Fisiologia de Sistemas
Mestrado Integrado em
Engenharia Biomédica
1º Semestre 2010/2011
2º Ano
Resumos por:
Tiago Silva e Sofia Faria
Página 1
Índice
Orgãos dos Sentidos – Introdução ……………………………………………………… 3
Sistema Nervoso………………………………………………………………………………… 5
Audição e Equilíbrio……………………………………………………………………………. 20
Visão………………………………………………………………………………………………….. 24
Os sentidos químicos: olfacto e paladar…………………………………………….. 27
Sistema Nervoso Autónomo………………………………………………………………. 30
Coração……………………………………………………………………………………………... 37
Sistema cardiovascular e circulatório ………………………………………………… 43
Electrocardiograma …………………………………………………………………………… 52
Sistema Respiratório …………………………………………………………………………. 55
Sangue ……………………………………………………………………………………………… 63
Equilíbrio Ácido-Base ………………………………………………………………………… 69
Sistema Renal ……………………………………………………………………………………. 71
Sistema Digestivo ………………………………………………………………………………. 80
Osso – Sistema Locomotor ……………………………………………………………….. 95
Agradecimentos ………………………………………………………………………………… 101
Página 2
Aula 1 – Orgãos dos Sentidos (Introdução)
Sentidos são as redes de informação (comunicação), entre o “mundo” exterior e o “mundo”
interior com o SNC. São fundamentais para a coordenação das diferentes funções
intercelulares e orgânicas. A complexidade destes órgãos é a do SNC.
A sua comunicação pode ser feita por um sistema:
Hormonal – lento, impreciso e genérico;
Neuronal (sistema eléctrico-quimico) – preciso, rápido e especifico;
Transdução
TERMO GENÉRICO, PARA O PROCESSO NO QUAL UMA FORMA DE ENERGIA É CONVERTIDA
NOUTRA (tipos de energia incidente possíveis no organismo: térmica, electromagnética,
mecânica, química).
A necessidade de transdutores no organismo advém do facto do
SNC só processar informação baseada em sinais electro-químicos e
é essencial que todo o tipo de informação seja codificado nesta
forma de informação.
Esta codificação vem da capacidade de diferentes transdutores
transformarem uma forma de energia como pressão arterial,
odores, sons, fotões, num sinal electro-químico de modo que
possa ser encaminhada e posteriormente processada pelo SNC.
Receptores sensoriais
Um receptor sensorial é a estrutura que reconhece um estímulo no ambiente interno ou
externo de um organismo.
Cada receptor é altamente sensível a um determinado estímulo (para aquele que foi criado) e
praticamente não responde a qualquer outro tipo de estímulos sensitivos (especificidade).
Uma vez activado, um receptor inicia os potenciais de acção na fibra sensitiva associada que
conduz estes estímulos até à medula espinal como uma linha marcada através de um nervo
periférico. Estes impulsos são semelhantes em todas as fibras sensitivas. Estas podem
manifestar diferenças qualitativas de amplitude ou frequência mas um potencial de acção
iniciado por um estímulo doloroso não é distinguível de qualquer outro iniciado por outro
receptor sensitivo ou modalidade sensitiva. O que nos permite diferenciar o tipo de sensação é
localizar de onde vêm e terminam as fibras do sistema nervoso.
Página 3
Todos os receptores sensoriais têm uma característica em comum. Qualquer que seja o tipo de
estímulo que excite o receptor, o seu efeito imediato é mudar o potencial eléctrico da
membrana. A isto chama-se um potencial receptor.
Tipos de receptores sensoriais:
Primário – Recebe o estímulo exterior e processa-o, transformando-o num potencial
de acção;
Secundário – Transmite o potencial de acção vindo do receptor primário;
Classificação:
1. Mecanoreceptores: detectam compressões ou estiramentos mecânicos do receptor
ou dos tecidos adjacentes.
2. Termoreceptores: detectam variações na temperatura, detectando alguns o frio e
outros o calor.
3. Nocireceptores: detectores de dor, detectando danos ocorridos em tecidos, quer
químicos, quer físicos.
4. Receptores electromagnéticos (Fotorreceptor): detectam luz na retina do olho.
5. Quimioreceptores: detectam variações químicas no corpo.
1 – Mecanoreceptores – 3 tipos:
Tacto: sensação do toque, pressão e vibração (podem ser terminações nervosas livres,
raízes pilosas, discos tácteis, corpúsculos tácteis, lamelados e de Ruffini);
Baroreceptores: detectam alterações de pressão nas paredes dos vasos sanguíneos e
tractos digestivos, reprodutivo e urinário;
Proprioreceptores: monitorizam a posição das articulações e músculos
2 – Termoreceptores: tem terminações nervosas livres localizadas na dermis, músculos
esqueléticos, fígado e hipotálamo. São conduzidas nas mesmas vias que conduzem dor.
5 – Quimioreceptores: podem estar localizados nos corpos carotídeos (na origem das carótidas
internas) e nos corpos aórticos (no arco aórtico). Monitorizam pH, níveis de CO2 e O2 no
sangue arterial.
Todos os receptores podem interpretar dor se forem demasiado estimulados!
Página 4
E quanto ao tipo de localização:
Extero-receptores – sensíveis a estímulos vindos de fora do corpo (toque, pressão,
dor, sentidos especiais)
Intero-receptores / (viscero-receptores) – respondem a estímulos de dentro do corpo
(vísceras) (alterações químicas, estiramento dos tecidos, temperatura)
Próprio-receptores – respondem a estímulos internos e localizam-se apenas nos
músculos esqueléticos, tendões, articulações e ligamentos.
Aula 2 – Sistema Nervoso motor e sensitivo
O sistema nervoso é constituído pelo sistema nervoso central (encéfalo e medula) e o sistema
nervoso periférico (pares cranianos e nervos espinhais).
Tecido Nervoso
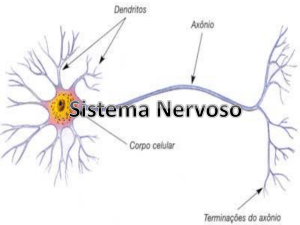
Neurónio: unidade funcional do sistema nervoso. Apresenta um corpo celular,
prolongamentos designados dendritos (receptores dos estímulos nervosos) e axónios
(responsáveis pela condução dos estímulos electro-químicos que partem do corpo celular) que
estabelecem ligações com outras células por intermédios de sinapses.
Células Gliais: unidades de sustentação, revestimento e defesa dos neurónios.
Os corpos dos neurónios agrupam-se à superfície, constituindo o córtex cerebral e em algumas
regiões da profundidade dos hemisférios, os núcleos cinzentos da base, e constituem a
substância cinzenta.
A substância branca, por seu lado, engloba os prolongamentos celulares dos neurónios,
envolvidos por células da glia.
Quando um neurónio entra em actividade alterando a carga eléctrica da parede celular há
progressão do impulso eléctrico até aos prolongamentos e à sinapse levando à libertação de
mediadores químicos que vão estimular a célula adjacente actuando num receptor que gera
um impulso eléctrico e faz progredir o estímulo. Uma característica especial destas sinapses é
que são unidireccionais, forçando o sinal a deslocar-se nas direcções requeridas, executando
funções nervosas específicas.
Sistema Nervoso Central:
Níveis principais das funções do SNC
O sistema nervoso humano herdou capacidades funcionais especiais do seu desenvolvimento
evolutivo. Deste modo, três níveis principais do SNC têm características funcionais específicas:
a) Nível da medula espinhal
Ao contrário do que se pensa, a medula espinhal é mais do que uma condutora de sinais da
periferia para o cérebro e vice-versa.
Página 5
Circuitos neuronais na medula podem causar:
1. Movimentos para andamento
2. Actos reflexos
3. Reflexos que alteram a posição das pernas para suportar o corpo
4. Reflexos que controlam vasos sanguíneos locais, movimentos gastrointestinais e
excreção urinária.
b) Nível subcortical (parte inferior do cérebro)
Grande parte do que se chamam actividades subconscientes do corpo é controlada nas áreas
inferiores do cérebro. Por exemplo, o controlo subconsciente da pressão arterial e respiração é
atingido principalmente no tronco cerebral. Controlo do equilíbrio é uma função combinada
do cerebelo com outras duas áreas inferiores do cérebro. Reflexos alimentares como a
salivação (tronco cerebral, amígdala e hipotálamo) e também comportamentos emocionais.
c) Nível cortical (parte superior do cérebro)
É um local de armazenamento de memória. O córtex cerebral nunca funciona sozinho mas
sempre em associação com centros inferiores do sistema nervoso. Sem o córtex cerebral, as
funções dos centros inferiores são imprecisas. O vasto armazenamento de informação cortical
converte essas funções imprecisas em funções precisas e determinadas.
Notas:
Nas variáveis fisiológicas existe um ponto de equilíbrio, sendo o sistema nervoso o
responsável pela sua manutenção, controlando os restantes sistemas (por feedback
negativo ou positivo), mantendo as suas funções normais.
O SN actua em conjunto com o sistema endócrino (que actua por libertação de
quantidades variáveis de hormonas no sangue), mas é mais preciso e rápido,
funcionando por emissão de potenciais de acção, sempre com a mesma intensidade e
duração. Transforma sinais em frequências de sinais, e quanto maior este número,
maior a intensidade.
O líquido encefaloraquidiano circula entre as meninges e serve para amortecer
pancadas no SNC, e para nutrição de certas células nervosas.
Organização do SNC:
O cérebro e a medula espinhal constituem o SNC. Os nervos espinhais saem do canal espinhal
ao nível da respectiva e contém as vias aferentes (raízes posteriores), que são sensitivas e as
vias eferentes (raízes posteriores), que são motoras e se projectam para a periferia.
Medula espinhal
Fazendo um corte transversal observa-se a substância cinzenta em forma de borboleta no
centro rodeada pela substância branca. As quatro asas da borboleta são os cornos (corte
transversal) ou colunas (corte longitudinal). Os cornos anteriores contêm neurónios motores
(projectados para os músculos), os cornos posteriores têm interneurónios. As fibras aferentes
têm os seus corpos celulares em gânglios espinhais. A substância branca contém os axónios
Página 6
dos tractos ascendentes e descendentes. As fibras nervosas aferentes (ou sensoriais)
transmitem a informação sensorial desde os órgãos dos sentidos, atingindo a medula no corno
posterior.
As fibras da raiz anterior transmitem informação motora desde a medula espinhal para os
órgãos efectores – fibras nervosas eferentes (ou motoras).
Encéfalo
É constituído pelo bulbo raquidiano, pons, mesencéfalo, cerebelo, diencéfalo e telencéfalo. O
bulbo, pons e mesencéfalo constituem o tronco cerebral. Estruturalmente é semelhante à
medula mas contém os núcleos dos corpos celulares dos nervos cranianos. O cerebelo é
importante para a coordenação motora.
Cérebro
O cérebro é constituído por dois hemisférios aparentemente semelhantes mas que hoje se
sabe não serem iguais nem anatómica nem funcionalmente. Do ponto de vista anatómico a
superfície externa do cérebro não é plana, apresenta circunvoluções e sulcos ou regos entre
elas e em cada hemisfério podem-se definir quatro lobos. O lobo frontal constitui a porção
anterior do hemisfério cerebral sendo limitado por baixo pelo rego de Sylvius e por trás pelo
de Rolando, atrás do qual se situa o lobo parietal. Abaixo do rego de Sylvius situa-se o lobo
temporal e o lobo occipital constitui a porção mais posterior dos hemisférios cerebrais. Na
superfície interna os hemisférios cerebrais apresentam igualmente circunvoluções e sulcos,
estruturas que os unem, como o corpo caloso, que permite troca inter-hemisférica de
informação, e o tronco cerebral que liga o cérebro à espinal medula.
Diencéfalo
O tálamo reúne muitas das vias aferentes como os olhos, ouvidos, pele, entre outras. O
hipotálamo é um centro nervoso autónomo mas desempenha também um papel importante
no controlo das hormonas libertadas pela hipófise.
Telencéfalo
Consiste no córtex cerebral e nos núcleos da base – corpo estriado: estriado (núcleo caudado e
putamen), globos pálidos (interno e externo), claustro; núcleos subtalâmicos; e substância
negra. O córtex cerebral divide-se em quatro lobos por fissuras. Este está dividido em dois
hemisférios, com diferentes funções, que estão interligados pelo corpo caloso.
Página 7
Cerebelo
Parte do encéfalo responsável pela manutenção do equilíbrio e pelo controle do tónus
muscular e dos movimentos voluntários, bem como pela aprendizagem motora. A função
cereberal é muito diferente da função cerebral: funciona só em nível inconsciente, dedica-se
exclusivamente ao serviço da motricidade (voluntária ou involuntária), mas nunca inicia ou
comanda qualquer movimento. A sua influência sobre os neurónios medulares é sempre
indirecta: ou através de tractos originados no tronco ou no cortéx.
Tronco Encefálico (ou Cerebral)
Porção do sistema nervoso central situada entre
a medula espinhal e o cérebro, sendo quase na sua
totalidade intracraniano (apenas uma porção do
bulbo é exocraniana). Ocupa a fossa craniana
posterior. É no tronco encefálico que se encontra
fixo o cerebelo.
Bulbo Raquidiano
Órgão condutor de impulsos nervosos. Estabelece
ligação entre o cérebro e a medula espinal. Bastante
relacionado com funções vitais como a respiração,
os batimentos do coração e a pressão arterial, e com
alguns
tipos
de
reflexos
como
mastigação, movimentos peristálticos, falar, piscar
de olhos, secreção lacrimal e vómito (mais específico
da área postrema). Por isso uma pancada nessa área
ou a sua compressão por parte do cerebelo, que se
encontra posteriormente, pode causar morte
instantânea,
paralisando
os
movimentos
respiratórios e cardíacos.
Página 8
Diferenças funcionais dos Lobos
1. Lobo frontal
Actividade motora
Linguagem
(hemisfério
esquerdo)
Pensamento
Concentração
Memória de trabalho
Comportamento
social
e
personalidade
2. Lobo parietal
Processamento da sensibilidade corporal
Capacidades visuo-construtivas
3. Lobo temporal
Processamento das informações auditivas
Memorização
4. Lobo occipital
Processamento das informações visuais
Diferenças funcionais dos Hemisférios Cerebrais
Hemisfério Esquerdo: Linguagem e outras informações de carácter simbólico (cálculo,
raciocínio abstracto);
Hemisfério Direito: integração global multi-sensorial (identificação de uma paisagem,
melodia de uma música, informação do sistema emocional);
Página 9
Sistema nervoso periférico
O sistema nervoso periférico (SNP), é a parte do sistema nervoso que se encontra fora
do sistema nervoso central (SNC), sendo constituído por nervos e gânglios nervosos. Os nervos
podem ser:
Nervos Sensitivos: são os nervos que têm o papel de transmitir os impulsos nervosos do órgão
receptor até ao SNC;
Nervos Motores: conduzem o impulso codificado no encéfalo (SNC), até ao órgão efector;
Nervos Mistos: tem o mesmo papel que os nervos sensitivos e motores ao mesmo tempo.
Os órgãos receptores são os órgãos dos sentidos (visão, audição, olfacto, paladar e corpúsculos
tácteis). Já os órgãos efectores são basicamente as glândulas e os músculos.
É graças a este sistema que o cérebro e a medula espinal recebe e envia as informações que
nos permite reagir face às diferentes situações que têm origem no meio externo ou interno.
O sistema nervoso periférico pode ser dividido em duas classes diferentes dependendo da
origem ou terminação dos terminais nervosos que o constituem. Se os nervos começarem ou
acabarem, no encéfalo denominamo-los de 'pares cranianos', mas se começarem na medula
espinal designam-se de 'nervos espinhais'.
Pares Cranianos (fazem a conexão com o encéfalo)
Nervos espinhais: existem 12 pares de nervos cranianos; 8 pares de nervos
cervicais; 12 pares de nervos torácicos; 5 pares nervos lombares e de
nervos sacrais e 1 par de nervos coccígeos.
Página 10
Sistema nervoso motor
Um dos papéis mais importantes do sistema nervoso é
controlar as diversas actividades corporais. Isto é feito
controlando:
Contracção de músculos esqueléticos apropriados
Contracção de músculos lisos nos órgãos internos
Secreção de substâncias quimicamente activas
por glândulas exócrinas e endócrinas.
Estas actividades são chamadas funções motoras do
sistema nervoso e músculos e glândulas são chamadas
efectores, pois são as estruturas anatómicas que
efectuam as funções do sistema nervoso.
Nota: Os músculos esqueléticos podem ser controlados
por diversos níveis:
1. Medula espinhal
2. Substância reticular da medula, mesencéfalo e
“pons”
3. Gânglios basais (núcleos da base)
4. Cerebelo
5. Córtex motor
Cada uma destas áreas desempenha um papel específico, estando as regiões mais abaixo
responsáveis pelas respostas musculares automáticas e instantâneas, enquanto as superiores
regulam movimentos complexos controlados pelo pensamento.
Córtex motor:
Página 11
O córtex motor primário tem uma representação somatotrópica, denominada de homúnculo
motor que mostra o tamanho relativo das regiões que representam as diversas partes do
corpo. É de referir que neste diagrama, mais de metade é ocupado por regiões responsáveis
pelo controlo dos músculos de mão e de fecho.
Níveis de organização do sistema motor somático
Conceito de neurónios motores superiores e inferiores
O ponto central para o diagnóstico em neurologia é definir se a lesão no paciente é nos
neurónios motores inferiores ou superiores. O neurónio motor é definido classicamente como
um neurónio cujo corpo celular e dendrites se localizam no SNC e cujos axónios se estendem
através dos nervos periféricos e vão fazer sinapse com as fibras musculares esqueléticas
extrafusais. Esta é a via final comum pela qual o SNC conduz comandos para os músculos
esqueléticos. Os neurónios que inervam os fusos musculares também são incluídos na
definição de neurónios inferiores.
Os neurónios motores superiores são todos os neurónios do SNC que influenciam os neurónios
motores inferiores. Subdividem-se em 3 subsitemas: os sistemas piramidal, extrapiramidal e o
cerebelo. Os neurónios motores superiores consideram-se como sistemas multineuronais que
se iniciam no cérebro e que enviam os seus axónios ao longo da medula espinhal ou no tronco
cerebral, para fazer sinapses com os neurónios motores inferiores.
Página 12
Vias motoras
Vias motoras nascem no córtex frontal, descem dentro do sistema nervoso e passam pelo
bulbo.
Via corticoespinhal ou piramidal: Esta via está envolvida na motricidade voluntária,
nomeadamente no controlo dos movimentos finos das extremidades e transporta informação
motora do córtex até às pontas anteriores da medula.
O
tracto
corticoespinhal,
constituído
maioritariamente por axónios motores, parte do
córtex motor primário e passa pelo tronco cerebral.
Ao nível do bulbo (parte inferior do tronco
cerebral), a maioria das fibras desse tracto cruzamse, dando origem à via corticoespinhal lateral, e as
restantes continuam a descida pela medula no
mesmo lado, formando a via corticoespinhal
anterior. Eventualmente, grande parte das fibras
desta última via cruzam a medula para o lado
oposto, quer seja na região do pescoço quer seja na
região torácica, pelo que quase 100% da
motricidade voluntária é cruzada. Os axónios das
vias lateral e anterior fazem depois sinapse com um
2º neurónio que controla os músculos esqueléticos.
O cerebelo recebe as informações sobre o
planeamento motor e dos sistemas sensoriais
informações auditivas, visuais, vestibulares e
somestésicas e realiza as correcções necessárias
entre o movimento pretendido e o executado,
suaviza as transições entre os movimentos
regulando actividades dos músculos agonistas e
antagonistas.
Página 13
Nos gânglios da base (grupo de núcleos no cérebro
interconectados com o córtex cerebral, tálamo e tronco
cerebral), a libertação do neurotransmissor dopamina (feito
nestes mesmos gânglios) vai determinar se o impulso nervoso
segue pela via directa ou indirecta dos gânglios da base. Se
ocorrer a libertação, o impulso nervoso segue a via directa que
consequentemente facilita o movimento, caso contrário, o
impulso nervoso a segue a via indirecta que acaba por inibir o
movimento. No final, o comportamento motor normal depende
do balanço entre as vias directa e indirecta dos gânglios da
base.
