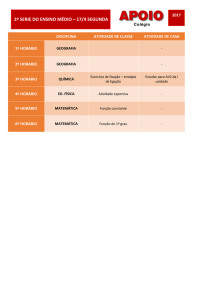
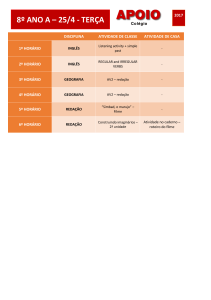
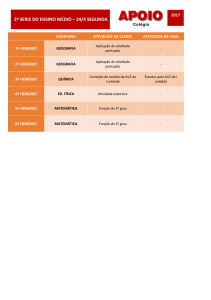
Bioquimica Fisiológica
Metabolismo de purinas e pirimidinas
Metabolismo de
purinas e pirimidinas
Introdução
Bases purínicas e pirimidínicas são compostos aromáticos heterocíclicos,
dos quais derivam os nucleótidos e nucleósidos.
Principais funções dos nucleótidos purínicos e pirimidínicos:
reacções de transferência do grupo fosfato dos NTPs
síntese de hidratos de carbono → UDP-glicose e UDP-galactose
síntese lipídica → CDP-acilglicerol
intermediários de alta energia para síntese de ligações covalentes
fazem parte de coenzimas como FAD, NAD, NADP, CoA, Sadenosilmetionina
funções reguladoras → efectores alostéricos
mensageiros secundários (cAMP, cGMP)
precursores de ácidos nucleicos e outros compostos
Estas bases absorvem radiação UV → luz UV é mutagénico.
As bases púrinicas e pirimidinas provêm da biossíntese e da dieta.
O metabolismo destas bases está associado ao ácido úrico e à gota.
O ácido úrico é insolúvel na urina ácida, assim os metabolitos xantina e
ácido úrico podem ocorrer em calculos urinários.
Composição e Estrutura das Purinas e Pirimidinas
Purinas e pirimidinas são compostos heterocíclicos, ou seja, são compostos
com anéis (cíclicos), com átomos de carbono e outros átomos (hetero),
em especial azoto.
Costumam ligar-se a uma pentose (açucar), ligada a um grupo NH por
uma ligação N-glicosídica.
Podem conter um ou mais grupos fosfato esterificados a um grupo OH da
pentose.
São moléculas essencialmente planas.
As mais importantes bases heterocliclicas dos ácidos nucleicos são as
purinas adenina e guanina e as pirimidinas citosina, timina (no DNA) e
uracilo (RNA)
Página 1 de 14
Bioquimica Fisiológica
Metabolismo de purinas e pirimidinas
Nomenclatura
São usados muitos sistemas de numeração para purinas e pirimidinas
As Pirimidinas possuem anéis menores (6 carbonos) e as purinas possuem
anéis maiores (9 carbonos).
Os átomos são numerados no sentido dos ponteiros do relógio na
pirimidinas e em sentido retrógrado nas purinas.
Nucleótidos
A maioria das purinas e pirimidinas existem na célula como nucleótidos
Nucleósidos = purina/pirimidina + açucar cíclico (D-ribose ou 2-desoxi-Dribose) com ligação -N-glicosídica entre o N-1 da pirimidina ou o N-9 da
purina.
Nucleótidos = nucleósidos fosforilados (mono, di ou tri)
Conformações
N-glicósidos heterocíclicos formam conformações Sin e Anti
Após se formar a base heterocíclica, não existe liberdade de rotação na
ligação N-glicosídica que une a base à pentose.
Deste modo, nucleótidos são estáveis, não interconvertíveis nas
conformações sin e anti → só são interconvertíveis pela ruptura e reformação na ligação glicosídica.
Ambas as conformações existem na natureza, mas a conformação anti
predomina, e participa na formação da dupla cadeia do DNA.
Bases secundárias ou incomuns
Ácidos nucleicos contém bases minor ou pouco usuais
5-metil-citosina → DNA
N6-metiladenina, N6,N6-dimetiladenina, N6,N7-dimetiladenina → mRNA
Vários no tRNA
Estas bases pouco usuais no DNA e RNA têm funções importantes no
reconhecimento de oligonucleótidos e regulação da meia-vida do RNA.
