BIOLOGIA – módulo 02
GENÉTI CA PÓS-MENDEL
A partir de 1900, com a descoberta dos trabalhos de Mendel, a
genética ganhou novo rumo. Novos experimentos eram feitos
com outras espécies de animais e plantas reproduzindo as
bases hereditárias deixadas por Mendel. Outros casos de
monoibridismo e diibridismo serão aqui relacionados com suas
peculiaridades, mas sempre relacionados às leis de Mendel.
Cabe a você distinguir as diferenças relacionadas a cada caso,
e memorizar o maior número possível de exemplos das novas
heranças.
Um outro exemplo está na anormalidade da proteína
hemoglobina dos indivíduos portadores ou acometidos pela
anemia falciforme ou sinclemia.
Portador
HbS HbA
Tipos de
hemoglobina
presentes
SeA
Anemia
falciforme
HbS HbS
S
Normal
HbA HbA
A
Fenótipo
Genótipo
Sangue
ambas
1. Dominância intermediária, Semidominância,
Dominância incompleta, ou Ausência de dominância.
Em 1906 o inglês William Bateson (1861-1926) fez
experimentos com a planta Mirabilis jalapa cuja flor
assemelha-se com a nossa “graxa”. Cruzando parentais de
flores vermelhas e brancas entre si Bateson obteve uma
geração F1 de plantas de flores róseas. Este tipo de fenótipo
intermediário entre os descendentes F1 não tinha sido relatado
por Mendel nos híbridos de ervilhas. Autocruzando a F1, o
cientista obteve uma progênie F2 com plantas de flores
brancas, vermelhas e róseas em proporções genotípicas
esperadas matematicamente.
3. Alelos múltiplos ou Polialelia.
A origem de variantes alélicas em um mesmo locus gênico é
provavelmente oriunda das mutações. A mutação altera a
seqüência de bases nitrogenadas de um gene ocasionando sua
incapacidade completa ou sua alteração culminando com uma
proteína diferenciada da proteína anterior. Nas variantes
alélicas de Mendel, só existiam duas possíveis para construção
de um par, ou seja, para o locus cor da semente, só existiam
os genes V e v, para textura, só R e r. Na polialelia, a mutação
origina outras variantes alélicas que podem ocupar o mesmo
locus gênico.
PELAGEM DE COELHOS
A pelagem de coelhos é condicionada pela combinação de
quatro variantes alélicas para o mesmo locus. Os fenótipos
produzidos são o selvagem ou aguti de coloração castanhoacinzentada, o chinchila de pelagem cinzento-prateada, o
himalaia de pelagem branca mais com extremidades negras, e
o albino totalmente despigmentado (branco) inclusive com
olhos vermelhos.
Assim como nos estudos mendelianos, a integridade dos alelos
foi mantida ao longo das gerações. Apenas nos híbridos ou
heterozigotos, nenhum dos alelos é dominante sobre o outro.
Ambos expressa-se em uma dosagem parcial.
Este tipo de herança caracteriza-se pela alteração nas
proporções fenotípicas, que passam a ser iguais às
proporções genotípicas na geração F2: 1 vermelha: 2 róseas:
1 branca. Vale lembrar: como não há dominância entre os
alelos, as letras do par não podem ser diferenciadas. Neste
caso, escolhe-se uma outra letra para associar os pares de
genes.
Ex:
Flor branca = IBIB; flor rósea = IBIV; flor vermelha = IVIV.
2. Coodominância.
Na coodominância, o indivíduo heterozigoto manifesta a dose
completa da característica de cada um dos genes presentes no
par. O exemplo mais comum está na tipagem sanguínea pelo
sistema ABO; o indivíduo sangue tipo AB possui os alelos IA e
IB e manifesta a proteína de ambos.
As mutações ocasionaram três variantes alélicas do gene C
que condiciona a pelagem selvagem. Estas variantes
apresentam uma relação de dominância como no esquema
abaixo:
C > cch > ch > ca
Na tabela abaixo vamos relacionar cada genótipo ao seu
respectivo fenótipo para nível de cruzamentos.