Sistema nervoso sensitivo (somestésico)
Somestesia é a capacidade que homens e animais têm de
receber informações sobre as diferentes partes do seu corpo.
Sensação: Qualquer alteração externa ou interna, que
provoca uma resposta fisiológica, ou comportamental num
organismo (como temperatura, dor, toque, pressão…).
Percepção:
Função cerebral que
atribui significado a estímulos sensoriais, a partir de vivências
passadas. Consiste na aquisição, interpretação, selecção e
organização das informações obtidas pelos sentidos.
Consciência da sensação.
Parte sensorial do sistema nervoso
Grandes partes das actividades do sistema nervoso são
iniciadas por excitações de receptores sensoriais, quer sejam
visuais, auditivos, tácteis, ou outros. Isto causa reacção
imediata do cérebro, ou memoriza a experiência para
determinar reacções corporais no futuro.
Página 14
A informação sensorial entra no sistema nervoso central através de nervos periféricos e é
conduzido imediatamente para múltiplas áreas sensoriais na:
Medula espinhal
Formação reticular na medula, mesencéfalo no cérebro
Cerebelo
Tálamo
Córtex cerebral
Transdução dos estímulos sensoriais em impulsos eléctricos
Os receptores traduzem um estímulo físico-químico num impulso nervoso. A fibra sensitiva
unida a cada receptor mostra um estímulo limiar. O potencial do receptor é proporcional à
intensidade do estímulo (quanto maior o estímulo, maior a frequência dos potenciais de
acção). Passado um tempo os receptores sensitivos adaptam-se ao seu próprio estímulo tanto
total como parcialmente e podem ser classificados como receptores de adaptação lenta
(transmitem sinais com pouca variação de frequência enquanto o estímulo está presente) ou
receptores de adaptação rápida (só se activam quando muda a intensidade do estímulo).
Classificação fisiológica dos receptores
Todas
as
fibras
nervosas
periféricas se dividem em:
- Tipo A: maiores e maior
velocidade de condução, dando
uma sensação de dor rápida
(injecção ou corte)
- Tipo C: menores e mais lentas,
dando sensação de dor lenta
(ardor, queimadura).
Intensidade de um estímulo
Nas fibras sensitivas a intensidade
representa-se
utilizando
as
características
de
sumação
espacial e temporal. Um tronco
nervoso sensitivo de um nervo
periférico contém geralmente
várias fibras, cada uma das quais
está relacionada com um número
variável de receptores na sua
extremidade distal. A soma de
todos os receptores e fibras
relacionadas com um único nervo
Página 15
definem o seu campo receptor. Um estímulo intenso que se estenda por todo o campo
activará todas as fibras, enquanto que um estímulo menos intenso activará apenas algumas.
Transmissão e processamento de sinal nos grupos neuronais
Qualquer agregado de neurónios, como os do córtex cerebral, tálamo ou um núcleo individual
do tálamo pode ser denominado um grupo neuronal. Geralmente cada grupo neuronal tem
um conjunto de vias aferentes que podem proporcionar um estímulo limiar ou sublimiar.
Divergência de sinais aferentes através de um circuito neuronal
Tipo amplificador – impulso nervoso espalha-se para um número crescente de
neurónios, à medida que passa através de neurónios sucessivos no mesmo caminho. É
o caso do tracto corticoespinhal no controlo dos músculos esqueléticos, em que uma
única célula do córtex motor é capaz de excitar várias fibras musculares.
Tipo múltiplos tractos – o sinal é transmitido para direcções distintas da rede neuronal.
Por exemplo, a informação transmitida às colunas dorsais da medula que segue duas
direcções diferentes na parte inferior do cérebro: cerebelo e partes mais internas
como o tálamo.
Convergência de sinais
Múltiplos impulsos nervosos unem-se para excitar um único neurónio.
Convergência de uma única fonte (múltiplos terminais de uma única fibra nervosa
terminam no mesmo neurónio). Isto é importante porque os neurónios quase nunca
são excitados por potenciais de um único terminal, mas de múltiplos, de modo a que
seja atingido o potencial sublimiar e portanto garantir o disparo do potencial de acção.
Convergência de várias fontes. Por exemplo os interneurónios da medula espinal
recebem sinais convergentes de fibras nervosas periféricas que entram na medula,
fibras proprioespinhais e fibras corticoespinhais do córtex cerebral.
Nestes circuitos observa-se atenuação do sinal.
Circuitos recorrentes ou oscilatórios
Circuitos mais importantes do SN e funcionam por feedback positivo, ou seja, dentro do
circuito existem fibras nervosas colaterais que voltam a excitar o próprio circuito. Desta forma,
após estimulado, o circuito pode estar continuamente a enviar sinais, durante algum tempo.
Vias Somáticas Sensitivas
Os circuitos neuronais das vias podem ser constituídos por 3 tipos de neurónios:
Neurónios de 1ª ordem: conduzem os impulsos dos receptores para a medula ou
tronco cerebral onde fazem sinapse com neurónios de 2ª ordem
Neurónios de 2ª ordem: transmitem impulsos ao tálamo ou cerebelo onde sinapsam
Neurónios de 3ª ordem: localizados no tálamo e conduzem impulsos ao córtex
somato-sensitivo do cérebro.
Página 16
Com poucas excepções, a informação sensitiva é
levada pelas fibras nervosas desde a superfície do
corpo (excluindo a cara), entram na medula
espinhal através dos cornos dorsais. Uma vez
dentro do SNC, os sinais separam-se em uma de
duas vias: via da coluna posterior (s. propioceptiva
consciente) e via anterolateral (s. termo-álgica).
Via da coluna posterior: transporta sensações de
toque com elevada localização espacial, sensações
que requerem gradação de intensidade e
sensações de pressão e vibração. Nesta via os
sinais são transmitidos pelas colunas dorsais da
medula até ao cérebro (neurónio de 1ª ordem). Ao
nível do bulbo os axónios sinapsam (neurónio 2ª
ordem) e cruzam para o lado oposto da medula,
continuando depois pelo tronco cerebral até ao
tálamo (neurónio de 3ª ordem), acabando no
córtex somato-sensitivo do cérebro.
Via anterolateral: transmite apenas
sensações de toque grosseiro, dor e
temperatura, pressão e comichão, que
não requerem uma localização muito
específica nem elevada gradação de
intensidade. Imediatamente depois de
entrarem na medula (1ª ordem), os
neurónios sinapsam com axónios da
substância cinzenta (2ª ordem), cruzam
na medula e ascendem num tracto
anterior (toque e pressão) e num tracto
lateral (dor e temperatura), pela matéria
branca, até terminarem no tronco
cerebral ou no tálamo (3ª ordem),
acabando o percurso no córtex somatosensitivo.
Página 17
Via espino-cerebelosa (s. proprioceptiva inconsciente)
Na via da propriocepção inconsciente há apenas dois
neurónios e não os três como vimos até então. O
neurónio de 3ªordem faria conexão entre o tálamo e a
região cortical correspondente à modalidade de
sensação, mas como esta via não chega ao tálamo, não
há terceiro neurónio. Esta via transmite ao cerebelo
informações sobre a posição dos músculos
esqueléticos, tendões e articulações.
Página 18
Protecção do SN
Líquor - fluido corporal estéril e de aparência clara que ocupa o espaço
subaracnóideo no cérebro (entre as membranas aracnóide e pia-máter das meninges).
É uma solução salina muito pura, pobre em proteínas e células, e age como um
amortecedor para o córtex cerebral e a medula espinal. O sistema ventricular secreta o
líquido cefalo-raquidiano, sendo os ventrículos espaços e túneis no centro do encéfalo.
Meninges – separam os ossos do crânio do cérebro. São três delicadas membranas
(dura-máter – mais espessa e externa; aracnóide – a do meio, pia-máter – mais
delgada e está aderente aos feixes nervosos) que revestem e protegem o Sistema
nervoso central, medula espinal, tronco encefálico e o encéfalo.
Barreira hemato-encefálica - membrana que protege o SNC de substâncias
químicas presentes no sangue e permite ao mesmo tempo a função metabólica
normal do cérebro. Esta membrana é composta por agrupamentos de células
endoteliais, anexados aos capilares cerebrais. Esta densidade aumentada restringe
muito a passagem de substâncias a partir da corrente sanguínea, muito mais do que as
células endoteliais presentes em qualquer outro lugar do corpo.
Ossos do crânio
Página 19
Audição e equilíbrio
Características de um som:
Tonia: Qualidade fisiológica: associada à frequência do som – altura (grave/agudo –
baixa ou alta)
Sonia: Não é a intensidade física mas a intensidade fisiológica da sensação;
intensidade (Forte/Fraco) amplitude
Timbre: espectro de frequências. Distingue sons com a mesma tonia e Sónia.
Audição:
- Ouvido externo: capta o som e direcciona-o para o ouvido médio;
- Ouvido interno: onde se encontram os receptores de som e equilíbrio.
No ouvido médio há uma abertura
que comunica com a nasofaringe, a
trompa de Eustáquio, e que
permite equilibrar as pressões no
ouvido médio (ex: variação da
altitude). Nesta região do ouvido
existem 3 ossículos: martelo,
bigorna e estribo, que estão
ligados entre si e que conectam a
membrana do tímpano à cóclea.
Estes ossículos aumentam (em 1.3)
a força das vibrações do tímpano e
aplicam-na numa área (17x)
menor, ao nível da janela oval
(deste modo, as vibrações do som são amplificadas e transformadas em ondas de pressão na
cóclea). O tímpano e o sistema de ossículos funcionam assim como um adaptador de
impedância (o meio de propagação do som no ar é diferente do que no interior do ouvido, já
que a perilinfa no interior do labirinto membranoso tem muito mais inércia que o ar, portanto
o aumento da pressão pelo sistema ossicular é necessário para produzir vibração na perilinfa).
Na ausência dos ossículos o som é meramente perceptível.
O ouvido interno tem uma parte óssea e outra parte membranar. O labirinto ósseo forma
canais preenchidos com perilinfa e nos quais se encontra o labirinto membranoso, que está a
endolinfa (endolinfa é o líquido transparente contido no labirinto membranoso do ouvido
interno e perilinfa é o líquido contido no ouvido, no espaço existente entre o labirinto
membranoso e o ósseo).
A cóclea (caracol) é parte do labirinto óssea e está dividida em 3 tubos: o ducto coclear (parte
do labirinto membranoso), a rampa vestibular que contém perilinfa e estão separados da
rampa timpânica pela membrana basilar (compartimento que contém endolinfa).
Página 20
No Orgão de Corti, localizado dentro do ouvido interno, é onde se gera os impulsos nervosos
em resposta à vibração da membrana basilar.
A membrana basilar vibra em locais
diferentes conforme a frequência do som.
Deste modo, o ouvido analisa os sons
complexos decompondo-os nas suas diversas
frequências de acordo com o local de
vibração da membrana (assim sons com o
mesmo timbre são distinguidos).
O local de maior vibração / de
reconhecimento é a porção da membrana
cuja frequência própria é igual à frequência
da onda sonora. (baixa frequência base da
membrana; alta frequência fim da
membrana)
Página 21
Equilíbrio
Para além da cóclea, o ouvido interno é também formado pelos canais semicirculares, o sáculo
e utrículo, que estão associados ao equilíbrio e formam o sistema vestibular.
O utrículo e o sáculo são duas câmaras que têm uma
pequena zona sensível, a mácula, responsável por
interpretar a aceleração linear determinar a orientação
da cabeça. As hair cells inclinam-se conforme a
orientação da gravidade (ou seja conforme a postura da
cabeça)
e
provocam
a
despolarização
ou
hiperpolarização dos neurónios.
Utriculo mácula horizontal orientação da cabeça
quando estamos de pé mais sensível à aceleração
horizontal;
Sáculo mácula vertical orientação deitado mais sensível à aceleração vertical
No sáculo e utrículo existem otólitos, cristais de CaCO3, que têm maior densidade que o
líquido circundante e provocam uma maior inércia que é necessária à estimulação das células
cíliadas dessas estruturas.
Os 3 canais semi-circulares estão colocados de tal forma que cada um deles corresponde a um
plano do espaço. No final dos canais há uma zona mais larga, a ampola, que está preenchida e
tem um órgão sensorial sensível à aceleração angular, ajudando assim a determinar a direcção
do movimento da cabeça.
Cada ampola tem uma área elevada, a crista ampullaris, onde estão localizadas célula pilosas.
Os cílios dessas células estão embebidos numa membrana gelatinosa, a cúpula, cujo
movimento activa as células receptoras.
Quando a cabeça inicia um movimento e devido à sua inércia, a endolinfa dos canais
semicirculares permanece estática, o que provoca um movimento relativo desse fluido no
sentido oposto ao da rotação da cabeça.
Depois das células ciliadas, são enviados sinais apropriados pelo nervo vestibular, para o SNC,
indicando-lhe uma mudança na rotação da cabeça e a velocidade de rotação.
Página 22
Para o equilíbrio de todo o corpo participam informações recolhidas pelo ouvido, olhos,
músculos, tendões, receptores cutâneos, etc.
Via auditiva
As fibras nervosas do gânglio espiral de Corti
entram no núcleo dorsal e ventral do bolbo
raquidiano. Neste ponto, fazem sinapse com
neurónios de segunda ordem, que passam para o
lado oposto do tronco cerebral para terminar no
“superior olivary nucleus”. Outros neurónios de
segunda ordem não chegam a cruzar para o lado
oposto e vão para o núcleo “superior olivary” do
mesmo lado. De ambos os lados, sobem para o
lemniscus lateral e daí para o coliculus inferior,
onde quase todas as fibras sinapsam. Daí, a via
segue para o núcleo geniculado medial onde
todas as fibras sinapsam. Finalmente, a via vai
para o córtex auditivo, localizado no lobo
temporal.
Nota: Muitas fibras colaterais da via auditiva passam directamente para o sistema de activação
reticular do tronco cerebral, que activa todo o sistema nervoso em resposta a sons elevados.
Outras fibras colaterais vão para o vérmix do cerebelo, que é activado instantaneamente no
caso de sons altos.
Página 23
Visão
Olho
A córnea é rígida e quase cristalina, e é transparente
a alguns comprimentos de onda (condição de
transparência). Tem uma forma esferocilíndrica com
algumas irregularidades – pontos de astigmatismo.
Perde água de forma activa:
¼ por evaporação canais de água;
¾ pelas camadas endoteliais. Essas camadas
não se renovam ao longo da vida e à medida
que morrem, diminuem a capacidade de
perda de água e a transparência da córnea.
A córnea tem uma capacidade dióptrica (≈ 44 dioptrias), ajudando a focar os raios de luz na
retina. Tem espessura variável entre 0,5 mm (região central) e 1 mm (região periférica) e é
muito enervada.
O cristalino é uma lente biconvexa transparente. No seu núcleo central tem células
embrionárias. Tem capacidade dióptrica ≈ 22 dioptrias mas tem potência dióptrica variável,
adaptando-se para focar a diferentes distâncias. Um conjunto de ligamentos, a zónula, liga o
cristalino ao resto da estrutura do olho. Esta estrutura associa-se ao corpo ciliar, que tem
fibras musculares circulares e longitudinais cuja contracção é regulada pelo sistema
parassimpático.
O cristalino funciona como uma lente biconvexa, convergente, que aumenta a sua potência
dióptrica, tornando-se mais convexo, permitindo a visão de perto. Esta capacidade denominase de acomodação e é a capacidade do cristalino de alterar a sua forma e, portanto, a sua
potência dióptrica. Com o avançar do tempo o cristalino vai perdendo a sua capacidade de
acomodação (originando a presbiopia), e o mesmo acontece com a reserva acomodativa. Isto
compensa-se com lentas positivas/convergentes. Também com o envelhecimento, o cristalino
pode perder a sua transparência, o que provoca “borramento” da visão (catarata).
Contracção das fibras relaxamento dos ligamentos cristalino mais esférico focagem
mais perto, maior poder refractivo.
A potência real do olho resulta da conjugação dos índices de refracção dos diferentes meios
que a luz atravessa (córnea, humor aquoso, cristalino,…), sendo o poder refractivo total 59
dioptrias.
Página 24
O fluido intraocular é constituído por:
Humor aquoso: líquido entre a córnea e o cristalino que nutre estas estruturas. É
constantemente secretado pelo corpo ciliar e forma-se por transporte activo ou
difusão facilitada a partir do plasma. É reabsorvido no canal de Schlemm e enviado
para a veia ocular. É o responsável pela pressão intraocular (mantém o olho
distendido). Glaucoma é uma doença associada ao aumento da pressão intraocular,
resultante do bloqueio do canal de Schlemm (ocorre acumulação de humor aquoso no
olho).
Humor vítreo: líquido gelatinoso entre o cristalino e a retina. Não é renovado. Não há
movimento de fluido mas pode haver difusão de substâncias.
A retina é sensível à luz e contém fotoreceptores que são excitados e enviam informações
para o cérebro via nervo óptico. Esses receptores actuam como transdutores, convertendo a
informação em impulsos nervosos.
Fotoreceptores:
Cones: reconhecem cores - cones azuis, verdes e vermelhos (absorvem o comprimento
de onda destas cores). Concentram-se na fóvea (centro da mácula).
Bastonetes: responsáveis pela visão a preto e branco e visão nocturna (retina).
Opsinas: proteínas fotossensíveis:
Bastonetes: escotopsina (505 nm)
Cones: fotopsinas azul (445 nm), verde (535 nm) e vermelho (570 nm).
Foto transdução: Os fotoreceptores têm receptores transmembranares. Quando os fotões
interagem com os receptores modificam a sua conformação e desbloqueiam uma enzima que
actua num segundo mensageiro, gerando uma onda de hiperpolarização. A resposta gerada
pelos fotoreceptores é assim uma resposta electrotónica (propagação do impulso dentro de
um neurónio, onde não contribuiem as mudanças de voltagem na condutância da membrana),
respondendo a diferentes intensidades de luz.
A interpretação do nível de estimulação de cada tipo de cone dá a ilusão de uma cor. Ex:
Laranja – 99% vermelho + 42% verde (x% significa nível de estimulação do cone).
Blind spot é o local da retina, na zona da mácula, onde parte o nervo óptico. Como neste local
não há fotoreceptores, a imagem aí focada “não se vê”. A fóvea, no centro da mácula, é o local
de maior acuidade visual.
Os neurónios e nervos associados aos cones são maiores e têm uma condução mais rápida que
os associados aos bastonetes.
Cones células bipolares células ganglionares
Bastonetes células bipolares células amácrimas células ganglionares
Página 25
À excepção das células ganglionares, que transmitem potenciais de acção ao nervo óptico, em
todas as outras células se dá condução electrotónica.
Via óptica
Os sinais nervosos partem da retina pelos nervos ópticos e
cruzam-se no quiasma óptico. Aí, as fibras que transmitem
informação do campo visual direito cruzam em direcção ao
hemisfério esquerdo e as fibras relativas ao campo visual
esquerdo cruzam para a direita, dando origem a dois tractos
ópticos, que seguem, um para cada hemisfério, em direcção
ao córtex visual primário.
Lesões em diferentes vias/zonas da via óptica podem ser
avaliadas/diagnosticadas através de um exame de campos
visuais. Outros testes ao nervo óptico incluem:
- reflexos da pupila
- tabelas
- visão cromática
- Estereoscopia
- campos visuais: testes de confrontação e campímetro de arco.
Página 26
Os sentidos químicos: paladar e olfacto
Paladar
É uma função principalmente ao cargo das papilas gustativas da língua, sendo
influenciado pelo olfacto. A identificação dos químicos específicos que excitam os
diferentes receptores do paladar não são todas conhecidas, mas os que se conhecem
agrupam-se em 5 categorias chamadas as sensações primárias do paladar: salgado,
doce, amargo, azedo e “umani” (palavra japonesa que designa delicioso).
Sabor azedo: é causado por ácidos, isto é, pela concentração de iões hidrogénio
e a intensidade da sensação é aproximadamente proporcional ao logaritmo da
concentração de iões.
Sabor salgado: é causado por sais ionizáveis, principalmente pela concentração
do ião sódio. A qualidade varia de sal para sal, pois alguns causam outras
sensações além de salgado.