Página 2 de 14
Bioquimica Fisiológica
Metabolismo de purinas e pirimidinas
Funções dos nucleótidos
1. NTP têm elevado potencial de transferência de grupos (ácido anídrico) →
fornecem energia a reacções de síntese
a) Derivados de adenosina:
ADP e ATP estão envolvidos na fosforilação oxidativa, e o ATP é o
maior transdutor biológico de energia livre (é muito abundante).
O AMPc é mensageiro secundário.
O adenosina 3-fosfato-5-fosfossulfato é um dador de grupos sulfato
para formação de proteoglicanos ou metabolitos urinários de
drogas.
S-adenosilmetionina é metionina activa, serve de dador de grupos
metil e fonte de propilamina para síntese de poliaminas.
b) Derivados de guanina:
Participa na conversão de succinil-CoA a succinato
GTP é activador da adenilato ciclase, regulador alostérico e fonte
de energia para síntese proteica.
GMPc é sinal intracelular ou 2º mensageiro
c) Derivados de hipoxantina:
IMP é um percursor dos nucleótidos purínicos, por desaminação do
AMP
Desfosforilação do IMP forma inosina, essencial para a recuperação
de purinas
d) Derivados de uracilo:
Derivados de UDP participam na epimerização de açúcares ex.:
glicose 1P → galactose 1P, com UDP-glicose
UDP-açúcar é dador de açúcar para a síntese de oligossacáridos de
glicoproteínas e proteoglicanos
UDP-glicuronato é dador de glicoronato para a conjugação da
bilirrubina e de drogas como a aspirina
e) Derivados de citosina:
CTP é necessária para a síntese de alguns fosfoglicéridos como a
esfingomielina.
2. Algumas coenzimas são derivados de nucleótidos
Nota: Podem ser usados nucleótidos análogos sintéticos para quimioterapia
Página 3 de 14
Bioquimica Fisiológica
Metabolismo de purinas e pirimidinas
Biossíntese dos nucleótidos púrinicos
A biossíntese de bases purinicas e pirimidinas é estritramente regulada,
dependendo da necessidade fisiológicas dos nucleótidos.
O fígado é o principal órgão da biossíntese dos nucleótidos purínicos.
Existe uma dependência parcial (cérebro - devido aos niveis baixos de
PRPP aminotransferase) ou exclusiva (leucócitos polimorfonucleares e
eritrócitos - devido à incapacidade de sintetizar 5-fosforibosilamina)
As reacções de síntese e degradação dos nucleótidos purínicos decorrem
no citosol.
A formação do PRPP (5-fosforibosil-1-pirofosfato) a partir de ribose-5fosfato, exige a dependência da via das fosfopentoses
O PRPP é intermediário comum na biossíntese de purinas e pirimidinas e
reacções de recuperação das purinas.
Existem 2 mecanismos de biossíntese dos nucleótidos purínicos:
1. Sintese“de novo”, a partir de intermediários anfibólicos;
2. Recuperação das purinas: fosforibosilação das purinas;
fosforilação de nucleósidos purínicos.
Síntese “de novo”
Requer a formação do anel de purina a partir de fragmentos de N e C de
aminoácidos (glicina, aspartato, glutamato), tetrahidrofolato (N5, N10metenil e N10 formil) e CO2.
11 reacções em sequência sintetizam o 1º produto da via: inosina 5’monofosfato (IMP)
O IMP é o precursor de dois mononucleótidos purínicos: AMP (adenosina
5’-monofosfato); GMP (guanosina 5’-monofosfato).
Formação de:
1. PRPP: 1º intermediário formado na síntese de novo das purinas e
intermediário na via de recuperação de purinas.