1
BIOLOGIA – módulo 02
Genótipos
CC, Ccch, Cch, Cca
cchcch, cchch, cchca
ch ch , ch ca
caca
Fenótipos
Selvagem
Chinchila
Himalaia
Albino
SISTEMA SANGUÍNEO ABO
O mérito da descoberta dos diferentes fenótipos para o tipo
sanguíneo é sem dúvida do austríaco Karl Landsteiner (18681943). Antes dele, as transfusões de sangue feitas não
passavam de expectativa. Algumas davam certo e o paciente
sobrevivia e outras não obtinham sucesso ocasionando, na
maioria das vezes, a morte do receptor. Usando macacos e
amostras de sangue humano, Landsteiner decifrou o enigma
das transfusões. Para uma transfusão com sucesso, além de
obedecer às combinações do sistema ABO, o sistema Rh (visto
a seguir) tem de ser obedecido.
Fenótipos
Genótipos
Aglutinogênio
ou antígeno
Aglutinina ou
anticorpo
A
IAIA, IAi
Proteína A
Anti – B
B
IBIB, IBi
Proteína B
Anti – A
AB
IAIB
Proteínas A e B
Ausentes
O
ii
Ausentes
Anti – A e Anti – B
A partir da tabela acima podemos esquematizar as possíveis
doações que podem ser feitas entre os diferentes tipos
sanguíneos.
Note que o sangue tipo O, por não possuir antígenos nas
hemácias, pode ser doado para cada um dos tipos sem
problemas de aglutinação (doador universal), enquanto o
2
Existem três variantes alélicas para o mesmo locus no sistema
ABO: IA, IB e i. Há uma relação de dominância completa entre
os alelos IA e IB em relação ao alelo i. Porém, há uma relação
de coodominância entre os alelos IA e IB. Quem possui
genótipo contendo um dos genes coodominantes acima possui
uma proteína na membrana dos glóbulos vermelhos do sangue
(hemácias) denominada de aglutinogênio ou aglutinógeno.
Esta proteína comporta-se como antígeno em um sangue
qualquer. Caso estas hemácias encontrem-se em um tipo de
sangue diferente do seu, haverá um anticorpo ou aglutinina
no plasma esperando para promover a reação antígenoanticorpo que ocasionará a aglutinação das hemácias
formando grumos ou pequenos coágulos dentro do sangue.
Isto pode promover o entupimento de capilares sanguíneos
levando o receptor da amostra sanguínea à morte.
O quadro abaixo representa os fenótipos possíveis, os
genótipos de cada fenótipo correspondente, e ainda os
aglutinogênios e aglutininas acompanhados da caracterização
esquemática de ambos com as possíveis aglutinações.
Hemácias
Plasma
Aglutinação
sem anticorpos
não oferece aglutinação
sangue tipo AB, por não possuir anticorpos em seu plasma,
pode receber de todos os outros tipos (receptor universal).
Para diagnosticar qual tipo de sangue você possui, basta
realizar um teste simples como abaixo:
BIOLOGIA – módulo 02
Os resultados possíveis são dados pela aglutinação diferencial
da amostra de sangue na lâmina.
Como na segunda gravidez o número de anticorpos é ainda
pequeno, a criança tem chance de nascer e se recuperar.
Porem, a cada novo parto, o perigo aumenta.
Fator Rh
O fator Rh também foi descoberto por Landsteiner em Macaca
rhesus. Este fator também está presente em hemácias
humanas e pode ocasionar problemas nas transfusões
sanguíneas. Veja o quadro de fenótipos, genótipos,
aglutinogênios e aglutininas abaixo:
Fenótipos
Genótipos Aglutinogênios Aglutininas
Rh+
DD, Dd
Proteína D
Ausente
Anti – D
Rhdd
Ausente
(caso seja
sensibilizado)
A diagnose para o fator Rh pode ser feita juntamente com a
da tipagem do sistema ABO.
Os problemas com eritroblastose podem ser evitados com
exames pré-nupciais e administração de anticorpos sintéticos
logo após as primeiras 72h pós-parto, destruindo hemácias
fetais pra que a mãe não seja sensibilizada em cada gravidez.