Sabor doce: não é causado por nenhuma classe específica de químicos. No
entanto, são essencialmente químicos orgânicos que causam este sabor. A
mudança de apenas um radical pode mudar a substância de doce para amargo.
Sabor amargo: é causado essencialmente por substâncias orgânicas.
Sabor “umani”: designa um sabor que causa uma sensação agradável. É
causado por alimentos contendo L-glutamato.
As papilas gustativas e sua função
A papila gustativa é composta por cerca de 50 células epiteliais,
sendo umas células de sustentação e outras células gustativas.
Estas últimas regeneram-se continuamente por mitose. As
superfícies apicais das células gustativas dispõem-se em redor de
um poro gustativo de onde saem microvilli, que são receptores
de paladar. Aí fibras nervosas gustativas, estimuladas por células
receptoras gustativas, rodeiam os corpos das células gustativas.
Formam-se muitas vesículas por baixo da membrana celular
perto das fibras, que se acredita terem neurotransmissores, libertados pela membrana
celular para excitar os terminais das fibras nervosas em resposta à estimulação
gustativa.
As papilas gustativas são encontradas nas papilas circunvaladas, papilas fungiformes e
papilas foliadas. Existem também no palato, nos pilares tonsilares na epiglote e no
esófago proximal.
Página 27
Transmissão de sinais gustativos para o SNC
Estes sinais originados na porção anterior da língua passam
primeiro para o nervo lingual, e depois, através da “chorda
tympani” para o nervo facial, e finalmente para o tracto
solitário localizado no tronco cerebral.
Sinais de paladar da parte posterior da língua, e outras regiões
posteriores da boca e faringe são transmitidos através do nervo
glossofaríngeo que vai até ao tracto solitário do tronco cerebral
(da base da língua e outras regiões da faringe, através do nervo
vago).
Todas as fibras sinapsam nos núcleos do tracto solitário (na
parte posterior do tronco cerebral). Estes núcleos enviam
neurónios de segunda ordem para uma pequena área do
núcleo medial posterior ventral do tálamo. Do tálamo,
neurónios de terceira ordem são enviados para o córtex
parietal.
Nota: Esta via é paralela à via somatossensorial da língua.
Do tracto solitário, muitos sinais de paladar são enviadas para os núcleos salivares
superiores e inferiores, que transmitem sinais para as glândulas submandibulares,
sublinguais e parótidas para ajudar na secreção de saliva.
Olfacto
Membrana olfactiva
Localiza-se na parte superior de cada narina. Os receptores
celulares são as células olfactivas, que são células nervosas
bipolares que derivam do SNC. Existem cerca de 100 milhões
destas células no epitélio olfactivo dispersas nas células
sustentaculares. A extremidade mucósica das células olfactivas
possui 4 a 25 cílios olfactivos que se projectam para a camada de
muco que cobre a cavidade nasal. Entre estas células existem as
glândulas de Bowman que segregam o muco.
Estimulação das células olfactivas
A porção destas células que responde ao estímulo é os cílios olfactivos. A substância
com odor, quando entra em contacto com a membrana olfactiva, primeiro difunde-se
no muco. De seguida, liga-se a proteínas receptoras na membrana de cada cílio. Cada
proteína receptora é uma molécula bastante longa, prolongando-se para dentro e fora
da membrana ciliar. A substância à parte exterior da proteína, causando uma alteração
Página 28
conformacional na parte interna, que por sua vez desencadeia um processo que abre
os canais de sódio, que causa um potencial de acção que é transmitido para o SNC pelo
nervo olfactivo.
A importância deste mecanismo é que multiplica bastante o efeito excitatório de uma
substância com “pouco cheiro”.
Transmissão de sinais de odor para o SNC
As fibras olfactivas que saem do bulbo olfactivo são chamadas
nervo cranial I, ou tracto olfactivo. No entanto, na realidade,
tanto o tracto olfactivo como o bulbo são o resultado do
crescimento de tecido cerebral anterior.
O bolbo olfactivo está sobre a placa cribriforme, que separa a
cavidade craniana da cavidade nasal (parte superior). Esta
placa tem múltiplas perfurações pequenas, através dos quais,
um igual número de pequenos nervos passa para cima da
membrana olfactiva na cavidade nasal, para o bulbo olfactivo,
na cavidade craniana.
Os axónios que levam o sinal olfactivo desde as células olfactivas terminam nos
glomérulos do bolbo olfactivo. Cada glomérulo é também o terminal das dendrites
vindas de células mitrais e células “tufo”. Os sinais são enviados ao longo do tracto
olfactivo até às zonas superiores do SNC.
Existem um sistema muito antigo de olfacto que assegura os reflexos olfactivos básicos
localizados anteriormente ao hipotálamo, um sistema olfactivo menos antigo de
controlo automático parcial de aversão a comida tóxica ou pouco saudável e um
sistema moderno utilizado na percepção consciente e análise olfactiva.
Página 29
Aula 3 – Sistema nervoso autónomo
Divisão do sistema nervoso (Resumo):
SNC:
- Processamento de informação
- Encéfalo e medula espinal
SNP:
- Rede de comunicação
- Nervos, receptores sensoriais e gânglios (12 pares de nervos cranianos, 31 pares de
nervos espinhais)
SN somático:
- Acções musculares voluntárias
- Neurónios sensoriais que recebem informações dos órgãos dos sentidos e neurónios
motores que controlam os músculos esqueléticos
SN autónomo:
- Acções musculares involuntárias
- Neurónios sensoriais que recebem informações de órgãos viscerais e neurónios
motores que controlam o músculo cardíaco, músculo liso e glândulas
- Activado por centros localizados na medula espinal, tronco cerebral e hipotálamo.
Sistema Nervoso Autónomo
Função:
Sistema de integração para a manutenção da
homeostase (regulação do corpo do ser vivo
para manter uma condição estável, face
múltiplos
ajustes
de
equilíbrio
dinâmico controlados por mecanismos de
regulação interrelacionados).
•
SNA regula extrinsecamente o músculo liso, músculo cardíaco e secreção glandular
•
Indirectamente os processos metabólicos
Este sistema têm uma enorme precisão na acção e têm uma elevada importância biólogica.
Está responsável pelo mecanismo de arco reflexo.
Página 30
Exemplos de funções do SNA:
O SNA está dividido em 3 partes:
Sistema nervoso simpático: (SNS)
o Mobiliza rapidamente energia e respostas a exercício e situações de resposta
rápida / emergência (stress)
o “Flight-or-fight response”
Sistema nervoso parassimpático: (SNP)
o Trabalha para a conservação da energia e promove actividades de não
emergência e funções de organismo, como a salivação, lacrimação, digestão…
o “Rest and digest response”
Sistema nervoso entérico:
o Rede neuronal no tracto intestinal
o Responsável pelos movimentos do intestino, pela actividade de glândulas
endócrinas e pela secreção de sucos gástricos.
Os órgãos internos são, em geral, enervados por fibras pertencentes aos dois primeiros
sistemas, sendo que normalmente um tipo de enervação tem sobre o órgão um efeito
antagonista àquele produzido pelo outro tipo - se o simpático estimular, o parassimpático
inibe e vice-versa. Como apontado, os dois sistemas diferem pela localização anatómica dos
gânglios e neurónios pré-ganglionares, assim como pelos neurotransmissores utilizados na
sinapse efectora - adrenalina no simpático e acetilcolina no parassimpático.
Página 31
Diferenças anatómicas:
S.N. Simpático
Neurónios pré-ganglionares curtos, contactam com gânglios longe dos órgãos
efectores.
Fibras pós-ganglionares longas
Fibras simpatéticas têm origem nos nervos espinhais que saem nos segmentos
T-1 a L-2 da coluna.
S.N. Parassimpático
Neurónios pré-ganglionares longos, contactam com gânglios perto dos órgãos
efectores
Fibras pós-ganglionares curtas
Fibras parassimpatéticas têm origem nos nervos cranianos III, VII, IX e X e nos
nervos que saem das vértebras sacrais 2 e 3 e, ocasionalmente, também da 1 e
4.
Página 32
Organização química
Os principais neurotransmissores do SNA são a acetilcolina e a noradrenalina, ou
norepinefrina.
Acetilcolina (transmissor parassimpático) sistema colinérgico (Rest and digest)
Noradrenalina (transmissor simpático) sistema adrenérgico (Stress)
Fibras pré-ganglionares: sempre colinérgicas
Fibras pós-ganglionares
o SN parassimpático: colinérgicas
o SN simpático:
Maioria adrenérgicas
Algumas colinérgicas (glândulas sudoríparas, músculos pilo-erectores,
alguns vasos sanguíneos)
Isto implica a existência de S.N. simpático adrenérgico e S.N. simpático
colinérgico.
Provas de avaliação do SNA
Função parassimpática
Respiração profunda (R6): estuda-se a variação da
frequência cardíaca com a inspiração profunda.
Durante a inspiração há um aumento do volume
torácico e diminuição da pressão intratorácica, o que
leva ao aumento do volume sistólico e,
consequentemente, ao aumento da pressão arterial.
É activado um mecanismo de compensação,
regulada pelo sistema parassimpático, que actua no
sentido de diminuir a pressão arterial através da
diminuição da frequência cardíaca.
PA – Pressão Arterial; R – Respiração; Vs – Volume
sistólico; FC – Frequência Cardíaca
𝑷𝑨 = 𝑹 ∗ 𝑽𝒔 ∗ 𝑭𝑪
Esquema:
Inspiração Volume tóracico Pressão intratorácica
Pressão arterial Compensação (SNP)
Volume sistólico
Frequência cardíaca Pressão arterial
Página 33
Manobra de Valsava (VR): respiração forçada. No início da expiração verifica-se um ligeiro
aumento do volume sistólico, devido ao maior fluxo de sangue para o coração, o que
provoca um ligeiro aumento da pressão arterial (1) e diminuição da frequência cardíaca.
De seguida, o aumento da pressão intratorácica devido à respiração contra uma resistência
(impede o ar de sair dos pulmões) leva a uma diminuição da PA (2). Para contrariar esta
tendência, o SNP promove o aumento da frequência cardíaca e a constrição de vasos,
aumentando a resistência (3), o que eleva a pressão arterial até níveis basais. Quando o ar
é finalmente expelido os vasos que rodeiam o coração dilatam causando um ligeiro
abaixamento no volume sistólico e na pressão arterial (4). Logo de seguida, o sangue
retorna ao coração e o Vs aumenta, provocando uma súbita elevação da Pressão (5). Para
manter a pressão em valores normais, a FC baixa bruscamente (6) para depois estabilizar.
Esquema:
(1) Expiração forçada / contra resistência ligeiro Vs ligeiro PA
da FC
(2) Pressão intratorácica (devido a (1)) Volume sistólico
PA
(3)
FC, constrição de vasos (contrariar a tendência (2))
PA
(4) Libertação do ar dilatação dos vasos ao pé do coração ligeiro Vs ligeiro
(5)
Vs, retorno do sangue PA
(6)
FC Estabilização
PA
PA
Ortostatismo activo (teste efectuado quando o paciente se levanta) (30:15)
Quando o paciente se levanta de repente o sangue tende a afluir para as pernas devido à
gravidade. Devido à diminuição do volume sistólico a pressão arterial baixa (1), o que leva
ao aumento da frequência cardíaca (A), até que a pressão atinja valores basais, seguida de
uma nova diminuição da frequência (B). Para estabilizar a pressão, a FC volta a aumentar,
atingindo um pico no 15º batimento (C) e decresce depois (D) até que ao 30º batimento
estabiliza.
Para verificar se o SN Parassimpático está a
funcionar correctamente calcula-se o índice
30:15, dividindo o valor do intervalo R-R ao
30º e 15º batimento.
Página 34
𝑅𝑅[30]
> 1,04 (𝑛𝑜𝑟𝑚𝑎𝑙)
𝑅𝑅[15]
𝑅𝑅[30] > 𝑅𝑅[15] => 𝐹𝐶[30] < 𝐹𝐶[15] (𝑜 𝑞𝑢𝑒 é 𝑠𝑢𝑝𝑜𝑠𝑡𝑜)
Ortostatismo passivo (teste efectuado quando se deita)
Função simpática adrenérgica:
o ΔP sistólica e diastólica ao ortostatismo activo e passivo (teste Tilt - exame que
avalia o comportamento do ritmo cardíaco e da pressão arterial em resposta a
uma variação da postura corporal)
o ΔP sistólica com manobra de Valsalva e frio cutâneo pressão sobe
gradualmente
o ΔP diastólica ao esforço isométrico a pressão vai subindo e desce quando o
esforço acaba. O esforço aumenta a resistência periférica, e
consequentemente aumenta ligeiramente a pressão diastólica e a pressão
sistólica.
Função simpática colinérgica:
o Estudo da vasomotricidade cutânea por fluxometria com Laser doppler
permite avaliar o fluxo da microcirculação (o SNS participa na vasoconstrição).
o Testes de função sudomotora: teste de termorregulação; teste de reflexo
axonal sudomotor (QSART); teste de impressão com moldes de silicone
Em resumo Doenças:
Tonturas, sincopes, disfunção eréctil, hipohidrose, pupilas mióticas e queda dos
valores da PA com o ortostatismo Disfunção autonómica
Página 35
Ataxia e dificuldade em realizar movimentos alternados rápidos Disfunção
cerebelosa
Tremor, rigidez e roda dentada parkinsonismo
Exemplos de Reflexos
Reflexos Oculares – Dividem-se em três tipos:
Fotomotor - (cuja via de aferência é composta pelo II par craniano - nervo óptico - e a
de eferência pelo III par craniano - motor ocular comum/oculomotor - sendo o centro
do reflexo o mesencéfalo) envolve a modificação do tamanho da pupila por resposta a
estímulos luminosos;
Acomodação - que envolve a distensão ou contracção da pupila, como o anterior, mas
devido à acomodação desta para focar pontos a diferentes distâncias;
Reflexo corneano, que equivale a um toque no olho, cuja resposta será o
encerramento do mesmo. (A via de aferência é composta pelo V par craniano trigémeo - e a via de eferência pelo VII par - facial - o centro do reflexo é a
protuberância).
Reflexo na laringe - reflexo faríngeo - de vómito
Neste caso, o IX par craniano - glossofaríngeo - é responsável pela via aferente e eferente e o
centro deste reflexo é no bulbo raquidiano.
Reflexos nos membros superiores
Estes são reflexos osteotendinosos e para os verificar, é utilizado um martelo especial para o
efeito. Conduzem a contracções musculares e dividem-se em:
Radial - ap. estilóide radial - activa C5-C6;
Bicipital - tendão - activa C5-C6;
Triciplital - tendão - activa C6, C7 e C8.
Reflexos do Abdómen
Estes reflexos podem ou não ser visíveis em pacientes muito obesos ou em mulheres que
tenham sofrido de múltiplas gravidezes. São também referidos como reflexos
cutâneoabdominais e dividem-se em:
Superior - D6-D7;
Inferior - D10, D11 e D12;
Médio.
Reflexos nos membros inferiores
Cremabtoriano - reflexo que afecta a face superior e interna das coxas;
Rotuliano - tendão da rótula - L2, L3 e L4;
Aquiliano - tendão de Aquiles;
Plantar - ou cutâneo-plantar - região plantar do pé, com flexão deste.
Página 36
Coração
1) Órgão com diversas funções, todas elas de importância vital.
Gera a pressão sanguínea
Dirige a circulação e assegura o seu fluxo unidireccional.
2)
Localização e forma:
Localizado no interior cavidade torácica, na zona do mediastino.
2/3 do seu volume encontram-se à esquerda da linha média.
Tem o tamanho de um punho fechado e a sua forma aproxima-se da de um cone,
terminando no apex, um vértice formado pelo ventrículo esquerdo.
A sua face posterior constitui a base do coração.
3) O coração é constituído por 4 câmaras ou cavidades:
2 aurículas (direita e esquerda): são as câmaras superiores do coração e localizam-se
numa posição posterior. Tem forma ovóide irregular, recebem o sangue que chega ao
coração e bombeiam o sangue para os ventrículos. Têm as paredes mais finas que os
ventrículos e são separadas pelo septo interauricular;
2 ventrículos (direito e esquerdo): são as câmaras inferiores do coração e localizam-se
numa posição anterior. Tem forma piramidal ou de cone, recebem o sangue das aurículas
e bombeiam-no para o resto do corpo. São maiores e têm paredes mais grossas do que as
aurículas. O ventrículo esquerdo é mais grosso que o direito pois é o responsável pela
circulação sistémica. Estão separados pelo septo interventricular.
No coração, as aurículas comportam-se de maneira diferente dos ventrículos. Em quanto que
nas aurículas a despolarização e a repolarização dão-se no mesmo ponto, nos ventrículos, a
repolarização começa no ponto onde acaba a despolarização.
4) Válvulas
Estruturas formadas basicamente por folhetos de tecido conjuntivo, chamados valvas.
Situadas entre aurículas e ventrículos bem como nas saídas das artérias aorta e artéria
pulmonar.
Permitem o fluxo do sangue em um único sentido não permitindo que este retorne
fechando-se quando o gradiente pressórico se inverte.
O que regula a abertura e fechamento das válvulas são as pressões dentro das câmaras
cardíacas.
As válvulas dentro do coração – válvulas AV:
o Mitral ou bicúspide – Possui dois folhetos. Permite o fluxo sanguíneo entre a
aurícula esquerda e o ventrículo esquerdo.
o Tricúspide - Três folhetos. Permite o fluxo sanguíneo entre a aurícula direita e o
ventrículo direito.
o A suspensão de ambas as válvulas mitral e tricúspide é mantida por acção dos
músculos papilares.
Página 37
As válvulas existentes à saída dos ventrículos – válvulas semilunares ou sigmóides:
o Aórtica - Permite o fluxo sanguíneo de saída do ventrículo esquerdo em direcção à
aorta.
o Pulmonar - Permite o fluxo sanguíneo de saída do ventrículo direito em direcção à
artéria pulmonar.
5) Nutrição e Circulação
Veias pulmonares direitas (sangue venoso) aurícula direita ventrículo direito artérias
pulmonares circulação pulmonar pulmões (sangue venoso => sangue arterial) veias
pulmonares esquerdas aurícula esquerda ventrículo esquerdo artéria aorta
circulação sistémica órgãos (sangue arterial => sangue venoso) …
O coração é nutrido pelas artérias coronárias direita e esquerda que executam a circulação
coronária ou cardíaca.
6) Tecido cardionector
Presente no endocárdio, é um tipo de tecido muscular cardíaco especializado na
produção e condução dos impulsos eléctricos.
O sistema cardionector é formado por:
o
Nódulo Sinusal: local na parede postero-superior da aurícula direita onde se
iniciam os impulsos. O esqueleto fibroso não permite a sua propagação directa
para os ventrículos, que se dá ao nível do feixe de His. Dispara 60 a 100 vezes
por minuto;
Página 38
o
o
o
Nódulo aurículo-ventricular: localiza-se na zona mediana da válvula
aurículoventricular direita e atrasa a propagação dos impulsos eléctricos
permitindo a separação temporal entre a contracção auricular e ventricular;
Feixe de His: tem origem no nódulo aurículo-ventricular e atinge o septo
interventricular, dividindo-se no ramo direito e esquerdo. O ramo esquerdo
estende-se por baixo do endocárdio e atinge a região apical dos ventrículos
onde termina na rede de Purkinje;
Rede de Purkinje: rede de fibras musculares cardíacas modificadas que
medeiam a contracção coordenada do miocárdio. As fibras de Purkinje
possuem células maiores que as do miocárdio, binucleadas, com citoplasma
extenso e não apresentam discos intercalares.
7) Propriedades do Coração
a) Automatismo / Cronotropismo
Na ausência de estímulo, a célula é capaz de gerar um potencial de acção
devido à abertura aos canais de sódio, mesmo em repouso.
As células encontram-se no nodo sino-auricular / sinusal
b) Condutibilidade / Dromotropismo
O estímulo gerado no modo sinusal transmite-se a todas as células cardíacas
c) Excitabilidade / Batmotropismo
As células cardíacas são capazes de responder a estímulos.