2. fosforibosilamina (N9 do anel de purina pela PRPP aminotransferase);
3. 5’fosforibosil-glicinamida (C4, C5 e N7, pela fosforibosil-glicinamida
sintetase);
4. 5’fosforibosil-formilglicinamida (C8, por formil transferase);
5. 5’-fosforibosil-formilglicinamidina (N3, por sintetase);
6. 5’-fosforibosil-5-amino-imidazol (encerramento do anel imidazol, por
sintetase);
7. 5’-fosforibosil
5-aminoimidazol-4-carboxilato
(C6,
por
carboxilase
independente da biotina);
8. 5’-fosforibosil 5-aminoimidazol 4-N succinocarboxamida (N1, por sintetase,
que incorpora aspartato);
9. 5’-fosforibosil-5-aminoimidazol-4-carboxamida (separação do fumarato,
por adenil-succinase);
10. 5’-fosforibosil-5-formamidoimidazol-4-carboxamida
(C2,
por
formil
transferase);
11. IMP (por encerramento do anel, por inosinicase).
Página 4 de 14
Bioquimica Fisiológica
Metabolismo de purinas e pirimidinas
Ribose 5P
Sintase
ATP
AMP
5-Fosforribosil 1-Pirofosfato (PRPP)
PRPP aminotransferase
Glutamina
Glutamato + PPi
5-Fosforribosil amina
Sintase
Glicina + ATP
ADP + Pi
5-Fosforribosil glicinamida
Formiltransferase
N10 Formil H4 Folato
H4 Folato
5-Fosforribosil formil glicinamida
Sintase
Glutamina + ATP
Glutamato + ADP + Pi
5-Fosforribosil formil glicinamidina
Sintase
ATP
ADP + Pi
5-Fosforribosil 5-aminoimidazol
Carboxilase
CO2
5-Fosforribosil 5-aminoimidazol 4-carboxilato
Sintase
Aspartato + ATP
ADP + Pi
5-Fosforribosil 5-aminoimidazol 4-succinocarboxamida
Adenil Succinase
Fumarato
5-Fosforribosil 5-aminoimidazol 4-carboxamida
Formiltransferase
N10 Formil H4 Folato
H4 Folato
5-Fosforribosil 5-formaminoimidazol 4-carboxamida
Inosinicase
NADH + H+ NAD+
Xantosina 5-monofosfato
GMP sintase
Glutamina + ATP
Glutamato + AMP + PPi
Guanosina 5-monofosfato
Nucleósido
cinase
GDP, GTP
IMP DH
H2
Aspartato + GTP
Inosina 5-monofosfato
GDP + Pi
AdenilSuccinato
Fumarato
Adenil Succinase
Adenosina 5-Monofosfato
Nucleósido
cinase
ADP,
ATP14
Página
5 de
Bioquimica Fisiológica
Metabolismo de purinas e pirimidinas
Notas:
Gasto de 6mol de ATP por mol de IMP formado
O tetrahidrofolato é transportador de Formil
As enzimas desta via estão organizadas em complexos – Proteinas
Multifuncionais.
Drogas antifolato ou análogos da glutamina bloqueiam a biossíntese de
nucleótidos púrinicos
Conversão do IMP em AMP e GMP
O IMP depois de formado pode seguir duas vias diferentes: sintese de AMP
ou GMP.
Ambos os processos dependem de energia: ATP para a biossíntese do
GMP; GTP, para a do AMP.
Transformação do IMP em AMP:
formação de adenil-succinato, por sintetase, através da adição de
aspartato
formação do AMP, por adenil-succinase, libertando fumarato
Transformação do IMP em GMP
formação da xantosina 5’-monofosfato, XMP, por desidrogenase NAD+
dependente
formação de GMP, por sintetase.
A transferência do grupo fosforil do ATP converte mononucleótidos a
nucleósidos di- e trifosfatos, catalisada 1º pela monofosfato nucleósido cinase
e depois pela difostato nucleósido cinase.
Recuperação das purinas
As reacções de recuperação requerem muito menos energia que a
sintese “de novo”.
A origem das bases pode ser exógena (da dieta) ou endógena (de ácidos
nucleicos degradados)
1. Fosforibosilação:
–
Envolve a fosforibosilação de uma purina livre por PRPP, formando purina
5-mononucleótido
A fosforibosilação da guanina em GMP e da hipoxantina em IMP é
catalisada por fosforibosiltransferases comuns: hipoxantina-guanina
fosforribosiltransferase (regulação por IMP ou GMP)
A fosforribosilação da adenina em AMP por fosforibosiltransferase
específica: adenina fosforibosiltransferase (regulação pelo AMP).
