula (f1-f2). Sendo f1 a frequência do
zeitgeber (1/79200 Hz) e f2 a frequência do ritmo em livre curso.
3. RESULTADOS
Uma inspeção visual dos actogramas e a geração de periodogramas a partir
do registro de atividade e temperatura indicam que todos os animais sincronizaram
118
ao ciclo CE de 24 horas (Figura 1). Todos os animais submetidos ao ciclo CE de 22
apresentaram dois componentes rítmicos de atividade locomotora:um com período
médio de 1320 min e o outro de 1500±10.13 min (Figura 2). Avaliando o ritmo
circadiano da temperatura, somente um animal apresentou dois componentes
rítmicos e sete animais apresentaram um único componente rítmico em livre-curso
com período de 1498.75±13.56.
Figura 1 – Gráficos representativos dos registros experimentais de atividade locomotora, temperatura
corporal e da simulação computacional em T24.
119
Figura 2 – Gráficos representativos dos registros experimentais de atividade locomotora, temperatura
corporal e da simulação computacional em T22.
A temperatura foi filtrada utilizando uma média móvel de 10 horas (figura 3A e
3B). Uma inspeção visual mostra que a variação na amplitude é menor em T24
quando comparado com T22 (figura 3). Esse resultado foi obtido nos registros
experimentais (figura 3A e 3B) e nas simulações computacionais (figuras 3C e 3D).
A partir desses valores calculamos o CV da amplitude da temperatura nas duas
condições de CE. Em T24 o CV foi igual a 0.1504±0.0454 e em T22 a média foi igual
a 0.2983±0.0423. O teste T mostrou que há uma diferença significante entre os dois
grupos (p < 0.001).
120
Figura 3. Registros experimentais da temperatura central filtrada pelo método de média móvel em
T24 (A) e em T22(B). Saída do oscilador dm-NSQ simulado em T24 (C) e em T22 (D). Tanto para os
dados experimentais e computacionais a variação na amplitude da temperatura é menor em T24 (A e
C) do que em T22 (B e D).
Calculamos o ângulo de fase entre as acrofases do ritmo circadiano da
temperatura central e do ritmo de atividade-repouso para cada período de 24 horas
para ambos os grupos. A média e desvio padrão do ângulo de fase foi igual a
1.21±0.36 h e 2.36±0.77 h para o ciclo CE de T24 e T22 respectivamente. O teste T
mostrou que há uma diferença significante entre os dois grupos (p<0.001). A partir
dos ângulos de fases dos animais calculamos a variância desses valores.
Encontramos que para T24 a média das variâncias foi de 2.50±1.28 h e para T22 foi
de 5.70± 2.34 h (p = 0.0019).
4. DISCUSSÃO
Neste trabalho, pudemos observar que ratos submetidos ao protocolo de
dessincronização forçada em ciclo CE simétrico de 22 horas, apresentam maior
variação na amplitude diária quando comparada aos animais expostos ao ciclo CE
121
de 24 horas, tanto do ritmo de atividade locomotora quanto do ritmo da temperatura
central. O mesmo resultado foi encontrado na simulação matemática do modelo.
Como acontece experimentalmente, em T24 ambos os osciladores se
sincronizaram ao estímulo externo confirmando a sincronização, ou o acoplamento,
entre as duas regiões dos NSQs sob o ciclo CE de 24 horas. Por outro lado, a
expressão de genes relógios durante o protocolo de dessincronização forçada (T22)
mostra que o oscilador vl-NSQ se sincroniza ao CE enquanto o dm-NSQ apresenta
um período semelhante aos animais em livre curso. Em nossas simulações, o
comportamento do oscilador vl-NSQ seguiu o achado experimental permanecendo
sincronizado a luz. Quanto à região dm-NSQ, os dois períodos encontrados na saída
deste oscilador não foram encontrados nos dados de la Iglesia et al. (2004) devido à
técnica utilizada. No referido trabalho, a expressão dos genes relógios foi analisada
em um único dia para cada animal. Essa amostragem impossibilita a detecção de
mais de um componente rítmico na expressão dos genes relógios no oscilador dmNSQ. Granada et al. (2011) mostraram que um único oscilador é capaz de gerar
diferentes componentes rítmicos e que isso deve ser uma explicação complementar
para a dessincronização forçada. Para os autores um oscilador pode apresentar dois
componentes espectrais independentes através da superposição linear de duas
ondas. A primeira seria referente ao período endógeno do oscilador e a segunda
dependente do ciclo CE. Se pensarmos que o dm-NSQ atua como principal saída do
NSQ, o padrão observado na simulação de T22 seria devido ao seu período
intrínseco somado a saída de vl-NSQ que está sincronizado ao ciclo CE de 22
horas.