0 Potencial de acção (abertura dos canais rápidos de sódio)
1 Pico de tensão
2 Fase plateau (abertura dos canais de cálcio no coração) Deve ser
relativamente longa para aumentar o tempo entre contracções, permitindo
que os ventrículos se encham.
3 Fase descendente
4 Potencial de repouso
Página 39
d) Contractibilidade / Inotropismo
As células cardíacas conseguem contrair-se;
Lei de Frank-Starling: Quanto maior o comprimento inicial da fibra miocárdica,
maior a força de contracção. Isto é só válido até aos 2,2 µm.
e) Tónus
O músculo cardíaco tem uma tensão-base;
f)
Lusitropia
Os ventrículos relaxam durante a diástole.
8) Ciclo Cardíaco
1.
2.
3.
4.
Válvula aurículo-ventricular fecha
Válvula aórtica abre-se
Válvula aórtica fecha-se
Válvula AV abre
A.
B.
C.
D.
E.
F.
Contracção isovolumétrica
Expulsão do sangue
Relaxamento isovolumétrico
Enchimento rápido
Diástase
Sístole Arterial
a) Sístole Ventricular
i. Encerramento das válvulas AV (1)
Página 40
ii. Contracção do miocárdio, aumentando a pressão porque o volume sanguíneo
mantem-se constante (as válvulas estão fechadas) – Contracção
isovolumétrica (A)
iii. Quando a pressão ventricular iguala a aórtica, as válvulas sigmóides abrem-se
(2), dando-se a expulsão do sangue. Inicialmente a expulsão é rápida e depois
fica lenta.
b) Diástole Ventricular
iv. Com a pressão a diminuir dá-se o encerramento das válvulas sigmóides (3).
v. O ventrículo continua a relaxar, mas o volume de sangue mantém-se
constante – Relaxamento Isovolumétrico (C)
vi. Quando a pressão é inferior à aurícula, dá-se a abertura das válvulas AV (4).
O sangue passa da aurícula ao ventrículo, primeiro rapidamente (D) e depois
lentamente (Diástase – E)
vii. Dá-se a sístole auricular (F) e o ciclo recomeça.
9) Circulação coronária
O sangue passa para as artérias coronárias durante a diástole, através das ósteas coronárias (2
orifícios que estão no inicio da aorta). O sangue venoso é drenado para a aurícula direita
através das veias cardíacas anteriores.
Isquémia – Falta de oxigenação dos tecidos por falta de irrigação de sangue.
Doença isquémica cardíaca:
Uma das principais causas de morte
Pode ser causada por oclusão coronária e enfarte do miocárdio.
Causa mais frequente para a diminuição do fluxo sanguíneo: aterosclerose – grandes
quantidades de colesterol vão ficando gradualmente depositados sobre o endotélio,
em diversos pontos em artérias por todo o corpo. Formam-se então as chamadas
placas arterioscleróticas que se prolongam para o lúmen, bloqueando parcialmente ou
totalmente o fluxo sanguíneo.
Um local comum para a formação destas placas é nas artérias coronárias.
Factores que influenciam o débito coronário:
1) Físicos
Pressão aórtica:
o Variações da pressão aórtica provocam variações do débito coronário no
mesmo sentido.
Resistência arteríolar (Factor físico determinante)
o Causada pelas variações da actividade metabólica do miocárdio.
Débito sistólico, que depende de:
o Contractibilidade ou inotropismo;
o Pré-carga (volume dentro do ventrículo antes da sístole – volume telesistólico)
Página 41
o
Pós-carga (Afterload) -> medida pela pressão arterial (resistência que a aorta
oferece à passagem do sangue)
Nota: Há mais irrigação nas coronárias durante a diástole, pois na sístole, as valvas da válvula
sigmodeia (aórtica) tapam parcialmente as ósteas coronárias.
Carácter físico do débito coronário (não tem débito constante).
o Compressão extravascular (maior na zona endocárdica)
o Vasos endocárdicos com maior débito na diástole.
o Em condições fisiológicas o débito endocárdico é igual ao débito epicárdico.
Nota: As células do endocárdio têm muitas características semelhantes ao endotélio vascular.
Em situações em que há diminuição da pressão diastólica das coronárias (hipotensão, estenosa
aórtica, oclusão arterial coronária), o débito endocárdico é menor do que o débito epicárdico.
Taquicardia (aumento da frequência cardíaca)
o Encurtamento da diástole;
o Diminuição do tempo de influxo sanguíneo em cada ciclo;
o Compensação pelos vasos de resistência que sofrem vasodilatação.
Bradicárdia (diminuição da frequência cardíaca)
o Aumento da diástole;
o Aumento do tempo de influxo sanguíneo em cada ciclo;
o Compensação pelos vasos de resistência que sofrem vasoconstrição.
2) Neuro-humurais
Estimulação simpática:
a) Aumento marcado do débito coronário;
b) Se eliminarmos os efeitos cronotrópicos e inotrópicos (β-bloqueantes
adrenégicos), o débito coronário diminui.
A acção primária do sistema nervoso central nos vãos de resistência coronários é a
vasoconstrição (estimulação alfa), mas o grande aumento da actividade metabólica sobrepõese causando vasodilatação.
Estimulação Parassimpática
a) Factores metabólicos.
Nota:
Pré-carga -> Pressão durante o enchimento do ventrículo
Pós-Carga -> Pressão arterial cujo ventrículo tem de igualar (para abrir a válvula)
Página 42
Sistema cardiovascular e circulatório
O Sistema Cardiovascular é constituído por:
Coração;
Sistema vascular sanguíneo: artérias elásticas, artérias musculares, arteríoloas,
capilares, vénulas, veias;
Sistema Linfático.
1) Funções:
Transporte de O2, nutrientes, hormonas, etc., a todas as células do organismo.
Transporte de produtos catabólicos para serem removidos
Regulação térmica
Defesa
2) Circulação
A circulação ocorre em vasos:
Artérias: elásticas, musculares;
Arteríolas;
Capilares: contínuos, fenestrados e sinusóides;
Vénulas;
Veias.
Uma vez que as veias têm maior calibre, existe mais sangue no sistema venoso que no
articular, sendo o primeiro sistema de vasos um de capacitância.
Volémia - quantidade de sangue no interior dos vasos. Se esta cair abruptamente, assim como
se tal ocorrer com a pressão, a constrição dos vasos contraria o seu efeito.
A função da circulação é manter um ambiente apropriado para o funcionamento dos diversos
tecidos, através do transporte de nutrientes, hormonas, oxigénio, etc.
A circulação divide-se em circulação pulmonar (pequena circulação) e circulação sistémica
(grande circulação ou circulação periférica).
Partes funcionais da circulação:
A função das artérias é transportar sangue sob elevada pressão, para os tecidos. Por esta
razão, têm paredes vasculares fortes, fluindo o sangue a alta velocidade. As arteríolas
funcionam como controladores da passagem de sangue para os capilares. Têm paredes
musculares fortes que permitem que a arteríola encerre completamente ou dilate, permitindo
desta forma controlar a passagem de sangue.
A função dos capilares é troca de fluidos, nutrientes, electrólitos, hormonas e outras
substâncias com os diferentes tecidos. Por causa desta função as paredes dos capilares são
finas e porosas.
Página 43
As vénulas recolhem o sangue proveniente dos capilares e conduzem-no através de veias de
calibre crescente. As veias funcionam como condutas para o transporte de sangue até ao
coração. Também serve como reservatório de sangue. Por causa da baixa pressão no sistema
venoso, as paredes das veias são finas. Mas apesar disso, têm músculo suficiente para
funcionar como uma reserva de sangue controlado.
3) Leis Gerais da Circulação
a) Lei da Pressão
A pressão que o sangue exerce na parede de um vaso é directamente
proporcional ao débito cardíaco e à resistência oferecida:
8𝜂𝑙
Δ𝑃 = 𝑄 ∗ 𝑅; 𝑄 = Δ𝑉 ∗ 𝑓; 𝑅 = 4
𝜋𝑟
Q Débito Cardíaco (Fluxo total da circulação);
ΔV Volume Sistólico (volume expulso por cada contracção ventricular);
f Frequência Cardíaca
R Resistência Oferecida
η Viscosidade do fluido
l Comprimento do vaso
r Raio do vaso
2 tipos de Fluxo: fluxo laminar – quando uma partícula está numa trajectória,
segue essa trajectória até ao fim. Neste fluxo, as diferentes trajectórias (linhas
de corrente) nunca se cruzam; fluxo turbulento – em que as trajectórias se
cruzam repetidamente e aparecem remoinhos.
o A tendência para se ter fluxo turbulento aumenta com a velocidade do
sangue v, diâmetro do vaso d e diminui com a viscosidade η:
𝑣. 𝑑. 𝜌
𝑟𝑒 =
𝜂
re é o número de Reynolds. Acima de 200-400, há fluxo turbulento.
À saída do coração o traçado da pressão é pulsátil
Pressão sistólica: Máxima
Pressão diastólica: Mínima
A pressão capilar já possui um traçado contínuo. Existem duas razões:
o Efeito de Windkessel
Encerramento de válvulas nas artérias durante a diástole de modo a
impedir o regresso do sangue ao coração. O sangue é forçado a
avançar para os capilares, perdendo a pulsatibilidade.
o Resistência oferecida pelas artériolas e esfíncter pré-capilares que
diminuem a pressão, tornando o fluxo contínuo.
b) Lei da Velocidade
A soma das secções dos ramos é superior à do tronco que as originou;
Página 44
Quanto maior a soma das secções, menor a velocidade do fluido que as
atravessa;
A velocidade é máxima à saída do coração e diminui até aos capilares onde é
mínima. No território venoso a capilaridade diminui, pelo que a velocidade
aumenta.
A velocidade nas artérias é superior à velocidade nas veias.
c) Lei do débito
Como o fluxo sanguíneo é constante, para qualquer secção transversal
completa, passam iguais quantidades de sangue em intervalos de tempo
iguais.
4) Princípios que controlam as funções do Sistema Cardiovascular
a) O fluxo de sangue para cada tecido é quase sempre controlado precisamente pela
necessidade do próprio tecido, através de dilatação ou constrição dos diferentes vasos
sanguíneos.
b) Em geral, a pressão arterial é controlada independentemente do controlo do fluxo do
sangue local ou do controlo do débito cardíaco. Se por exemplo, a pressão baixar
significativamente abaixo dos 100mmHG, dentro de segundos o sistema circulatório
sofre modificações para aumentar a pressão:
i) Aumenta a força de contracção do coração.
ii) Causa a contracção dos reservatórios venosos de sangue, para aumentar a
quantidade de sangue que chega ao coração.
iii) Causa constrição de grande parte das arteríolas, de modo que se acumule mais
sangue nas artérias de maior calibre, aumentado a pressão arterial.
Os rins após um maior período de tempo libertam hormonas que controlam a pressão, e
regulam o volume sanguíneo.
Pressão sanguínea – Significa a força exercida pelo sangue sobre uma determinada área de
parede de um vaso. À força chamamos tensão. A pressão média pode ser medida segundo a
fórmula:
𝑃𝑀 =
2𝑃𝑠 + 𝑃𝑑
3
Resistência à passagem do sangue – Não pode ser medida directamente. Tem de se medir o
fluxo e a diferença de pressões num determinado vaso, e utilizada a relação. 𝑅 =
∆𝑃
𝐹
Ondas de Pulso
Quando se dá a ejecção de sangue, intermitentemente, do coração para a aorta, esta enche e
gera-se uma onda de pressão e de débito que se vai propagar através das paredes de todas as
artérias, muito mais rapidamente do que o sangue (tem uma velocidade cerca de 10 vezes
Página 45
maior que a do sangue). A velocidade das ondas de pulso depende do módulo da elasticidade
das artérias e depende portanto, da idade do indivíduo: em indivíduos mais idosos, as artérias
são mais rígidas, e portanto, a velocidade é maior.
A velocidade das ondas de pulso aumenta em caso de doença que diminua a elasticidade dos
vasos.
A onda de pulso vai sofrendo alterações à medida que se desloca do centro para a periferia.
Estas alterações têm como base o aumento gradual da rigidez das artérias mais periféricas
(devido ao aumento do ratio espessura da parede/diâmetro do vaso) em relação às artérias
centrais (em que este ratio é menor). Assim é de esperar que em artérias progressivamente
mais rígidas aumente o declive do ramo ascendente da onda de pulso:
Aumenta o valor do pico de pressão máxima (sistólica);
Diminui o valor de pressão diastólica.
É de referir que não é a pressão arterial que aumenta à medida que caminhamos para as
periferias, mas sim a pressão de pulso.
A onda de pulso é então afectada pela distensibilidade das artérias e débito cardíaco.
5) Mecanismos de comtrolo da pressão arterial e débito cardíaco e hipertensão
arterial
Medição da Pressão Arterial. (PA)
A medição da pressão arterial pode ser avaliada de forma directa através da introdução de um
cateter conectado a um transdutor de pressão. Conduto, a PA determinada, por rotina, de
forma indirecta, utilizando um esfigmomanómetro. Este aparelho é composto por um braçal
com um balão insuflável no seu interior, por um manómetro de pressão (coluna de Hg) e por
uma bomba de ar com uma válvula.
A pressão arterial média é uma variável fisiológica fundamental porque representa a média da
pressão efectiva que conduz o sangue aos órgãos sistémicos. A pressão arterial média é dada
por: 𝑃𝐴𝑚 = 𝐷𝐶 × 𝑅𝑝𝑒𝑟𝑖𝑓é𝑟𝑖𝑐𝑎 em que DC= débito cardíaco = volume sistólico * frequência
cardíaca
Pressão de pulso = Psistólica-Pdiastólica
Regulação
A regulação da pressão arterial prende-se, como já foi referido com a frequência cardíaca, com
o volume sistólico e com a resistência dos vasos. Mas existem mecanismos mais básicos, que
controlam os referidos e são divididos em regulação nervosa e humoral.
Página 46
Regulação nervosa
O sistema nervoso controla a circulação através do sistema nervoso autónomo (principalmente
o sistema nervoso simpático). O sistema parassimpático também contribui, especificamente na
regulação da actividade cardíaca.
Nota:
Sistema nervoso simpático –> aumenta a actividade cardíaca.
Sistema nervoso parassimpático –> diminui a actividade cardíaca.
Uma das principais funções do controlo nervoso da circulação é a sua capacidade de aumentar
rapidamente a pressão arterial. Para este fim, todas as funções vasoconstritoras e
cardioaceleradores são actividadas. Ao mesmo tempo há inibição da acção do sistema
parassimpático. Assim, ocorre 3 grandes mudanças simultaneamente:
i.
ii.
iii.
Constrição de quase todas as arteríolas do corpo, aumentando a resistência periférica
total, e portanto, aumentando a Pressão Arterial.
Há constrição das veias e de outros grandes vasos da circulação. Deste modo, há maior
quantidade de sangue a chegar ao coração, e portanto, por aumento do débito cardíaco,
aumenta-se a Pressão Arterial.
O coração é directamente estimulado pelo sistema nervoso autónomo, aumentando a
frequência cardíaco e a contractilidade do coração, aumentando o volume de sangue
ejectado a cada sístole. Daí que o débito cardíaco aumente e consequentemente a pressão
arterial.
Além das funções aliadas ao exercício e stress (em que o S.N. aumenta a pressão arterial),
existe outros mecanismos de controlo nervoso, que estão sempre activos, mantendo a pressão
arterial controlada.
Sistema de controlo Baroreceptor de pressão arterial:
Funciona pelo chamado reflexo baroreceptor e por feedback negativo. Um aumento de
pressão arterial faz com que baroreceptores localizados e pontos específicos das paredes de
grandes vasos transmitam sinais para o sistema nervoso central. Sinais de “feedback” são
então enviados através do sistema nervoso autónomo para a circulação, diminuindo a pressão
arterial para o nível normal.
Principais baroreceptores:
Nos grandes vasos, destacando-se baroreceptores da aorta;
Baroreceptores localizados na bifurcação das carótidas.
Reflexo circulatório iniciado pelos baroreceptores:
Quando PA aumenta:
São enviados sinais para a medula
Excitação do centro parassimpático
Página 47
Dilatação das veias e arteríolas na circulação periférica (Redução da
resistência periférica)
Diminuição da frequência cardíaca e contractabilidade do coração
(diminuição do débito cardíaco)
Uma diminuição da pressão arterial terá o efeito contrário, traduzindo-se num aumento da
pressão arterial através da estimulação do sistema nervoso simpático.
Controlo da Pressão Arterial através dos quimio-receptores carotídeos e aórticos:
Associado ao controlo da pressão por baroreceptores, existe um reflexo de quimioreceptores
que funciona basicamente da mesma forma que o reflexo baroreceptores, excepto que são os
quimioreceptores a iniciar.
Quimioreceptores
Células quimiosensíveis, detectam falta de O2 e excesso de CO2 e H+.
Ex: Corpos carotídeos e corpos aórticos.
Tão eficazes como o reflexo baroreceptor
Sempre em contacto com o sangue arterial
Quando PA baixa:
Diminui a quantidade de O2 e aumenta a de CO2 e H+ que não são
removidos
São emitidos sinais que excitar o centro vasomotor (SNA simpático)
Aumento da PA
Regulação humoral
Nos vários órgãos de todos os sistemas, e também no sistema cardiovascular, existem diversos
receptores que possibilitam o controlo pelo sistema nervoso autónomo. Verificam-se, no
sistema cardiovascular:
Receptores colinérgicos;
Receptores adrenérgicos:
o Alfa (vasos sanguíneos):
alfa 1 adrenérgicos - são os principais do sistema cardiovascular,
existindo em quase todos os vasos sanguíneos, causando a
vasoconstricção;
alfa 2 adrenérgicos - existem em menos vasos e potenciam a
vasodilatação;
o Beta (coração e brônquios):
beta 1 - existem em grande número no coração, aumentam a força de
contracção, a velocidade e frequência e a condução nervosa, sendo
muito importantes no aumento da pressão arterial;
beta 2 - causam broncodilatação.
Página 48
Significa controlo através de substâncias segregadas ou absorvidas nos fluidos corporais
(Hormonais, iões entre outros). Algumas destas substâncias são formadas por glândulas
especiais e transportadas para todo o corpo através da circulação.
As mais importantes são:
Agentes vasoconstritores:
o Norepinefrina e epinefrina –> a primeira é um potente vasoconstritor,
enquanto o segundo é mais fraco, causando vasodilatação em alguns tecidos.
(é a epinefrina que é responsável pela vasodilatação das artérias coronários
durante um aumento da actividade cardíaca). A norepinefrina aumenta a
actividade cardíaca e causa a constrição das veias e arteríolas.
o Angiotensina II –> É dos mais potentes vasoconstritores do organismo. Causa
a constrição de pequena arteríolas por todo o corpo, aumentando assim a
resistência total periférica, e portanto, aumenta a pressão arterial.
o Vasopressina -> Um dos mais potentes vasoconstritores do organismo (mais
potente que a angiotensina II). Consegue aumentar grandemente a pressão
arterial e promove a reabsorção de água nos túbulos renais, sendo portanto
considerados uma hormona anti-diurética.
Controlo vascular por iões:
o O aumento da [Ca2+] causa vasoconstrição. Isto porque o cálcio promove a
contracção do músculo liso.
o Aumento da [K+] causa vasodilatação, uma vez que inibe a contracção do
músculo liso.
o Aumento da [Mg2+] causa vasodilatação, uma vez que inibe a contracção do
músculo liso.
6) Controlo da Pressão pelo sistema Renina-Angiotensina.
Renina é uma proteína enzima libertada pelos rins, quando a pressão arterial diminui muito,
para a circulação, permanecendo parte dela nos rins para iniciar diversas funções intrarenais.
A renina é uma enzima e não uma substância vasoactiva. Ela vai actuar sobre uma globulina
denominada angiotensinogénio libertando um polipéptido com 10 aminoácidos: angiotensina
I. A angiotensina I tem algumas propriedades vasoconstritoras, mas nada de significativo.
Segundos a minutos, após a formação da angiotensina I, dois aminoácidos são retirados da
cadeia da cadeia, formando a angiotensina II, um potente vasoconstritor. Esta conversão
ocorre nos pulmões, enquanto o sangue passa nos capilares pulmonares, e é catalisada pela
enzima de conversão presente no endotélio dos vasos dos pulmões.