PRPP PPi
Adenina
Adenosina 5-mononucleótido
Adenina fosforribosil transferase
Página 6 de 14
Bioquimica Fisiológica
Metabolismo de purinas e pirimidinas
PRPP PPi
Hipoxantina
Inosina 5-mononucleótido
PRPP PPi
Guanina
Guanosina 5-mononucleótido
Hipoxantina-guanina fosforribosil transferase
2. Fosforilação:
A fosforilação de nucleósidos purínicos pelo ATP, envolve a adenosina
cinase (adenosia AMP ou deoxiadenosina dAMP) e a deoxicitidina
cinase (deoxicitidina CMP; deoxiadenosina dAMP; deoxiguanosina
dGMP)
ATP ADP
desoxiGuanosina
desoxiGuanosina-mononucleótido
ATP ADP
desoxiAdenosina
desoxiAdenosina mononucleótido
ATP ADP
desoxiCitidina
desoxiCitidina mononucleótido
Desoxicitdina cinase
ATP ADP
Adenosina
Adenosina-mononucleótido
ATP ADP
desoxiadenosina
Desoxiadenosina 5-mononucleótido
Adenosina cinase
Página 7 de 14
Bioquimica Fisiológica
Metabolismo de purinas e pirimidinas
Regulação da síntese dos nucleótidos purínicos
O PRPP é o determinante da velocidade da sintese “de novo” daas
purinas.
A síntese do PRPP depende da actividade da sintetase do PRPP e da
disponibilidade de ribose-5-fosfato.
A PRPP sintetase é sensível à concentração de fosfato e às concentrações
de ribonucleótidos de purina que actuam como reguladores alostéricos.
A principal etapa reguladora é a catalisada pela aminotransferase da
fosforibosilamina, que é sensivel à inibição competitiva por nucleótidos
púrinicos (AMP e GMP).
A inibição da PRPP aminotransferase, transforma esta enzima num dímero
inactivo. A libertação do dímero, activa a enzima.
O AMP e o GMP regulam a sua própia formação a partir do IMP, por
retroinibição.
A formação do AMP é regulada a nível da IMP desidrogenase (inibição
competitiva pelo GMP) e formação do GMP é controlada na adenilsuccinato sintetase (inibição competitiva pelo AMP).
5-Fosforribosil 1-Pirofosfato (PRPP)
PRPP aminotransferase
Guanosina 5-monofosfato
Inosina 5-monofosfato
Adenosina 5-Monofosfato
Adenilsuccinato sintase
IMP DH
GTP
ATP
Transformação de nucleósidos púrinicos em
desoxiribonucleósidos
A redução, no carbono 2 dos ribonucleótidos púrinicos é catalizada pelo
complexo
da
nucleótido-redutase,
formando
difosfatos
de
desoxiribonucleósidos.
A enzima só é activa quando a célula sintetiza activamente DNA, em
preparação para a divisão celular.
A redução requer tio-redoxina, redutase e NADPH.
Ribonucleósido-DP
NDP redutase
Tioredoxina reduzida
Tioredoxina oxidada
NADP+
Redutase
NADPH + H+
Desoxirribonucleósido-DP
Página 8 de 14
Bioquimica Fisiológica
Metabolismo de purinas e pirimidinas
Catabolismo dos nucleósidos purínicos
Os nucleósidos púrinicos são convertidos a ácido úrico
Catabolismo dos nucleósidos por nucleotidases;
A adenosina é desaminada a inosina, por desaminase
Ocorre fosforólise de ligações N-glicosidicas da inosina e guanosina,
catalisada pela purina nucleósido fosforilase, formando hipoxantina e
guanina.
A hipoxantina e guanina formam xantina pela xantina oxidase e guanase,
respectivamente,
A xantina é oxidada a ácido úrico (pela xantina-oxidase).