4. Interação Gênica.
Este tipo de interação refere-se as relações existentes entre
pares de genes não alelos. A genética mendeliana, a princípio,
não teve problemas com interações pois, na 2ª Lei, os pares
de genes não alelos se segregavam independentemente entre
si exibindo proporções fenotípicas na F2 de 9:3:3:1.
Para diferenciarmos o diibridismo mendeliano com a interação
gênica é preciso explicar melhor os tipos de interações
existentes entre os diferentes pares de genes.
Os resultados são:
As doações são possíveis apenas de indivíduos Rh- para Rh+.
Ao contrário, haverá desenvolvimento de anticorpos anti – D,
ocorrendo aglutinação das hemácias.
4.1 Interação não epistásica: é aquela onde a
proporção típica mendeliana é mantida inalterada (9:3:3:1),
embora os fenótipos sejam sempre mais diversificados.
Ex:
Genes complementares para forma da crista de galinhas:
Após diversos cruzamentos experimentais o inglês W. Bateson
concluiu que a forma da crista das galinhas era determinada
por dois pares de genes que se interagiam entre si. Ao cruzar
duas galinhas puras com cristas ervilha e rosa, o cientista
alcançou uma geração F1 com crista inteiramente diferenciada
chamada de crista noz. Cruzando entre si indivíduos da
geração F1, Bateson obteve a proporção fenotípica mendeliana
para cruzamentos diíbridos.
ERITROBLASTOSE FETAL OU DHNR (Doença Hemolítica do
Recém-Nascido).
A eritroblastose fetal é uma incompatibilidade sanguínea
materno-fetal. Ela só acontece quando a mãe é Rh- e o pai é
Rh+. Mesmo o pai sendo Rh+, ainda há uma chance de ser
heterozigoto (Dd) e ter apenas 50% de chance de ter filhos
com a doença.
A doença ocorre porque a mãe, durante a primeira gravidez
recebe algumas hemácias Rh+ do filho acidentalmente
durante o parto. Sendo sensibilizada, apenas na segunda
gestação é que os problemas poderão acontecer. A mãe
sensibilizada constrói anticorpos anti-D no seu plasma. Como
os anticorpos maternos têm livre acesso pela barreira
placentária, o feto terá destruídas todas as hemácias com
proteína Rh+ que ele fabricar, ainda na vida intra-uterina.
Caso nasça vivo, o bebê apresentará icterícia (pigmentação
amarelada nas mucosas) devido à hemoglobina liberada pelas
hemácias destruídas.
3
BIOLOGIA – módulo 02
A interação entre os genes dominantes E da crista ervilha e R
da crista rosa dão o fenótipo noz. Na geração F2 a presença
de apenas um gene dominante em cada par, ao mesmo
tempo, dará crista noz, a presença de apenas um dominante
em apenas um par, dará crista ervilha ou rosa, e a ausência
de um gene dominante entre os dois pares formará a crista
simples, um outro fenótipo.
Reveja as interações no quadro abaixo:
Exemplo de epistasia recessiva:
Neste caso, um dos pares de genes, quando em homozigose
recessiva, inibem a manifestação do caráter do outro par
independentemente de sua combinação.
Nas raças de cães labradores, existem três tipos de cor na
pelagem. Os cães podem ter pelagem preta ou marrom ou
dourada. O gene C condiciona a pigmentação da pelagem e é
dominante sobre o seu alelo c que inibe a manifestação da cor
apresentando pelagem dourada. Num outro par de genes, o
gene M determina pêlos pretos e é dominante sobre seu alelo
m que condiciona a cor de pêlos marrons.
4.2 Interação epistásica: é aquela onde a
proporção fenotípica do diibridismo memdeliano é alterada.
O termo epistático é dado a qualquer gene ou par de genes
que iniba ou empeça a manifestação da característica de um
outro par de genes não alelos. O par de genes inibido passa a
ser chamado de hipostático.
A tabela abaixo apresenta os principais tipos de interações
epistásica entre pares de genes não alelos.