A angiotensina II é rapidamente degradada, mas antes tem dois principais efeitos:
Causa vasoconstrição de muitas arteríolas em todo o corpo.
Aumento do volume de líquido extracelular, por diminuição da excreção de sais e
água.
Página 49
Diminuição da Pressão Arterial
Produção de Renina pelos rins
Angiotensinogénio (substrato de renina)
Angiotensina I
Enzima (pulmão)
Angiotensina II
Vasoconstrição
Retenção renal de sais e água
Aumento da Pressão Arterial
Estes dois efeitos aumentam a pressão arterial.
A angiotensina II promove ainda a libertação de aldosterona (produzida no córtex da glândula
supra renal) que actua sobre o tubo contornado distal do rim, promovendo a reabsorção de
água e sais, aumentando a volémia e o volume sistólico.
Para além desta regulação, os rins podem ainda, tendo o mesmo efeito sobre a pressão
arterial, ser regulados de outra forma:
Este sistema renal para controlo do PA é muito simples: quando o corpo contém muito líquido
extracelular, o volume de sangue e a pressão arterial aumenta. Este aumento de pressão, por
sua vez, tem um efeito directo nos rins, causando uma maior excreção de líquidos, fazendo a
pressão arterial regressar ao normal.
O contrário traduz-se numa diminuição de excreção, aumentando o volume de sangue e
consequentemente, e pressão arterial.
O aumento do volume urinário, em resposta a um aumento de pressão arterial, é um
fenómeno chamado pressão diurética.
O mecanismo geral pelo qual um aumento do volume de líquido extracelular se traduz num
aumento da pressão arterial é o seguinte:
Aumento do volume de líquido extracelular.
Aumento o volume de sangue.
Aumenta a pressão circulatória média.
Aumenta o retorno de sangue venoso para o coração.
Página 50
Aumenta o débito cardíaco.
Aumenta a pressão arterial.
É de referir que para passar do 5) para o 6) existe duas vias:
Efeito directo do débito cardíaco aumentado, que aumenta a pressão arterial.
Efeito indirecto, que aumenta a resistência vascular periférica, através de autoregulação do fluxo sanguíneo, isto é, por aumento da volémia, causa a constrição dos
vasos sanguíneos, ao longo de todo o corpo, que causa um aumento da resistência
periférica.
Lembrando que PA=DC × Rperiférica. Um aumento do volume de líquido extracelular causa um
aumento da Pressão Arterial.
Mecanismos de controlo local do fluxo sanguíneo:
O controlo local pode ser dividido em duas fases:
I.
Controlo agudo: mudanças rápidas em vasodilatação ou vasoconstrição local das
arteríolas, metarteríolas (terminais as arteríoldas e início dos capilares) e esfíncteres précapilares, ocorrendo entre segundos a minutos, para providenciar um fluxo de sangue
local apropriado.
II.
Controlo a longo prazo (“long-term”): mudanças controladas, lentamente do fluxo, num
período de dias, semanas ou até mesmo meses. Geralmente, estas modificações de longo
prazo permitem um melhor controlo do fluxo em proporção com as necessidades dos
tecidos. As mudanças resultam num aumento ou diminuição no tamanho físico do vaso e
do número real de vasos sanguíneos que “alimentam” os tecidos.
Nota: quanto maior a taxa de metabolismo ou menos oxigénio disponível (ou outro
nutriente necessário ao tecido), verifica-se um aumento da formação de substâncias
vasodilatadoras nas células do tecidos. Estas substâncias actuam nas arteríolas,
metarteríolas e esfíncteres pré-capilares, causando dilatação. Exemplos de substâncias
dilatadoras: adenosina, dióxido de carbono, compostos de adenosina fosfato, histamina.
Desenvolvimento de uma circulação colateral:
É um fenómeno de controlo de longo prazo. Quando uma artéria ou veia é bloqueada, um vaso
novo desenvolve-se à volta do bloqueio para permitir a passagem de uma pequena quantidade
de sangue.
Página 51
Desta forma, as necessidades do tecido são satisfeitas. A primeira fase é dilatação dos
pequenos arcos vasculares que fazem conexão acima do bloqueio. (controlo agudo) Dá-se uma
maior abertura através do controlo a longo prazo.
Electrocardiograma
1) Composição
O electrocardiograma normal é composto por uma onda P, um complexo QRS e uma onda T.
Onda P Causada pelos potenciais eléctricos causados pela despolarização auricular.
Complexo QRS Causado pelos potenciais eléctricos causados pela despolarização
ventricular.
Onda T Causado pelos potenciais eléctricos gerados pela repolarização ventricular.
A onda P é causada pela propagação da despolarização através das aurículas, e é seguido pela
contracção auricular, o que causa um aumento na pressão auricular.
O complexo QRS surge como resultado da despolarização dos ventrículos, iniciando assim a
contracção auricular, o que causa o aumento da pressão ventricular. Deste modo, o complexo
QRS começa ligeiramente antes da sístole ventricular.
A sístole designa o fenómeno de contracção do músculo cardíaco, e a diástole o fenómeno de
relaxamento. São usados os seguintes prefixos com estes termos: “proto” – início, “meso”meio e “tele” – fim.
Nota 1: Quando o impulso cardíaco passa através do coração, a corrente eléctrica atravessa os
tecidos adjacentes e uma pequena porção chega às superfícies do corpo, a qual pode ser
captada por eléctrodos, criando assim um electrocardiograma.
Nota 2: A repolarização auricular também gera uma onda de repolarização T, mas é “tapada”
pelo complexo QRS de bastante maior intensidade. Daí que raramente é captado no
electrocardiograma.
2) Localização dos eléctrodos:
Derivações pré-cordiais de Wilson (unipolares)
V1 -> 4º Espaço Inter Costal (EIC) junto ao bordo esternal direito.
V2 -> 4º EIC junto ao bordo esternal esquerdo.
Página 52
V3 -> Equidistante de V2 e V4.
V4 -> 5º EIC, junto à linha média-clavicular.
V5 -> 5º EIC, ao nível da linha axilar anterior esquerda.
V6 -> 6º EIC, ao nível da linha axilar média esquerda.
Derivações bipolares dos Membros (DI, DII e DIII)
Braço Direito: fio Vermelho
Braço Esquerdo: fio Amarelo
Perna Esquerdo: fio Verde
Perna Direita: Fio Preto (Terra)
As derivações bipolares dos membros entre o braço direito, braço esquerdo e perna esquerda
formam um triângulo à volta do coração conhecido por Triângulo de Einthoven.
a) Braço direito
b) Braço esquerdo
c) Perna esquerda
A amplitude é positiva se a despolarização se desloca em
direcção ao eléctrodo positivo.
Lei de Einthoven:
Estabelece simplesmente que se os potenciais eléctricos de duas das três derivações bipolares
dos membros forem conhecidos, em qualquer momento, o da terceira pode ser determinado
automaticamente pela simples soma dos dois primeiros, tendo em conta o sinal positivo ou
negativo da derivação.
Derivações aumentadas de Golberg (aVR, aVF, aVL) são derivações unipolares medidas em
relação a uma terra virtual (central terminal de Wilson). São o registo aumentado dos
potenciais eléctricos registados nos membros.
aVR –> fio vermelho (braço direito)
aVL –> fio amarelo (braço esquerdo)
aVF –> fio verde (Perna esquerda)
Sistema de referência Hexoaxial:
Derivações do plano frontal:
I, II, III, aVR, aVL e aVF
Página 53
Neste sistema, as seis derivações dos membros têm uma origem comum. As 6 linhas (metade
contínua, positivo, metade negativa, descontínua) estão separadas entre si 30º.
Determinação do eixo eléctrico do Coração:
Pode determinar-se a partir de duas derivações bipolares dos membros. O eixo anatómico do
coração é o ângulo do coração no corpo, desde a base até ao ápice. O eixo eléctrico não é
sinónimo de posição anatómica do coração e representa a direcção principal do fluxo da
corrente eléctrica gerada durante a despolarização ventricular.
Obtém-se da seguinte forma:
. Obter o registo do ECG em duas derivações bipolares dos membros (Ex: DI e DII)
. Determinar a amplitude do complexo QRS, e também a polaridade.
. Marcar os valores obtidos nos respectivos eixos das derivações. Perpendiculares de cada eixo
são traçados nestes pontos. O ponto de intersecção destas duas perpendiculares representa a
direcção do vector resultante.
. O comprimento do vector representa o potencial eléctrico médio gerado pelos ventrículos
durante a despolarização e a direcção do vector, o eixo eléctrico do coração.
Página 54
Sistema Respiratório
1) Funções:
Ventilação – trocas gasosas a nível pulmonar (vulgarmente designado por
respiração, uma vez que não implica consumo de O2 a nível celular);
Trocas gasosas de O2 (essencial à função celular) e CO2 (gás final das reacções de
metabolismo celular);
Transporte: de O2 e CO2 desde da atmosfera até pulmões e depois ao nível
sanguíneo.
Intercâmbio: O2 fornecido e CO2 devolvido Hematose
Regulação do pH: os níveis de CO2 do sangue estão associados ao seu pH
Fonação: o som é produzido nas cordas vocais através da passagem de ar
Olfacto/Paladar: quando moléculas em suspensão atravessam a cavidade nasal ou
a língua produzem a sensação de cheiro, que também está associada ao paladar
Defesa contra agentes patogénicos e irritantes: o ar é filtrado quando passa nas
vias respiratórias
Reservatório de sangue
Regulação da concentração hormonal no sangue
Aquecimento: à medida que passa do aparelho respiratório o ar é aquecido e
misturado
Árvore Traqueobrônquica: todo o seu epitélio é ciliado, de modo a remover secreções e
corpos estranhos do conducto aéreo, que passam para o sistema digestivo. Outra função é
aquecer e humidificar o ar atmosférico, uma vez que as trocas ao nível dos alvéolos são feitas
na presença de vapor de água.
Traqueia Brônquios Brônquíolos Brônquíolos Terminais
- zona condutora
Sacos Alvéolares Ductos Alvéolares Brônquíolos Respiratórios
- zona respiratória
2) Mecânica Pulmonar
Barreira Alvéolo-Pulmonar: é muito importante porque é onde se dão as trocas gasosas entre
o ar dos alvéolos, situados na parte mais profunda do pulmão (no parênquima pulmonar), e o
sangue; é formada pelo epitélio dos alvéolos, o espaço intersticial e o endotélio dos capilares
(camada única). Os capilares formam uma rede de tal forma densa sobre o pulmão, que é
quase como se existisse uma camada de sangue em torno do mesmo.
Movimentos respiratórios criam diferenças de pressão:
Inspiração:
Diafragma contrai por acção do nervo pélvico; contrai e vem para baixo; as
costelas e o externo são levantados por acção do diafragma e dos músculos
intercostais externos;
Diâmetro lateral transversal da caixa toráxica aumenta assim como o anteroposterior;
Maior volume da caixa toráxica e menor pressão intrapleural;
Página 55
Pulmões são expandidos.
Expiração:
Relaxamento de todos os músculos da caixa toráxica;
Menos volume da caixa toráxica e maior pressão pleural e alveolar;
Ar sai para o exterior.
A pleura é constituída por dois folhetos entre os quais existe o líquido pleural que
funciona como lubrificante permitindo a expansão da caixa torácica.
Para que exista ventilação, é necessário que se crie uma diferença de pressão entre o
ar exterior e os alvéolos. Onde na Inspiração Palv.<Pext. e na Expiração Palv.>Pext.
A pressão pleural é negativa de modo a haver entrada de ar nos pulmões. No inicio da
inspiração a pressão é cerca de (-5cm H2O) e quando o volume pulmonar aumenta,
esta decresce aos (-7,5 cm H2O). Na expiração, o processo é inverso.
Existe também uma pressão alveolar, nula quando a glote está fechada. Na inspiração
a pressão alveolar cai para (-1 cm H2O), forçando a entrada de 0,5L de ar nos pulmões.
Na expiração esta aumenta para 1 cm H2O, expulsando igual quantidade de ar.
o A diferença entre a pressão pleural e alveolar designa-se de pressão
transpulmonar e é uma medida de força elástica dos pulmões. Esta pressão
aumenta na inspiração.
A Compliance pulmonar relaciona a variação de volume com a variação da pressão
transpulmonar. 𝐶 =
𝑑𝑉
𝑑𝑃
. Esta grandeza depende da elasticidade dos tecidos das
propriedades tensoactivas dos surfactantes e do volume pulmonar.
Nos alvéolos existem os pneumócitos tipo II que produzem o surfactante pulmonar.
Este baixa drasticamente a tensão superficial impedindo o colapso alveolar, mantém
os alvéolos secos, diminui o trabalho na dilatação dos pulmões e previne a passagem
de líquidos dos capilares para o espaço alveolar.
3) Espirometria e ventilação pulmonar
A espirometria mede o volume de ar que
entra/sai dos pulmões durante a
inspiração/expiração. Com este é possível
medir os diferentes volumes pulmonares. O
indivíduo respira para uma peça bucal; temse água e uma campânula com O2, donde
ele extrai oxigénio, libertando anidrido
carbónico; quando este volume varia, um
sistema de roldanas faz variar o registo num
cilindro e portanto observam-se as
variações de volume que se deram durante
a respiração.
Página 56
a) Volumes pulmonares
Volume corrente (VC – tidal volume): Volume de ar mobilizado por uma pessoa em
repouso (500 mL).
Volume de Reserva Inspiratória (VRI – IRV(inglês)): Volume de ar que pode ser
inspirado profundamente depois de uma inspiração normal (3000 mL)
Volume de Reserva Expiratória (VRE – VER): Volume de ar que pode ser expirado
depois de uma expiração em repouso (1100 mL)
Volume Residual (VR – RV): Volume de ar que permanece nos pulmões depois de uma
expiração máxima. Não pode ser medido por espirometria (1200 mL).
b) Capacidades pulmonares
Capacidade Vital (CV – VC) = VRI + VC + VRE
Capacidade Inspiratória (CI – IC) = VRI + VC
Capacidade Residual Funcional (CRF – FRC) = VR + VRE
Capacidade Pulmonar Total (CPT) = VRI + VC + VRE + VR
c) Débito Ventilatório: Grandeza que mede o volume de ar mobilizado por unidade de
tempo. É o produto do volume Inspiratório (ou Expeiratório) pela frequência/minuto.
4) Difusão gasosa na membrana pulmonar
A membrana respiratória (colectivo das membranas) é constituída por:
Camada de surfactante pulmonar no alvéolo
Epitélio alveolar (pavimentoso simples)
o Pneumócitos tipo I – células epiteliais de revestimento, muito finas e
através das quais se efectuam as trocas gasosas;
o Pneumócitos tipo II – células glandulares, menores e mais grossas,
possum microvilosidades dirigidas ao interior do alvéolo que
sintetizam surfactante (contém fosfolípidos e lipoproteínas)
o Macrófagos alveolares - células do sistema imunitário, fagocitam
organismos estranhos que não foram envolvidos pelo muco ou
bloqueados por pêlos e cílios;
Membrana epitelial base
Espaço intersticial entre a membrana base e a membrana capilar
Membrana capilar base que se funde em muitos locais com a alveolar base
Membrana capilar endotelial
o A troca gasosa dá-se por um processo de difusão simples (não
mediado e a favor do gradiente de concentração), tanto a nível dos
alvéolos como das restantes células.
Página 57
o
o
o
A pressão de CO2 nos alvéolos é superior à do ar porque há mistura do
ar inalado com o do espaço fisiológico morto [soma do espaço morto
anatómico (é o ar existente nas vias condutoras que não participa nas
trocas gasosas) e o alveolar (ar que não participa das trocas existente
nas áreas de trocas gasosa dos pulmões: próximo a 0 nos indíviduos
normais)] resultante do ciclo anterior;
Os capilares pulmonares são muito finos, pelo que os eritrócitos têm
de se “espremer” para passarem neles. Assim, é comum a membrana
que contem glóbulos vermelhos tocar a parede capilar, para que o O2
e CO2 não precisem de atravessar quantidades significativas de
plasma, o que provoca um aumento da rapidez da difusão.
Existe uma relação entre o ritmo de ventilação alveolar e as pressões
parciais dos gases.
Se P(O2) aumentar, o ritmo aumenta
Se P(CO2) diminuir, o ritmo aumenta
Se a produção de CO2 das células
aumentar, a ventilação alveolar
aumenta para manter a pressão
Se o consumo de O2 pelas células
aumentar, a ventilação aumenta
para manter a pressão
Assim sendo a concentração e pressão dos dois gases nos
alvéolos são determinadas pelas taxas de absorção/excreção
dos gases e também pela ventilação.
Taxa de difusão influenciada por:
o Espessura da membrana
o Área da difusão
o Coeficiente de difusão
o Gradiente de concentração
5) Transporte de gases
a) Transporte de O2
O2 pode ser transportado ao dissolver-se no plasma ou ao ligar-se à
hemoglobina através de Ferro formando um composto (Oxihemoglobina)
A pressão de O2 varia ao longo da circulação
Página 58
A pressão de O2 nos capilares pulmonares aumenta até igualar a dos
alvéolos (104 mmHg)
A pequena redução da pressão na passagem ao sistema arterial deve-se à
mistura do sangue arterial com o venoso que alimentou previamente o
tecido pulmonar
A pressão a nível dos capilares sistémicos é de 95 mmHg, muito superior à
do tecido circundante, favorecendo a difusão de O2 para este. A pressão
cai até aos 40 mmHg
A saturação de hemoglobina e a pressão de O2 estão relacionados:
O nível de saturação aumenta com a PO2
A nível das artérias sistémicas a PO2 é de cerca de 95 mmHg, conduzindo a
uma saturação de 97%
A nível do sangue venoso, a PO2 é aproximadamente 40 mmHg,
representado uma saturação de 75%.
A afinidade do O2 à hemoglobina também depende de:
Temperatura – enfraquece as ligações
pH – desnaturação;
DPG (fosfoglicerato) – aumento de DPG reduz a afinidade
PCO2
b) Transporte de CO2
O CO2 (produto do ciclo de Krebs) tem 3 vias de transporte:
Dissolução no plasma
Hemoglobina
(i) Cerca de 23% de CO2 reage directamente com radicais amino da
hemoglobina, formando carboxiemoglobina (CO2Hb). Se existir muita
carboxiemoglobina, a hemoglobina fica saturada de CO2 e não pode
transportar O2
Forma de bicarbonato (mais importante). Este processo engloba os
seguintes passos (este ocorre nos capilares sistémicos):
(i) Dissolução de CO2 no eritrócito
(ii) CO2 + H2O
H2CO3
(iii) H2CO3
H+ + HCO3-
Página 59
(iv) HCO3- passa para o plasma (troca com o Cl -) devido à acção da enzima
anidrase carbónica
(v) H+ liga-se à hemoglobina (base forte)
(vi) Nos capilares pulmonares dá-se o processo inverso até à expiração de
CO2
6) Circulação Pulmonar e Circulação Sistémica:
Note-se que as pressões são muito mais baixas na artéria pulmonar que nas grandes artérias
sistémicas. Na circulação pulmonar, o parênquima (o parênquima é uma camada que cobre os
grandes vasos pulmonares) do pulmão tende a fazer dilatar os vasos e as forças elásticas dos
vasos e as forças de contracção muscular opõem-se, ocorrendo um equilíbrio de forças.
Geralmente o aumento do volume do pulmão provoca dilatação dos vasos e o aumento da
pressão alveolar provoca uma diminuição do diâmetro capilar, o que leva a um colapso.
Pressão crítica de abertura: valor de pressão que a artéria pulmonar tem de atingir para que
se passe a dar a circulação através do pulmão quando este se encontra colapsado.
A variação da pressão pulmonar não é muito grande para um débito normal, mas para valores
de débito elevados aumenta muito e para valores baixos diminui muito.
Zonas 1, 2, e 3 da Circulação Pulmonar:
Zona 1: a pressão arterial pode chegar para que os capilares não colapsem por acção da
pressão alveolar e passe sangue. Parterial > Palveolar > Pvenosa
Se a pressão arterial baixar por hemorragia, ou se a pressão alveolar aumentar porque
demos O2 ao individuo o capilar colapsa e não há irrigação Espaço morto alveolar
Zona 2: Pvenosa < Ppulmonar, o que vai controlar a circulação é a resistência.