O ácido úrico é um ácido fraco, logo as proporções não dissociadas e a
sua base conjugada (urato) dependem do pH.
A dissociação ocorre a pH = 5,8, logo existe urato na urina.
Os uratos são muito mais solúveis em água do que o ácido úrico.
O ácido úrico torna-se a forma predominante na urina normal, assim, os
cristais do tracto urinário são de urato de sódio, em qualquel local próximo
do sítio de acidificação do tubulo distal e nos ductos colectores, mas o
ácido úrico estará presente em qualque local distante destes sítios.
A formação de cálculos pode ser reduzida através da alcalinização da
urina.
Adenosina
H2 O
NH4+
Desaminase
Guanosina
Inosina
Pi
Ribose 1P
Hipoxantina
H2O + O2
H2O2
Pi
Purinanucleósido fosforilase
Ribose 1P
Guanina
Xantina
oxidase
Guanase
NH3
Xantina
H2O + O2
H2O2
Ácido Urico
Página 9 de 14
Bioquimica Fisiológica
Metabolismo de purinas e pirimidinas
Defeitos metabólicos
Hiperuricémia: os nivés séricos de urato de sódio excedem o limite de
solubilidade.
Cristais de urato de sódio podem ser formados nos tecido moles e nas
articulações o que pode levar a processos inflamatórios.
Doença
Gota
Síndrome de
Lesch-Nyahn
Imuno-deficiência
Imuno-deficiência
Litiase Renal
Xantinúria
Enzima Deficiente
PRPP sintetase
ou
Hipoxantina-guanina
fosforribosiltransferase
(parcial)
Hipoxantina-guanina
fosforribosiltransferase
(completa)
Adenosina desaminase
Purina nucleósido
fosforilase
Adenina fosforribosil
transferase
Xantina Oxidase
Caracteristicas
Padrão de
Herança
Recessivo
ligado ao X
superprodução e excreção
acentuada de purina
superprodução e excreção
acentuada de purina; paralesia
cerebral e automutilação
Imunodeficiência combinada
(células T e célula B),
desoxiadenosinúria
Deficiância das células T,
inosinúria, desoxiguanosinúria,
hipouricémia
Litiase renal de 2,8diidroxiadenina
Litiase renal da xantina,
hipouricémia
Recessivo
ligado ao X
Autossómica
recessiva
Autossómica
recessiva
Autossómica
recessiva
Autossómica
recessiva
Página 10 de 14
Bioquimica Fisiológica
Metabolismo de purinas e pirimidinas
Biossíntese dos nucleótidos pirimidinicos
Síntese “de novo”
A síntese “de novo” de núcleósidos purinicos e pirimidinicos envolve vários
percursores comuns: PRPP, glutamina, CO2, aspartato e derivados de
H4folato (para os derivados de Tiamina)
Uma diferença entre estes 2 processos é que enquanto a ribose fosfato é
uma parte integrante da molécula precursora inicial na síntese de
nucleótidos de purina, a ligação do grupo ribose fosfato ao N-3 da base
pirimidina ocorre as últimas etapas da biossíntese.
1. Formação do carbamoil-fosfato (C2, N3- carbamoil sintetase II)
2. Condensação com aspartato formando carbamoil aspartato (N1, C4, C5,
C6 - transcarbamilase ou carbamoil-transferase)
3. Encerramento do anel (dihidro-orotase)
4. Formação de orotato (desidrogenase de dihidro-orotato, mitocondrial;
todos as restantes etapas decorrem no citosol)
5. Fosforibosilação (com PRPP) do orotato, originando a orotidina monofosfato
6. Descarboxilação em UMP (uridina monofosfato)
7. Transformação em UDP e UTP (por fosforilação com ATP)
8. Aminação do UTP em CTP (com a glutamina e ATP) pela sintetase do CTP.
9. A transformação de nucleósidos em desoxiribonucleósidos pirimidínicos é
idêntica ao das purinas, por complexo da redutase da tio-redoxina;.