Tipo de interação
Proporção clássica
Tipo de interação
Proporção clássica
Epistasia dominante
Epistasia recessiva
Duplo-recessiva
Interação dominante-recessiva
Dupla-dominante
A_B_
9
A_B_
9
9
9
Genótipos
A_bb
aaB_
3
3
aabb
1
Genótipos
A_bb
aaB_
aabb
3
3
1
12
3
1
3
4
7
13
3
15
1
A seguir, vamos exemplificar apenas alguns tipos de
interações epistásica.
Exemplo de epistasia dominante:
Ocorre quando um alelo dominante de um dos pares é
epistásico sobre a manifestação do outro par.
Em cavalos, o gene W inibe a manifestação de cor e é
dominante sobre seu alelo w. O gene B determina pêlos
pretos é dominante sobre seu alelo b que condiciona pêlos de
cor marrom. Sendo o gene W epistático sobre os genes B e b,
quando ele ocorre no genótipo, a pelagem do animal é branca.
4
Exemplo de epistasia duplo-recessiva:
Este tipo de herança envolve a inibição de uma característica
quando quaisquer dos pares de genes encontram-se em
homozigose recessiva.
Na espécie humana, a surdez hereditária é ocasionada pela
presença em homozigose dos alelos d e e que são epistáticos
em relação aos genes alelos D e E; estando ambos presentes,
estes genes condicionam fenótipo normal.
4.3
Herança
quantitativa,
multifatorial, aditiva ou cumulativa.
poligênica,
A herança das características até agora estudadas retrata
fenótipos opostos e estáticos. Citando como exemplo a altura
da planta de ervilha estudada por Mendel, só existiam plantas
altas e plantas baixas. Não há referência à fenótipos
intermediários entre elas. Este tipo de herança é chamada de
descontínua. Porém, é possível existir um tipo de herança
onde haja fenótipos intermediários entre os extremos obtidos,
levando ao aparecimento contínuo ou gradativo da expressão
fenotípica. Exemplos disto podem ser visualizados na nossa
própria espécie como cor de olhos, altura, cor de pele,
inteligência etc.
BIOLOGIA – módulo 02
Na herança quantitativa os pares de genes que interagem
entre si possuem uma relação diferente da dominância e da
recessividade já estudadas. Os genes representados por letras
maiúsculas serão chamados de efetivos, ou seja, eles
adicionam sua dose ao fenótipo. Quando representados por
letras minúsculas, ou genes não-efetivos, estes genes não
acrescentarão nada ao fenótipo final do indivíduo.
A representação gráfica da freqüência dos fenótipos para a
herança quantitativa é a curva de distribuição normal, em
forma de sino.
5. Herança Sexual ou Ligada ao sexo.
Relembrando um pouco o cariótipo humano já estudado no
módulo anterior, podemos constatar a presença, na espécie
humana, de 23 pares de cromossomos homólogos. Dentre
estes pares, 22 deles, idênticos entre homens e mulheres são
denominados de cromossomos autossômicos. O último par é
formado pelos cromossomos sexuais cuja combinação é
diferente entre homens (heterogaméticos - XY) e mulheres
(homogaméticas - XX); são denominados de cromossomos
alossômicos ou sexuais. A determinação do sexo do
indivíduo é sempre dada pela presença no espermatozóide de
um cromossomo X ou Y, pois o óvulo materno só apresenta
um cromossomo X sempre para formar o par sexual.
Exemplo de herança quantitativa:
A cor da pele humana parece ser determinada pela presença
de no mínimo dois pares de genes não alelos. Caracterizando
estes genes como N, n, B e b, poderíamos promover um
cruzamento imaginário entre homem e mulher híbridos para
estudar a possível freqüência fenotípica da geração F2.
Os gametas formados por cada um dos indivíduos do casal
levando em conta a segregação independente seria NB, Nb,
nB e nb.
Analisando o quadrado de Punnét na página seguinte, a
proporção mendeliana típica é, em grande parte, alterada.