A superposição linear de dois períodos gera um fenômeno chamado
batimento que é típico de osciladores no limite de sincronização (Granada et al.
2011). O batimento gera uma modulação lenta na saída do oscilador com uma
frequência igual à composição das frequências do zeitgeber e do oscilador. Os
valores encontrados para o período dessa modulação na atividade foi próximo ao
calculado, com um erro de 10+2%. A variação encontrada na amplitude da
temperatura em T22 pode ser explicada por esse efeito de batimento e também
ocorre no ritmo de secreção de melatonina (Schwartz et al., 2009). A compressão e
122
descompressão na amplitude da curva de secreção de melatonina persistem em
escuro constante. Com isso, é possível concluir que o ritmo de secreção de
melatonina é controlado pelo oscilador dm-NSQ, cuja saída no modelo apresentou
uma maior variação na amplitude. Assim foi descartada a hipótese de que a variação
na concentração de melatonina é somente dependente do mascaramento pela luz.
Figura 4. Influência do NSQ sobre os osciladores centrais e periféricos na construção dos ritmos
circadianos. Em T22 a saída da região dm-NSQ simulada é apresentada em (A) com uma variação
na amplitude tendo o seu máximo em 1 e o mínimo em 2. Essa variação altera a estabilidade na
sincronização que ocorre entre os múltiplos osciladores que formam o sistema de temporização
circadiano (B), sendo que em 1 a sincronização do ritmo do NSQ é maior do que em 2. Quando a
saída do dm-NSQ é baixa a diferença de fase entre os ritmos deve ser maior como demonstrado em
(C).
Através do nosso modelo computacional reproduzimos essas condições e
encontramos essa variação na amplitude da saída de dm-NSQ em T22. O modelo
computacional utilizado por Schwartz et al. (2009) apresentou resultados
semelhantes aos nossos e como ambos os modelos não simulam o efeito inibitório
da luz, reforçamos a ideia de que essa variação na amplitude dos ritmos tem uma
relação com a saída do oscilador. A alta variabilidade na amplitude da saída do NSQ
em T22 deve gerar uma instabilidade na fase dos ritmos circadianos como
temperatura central, ciclo sono-vigília, liberação de hormônios e outros, dificultando
a sincronização, como mostrado na figura 4. Acreditamos que quando a saída do
123
NSQ é reduzida há uma perda na capacidade desse marcapasso em sincronizar os
osciladores circadianos no regime CE de 22 horas (figura 4B). Da mesma forma a
maior estabilidade na relação de fase dos ritmos em T24 se deve a pouca variação
na amplitude da saída do NSQ, facilitando a sincronização do oscilador (a geração
dos ritmos) com os osciladores periféricos.
O sistema circadiano parece estar envolvido também na regulação de
processos cognitivos (Tapp e Holloway, 1981; Valentinuzzi et al, 2008; Craig e
McDonald, 2008; Loh et al., 2010). Especificamente, mostramos que os animais com
dissociação do ritmo circadiano apresentam déficits de memória em um tipo
específico de tarefa, a esquiva passiva (Neto et al., 2008). Talvez, a
dessincronização circadiana ou o desalinhamento entre ritmos circadianos, os seja,
a desordem temporal interna, prejudique funções cognitivas e também a saúde
numa forma geral, uma vez que várias evidências também mostram que a
dessincronização circadiana pode estar ligada ao desenvolvimento de doenças
(Martino et al., 2008; Preuss et al., 2008)
Consequências da alteração/dessincronização circadiana têm sido reportadas
também em seres humanos (Cho et al., 2000). Além disso, transtornos psiquiátricos,
por exemplo, apresentam uma série de sintomas relacionados a distúrbios de ritmo,
como a dessincronização do ciclo sono-vigília que é observada em praticamente
80% dos indivíduos acometidos (Maurizi et al., 2000). Os pacientes depressivos
apresentam um avanço da fase do sono e acarretam uma relação de fase anormal
com diversos outros ritmos, quase sempre resultando em um episódio depressivo
(Wehr et al., 1979). Esses estudos, entre outros, mostram a estreita relação entre o
sistema de temporização com graves transtornos como os psiquiátricos, ressaltando
assim a relevância da ritmicidade circadiana na saúde e no bom funcionamento do
organismo. Em alguns casos os seres humanos são submetidos a mudanças no
ciclo CE que os colocam em seu limite de sincronização como constantes jet-lag e
trabalho em turno por exemplo. As alterações fisiológicas que acontecem podem
estar relacionadas à variação na saída do oscilador principal e na instabilidade de
fase entre os ritmos circadianos.
124
REFERÊNCIAS
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
Albus H, Vansteensel MJ, Michel S et al. A GABAergic mechanism is necessary for coupling dissociable
ventral and dorsal regional oscillators within the circadian clock. Curr Biol 2005; 15:886–893.