𝑆 = 𝑃𝑣𝑒𝑛𝑜𝑠𝑎 − 𝑃𝑎𝑙𝑣𝑒𝑜𝑙𝑎𝑟 Apenas existe fluxo de sangue nos picos de pressão arterial,
porque a pressão sistólica é maior que a pressão alveolar, mas a pressão diastólica é
inferior à alveolar.
Zona 3: o fluxo é continuo porque a pressão capilar é sempre maior que a alveolar durante
todo o ciclo cardíaco.
Nota: As situações normais são a 2 e 3. Valores muito elevados de pressão nas artérias
pulmonares causam edemas pulmonares.
Modificação do calibre dos vasos pulmonares:
Vasoconstrição Arteriolar :
a) Hipóxia
b) Hipercápnia
c) Acidose Respiratória
d) Noradrenalina
e) Adrenalina
Página 60
f)
Serotonina
Vasodilatação Arteriolar:
a) Isoproterenol
b) Acetilcolina
Vasoconstrição de veias e vénula:
a) Serotonina
b) Histamina
c) Aloxana
Vasodilatação em pulmões isolados:
a) Aminofilina
b) Nitritos
c) Papaverina
7) Regulação da Respiração
a) A respiração é controlada por um conjunto de 6 neurónios:
Neurónios inspiratórios precoces – são estimulados antes da respiração
e param antes do fim desta
Neurónios da rampa inspiratória – começam antes da inspiração e
mantém-se após a paragem dos neurónios respiratórios precoces. Não
acabam a inspiração.
Neurónios da rampa inspiratória tardia – estimulados perto do fim da
actividade dos neurónios da rampa inspiratória. Terminam a inspiração.
Neurónios pós-inspiratórios – têm grande actividade e são os grandes
responsáveis na 1ª fase da expiração
Neurónios da rampa expiratória – Iniciam a actividade durante a
inspiração e continuam na 2ª fase da expiração
Neurónios pré-inspiratórios – Iniciam a actividade antes dos neurónios
inspiratórios precoces.
b) Quimiorreceptores
i)
Centrais:
Mitchell
Schlaefler
Loeschke
ii) Periféricos
Corpos aórticos
Seio carotídeo
Detectam alterações de pH
iii) Pulmonares (regulam a respiração mecânica)
Página 61
Receptores de estiramento pulmonar (Reflexo Hering – Bauer – Desligam a
rampa inspiratória/expiratória quando os músculos atingem o limite)
Receptores de irritação – Provocam tosse/espirro;
Receptores C (proteína transmembranar cujo terminal-N está localizado
extracelularmente).
Receptores do nariz e vias aéreas superiores – espirro/tosse
Receptores J – Activos quando existe um excesso de líquido nos alvéolos.
Induzem hiperventilaçºão e drenagem linfática
Receptores articulares e musculares – induzem uma respiração mais funda no
exercício físico.
Dor e Temperatura Aumentam a frequência respiratória
Baroreceptores arteriais Condicionam a respiração
Sistema Gama – Impede o rompimento das fibras do diafragma ao parar a
inspiração profunda
c) Regulação humoral
A frequência respiratória aumenta com o aumento de PCO2 e diminuição de
PO2
Os quimioreceptores periféricas são mais sensíveis à PO2 que os centrais
d) Regulação Reflexa
Seios aórticos e carotídeos (é dominante quando o volume sanguíneo
diminui)
Aumento da pressão sanguínea inibe o centro respiratório
Aumento de O2 inibe o centro respiratório
Zona de reflexo de Bainbridge (é dominante quando o volume sanguíneo
aumenta)
Aumento da pressão sanguínea estimula o centro respiratório
Contrário aos seios aórticos e carotídeos
e)
Resposta ao exercício:
Exercício físico, baixa pH e PCO2 aumenta estimulando a ventilação
A temperatura aumenta estimulando a ventilação
Página 62
Sangue
Principais funções do sangue:
Respiração
Nutrição
Excreção
Balanço hidroelectrolítico
Balanço ácido-base
Temperatura corporal
Defesa imunológica
Transporte de hormonas e regulação metabólica
Hemostase
Constituintes do sangue:
Plasma - fluido onde circulam electrólitos, nutrientes, metabolitos, vitaminas,
hormonas, gases e proteínas dissolvidas
Elementos figurados
o Eritrócitos (RBC’s), glóbulos vermelhos ou hemácias – transportam o O2 e
desempenham um papel importante na regulação do pH
o Leucócitos (WBC’s), glóbulos brancos
o Plaquetas, trombócitos – necessários à hemostase.
Hematócrito (Hct) é a percentagem ocupada por eritrócitos no volume total do sangue.
As proteínas plasmáticas estão envolvidas na defesa imunitária e regulação da pressão
oncótica, o que mantém o volume sanguíneo constante. Por ligação às proteínas plasmáticas,
compostos insolúveis em água podem ser transportados no sangue e muitas substâncias
podem ser protegidas de se desintegrarem e da excreção renal. A ligação de pequenas
moléculas às proteínas plasmáticas reduz a sua eficácia osmótica. Muitas delas estão
envolvidas na coagulação e fibrinólise. Para além disso ainda intervém na resposta
inflamatória, hormonas, enzimas e antiproteases.
Eritrócitos
A principal função dos eritrócitos é transportar a hemoglobina que, por sua vez, leva o
oxigénio desde os pulmões até aos tecidos Os eritrócitos normais são bicôncavos, apesar de o
seu aspecto poder variar bastante quando passam nos capilares. Os eritrócitos normais têm
um excesso de membrana celular para a quantidade de conteúdo que têm. Por isso, a
deformação destas células não exige um grande estiramento da membrana e,
consequentemente, as células não lisam. A quantidade média de eritrócitos por milímetro
cúbico é de 5 200 000.
Página 63
Os eritrócitos têm a faculdade de concentrar a
hemoglobina. Nas pessoas normais, a quantidade
de hemoglobina está quase sempre próxima do
nível máximo em cada célula. Cada grama de
hemoglobina pode combinar-se com 1039 mm de
O2.
Génese das células sanguíneas. Todas as células
sanguíneas derivam das chamadas células-mãe
hematopoiéticas pluripotentes. A partir da
diferenciação das ditas células vão-se formando as
células sanguíneas circulantes. À medida que estas
células se reproduzem, uma pequena percentagem
permanece exactamente igual às células
pluripotentes originais e ficam retidas na medula
óssea, mantendo-se assim uma reserva constante. A
primeira geração de novas células não se pode
diferenciar como pertencente a um tipo concreto de células sanguíneas, apesar de
já estarem encaminhadas a originar um determinada linha celular: são chamadas
as células mãe comprometidas.
A proliferação e reprodução de diferentes células mãe estão sob o controlo de
múltiplas proteínas chamadas indutores de proliferação, que activam a dita
proliferação mas não a sua diferenciação. Esta activa-se pelo grupo de proteínas
denominadas indutores da diferenciação. Cada proteína indutora provoca a
diferenciação da célula mãe, em um ou mais passos, para formar um dos tipos de
células sanguíneas adultas. A formação de indutores de diferenciação está
controlada por factores externos à medula óssea. No caso dos eritrócitos, a
exposição do organismo a baixas concentrações de O2 durante um período
prolongado estimula a proliferação, a diferenciação e a produção de um número
muito elevado de eritrócitos.
Regulação da produção de eritrócitos: função da eritropoietina
A massa total de eritrócitos no sistema circulatório está regulada entre limites estreitos.
Qualquer circunstância que reduza abaixo do normal a quantidade de oxigénio que se
transporta aos tecidos, aumenta a produção de eritrócitos. O principal factor que estimula a
produção de eritrócitos é a eritropoietina. Esta é formada principalmente nos rins (80-90%)e
no fígado. Quando se removem os dois rins ou uma doença renal os destrói, surge uma
anemia grave, já que a quantidade de eritropoietina que se forma nos tecidos não renais
apenas permite a formação de 1/3 a ½ dos eritrócitos que o organismo necessita.
A vitamina B12 e o ácido fólico são importantes para a maturação final dos eritrócitos. A
falta de qualquer uma destas vitaminas produz uma diminuição na quantidade de DNA
(síntese) e, consequentemente, provoca uma fraqueza na maturação e divisão celular. Para
além disso, os eritrócitos tornam-se maiores que o normal, formando megaloblastos. Estas
Página 64
células têm fórmulas irregulares e uma membrana muito fina: são capazes de transportar O2
normalmente, mas a sua fragilidade diminui o seu tempo de vida a metade.
Formação da hemoglobina
A síntese de hemoglobina começa quando os eritrócitos estão ainda num estado de
proeritroblastos e continua até à fase de reticulócitos, quando as células abandonam a medula
óssea e entram na corrente sanguínea. Quatro cadeias de hemoglobina juntam-se para que
esta se torne funcional. A sua característica mais importante é a capacidade de se unir
reversivelmente com o oxigénio. O átomo de O2 une-se laxamente a uma das ligações de
coordenação do átomo de ferro que existe na hemoglobina.
Anemias
Anemia - défice de eritrócitos e pode dever-se a
uma perda rápida ou a uma produção lenta dos
mesmos.
Anemia por perda de sangue – ocorre
após uma hemorragia grave. O organismo
é capaz de repor o plasma em 1 a 3 dias,
mas a concentração de eritrócitos continua
baixa. Para normalizar o número de
eritrócitos são necessários 3 a 4 semanas.
Anemia aplástica – ocorre por falta função
da medula óssea. Pode dever-se a uma
exposição à radiação gama, alguma
substância industrial química tóxica ou
uma reacção adversa a um fármaco.
Anemia megaloblástica – ocorre por falta de vitamina B12, ácido fólico ou de factor
intrínseco. A carência de alguma das ditas substâncias provoca uma reprodução lenta
dos eritrócitos na medula óssea. Assim os eritrócitos tornam-se megaloblastos.
Anemia hemolítica – ocorre como consequência da fragilidade dos eritrócitos, que
lisam ao passar os capilares. O número de eritrócitos é normal ou maior do que o
normal, no entanto, o seu tempo de vida é muito curto.
Anemia de células falciformes – é um tipo de anemia hemolítica devida à composição
anormal das cadeias de hemoglobina. Quando esta anemia se expõe a uma [O2] baixa,
precipita no interior dos eritrócitos formando cristais, fazendo com que as células
fiquem em forma de foice e sejam muito frágeis.
Resistência do organismo à infecção
Leucócitos, granulócitos, sistema monócito – macrófago, inflamação
Os leucócitos são elementos móveis no sistema de protecção do organismo. Formam-se na
medula óssea e no tecido linfático, e logo se transportam no sangue até às zonas
Página 65
inflamadas, onde realizam uma defesa rápida e poderosa contra os agentes infecciosos
que possam existir. Existem vários tipos de leucócitos:
Neutrófilos
Eosinófilos
Basófilos
Monócitos
Linfócitos
Os três primeiros são polimorfonucleares, têm um aspecto granular pelo que se chamam
granulócitos/”polis”.
Os granulócitos e os monócitos protegem o
corpo de organismos invasores, ingerindo-os
por fagocitose. Os linfócitos actuam geralmente
junto ao sistema imunitário agregando-se de
maneira específica com o organismo invasor e
destruindo-o.
Génese dos leucócitos. A partir das células mãe
hematopoiéticas pluripotentes formam-se
duas estirpes de leucócitos: a linha mielocítica
e a linha linfocítica. Os granulócitos e os
monócitos pertencem à primeira e os linfócitos
à segunda. Os primeiros só se formam na
medula óssea. Os linfócitos produzem-se nos
diferentes órgãos linfáticos, como os gânglios
linfáticos, o baço e o timo. A esperança de vida
dos leucócitos é bastante variável.
Propriedades defensivas dos neutrófilos e dos macrófagos
Os neutrófilos e os monócitos são os principais responsáveis pelo ataque e destruição
das bactérias, vírus e outros agentes nocivos. Os neutrófilos são células maduras que
podem atacar e destruir bactérias e vírus no sangue circulante. Já os monócitos apenas
atingem maturação nos tecidos e transformam-se em macrófagos tecidulares. São
atraídos para as zonas inflamadas por quimiotaxis. A função mais importante dos
neutrófilos e macrófagos é a fagocitose. Quando ocorre uma lesão, existem
substâncias que circulam no sangue e aumentam a permeabilidade dos capilares,
produzindo-se a saída de grande quantidade de líquido para o espaço intersticial e
uma migração dos granulócitos e monócitos até aos tecidos afectados.
Eosinófilos - Combatem os efeitos da histamina na resposta alérgica; actuam por fagocitose
ou libertação do conteúdo dos seus grânulos.
Basófilos - Libertam heparina, histamina e serotonina, que actuam em reacções alérgicas e
anti-parasitárias.
Página 66
Imunidade inata e imunidade adquirida
A imunidade é a capacidade do corpo para resistir aos ataques de quase todos os organismos e
toxinas que podem causar danos. A maioria dos organismos têm uma imunidade inata, que
consistem em acções comuns, como a fagocitose de bactérias, a destruição de organismos
patogénicos pelas secreções ácidas e enzimas digestivas, a resistência da pele à invasão e a
presença de certos compostos químicos no sangue que atacam e destroem os organismos
estranhos e as toxinas. A imunidade adquirida é a capacidade do corpo para apresentar
mecanismos muito específicos e eficazes contra agentes invasores concretos, como certas
bactérias, vírus e toxinas potencialmente mortais, ou mesmo contra outros tecidos estranhos
provenientes de outros organismos. A imunidade adquirida inicia-se por antigénios. Os
linfócitos são a base deste tipo de imunidade. Podem ser encontrados em vários tecidos
linfáticos que estão distribuídos por todo o corpo, de maneira a poder interceptar organismos
invasores e toxinas que se disseminam rapidamente.
Há duas populações de linfócitos. Os linfócitos T maturam-se no timo e são responsáveis pela
imunidade mediada por células. Os linfócitos B modificam-se no fígado e depois na medula
óssea durante a vida fetal, sendo responsáveis pela imunidade humoral.
O timo processa linfócitos T. Os linfócitos dividem-se rapidamente e distribuem-se pelos
tecidos linfáticos de todo o corpo. O fígado e a medula óssea processam os linfócitos B, que se
diferenciam dos T por segregarem anticorpos activamente, que são grandes moléculas
proteicas que se combinam com o antigénio e destroem-no. A diversidade de linfócitos B é
muito maior que de linfócitos T e uma vez processados migram para todos os tecidos linfáticos
onde se armazenam.
Hemostasia e Coagulação do Sangue
O termo hemostasia significa prevenção da perda de sangue. Quando um vaso se lesiona ou
rompe, a hemostasia consegue-se por meio de: 1) um espasmo vascular; 2) a formação de um
tampão plaquetário; 3) a formação de um coágulo sanguíneo devido à coagulação do sangue, e
4) a proliferação final de tecido fibroso para fechar de maneira permanente a lesão do vaso.
Página 67
Hemostase Primária
Quando há uma lesão vascular, as plaquetas aderem ao colagénio e ao factor Van
Willebranz existente no endotélio do vaso sanguíneo, incham e adquirem formas
irregulares e libertam grandes quantidades de serotonina, tromboplastina e ADP que
atraem outras plaquetas, acabando por se formar o tampão plaquetar. Para evitar a
formação de um tampão demasiado grande, o endotélio liberta algumas substâncias,
como NO e as prostacilinas.
Hematose Secundária
Na hemostase secundária há formação de um coágulo sanguíneo em 3 passos essenciais:
1) Activação sequencial de vários factores de coagulação, muitos deles sintetizados no
fígado e dependentes da vitamina K , originando o complexo activador da protombina
2) O complexo activador cataliza a conversão da protombina em trombina
3) A trombina actua sobre o fibrinogénio convertendo-se em fibras de fibrina que
formam uma rede entre as plaquetas, células sanguíneas e plasma, dando origem ao
coágulo sanguíneo.
Finalmente dá-se a dissolução do coágulo, (quando o tecido está reparado) através da
fibrinólise. A fibrina é degradada pela plasmina, levando à produção de fragmentos circulantes
que são depois destruídos por outras proteinases ou pelos rins e fígado.
O plasminogénio é convertido em plasmina por acção de diversos activadores tecidulares,
como o tPA e o pro UK.
Página 68
Equilíbrio Ácido-Base
Este é um capítulo auxiliar de modo a compreender termos que se são dados ao longo do
tempo. Perguntas específicas sobre isto não saiem.
pH fisiológico: 7,35 – 7,45
Equação de Henderson-Hasselbach
[A− ]
𝑝𝐻 = 𝑝𝐾𝑎 + log (
)
[HA]
[HCO− ]
3
𝑝𝐻𝑓𝑖𝑠𝑖𝑜𝑙ó𝑔𝑖𝑐𝑜 = 6,1 + log (P (CO )∗0,0301
)
a
2
Pa(CO2) – Pressão Alveolar
Acidémia: valor de pH do sangue abaixo do valor de referência 7,35.
Acidose: situação que leva a uma diminuição do pH abaixo de valores fisiológicos.
Pode ser compensada, restabelcendo-se o valor de pH normal mas permanecem
alterações em [HCO3-] e Pa(CO2).
Alcalémia: valor de pH do sangue acima do valor de referência 7,45.
Alcalose: semelhante à acidose mas para valores de pH superiores ao fisiológico.
Alterações primárias do numerador e denominador da equação:
Alterações metabólicas alterações no ião bicarbonato (alterações no numerador):
o [HCO3-] => pH => alcalose
o [HCO3-] => pH => acidose
Valores de [HCO3-] são controlados principalmente ao nível dos rins, onde se dá:
o
o
o
Reabsorção de HCO3-;
Formação de ácidos tituláveis;
Excreção de NH4+ (amónia).
Alterações respiratórias alterações na pressão parcial de CO2 (alterações no
denominador):
o Pa(CO2) => pH => acidose
o Pa(CO2) => pH => alcalose
Pa(CO2) é modificável pela respiração
Na gasimetria arterial medem-se valores de pH do sangue, as concentrações de HCO3-, Pa(CO2)
e Pa(O2), e ainda o “anion gap”.
Anion gap, ou hiato iónico, representa os aniões que não são medidos directamente, como
proteínas aniónicas, fosfatos, sulfatos e aniões orgânicos. O seu valor ronda os 10-12 mmol/L e
calcula-se através da expressão:
Página 69
𝐴𝐺 = 𝑁𝑎+ − (𝐶𝑙 − + 𝐻𝐶𝑂3− )
Na+ ião mais importante medido na gasimetria
Os distúrbios ácido-base podem ser simples (maioria das situações) ou mistos (bastante
complexos e difíceis de diagnosticar). O nosso organismo tem mecanismo de compensação
que tentam manter pH do sangue dentro dos valores fisiológicos.
Nota: Os mecanismos de compensação regulam o pH mas não corrigem os problemas
que levam aos próprios distúrbios ácido-base.
Predições de compensação:
Acidose metabólica
o PaCO2 baixa 1,25 mm Hg por cada 1 mmol/L de [HCO3-] que baixa;
o
1 [HCO3-] => 1,25 PaCO2
Alcalose metabólica
o PaCO2 sobe 0,75 mm Hg por cada 1 mmol/L de [HCO3-] que aumenta;
o
1 [HCO3-] => 1,25 PaCO2
Acidose respiratória
o Aguda: [HCO3-] aumenta 1 mmol/L por cada aumento de 10 mm Hg na PaCO2
o Crónica: [HCO3-] aumenta 4 mmol/L por cada aumento de 10 mm Hg na PaCO2
Alcalose respiratória
o [HCO3-] diminui 2 mmol/L por cada decréscimo de 10 mm Hg na PaCO2
o [HCO3-] diminui 4 mmol/L por cada decréscimo de 10 mm Hg na PaCO2
Causas de acidose metabólica com anion gap aumentado:
Insuficiência renal
mais frequentes
Diabetes
Acidose láctica (desportista e doentes terminais)
Cetoacidose (tipo de acidose metabólica que é causada por altas concentrações
de cetoácidos, formados pela deaminação dos aminoácidos) diabética ou alcoólica
Toxinas (etilenoglicol, metanol, ou salicilatos)
Causas de acidose metabólica sem anion gap aumentado:
Perda gastrointestinal de bicarbonato
o Diarreia
o Drenagem pancreática ou do intestino delgado
o Uretero-sigmoidostomia, ansa jejuanl, ansa ileal
o Fármacos (cloreto de cálcio, sulfato de magnésio, colestiramina)
Acidose Renal
o Distúrbios dos tubos renais
o Aumento do ião K+ (hipercaliémia)
Hipercaliémia (grande [K+] no sangue) induzida por fármacos
Outras
Página 70
Sistema Renal
1. Funções do rim
Filtro – Remove produtos metabólicos e toxinas (excreta ureia, creatinina,
bilirrubina, substâncias tóxicas ingeridas, etc.)