Para obter o fenótipo de cada indivíduo esperado na
população ideal de 16 possibilidades, você deve observar o
número de genes efetivos e não-efetivos que cada genótipo
possui. Assim, a proporção fenotípica esperada será como no
quadro a seguir:
Genótipos
NNBB
NNBb, NnBB
NNbb, nnBB, NnBb
Nnbb, nnBb
nnbb
Fenótipos
Negro
Mulato escuro
Mulato médio
Mulato claro
Branco
Os insetos dípteros (duas asas) como as moscas apresentam
cariótipo diferente dos seres humanos, mas sua determinação
sexual é idêntica. Por ter ciclo de vida curto, a “mosquinhadas-frutas” ou Drosophila melanogaster (2n = 8) é um dos
animais mais estudados pela genética desde o início do século
passado.
Proporção fenotípica
1:16
4:16
6:16
4:16
1:16
Atenção! existem espécies de animais que possuem
determinação sexual diferente do homem e dos insetos
dípteros. Algumas aves, insetos lepidópteros e répteis
apresentam uma inversão na determinação sexual.
Nestes grupos, a fêmea é heterogamética (ZW) e o
macho é homogamético (ZZ). Sendo assim, é a fêmea
que determina o sexo de sua prole
5
BIOLOGIA – módulo 02
Herança dos cromossomos sexuais:
As características até então estudadas encontravam-se
distribuídas entre pares de cromossomos homólogos
autossômicos. Na herança sexual vamos reconhecer o
comportamento de alguns caracteres cujos genes estão
localizados em partes dos cromossomos sexuais. O primeiro
ponto a ser analisado é a posição deste gene, já que os
cromossomos sexuais, no caso do homem, não apresentam
homologia completa quando comparados aos da mulher.
No emparelhamento dos cromossomos sexuais masculinos,
podem se distinguir três tipos básicos de comportamento dos
genes:
Região homóloga entre X e Y: os genes posicionados nesta
região comportam-se como todos os genes até então
estudados. Eles se segregam independentemente um do outro
durante a meiose.
Região do cromossomo Y não homóloga a X: estes genes
representam características jamais presentes no sexo
feminino. A maior parte destes genes (holândricos) está ligada
a fatores de determinação testicular.
Região do cromossomo X não homóloga a Y: os genes
encontrados nesta região comportam-se diferentemente entre
os dois sexos; isto porque, no homem há apenas um gene e
não um par (hemizigoto). Na mulher, a presença de dois
cromossomos X inteiramente homólogos, garante a formação
dos pares, podendo ser homozigota ou heterozigota.
As heranças para os tipos de genes citados acima são
comumente chamadas, respectivamente, de herança
parcialmente ligada ao sexo, herança restrita ao sexo e
herança ligada ao sexo.
Entre os genes da herança ligada ao sexo, estão: o responsável
pela distrofia muscular do tipo Duchenne, o gene responsável
pela hemofilia e o daltonismo, e os genes responsáveis pela
cor dos olhos em drosófilas. A maior parte da herança para
estas características comporta-se como recessiva.
Veja alguns genótipos e fenótipos abaixo:
Mulheres
Genótipo
Fenótipo
XDXD
Normal
Normal
D d
X X
portadora
XdXd
Daltônica
Homens
Genótipo
Fenótipo
XDY
Normal
XdY
Daltônico
Por ser hemizigoto, o homem tem apenas uma chance em
duas de não apresentar a anomalia. As mulheres têm o dobro
de chance de não apresentar a doença. A freqüência de
homens com daltonismo (incapacidade de destinguir cores
como o vermelho e o verde) é de 1/2500, enquanto, nas
mulheres, é de 1/5000.
Veja o exemplo do um cruzamento a seguir:
6
Em drosófilas, fêmeas e machos selvagens possuem olhos
vermelhos e são portadores de pelo menos um alelo w+. O
alelo que determina a cor branca dos olhos é representado
apenas pela letra w.