Benedetti, F., Dallaspezia, S., Colombo, C., Pirovano, A., Marino, E., Smeraldi, E., 2008. A length
polymorphism in the circadian clock gene Per3 influences age at onset of bipolar disorder. Neurosci.Lett.
445, 184–187.
Benedetti, F., Serretti, A., Colombo, C., Barbini, B., Lorenzi, C., Campori, E., Smeraldi, E., 2003.
Influence of CLOCK gene polymorphism on circadian mood fluctuation and illness recurrence in bipolar
depression. Am. J. Med. Genet. B Neuropsychiatr. Genet. 123B, 23–26.
Cambras T, Chiesa J, Araujo J, Díez-Noguera A. Effects of photoperiod on rat motor activity rhythm at
the lower limit of entrainment. J Biol Rhythms.2004;19(3):216-25.
Cambras T, Weller JR, Anglès-Pujoràs M, Lee ML, Christopher A, Díez-Noguera A, Krueger JM, de la
Iglesia HO. Circadian desynchronization of core body temperature and sleep stages in the rat. Proc Natl
Acad Sci U S A. 2007;104(18):7634-9. Epub 2007 Apr 23.
Campos, T.F., Silva, S.B., Fernandes, M.G., Araujo, J.F., Menezes, A.A.L., 2001. Diurnal variation in a
visual-motor coordination test in healthy humans. Biological Rhythm Research, 32, 255-262.
Campuzano A, Vilaplana J, Cambras T, Dıez-Noguera A. Dissociation of the rat motor activity rhythm
under T cycles shorter than 24 hours. Physiol. Behav. 1998;63:171–176.
Cho K, Ennaceur A, Cole JC, Suh CK. Chronic jet lag produces cognitive deficits. J Neurosci, 2000; 20
(RC66):1-5.
Craig LA, McDonald RJ. Chronic disruption of circadian rhythms impairs hippocampal memory in the rat.
Brain Res Bull, 2008; 76: 141-51.
de la Iglesia HO, Cambras T, Schwartz WJ, Dı A. Forced Desynchronization of Dual Circadian
Oscillators within the Rat Suprachiasmatic Nucleus. Current. 2004;14(Ld):796-800.
Gonçalves BSB, Carneiro BTS, Silva CA, Fernandes DAC, Fortes FS, Ribeiro JMG, Cerqueira RC,
Rolim SAM, Araújo JF. Mathematical model of the interaction between the dorsal and ventral regions of
the suprachiasmatic nucleus of rats. Sleep Sci. 2010;3(1): 40–44.
Granada AE, Cambras T, Díez-Noguera A, Herzel H. Circadian desynchronization. Interface Focus.
2011;1(1):153-66. Epub 2010.
Kondo, T., Strayer, C.A., Kulkarni, R.D., Taylor, W., Ishiura, M., Golden, S.S. & Johnson, C.H. (1993).
Circadian rhythms in prokaryotes: Lucyferase as a reporter of circadian gene expression in
cyanobacteria. Proceedings of the National Academy of Sciences of the United States of America, 90,
5672-5676.
Lavebratt, C., Sjoholm, L.K., Soronen, P., Paunio, T., Vawter, M.P., Bunney, W.E., Adolfsson, R.,
Forsell, Y., Wu, J.C., Kelsoe, J.R., Partonen, T., Schalling, M., 2010. CRY2 is associated with
depression. PLoS One 5, e9407.
Leak RK, Card JP, and Moore RY. Suprachiasmatic pacemaker organization analyzed by viral
transynaptic transport. Brain Research 1999;819:23-32.
Lewy, A.J., Lefler, B.J., Emens, J.S., Bauer, V.K., 2006. The circadian basis of winter depression. Proc.
Natl. Acad. Sci. U. S. A. 103,7414–7419.
Loh DH, Navarro J, Hagopian A, Wang LM, Deboer T, Colwell CS. Rapid changes in the light/dark cycle
disrupt memory of conditioned fear in mice. Plos One, 2010; 5: e12546.
Mansour, H.A., Talkowski, M.E., Wood, J., Chowdari, K.V., McClain, L., Prasad, K., Montrose, D.,
Fagiolini, A., Friedman, E.S., Allen, M.H., Bowden, C.L., Calabrese, J., El-Mallakh, R.S., Escamilla, M.,
Faraone, S.V., Fossey, M.D., Gyulai, L., Loftis, J.M., Hauser, P., Ketter, T.A., Marangell, L.B., Miklowitz,
D.J., Nierenberg, A.A., Patel, J., Sachs, G.S., Sklar, P., Smoller, J.W., Laird, N., Keshavan, M., Thase,
M.E., Axelson, D., Birmaher, B., Lewis, D., Monk, T., Frank, E., Kupfer, D.J., Devlin, B., Nimgaonkar,
V.L., 2009. Association study of 21 circadian genes with bipolar I disorder, schizoaffective disorder, and
schizophrenia. Bipolar Disord. 11, 701–710.