Manutenção do equilíbrio electrolítico e ácido-base – Regula
concentrações iónicas (Na+, K+,…) e regula o pH fisiológico através da
regeneração de HCO3- e eliminação de H+
Produção hormonal:
o Renina – Controlo de pressão arterial a longo/médio prazo
o Vitamina D3 – aumenta o metabolismo (fixação) de Fósforo e
Cálcio
o Eritropoeitina – Estimula a formação de eritrócitos (eritropioese)
o Prostoglandinas e bradiquininas – Controlo da pressão arterial
local
Gliconeogénese
2. Anatomia fisiológica do Rim
Os rins são órgãos retroperitoniais divididos em 2
grandes regiões:
Parte mais periférica é compacta – córtex;
Parte medial (interior) designada por
medula;
A medula está dividida em múltiplas massas de
tecidos em forma de cone denominadas pirâmides
renais. A sua base está na fronteira entre o córtex e a
medula e termina na “papilla”, que se projecta para a
pélvis renal (espaço em forma de túnel) e é a
continuação da parte superior do uréter.
O bordo exterior da pélvis está dividido em cálices maiores e cálices menores que recolhem a
urina e a conduzem ate à bexiga.
Os rins recebem cerca de 20% do débito cardíaco através da artéria renal que entra no rim
pelo Hilo e se vai ramificando progressivamente até formar artérias interlobares, arteríolas
aferentes que se ramificam originando os capilares glomerulares onde se dá a filtração.
Os capilares unem-se para formar a arteríola eferente que origina uma rede de capilares
secundária: os capilares peritubulares.
Página 71
Unidade funcional do rim: nefrónio
A cápsula de Bowman envolve o novelo e é
para onde vai o plasma. O sangue chega ao
nefrónio pelo arteríola aferente e sai pela
arteríola eferente.
São os nefrónios os responsáveis pela
produção de urina. Cada um contém uma
rede capilar (glomérulo) sujeita a elevadas
pressões hidrostáticas, dando-se a
filtração. É aqui que se inicia a produção de
urina.
O filtrado (supostamente sem proteínas)
passa para a cápsula de Bowman. Grande
parte das substâncias no plasma (excepto
as proteínas) são livremente filtradas de
modo que a sua concentração no filtrado
na cápsula de Bowman é praticamente a
mesma que no plasma.
Á medida que o filtrado passa da cápsula de Bowman para os tubos contornados proximais,
sofre modificações por reabsorção de água e solutos específicos de volta à circulação, e
também por secreção de outras substâncias para os túbulos por parte dos capilares
peritubulares.
Em geral, a reabsorção tubular é mais importante que as secreções tubulares, na formação
de urina. Mas, a secreção desempenha um papel importante na determinação das
quantidades de iões potássio, hidrogénio e outras substâncias que são excretadas na urina.
Grande parte das substâncias que precisam de ser libertadas no sangue (especialmente
produtos metabólicos como ureia, ácido úrico ente outras) são pouco reabsorvidas, sendo
excretadas em elevadas quantidades na urina.
Outras substâncias estranhas como drogas, também são reabsorvidas no sistema tubular mas,
além disso, são segregadas do sangue para os túbulos, pelo que as suas taxas de excreção
também são elevadas.
Cada um destes processos – filtração glomerular, reabsorção tubular e secreção tubular – é
regulado de acordo com as necessidades do corpo.
Nota: cerca de 20% do plasma que passa pela arteríola aferente é filtrado para a cápsula de
Bowman e deste filtrado cerca de 99% é reabsorvido para a circulação.
Excreção urinária = (Filtração – reabsorção) + secreção
Página 72
3. Micção
Processo de esvaziamento da bexiga:
i.
Bexiga enche-se progressivamente de urina e a pressão no seu interior vai
aumentando
ii.
Sensores de pressão que existem nas suas paredes enviam sinais sensoriais
pelos nervos pélvicos até à porção sacral da medula, iniciando assim o
reflexo de micção, e um estímulo de contracção da bexiga é enviado por
fibras parassimpáticas
iii.
Estes reflexos dão-se constantemente mas só quando a pressão da bexiga
já é demasiado alta e os reflexos muito frequentes e fortes é que é
desencadeada o relaxamento voluntário do esfíncter externo e a bexiga é
finalmente esvaziada
4. Filtração glomerular
Barreiras de filtração glomerular: A membrana de filtração glomerular tem 3
camadas diferentes que funcionam como uma barreira para vários tipos de
substâncias/moléculas:
o Endotélio fenestrado (revestimento interno da membrana) – funciona
como uma barreira mecânica retendo substâncias de acordo com o seu
tamanho. Retém os elementos figurados do sangue e proteínas.
o Membrana basal – formada por glicoproteínas de carga negativa. Deixa
passar livremente líquido mas retém substâncias de acordo com a sua
dimensão e sobretudo de acordo com a sua carga eléctrica – cargas
negativas não retidas. Principal barreira à filtração à de macromoléculas
(principalmente proteínas).
o Podócitos – última camada e para além de ajudar na filtração de proteínas
forma uma “malha” em torno do glomérulo e mantém a sua integridade.
Débito Renal (DR) – quantidade de sangue que passa pelo rim;
Taxa de filtração glomerular (TFG) – quantidade de sangue filtrado a nível dos
glomérulos capilares na cápsula de Bowman;
A TFG tem alguns determinantes:
o Diferença entre as pressões hidroestáticas intracapilar (55 mm Hg,
favorável à filtração) e no espaço de Bowman (- 15 mm Hg, desfavorável –
espaço de bowman é o interior da cápsula). Representa-se ΔP e o seu
balanço é favorável à filtração (ΔP= 55-15 =40 mm Hg)
o Diferença entre a pressão oncótica (determinada pela quantidade de
proteínas do fluido) intracapilar (30 mm Hg, desfavorável) e no espaço de
Bowman (≈ 0 mm Hg, favorável). O balanço destas pressões é desfavorável
e representa-se por Δπ
o Área e permeabilidade da superfície de filtração, o que se dá o nome de
coeficiente de filtração glomeral e representa-se por Ks.
A TFG pode ser calculada matematicamente em mL/ min, através da expressão:
𝐾𝑠 ∗ (𝛥𝑃 − 𝛥𝜋) = 𝐾𝑠 ∗ (40 − 30) = 10 ∗ 𝐾𝑠
Alterações na permeabilidade das membranas ou nas várias pressões
mencionadas podem alterar a TFG a favor ou contra a filtração.
Página 73
a) Mecanismos de regulação da TFG e DR externos
A activação do sistema nervoso simpático estimula a diminuição do DR e da TFG
Só acontece em casos mais severos de diminuição da pressão arterial (PA)
Esta estimulação é conseguida através da vasoconstrição da arteríola
aferente (aumenta resistência arteriolar), reduzindo o fluxo sanguíneo
(impede a diminuição de volémia e diminui a pressão hidroestática – poupa
água) e aumenta a pressão.
Regulação hormonal
Angiotensina II
Vasoconstritor arteriolar
Previne redução do TFG em PA baixa
Aumenta a absorção de Na+ e H2O
Prostaglandina
Vasodilatador
Previne a redução acentuada de TFG
b) Mecanismos de autoregulação do DR e TFG
A PA influencia pouco a TFG devido aos
mecanismos de autoregulação;
O aparelho justaglomerular detecta variações na composição do fluido filtrado
(este controlo denomina-se de retrocontrolo tubulo-glomerular);
Complexo formado por 3 tipos de células: extraglomerulares,
justaglomerulares e mácula densa
Célula da Mácula densa – células especializadas da junção da Ansa de Henle
com o tubo contornado distal e contactam com as arteríolas aferente e
eferente. Funcionam como um sensor para o feedback/retrocontrolo
tubuloglomerular que permite o ajuste da TFG:
o Quando a TFG aumenta a reabsorção de Na+ e Cl- não é tão eficaz.
Consequentemente as células da mácula densa vão detectar um
aumento da [NaCl] no filtrado e vão induzir a contracção da arteríola
aferente, provocando uma diminuição da TFG (mecanismo de feedback
negativo).
o Se TFG decresce, provoca uma maior reabsorção de Na+ e Cl - na porção
ascendente da ansa de Henle => redução de [NaCl] na mácula densa
causa a dilatação das arteríolas aferentes (através da prostaglandina) e
aumento da produção de renina (ciclo na página seguinte).
Autoregulação miogénica
Capacidade dos vasos sanguíneos de resistirem à distensão durante
períodos de PA aumentada;
Dilatação aumenta a passagem de Ca2+ do fluido extracelular para a célula,
causando uma contracção do músculo liso
Impede o aumento do TFG nestas condições
Página 74
Pressão Arterial
inibe
inibe
ΔP glomerular
TFG
Reabsorção
de NaCl proximal
[NaCl] na mácula densa
Renina
Resistência
arteriolar eferente
Angiotensina I
Angiotensina II
Resistência arteriolar eferente
5. Reabsorção Tubular (Mecanismos de transporte tubular)
Transporte máximo: taxa máxima à qual um soluto pode ser transportado.
Como o transporte da maioria das substâncias para os túbulos está associado a
sistemas específicos (passivos ou activos), a determinada altura pode ocorrer
saturação das proteínas/ sistemas transportadores. Neste momento atingiu-se o
transporte máximo
Movimento do material filtrado do lúmen para o plasma pelos capilares
peritubulares
a) Vias de reabsorção: 3
Transporte mediado (activo)
o O Na+ é reabsorvido activamente através dos canais de sódio devido à
sua elevada concentração no lúmen.
o Responsável pelo elevado consumo energético do rim e permite
estabelecer gradientes de concentração de Na+ necesserários ao
transporte de outras substâncias.
o Existem também bombas que fazem o transporte para K+, H+, Ca2+
Difusão
o Devido à absorção de Na+ (activamente), cria-se um gradiente de
potencial, pelo que o Cl- se difunde passivamente para a corrente, e
reguladas por hormonas
o Água é sempre reabsorvida por osmose (transporte de água é
assegurado por canais próprios – aquapurinas – e está sempre
dependente do gradiente de diversos iões, como o Na+ fornece a
Página 75
o
o
o
o
força necessária para o movimento da água por osmose a partir do
túbulo renal).
60%-70% da reabsorção de água dá-se no tubo contornado proximal
(TCP) e depende do gradiente de Na+. No TC distal a permeabilidade à
agua é regulada pela hormona ADH.
A difusão de H2O no TCP cria um excesso de concentração de Cl- no
lúmen tubular, favorecendo a sua difusão a favor do gradiente
eléctrico. A ureia é reabsorvida pela mesma razão.
Difusão simples: CO2 e ureia
Difusão facilitada - através de canais iónicos: Na+, Cl-, K+
Transporte paracelular
o Os produtos reabsorvidos atravessam espaços intercelulares
o É a via utilizada pelas proteínas, Ca2+, Cl-, H2O, etc, em alguns
segmentos do nefrónio
I – transporte activo;
II – difusão;
III – transporte paracelular
Página 76
b) Reabsorção no tubo contornado proximal
Na 1ª metada dá-se a reabsorção de Na+ por co-transporte de glucose,
aminoácidos.
o Na membrana apical existem transportadores específicos que utilizam
a energia do transporte de Na+ a favor do seu gradiente (para o interior
da célula) para transportar uma molécula de glucose contra o seu
gradiente electroquímico (também para dentro da célula). Uma vez no
meio intracelular a glicose é difundida passivamente para o fluido
intersticial.
o O transporte de glucose está assim dependente da actividade da
bomba Na+/K+ ATPase que é responsável por eliminar sódio do meio
intracelular, em troca de potássio
Na 2ª metade dá-se a reabsorção de Cl-.
c) Reabsorção na Ansa de Henle
A ansa de Henle divide-se em 3 partes funcionais que desempenham
funções diferentes:
i. Porção fina descendente
Bastante permeável a H2O e moderadamente permeável a alguns
solutos
Permite a difusão simples de substâncias através das suas paredes
Página 77
ii. Porção grossa ascendente
Bastante impermeável a água
Possui células epiteliais com elevada actividade metabólica
Estas células permitem a absorção activa de Na+, Cl- e K+, e de
outros solutos como Ca2+, HCO3-, Mg2+
Secreta-se H+
HCO3- é totalmente filtrado no glomérulo. No lúmen dos túbulos
renais HCO3- combina-se com H+ que foi excretado e forma H2CO3.
Este ácido dissocia-se em H2O e CO2 por acção da enzima anidrase
carbónica e o CO2 é difundido para o interior da célula. Aí volta a
associar-se a H2O e forma outra vez H2CO3 que, por sua vez forma
H+ + HCO3-. Os iões bicarbonato podem então retornar à corrente
sanguínea.
iii. Porção fina ascendente
Bastante impermeável a água
Não se dá reabsorção significativa
d) Reabsorção no tubo contornado distal e ducto colector
Ambos impermeáveis à ureia
Permitem a reabsorção de Na+ (e outros iões) activamente
Secretam H+ (têm papel fundamental no equilíbrio ácido-base)
Sujeitos a acção de hormonas, nomeadamente à aldosterona e ADH
o Aldosterona estimula a absorção distal de Na+ e eliminação de K+ e H+
o ADH controla a permeabilidade destes segmentos à água, controlando
assim o grau de diluição da urina:
Elevadas quantidades de ADH TCD torna-se permeável a água,
permitindo a sua reabsorção urina muito concentrada
Ausência de ADH permeabilidade dos túbulos é baixa, e a urina
torna-se mais diluída
6. Mecanismo de Contra-Corrente
A multiplicação em contracorrente é o mecanismo pelo qual o conteúdo de dois tubos
adjacentes se desloca em direcções opostas originando concentração de solutos
progressivamente maiores. O gradiente de concentração entre os dois ramos da ansa é
estabelecido com gasto de energia e tem por base 3 propriedades:
a. Transporte activo de sódio e co-transporte de potássio, cloreto e outros iões ao longo
dos ramos ascendentes (porção grossa);
b. Baixa permeabilidade à água dos ramos ascendentes;
c. Alta permeabilidade à água dos ramos descendentes.
Página 78
Para que a maior parte da água filtrada seja reabsorvida e se excrete uma urina concentrada é
fundamental manter uma pressão osmótica (concentração de solutos) elevada na medula
renal. Para tal facto contribui o mecanismo de contra-corrente da ansa de Henle.
Paralela à ansa existe um conjunto de vasos (vasa recta) que se dispõe em contracorrente em relação ao fluxo do túbulo renal e que ajuda a manter a
hiperosmolaridade da medula e o gradiente da ansa:
o No ramo ascendente da vasa recta vai haver entrada de água e saída de sais,
o que torna a medula hiperosmótica e contribui para a reabsorção de água no
ramo descendente da ansa de Henle.
o No ramo ascendente da ansa circula um fluido hiposmótico devido à
reabsorção de sais e à impermeabilidade à água. No ramo correspondente da
vasa recta (ramo descendente) vai verificar-se saída de água e a entrada de
sais, ficando o sangue progressivamente mais concentrado.
7. Clearance (depuração) de creatina
Forma de avaliar a função renal.
Creatina - substância excretada para os tubulos renais que não volta a ser reabsorvida. Se os
rins funcionam mal, [creatina] aumenta no sangue.
Clearance – quantidade de sangue que contem determinada substância, por Δt. Se tiver uma
substância que não seja reabsorvida nem excretada pelos túbulos renais, consegue-se saber a
clearance que foi excretada e então saber que essa é a quantidade de substância que passou
do sangue para as cápsulas de Bowman.
𝑐𝑙𝑒𝑎𝑟𝑎𝑛𝑐𝑒 ∗ [ ] 𝑃𝑙𝑎𝑠𝑚á𝑡𝑖𝑐𝑎 = [ ] 𝑈𝑟𝑖𝑛á𝑟𝑖𝑎 ∗ 𝑉𝑜𝑙𝑢𝑚𝑒 𝑈𝑟𝑖𝑛á𝑟𝑖𝑜
Página 79
Sistema Digestivo
Funções Principais
Fornecer continuamente água, electrólitos e nutrientes.
São necessários
Movimento do conteúdo ao longo do tubo digestivo;
Secreção de sucos digestivos e digestão dos alimentos;
Absorção de água, electrólitos e produtos da digestão;
Circulação de sangue para absorção e controlo;
Controlo por sistemas locais, nervosos e hormonais.
Anatomia do sistema digestivo
Princípios gerais da mobilidade gastrointestinal
Características Gerais da parede intestinal
As funções motoras do tubo digestivo realizam por diversas camadas de músculo liso. A
parede intestinal é composta por, de fora para dentro:
Página 80
1.
2.
3.
4.
5.
Serosa
Camada muscular longitudinal
Camada muscular circular
Submucosa
Mucosa
A zona profunda da mucosa contém fibras dispersas de
músculo liso, a muscularis mucosae.
O músculo liso gastrointestinal funciona com um síncicio. As fibras lisas dos músculos
longitudinais e circulares estão conectadas electricamente por vários gap junctions que
permitem a passagem de iões de uma célula para as outras. Cada camada muscular funciona
como um síncicio: e quando em algum ponto da massa muscular aparece um potencial de
acção, propaga-se em todas as direcções do músculo. A distância percorrida depende da
excitabilidade do músculo. Existem ainda algumas conexões entre as duas camadas, o que
pode provocar a excitação de uma a partir da outra.
Actividade eléctrica do músculo liso gastrointestinal
Este músculo liso é excitado de forma quase contínua. Esta
actividade é feita de duas formas: ondas lentas e ondas rápidas. As
ondas lentas ocorrem ritmicamente, e não são potenciais de acção.
São geradas por interacções complexas entre as células do músculo
liso e as células de Cajal, que se pensa serem “pacemakers” para as
células do músculo liso. As ondas rápidas, ou spikes, são potenciais
de acção, e acontecem quando o potencial da célula sobe acima de
-40 mV. Quanto mais alto for o potencial da célula, maior a
frequência destas ondas.
Em repouso, o potencial da membrana é normalmente -56 mV.
Quando o potencial sobe acima deste valor (despolarização) as
fibras ficam mais excitáveis. Quando o potencial desce abaixo deste
valor (hiperpolarização) as fibras ficam menos excitáveis.
Factores de despolarização: distensão do músculo, estimulação com acetilcolina,
estimulação de nervos parassimpáticos que segregam acetilcolina nas suas
terminações, estimulação por hormonas gastrointestinais
Factores de hiperpolarização: efeito da noradrenalina ou adrenalina sobre a
membrana muscular, estímulo de nervos simpáticos que segregam acetilcolina nas
suas terminações
Página 81
Controlo Neuronal da função gastrointestinal: Sistema Nervoso Entérico
O sistema nervoso está totalmente alojado nas paredes do
tracto alimentar, começando no esófago e terminando no ânus.
Alberga aproximadamente 100 milhões de neurónios. Este
sistema controla especialmente os movimentos e secreções
gastrointestinais.
O SNE é constituído por 2 plexos:
o plexo exterior (ou plexo mesentério / plexo de
Auerbach), situado entre as duas camadas musculares.
A sua estimulação produz: 1) aumento do tónus da
parede intestinal; 2) maior intensidade das contracções
rítmicas; 3)aumento da frequências das contracções; 4)
maior velocidade de condução.
o plexo interior (ou plexo submucosal / plexo de Meissner), situado na submucosa.
Controla as secreções e o fluxo sanguíneo.