10. Formação do dUTP pela aceitação de um fosfato do ATP pelo dUMP
11. Transformação do dUMP em TMP.
Glutamina + CO2
ATP
Carbamoil
Sintase II
Glutamato + ADP
Carbamoil-P
Aspartato
Aspartato carbamoil
sintase
Pi
Carbamoil Aspartato
Dihidroorotase
Dihidroorotato
NAD+
DH
NADH + H+
Orotato
Fosforribosil
Transferase
PRPP
PPi
Orotidina monofosfato
ADP
ATP
Descarboxilase
UTP
Sintase
CTP
CO2
Tioredoxina redutase
Uridina monofosfato
Glutamina + ATP
dUMP
N5N10metileno H4Folato
Glutamato + ADP + Pi
NADH + H+
NAD+
Timidilato
sintase
H2Folato
Página 11 de 14
dTMP
Bioquimica Fisiológica
Metabolismo de purinas e pirimidinas
Recuperação das pirimidinas
Existem reacções de recuperação que convertem os ribonucleósidos de
pirimidina (uridina e citidina) e desoxirribonucleósidos (timidina e
desoxicitidina) aos seus respectivos nucleótidos.
As reacções de recuperação incluem a fosforibosilação das pirimidinas
(por fosforibosiltransferase) e fosforilação (por cinases).
A oroato fosforibosiltransferase é capaz de poupar o oroato, convertendoo a OMP.
1. Fosforribosilação
PRPP PPi
Uracilo
Uridina 5-mononucleótido
PRPP PPi
Tiamina
Timidilato 5-mononucleótido
Fosforribosil transferases
2. Fosforilação:
ATP ADP
Uridina
Uridina-mononucleótido
ATP ADP
Citidina
Citidina mononucleótido
Uridina-citidina cinase
ATP ADP
Timidina
Timidilato mononucleótido
Timidina cinase
ATP ADP
desoxiCitidina
desoxiCitidina mononucleótido
dCitidina cinase
ATP ADP
CMP
CDP
ATP ADP
UMP
UDP
ATP ADP
dCMP
dCDP
Pirimidina nucleótido monofosfocinase
ATP ADP
dTMP
dTDP
Timidilato cinase
Página 12 de 14
Bioquimica Fisiológica
Metabolismo de purinas e pirimidinas
Regulação da biossíntese de pirimidinas
A regulação é a nível alostérico e genético.
As 2 primeiras enzimas do processo biossíntético são reguladas
alostericamente.
A nivel genético são controladas as 3 primeiras e 2 últimas enzimas do
processo, através de mecanismos de repressão e desrepressão
coordenadas.
A actividade da carbamoil-sintetase II é inibida pela UTP, mas activada
por PRPP
A aspartato transcarbamoilase é inibida por CTP e activada por ATP.
A OMP descarboxilase é regulada pelo UMP e CMP
A sintetase do CTP é inibida pelo CTP.
Glutamina + CO2
Carbamoil Sintase II
PRPP
Carbamoil-P
Aspartato carbamoil sintase
Carbamoil Aspartato
Orotidina monofosfato
Descarboxilase
Uridina monofosfato
UTP
Sintase
CTP
Página 13 de 14
Bioquimica Fisiológica
Metabolismo de purinas e pirimidinas
Catabolismo dos nucleótidos pirimidinicos
Os produtos finais do catabolismo das pirimidinas são altamente
hidrossolúveis: CO2, NH3, β-aminoisobutirato e β-alanina.
A excreção de β-aminoisobutirato aumenta na leucémia e apóes a
exposição a raios X, devido a uma aumentada destruição de DNA.
Defeitos metabólicos
Sindrome de Reye: é acompanhado por acíduria orótica,
incapacidade da mitocôndria severamente danificada, de
carbamoil-fosfato que fica disponível para a superprodução de
citosol.
Acidúria Orótica I: deficiência na oroato fosforibosiltransferase e
descarboxilase.
Acidúria Orótica II: deficiência na orotidilato descarboxilase.
Citosina
Uracilo
devido à
utilizar o
oroato no
orotidilato
Timina
NADPH + H+
NADP +
Dihidrouracilo
β-alanina
Dihidrotimina
CO2, NH4+
β-aminobutirato
Página 14 de 14