Fêmeas
Genótipo
Fenótipo
Xw+Xw+
Selvagem
Selvagem
Xw+Xw
portadora
XwXw
Branco
Machos
Genótipo
Fenótipo
Xw+Y
Selvagem
XwY
Branco
6. Ligação fatorial, Vinculação gênica ou “Linkage”.
A partir de 1910, um cientista chamado Thomas Hunt Morgan
(1866-1945), trabalhando com drosófilas selvagens e
mutantes, descobriu um tipo de herança bastante diferente da
segregação independente dos caracteres postulada na 2ª lei
de Mendel.
Cruzando drosófilas puras para cor do corpo e tamanho da asa
Morgan obteve uma progênie homogênea de moscas híbridas
para dois caracteres. Em drosófilas, a cor cinza do corpo,
condicionada pelo alelo P, é dominante sobre a cor preta
presente apenas em homozigotos para o gene p. A forma da
asa longa ou normal é determinada pelo gene V e é
dominante sobre seu alelo v responsável, em homozigose,
pela asa vestigial. Fazendo um cruzamento-teste ou “back
cross” o qual consiste em cruzar indivíduos com características
dominantes com indivíduos duplo-recessivos, Morgan obteve
um comportamento diferenciado entre os sexos ao retrocruzar
a geração F1. Machos híbridos cruzados com fêmeas duplorecessivas originaram indivíduos com corpo cinza e asa normal
(50%) e indivíduos com corpo preto e asa vestigial (50%).
Duas das classes fenotípicas da segregação mendeliana (corpo
cinza e asa vestigial e corpo preto e asa normal) estavam
ausentes na descendência.
Ao retrocruzar fêmeas da geração F1 com machos duplorecessivos, Morgan obteve as quatro classes fenotípicas
esperadas, porém não obedeciam a estatística mendeliana.
41,5% cinza de asas normais;
41,5% pretos de asas vestigiais;
8,5% cinza de asas vestigiais; e
8,5% pretos de asas normais.
Como os indivíduos do cruzamento-teste duplo-recessivos só
produzem um tipo de gameta pv, a conclusão de Morgan foi
de que machos da F1 podiam produzir apenas gametas PV e
pv em proporções iguais, e fêmeas podiam produzir gametas
PV (41,5%), pv (41,5%), Pv (8,5%) e pV (8,5%) distribuídos
em proporções diferentes.
BIOLOGIA – módulo 02
Os gametas ausentes nos machos não aconteciam porque os
genes P e V estavam em “linkage” ou vinculados, ou seja, no
mesmo cromossomo.
A partir das novas descobertas de Morgan sobre a segregação
de mais de 85 mutações entre os 4 pares de cromossomos de
drosófila, foi possível estabelecer um mapeamento dos genes
nos cromossomos pelos percentuais de permutação existentes
entre genes ligados.
Faltava explicar ainda o motivo da segregação existente na
formação dos gametas das fêmeas da F1 e o porque da
freqüência diferenciada entre eles.
Em 1909, o citologista F. Janssens (1863-1924) confirmou as
suposições de Morgan sobre o “crossing-over” ou permutação
gênica entre cromátides homólogas (Prófase I). A permuta de
genes entre cromátides permitiria que genes ligados
pudessem ser trocados gerando os gametas diferenciados.
Porém, como na figura a seguir, se houvesse “crossing” em
todos os gametas formados, o número de indivíduos da
progênie seria homogêneo entre recombinantes e parentais.
Mais tarde com outros experimentos Morgan associou a soma
dos permutantes como a distância entre os genes no mesmo
cromossomo. Quanto mais distantes fossem os genes entre si,
maior seria a freqüência de permuta entre cromátides
homólogas. No exemplo anterior, a soma das combinações
novas (8,5 + 8,5 = 17%) representaria a distância entre os
genes no cromossomo; 17 unidades de mapa ou
morganídeos, em homenagem a Morgan.
Outra constatação do cientista foi a de que machos de
drosófilas jamais fazem “crossing” ou permuta.