Martino TA, Oudit GY, Herzenberg AM, Tata N, Koletar MM, Kabir GM, Belsham DD, Backx PH, Ralph
MR, Sole, MJ. Circadian rhythm disorganization produces profound cardiovascular and renal disease in
hamsters. . Am J Physiol Regul Integr Comp Physiol, 2008; 294: R1675-R1683.
Maurizi CP. A preliminary understanding of mania: roles for melatonin, vasotocin and rapid-eyemovement sleep. Med Hypotheses. 2000 Jan; 54(1): 26-9.
Mirmiran, J. Overdijk, W. Witting, A. Klop and D.F. Swaab. A simple method for recording and analysing
circadian rhythms in man. Journal ofNeuroscience Methods, 25 (1988) 209-214
Miyake S, Sumi Y, Yan L, Takekida S, Fukuyama T, Ishida Y, Yamaguchi S, Yagita K, Okamura H.
Phase-dependent responses of Per1 and Per2 genes to a light-stimulus in the suprachiasmatic nucleus
of the rat. Neurosci Lett. 2000;294(1):41-4.
125
23. Neto SPD, Carneiro BTS, Valentinuzzi VS, Araujo JF. Dissociation of the circadian rhythm of locomotor
activity in a 22 h light-dark cycle impairs passive avoidance but not object recognition memory in rats.
Physiol Behav, 2008; 94: 523-527.
24. Preuss F, Tang Y, Laposky AD, Arble D, Keshavarzian A, Turek FW. Adverse effects of chronic
circadian desynchronization in animals in a “challenging” environment. Am J Physiol Regul Integr Comp
Physiol, 2008; 295: R2034-R2040.
25. Ralph MR, Foster RG, Davis FC, Menaker M. Transplanted suprachiasmatic nucleus determines
circadian period. Science. 1990;247(4945):975-8.
26. Schibler U, Ripperger J, Brown SA. Peripheral circadian oscillators in mammals: time and food. J Biol
Rhythms. 2003;18(3):250-60
27. Schwartz MD, Wotus C, Liu T, et al. Dissociation of circadian and light inhibition of melatonin release
through forced desynchronization in the rat. Proceedings of the National Academy of Sciences of the
United States of America. 2009;106(41):17540-5.
28. Schwartz MD, Wotus C, Liu T, Friesen WO, Borjigin J, Oda GA, de la Iglesia HO. Dissociation of
circadian and light inhibition of melatonin release through forced desynchronization in the rat. Proc Natl
Acad Sci U S A 2009;1-6.
29. Schwartz WJ. Suprachiasmatic nucleus. Curr Biol. 2002;12(19):R644.
30. Sokolove PG, Bushell WN. The chi square periodogram: its utility for analysis of circadian rhythms. J
Theor Biol. 1978;72(1):131-60.
31. Soria, V., Martinez-Amoros, E., Escaramis, G., Valero, J., Perez- Egea, R., Garcia, C., Gutierrez-Zotes,
A., Puigdemont, D., Bayes, M., Crespo, J.M., Martorell, L., Vilella, E., Labad, A., Vallejo, J., Perez, V.,
Menchon, J.M., Estivill, X., Gratacos, M., Urretavizcaya, M., 2010. Differential association of circadian
genes with mood disorders: CRY1 and NPAS2 are associated with unipolar major depression and
CLOCK and VIP with bipolar disorder. Neuropsychopharmacology 35, 1279–1289.
32. Tapp WN, Holloway, FA. Phase shifting circadian rhythms produces retrograde amnesia. Science, 1981;
211: 1056-8.
33. Valentinuzzi VS, Neto SPD, Carneiro BTS, Santana KS, Araujo JF, Ralph MR. Memory for time of
training modulates performance on a place conditioning task in marmosets. Neurobiol Learn Memory,
2008; 89: 604-607.
34. Vilaplana J, Cambras T, Campuzano A, Díez-Noguera A. Simultaneous manifestation of free-running
and entrained rhythms in the rat motor activity explained by a multioscillatory system. Chronobiol Int.
1997;14(1):9-18.
35. Wehr TA, Wirz-Justice A, Goodwin FK, Duncan W, Gillin JC. Phase advance of the circadian sleep-wake
cycle as an antidepressant. Science 1979;206(4419):710-3.
36. Wright KP, Lowry CA, LeBourgeois MK. Circadian and wakefulness-sleep modulation of cognition in
humans. Front Mol Neurosci, 2012; 5: 50.
126