Freqüência de
permutação
=
Soma das freqüências das
classes recombinantes
Freqüência de
permutação
=
distância entre os genes ligados
no mesmo cromossomo
MUTAÇÕES
O termo mutação tem o mesmo significado da palavra
mudança. Qualquer modificação na molécula de DNA, por
mais simples que seja, é caracterizada como uma mutação. As
mutações podem ser caracterizadas em dois grandes grupos:
as gênicas, que ocorrem na estrutura de um gene e envolve
a substituição, adição ou perda de bases nitrogenadas, e as
cromossômicas que estão relacionadas a mudanças no
número ou na estrutura de um ou mais cromossomos de uma
espécie.
Embora as mutações sejam quase sempre sinônimas de
aberrações, deficiências ou má formação de proteínas e
estruturas protéicas, elas também representam fonte de
variabilidade primária para objeto da seleção natural como
veremos no módulo posterior. Um exemplo disto está em volta
de nós mesmos quando observamos indivíduos da nossa
própria espécie; embora tenhamos, na maioria, estruturas
iguais como braços, pernas, cabelos etc, são de infinitas
formas e, profundamente diferenciadas de nós.
Mutações gênicas:
Ao longo de nosso estudo, nos deparamos com muitas das
prováveis mutações desta natureza. Com as ervilhas de
Mendel, para cada característica, existe um par de genes e de
qualidades diferentes (Ex: V e v, R e r). Na herança dos alelos
múltiplos, mais de um par eram variantes para um mesmo
locus (Ex: IA, IB e i).
7
BIOLOGIA – módulo 02
As mutações gênicas podem ser fruto da ineficiência das
estruturas de reparo da molécula de DNA, mas também
podem ser induzidas através de agentes mutagênicos como
substâncias químicas (íons, pesticidas, radicais livres etc.) ou
agentes físicos como as radiações ultra-violeta. Entre os
exemplos de modificações gênicas está a anemia falciforme,
produto da substituição no gene da hemoglobina de uma
única base (A=T por T=A) modificando um aminoácido (ácido
glutâmico por valina) e alterando a forma da proteína final e
também das hemácias.
Atenção! As mutações gênicas podem acontecer nas
células somáticas e nas células germinativas. Porém, só as
mutações das células germinativas poderão ser passadas
para a próxima geração.
Mutações cromossômicas ou aberrações:
Este tipo de mutação possui uma classificação para as
diferentes modalidades de alterações cromossômicas. Veja na
tabela e na figura abaixo:
Mutações estruturais
deficiência ou deleção
duplicação
inversão
translocação
Mutações numéricas
Aneuploidias
Euploidias
A deleção é a perda de um pedaço de um cromossomo.
Como existem vários genes em um único cromossomo, as
mutações neste nível, são sempre mais radicais e perigosas
quando comparadas com as mutações gênicas.
A duplicação é quando o produto da deleção de um
cromossomo une-se ao outro cromossomo do mesmo par.
Como os genes entre os homólogos são para as mesmas
características, este cromossomo recebedor do pedaço, está
duplicado.
A inversão trata-se da fragmentação do cromossomo, e a
conseguinte soldagem do fragmento em sentido invertido.
A translocação é quando o produto da deleção de um par de
homólogos une-se a um outro par diferente.
Uma translocação entre cromossomos de ancestrais humanos
parece ter sido responsável pela origem de seres humanos e
chimpanzés, nossos parentes mais próximos.
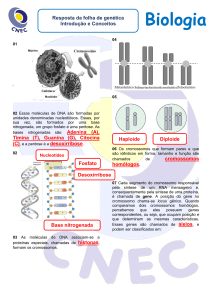
As mutações
classificações.
Euploidias
Aneuploidias
numéricas
também
Tipo de aberração
Haploidia
Triploidia
Tetraploidia
Nulissomia
Monossomia
Trissomia
Tetrassomia
possuem
grupos
de
Nº de cromossomos
n
3n
4n
2n – 2
2n – 1
2n + 1
2n + 2
Entre as aneuploidias humanas mais conhecidas está a
trissomia do cromossomo 21, ou síndrome de Down (2n=47),
a trissomia no par sexual ou síndrome de Klinefelter (2n=47),
e a monossomia no par sexual ou síndrome de Turner
(2n=45).
*****
8