Controlo autónomo do sistema gastrointestinal
Os nervos parassimpáticos aumentam a actividade do sistema nervoso entérico. Por sua vez,
este intensifica a actividade da maioria das funções gastrointestinal. A inervação
parassimpática do tubo digestivo é formada pelas divisões craneal e sacra.
O sistema nervoso parassimpático craneal inerva, através dos nervos vago, o esófago,
o pâncreas e a primeira parte do intestino grosso.
O sistema nervoso parassimpático sacro inerva, através dos nervos pélvicos, a parte
distal do intestino grosso. O cólon sigmóide, o recto e o ânus estão especialmente
inervados por fibras parassimpáticas, que intervêm nos reflexos da defecação.
Reflexos gastrointestinais
Existem 3 tipos de reflexos essenciais:
Reflexos que estão totalmente integrados na parede do tracto intestinal, no SNE
controlam a secreção gastrointestinal, o peristaltismo, as contracções de mistura, os
efeitos locais inibidores, etc.
Reflexos desde o tracto alimentar até aos gânglios simpáticos e de novo até ao tracto
alimentar transmitem sinais a longa distância, como os do estômago que provocam a
evacuação do cólon (reflexo gastrocólico), do cólon e intestino delgado que inibem a
mobilidade e secreções gástricas (reflexo enterogástrico) e reflexos originados no
cólon que inibem o esvaziamento do conteúdo ileal e no cego (reflexo cólico-ileal)
Reflexos desde o tracto alimentar até à medula espinhal ou tronco cerebral e para trás
de novo consistem especialmente em: 1) reflexos originados no estômago e duodeno
-> tronco cerebral -> estômago através do nervo vago, que controlam a actividade
motora e secretora; 2) reflexos dolorosos que provocam uma inibição geral da
totalidade do aparelho digestivo; e 3) reflexos de defecação que viajam do cólon e
recto -> medula espinhal e regressão para produzir fortes contracções do cólon, recto
e abdómen.
Página 82
Hormonas gastrointestinais
Secretina
Gastrina
Colecistoquinina
Péptido inibidor gástrico (pepsinogénio?)
Tipos Funcionais de movimentos no tubo digestivo
Dividem-se em dois tipos:
Propulsivos (Peristalse). Os movimentos progressivos ou peristálticos, são responsáveis
pela progressão dos alimentos ao longo do tracto gastrointestinal a uma velocidade
que permita a sua digestão e absorção. Dão-se principalmente no esófago e intestino e
caracterizam-se pelo aparecimento de anéis contrácteis ao longo do músculo.
Contracções envolventes. São responsáveis por manter o alimento e os sucos
digestivos bem misturados e homogeneizados. Por vezes, os próprios movimentos
peristálticos desempenham esta função.
Ao longo do tubo digestivo as características específicas de cada tipo de movimento vão
variando de acordo com a função e necessidade de cada órgão / região.
Trânsito e mistura dos alimentos no tubo digestivo
Ingestão dos alimentos
Depois de ingerida, a comida segue os dois processos seguintes
até chegar ao estômago:
1. Mastigação
A maior parte dos músculos envolvidos neste processo são
inervados pelo ramo motor do 5º nervo cranial. A maior parte da
mastigação deve-se a reflexo mastigativo.
2. Deglutição
Este processo pode ser dividido em duas fases: fase voluntária e
fase involuntária. Na primeira, a comida é voluntariamente
enviada para a faringe por pressão da língua contra o palato. A
segunda, que é involuntária, ocorre do seguinte modo: a traqueia
é encerrada momentaneamente e o esófago aberto, e uma onda
rápida peristáltica iniciada pelo sistema nervoso na faringe força o bolo alimentar para o
esófago, sendo que este processo leva menos de 2 segundos a ocorrer. No esófago, a comida é
conduzida até ao estômago por dois tipos de movimentos peristálticos: primários e
secundários, sendo que os primários são continuações das ondas peristálticas que começam
na faringe, e os secundários resultam da distensão do esófago causada por comida retida. Ou
seja, os movimentos secundários iniciam-se quando os primários não são suficientes.
Página 83
Funções motoras do estômago
O estômago tem 3 funções essenciais: armazenamento de
grandes quantidades de comida até que possa ser processada,
envolvimento da comida com os sucos gástricos até à formação
do quimo, e saída do quimo para o duodeno a um ritmo que
permita a sua absorção.
1. Função de armazenamento
À medida que a comida entra no estômago, vai-se depositando de
modo que a comida mais recente se encontre mais próxima do
esófago e a mais antiga se encontre mais próxima da parede
oposta do estômago. Quando a comida começa a ser demasiada
no estômago, o reflexo vagovagal que vai do estômago até ao
tronco cerebral reduz o tónus muscular no estômago, o que causa o seu relaxamento, para
que caiba mais comida.
2. Envolvimento da comida
A retropulsão é um mecanismo importante de mistura. Cada vez que uma onda peristáltica
passa pelo antro até ao piloro, o músculo pilórico se contrai, dificultando ainda mais o
esvaziamento através do piloro. A maior parte do conteúdo do antro é comprimido e
retrocede através do anel peristáltico até ao corpo do estômago. Os sucos digestivos são
secretados pelas glândulas gástricas. Estas secreções entram imediatamente em contacto com
as porções de comida mais próximas da parede do estômago. No estômago, as ondas
constrictoras peristálticas fracas (ou ondas envolventes) ajudam no envolvimento da comida
com os sucos. No final do envolvimento com os sucos, está formado o quimo.
3. Saída do quimo
O esvaziamento do estômago é promovido por intensas contracções peristálticas na base do
estômago. Ao mesmo tempo, este esvaziamento é contrariado por vários graus de resistência
à passagem do quimo pelo piloro.
Regulação do esvaziamento do estômago
Durante o esvaziamento do estômago dão-se intensas contracções do antro do estômago e a
passagem de alimentos para o duodeno é controlada pelo nível de relaxamento / contracção
do esfíncter pilórico. Este, por sua vez, é controlado por reflexos hormonais e nervosos
provenientes do estômago e principalmente do duodeno.
O grau de enchimento do estômago e a secreção de gastrina estimulam os movimentos
peristálticos e promovem o esvaziamento.
Por outro lado, reflexos enterogástricos e hormonas segregadas pelo duodeno, como a
colecistoquinina e a secretina, inibem o processo de esvaziamento quando já existe quimo
suficiente no intestino ou quando este é excessivamente ácido, é hiper / hipotónico ou quando
contém demasiadas gorduras ou proteínas mal processadas.
Página 84
Movimentos do intestino delgado
Os movimentos do intestino delgado têm como função a
fragmentação e digestão do bolo alimentar e a sua progressão ao
longo do intestino. Estes movimentos devem ser lentos, para
permitir a correcta digestão e absorção, e podem dividir-se em 4
tipos:
As contracções segmentares são responsáveis por fragmentar o bolo
alimentar e misturá-lo com as enzimas digestivas. Provocam, ao
longo do intestino, zonas de contracção intercaladas com zonas de dilatação e podem ser
isoladas, regularmente espaçadas, irregularmente espaçadas ou fracas regularmente
espaçadas.
Os movimentos peristálticos são de curta duração e têm pouco poder de progressão, sendo
necessárias 3 a 5 horas para que o quimo passe desde o piloro até à válvula ileocecal. Esta
actividade é grandemente induzida após uma refeição. Isto é causado parcialmente pela
entrada do quimo no duodeno, mas também devido ao reflexo gastrointestinal que é iniciado
pela distensão do estômago e conduzido pelo plexo mesentérico até ao intestino. No entanto,
estes movimentos podem também ser controlados por: gastrina, CCK, insulina, motilina e
serotina.
As contracções da mucosa muscular mantém o intestino pragueado e têm como principal
função aumentar a superfície de absorção.
Por último, a contracção das vilosidades intestinais ajuda a misturar o bolo alimentar com as
secrecções e a escoar o conteúdo dos vasos linfáticos.
Função da válvula ileocecal
A principal função desta válvula é impedir o refluxo dos componentes fecais do cólon para o
intestino delgado. Esta válvula encerra quando existe demasiada pressão sobre ela e tenta
empurrar os componentes fecais para o interior do cólon.
Movimentos do cólon
No cólon dá-se essencialmente a absorção de água e
electrólitos e o armazenamento das fezes até que estas sejam
expulsas. Os movimentos que ocorrem neste órgão são de 3
tipos:
Os movimentos de mistura, ou hausteração, provocam
contracções ao longo do cólon que são responsáveis por
remexer as fezes e mantê-las em contacto com a mucosa para
que a água e outras substâncias possam ir sendo absorvida. Em
conjunto com as ondas de contracção dos movimentos
peristálticos possibilitam a progressão da massa fecal.
Os movimentos em massa são movimentos peristálticos modificados que levam ao
desaparecimento das haustras (segmentação) e ao movimentos das fezes até ao recto para
posterior defecação.
Página 85
Quando as fezes pressionam o interior do recto, existe desde logo o desejo de defecar. Isto
inclui o reflexo de contracção do recto e o relaxamento dos esfíncteres anais. A saída das fezes
é impedida de duas formas: um esfíncter anal interno, situado no interior do ânus, e um
esfíncter anal externo, composto de músculo estriado, e cuja acção é voluntária.
Funções secretoras do tracto alimentar
Funções das glândulas secretoras:
A maioria das regiões produzem enzimas digestivas
As glândulas mucosas segregam muco para lubrificar e proteger todas as regiões do
tubo digestivo
Princípios gerais da secreção no tubo digestivo
O contacto dos alimentos com o epitélio estimula a secrecção. Esta estimulação activa o SNE.
Estes estímulos podem ser: 1) tactéis; 2) irritação química; 3) distensão da parede intestinal.
A estimulação parassimpática aumenta os índices de secreção glandular. Em particular, os
nervos glossofaríngico e vago estimulam as glândulas do tracto superior. A estimulação
simpática pode ter um duplo efeito sobre a secreção glandular, podendo aumentá-la ou
diminuí-la. Diversas hormonas, como a gastrina, a colecistoquinina ou a secretina, são
secretadas no tracto GI e lançadas na corrente sanguínea actuando depois como
estimuladores ou inibidores em diversos órgãos.
Existem 3 tipos fundamentais de secreção:
Na secreção endócrina as células / glândulas produzem hormonas que são lançadas
para a corrente sanguínea e postas em circulação podendo actuar em tecidos muito
distantes.
Na secreção parácrina as células segregam mensageiros químicos que actuam em
células adjacentes, não entrando em circulação.
Na secreção autócrina as substâncias secretadas para o meio extracelular actuam na
própria célula secretora.
Existem essencialmente 4 tipos de glândulas:
1. Glândulas mucosas unicelulares chamadas células mucosas, localizadas na superfície
do epitélio na maior parte do tracto alimentar
2. Invaginações do epitélio para a submucosa, localizadas em vários locais do tracto
alimentar
3. Glândulas tubulares, típicas do estômago e duodeno superior
4. Glândulas salivares, pâncreas e fígado, as chamadas glândulas complexas, que estão
associadas ao tracto alimentar
Secreção de substâncias orgânicas
A secreção processa-se da seguinte forma:
1. O nutriente necessário para a formação da secreção tem de chegar à base da célula
glandular;
2. As mitocôndrias usam a energia do nutriente para formar ATP;
Página 86
3. O ATP é usado para sintetizar as substâncias secretoras
orgânicas;
4. Estas substâncias são transportadas para o retículo
endoplasmático, e depois para o complexo de Golgi;
5. Aqui, estes materiais são transformados, e reenviados
sob a forma de vesículas secretórias;
6. Estas vesículas permanecem inactivas até chegar o sinal
nervoso que causa a libertação do seu conteúdo através
da superfície da célula;
Secreção de água e electrólitos;
A secreção processa-se da seguinte forma:
1. Estimulação nervosa causa o transporte de iões cloreto para o interior da célula;
2. O aumento de electronegatividade causa a entrada de iões positivos, como o sódio
3. O excesso de cargas negativas e positivas causa, por osmose, a entrada de água na
célula;
4. A pressão na célula aumenta, e logo a seguir começa a diminuir, através da saída
tanto de água como de electrólitos para o exterior da célula;
Propriedades protectoras e lubrificantes do muco
O muco é uma mistura de água, electrólitos e glicoproteínas, que por sua vez são constituídas
por polissacarídeos e pequenas quantidades de proteínas. Este muco tem a capacidade de
permitir um movimento mais fácil da comida ao longo do tubo digestivo e evita que esta cause
danos nas paredes do tracto alimentar.
Secreção da saliva
A saliva é secretada pelas glândulas parótidas, submandibulares
e sublinguais. Na sua constituição entra: a ptialina (ou amilose
salivar), que digere polímeros de glícidos, como o amido; muco
que lubrifica o bolo alimentar e protege a mucosa da boca;
grandes quantidades de iões potássio e bicarbonato; e iões sódio
e cloro em concentrações bastante inferiores às do plasma
sanguíneo.
A produção de saliva é controlada pelo SN parassimpático e, em
menor grau, pelo SN simpático. Estímulos olfactivos e tácteis
provenientes da língua, boca e faringe estimulam o núcleo
salivatório do tronco cerebral e promovem o aumento da
secreção salivar.
Secreção esofágica
As secreções no esófago são apenas mucosas e servem para lubrificar os elementos que
intervém na deglutição. Ao longo do esófago existem portanto glândulas mucosas simples.
Página 87
Mais abaixo, perto do estômago, existem também glândulas mucosas compostas, cujas
secreções previnem escoriações no esófago por parte da comida.
Secreção gástrica
Características da secreção gástrica
A mucosa estomacal tem dois tipos principais de glândulas:
Gástricas: são constituídas por 3 tipos de células: mucosas, pépticas e parietais. As
primeiras segregam muco, as segundas pepsinogénio e as terceiras ácido clorídrico. O
pepsinogénio necessita de activação para formar a pepsina, activação essa que é feita
em contacto com o ácido clorídrico.
Pilóricas: são estruturalmente semelhantes às anteriores, mas contém poucas células
pépticas e quase nenhumas parietais. Assim, a sua função é praticamente apenas a
segregação de muco para protecção da mucosa pilórica.
Células mucosas superficiais
A superfície do estômago está coberta células mucosas superficiais, que segregam grandes
quantidades de muco, que cobre a mucosa do estômago, e o protege contra a acidez do
ambiente dentro deste, já que este muco é alcalino.
Estimulação da secreção do ácido gástrico
A segregação deste ácido é sobretudo controlada por sinais endócrinos e nervosos. Para além
disso, as células parietais operam em conjunto com as células ECL, que segregam histamina.
A produção de HCL dá-se nas células parietais do estômago, onde ocorre o transporte activo
de iões Cl- do citoplasma para o lúmen e de iões Na+ no sentido oposto. Consequentemente,
cria-se um potencial negativo no lúmen que promove a difusão de K+ e Na+.
Simultaneamente no interior da célula a
água dissocia-se em OH- e H+. H+ é
transportada para o lúmen em troca de K+
(via H+ / K+ ATPase) e reage com o cloro
dando origem a HCL. Os iões OH- combinamse com CO2, que entra na célula e são
difundidas para o fluido extracelular sob a
forma de HCO3-. O HCl produzido é então
secretado do lúmen das células parietais
para o estômago.
Função e regulação da gastrina
A gastrina é uma hormona produzida pelas células G do antro do estômago e duodeno. A sua
principal função é estimular a secreção de ácidos pela mucosa gástrica mas também actua no
Página 88
sentido de promover a motilidade do estômago e do intestino delgado, os movimentos de
massa do cólon e o crescimento do estômago, intestino e pâncreas exócrino.
A sua produção é estimulada pela presença de a.a. e proteínas no estômago, pela actividade
do vago e por factores circulatórios. Por outro lado a presença de ácido inibe a sua secrecção.
Regulação da secreção de pepsinogénio
Esta regulação ocorre em resposta a dois sinais: estimulação das células pépticas por
acetilcolina libertada pelos nervos vagos plexos nervosos entéricos gástricos, ou em resposta à
presença de ácido no estômago. Assim, a presença de pepsina no estômago depende da
quantidade de acido lá existente.
Fases da secreção gástrica
A secreção gástrica divide-se em 3 fases:
1. Fase cefálica: ocorre antes da comida entrar no
estômago, e esta secreção é resultado de sinais
neurológicos que chegam ao estômago
principalmente enquanto a comida está na boca
ou no esófago estimulando o nervo vago e levam
à produção de 20% da secreção total gástrica.
2. Fase gástrica: ocorre assim que a comida entra
no estômago, por reflexos vagovagais, reflexos
entéricos locais, e por mecanismos gástricos.
Todos causam a secreção de suco gástrico e
representam 70% da secreção total gástrica.
3. Fase intestinal: ocorre quando o quimo entra no
duodeno no qual é estimulada a secreção de
sucosgástricos, de secretina e de colecistoquinina.
Secreção pancreática
O suco pancreático, produzido no centro acinar pancreático, juntamente com grandes
quantidades de iões bicarbonato, é conduzido através do ducto pancreático que depois
desagua no ducto hepático antes de despejar o seu conteúdo no duodeno. Este suco é
segregado em resposta à presença de quimo na porção superior do intestino delgado.
Enzimas digestivas pancreáticas
As mais importantes são a tripsina, a quimotripsina, e a carboxipolipeptidase. A mais
abundante é de longe a tripsina. A enzima que degrada os carbohidratos produzida no
pâncreas é a amilase pancreática, e as que degradam os lípidos são a lipase pancreática, a
colesterol esterase e a fosfolipase. Quando são sintetizadas, estas encontram-se inactivas, e
são activadas assim que chegam ao tracto intestinal por enzimas específicas.
Página 89
Regulação da secreção panceática
A produção de enzimas é estimulada principalmente estimulada
pela presença de péptidos no intestino; gorduras que levam à
secreção de quantidades semelhantes de água, HO3- e enzimas;
o aumento da acidez provoca principalmente produção de HCO3. Ácidos gordos e aminoácidos levam À libertação de
colicistoquinina (CCK) (estimula a secreção enzimática) e HCl leva
a um aumento da secretina (estimula a secreção de HCO3-)
A secreção pancreática dá-se em 3 fases: cefálica, gástrica e
intestinal. Nas duas primeiras há secreção maioritariamente de
enzimas, estimulada hormonalmente e pelo nervo vago. No
entanto, Sá na fase intestinal, com a secreção de grandes
quantidades de HCO3-, agua e electrólitos em resposta à chegada
do quimo é que grande parte das enzimas flui para o intestino.
Função da secretina
A secretina é uma hormona produzida no duodeno e no jejuno aquando da presença de ácido
gástrico nestas regiões. Tem um efeito inibidor na secreção de ácidos e na motilidade do
estômago e estimula a produção de bicarbonato pelo pâncreas e fígado, o que ajuda a
neutralizar o ácido. Promove também a secreção de enzimas, pois potencia a acção de CCK, e o
crescimento da porção exócrina deste órgão.
Função da colecistoquinina
A CCK é uma hormona produzida no duodeno e jejuno aquando da presença de péptidos ou
ácidos gordos nestas regiões.
A CCK inibe a secreção de ácido e o esvaziamento gástrico. Por outro lado estimula a secreção
de bicarbonato pelo fígado e pâncreas; promove a secreção de enzimas pelo pâncreas e o
crescimento da sua porção exócrina; e estimula a contracção da vesícula biliar e o relaxamento
do esfíncter de Oddi, permitindo a libertação de bílis e sucos pancreáticos.
Secreção e regulação de bicarbonato
Iões bicarbonato são secretados para o duodeno a partir de células do
epitélio glandular. CO2 é difundido a partir do sangue para o interior das
células combina-se com a água, numa reacção catalisada pela anidrase
carbónica, para dar origem a H2CO3 que logo se dissocia em HCO3- e H+.
Os HCO3- são transportados activamente para o lúmen dos tubos
pancreáticos em conjunto com o Na+ «. A concentração destes iões no
meio intracelular é reposta pela troca activa de H+ (sai) e Na+ (entra)
entre a célula e o sangue. O movimento de Na+ e HCO3- cria um
gradiente osmótico no lúmen que leva à passagem de água por osmose
para os ductos e à criação de uma solução osmótica.
Cerca de 70% da secreção pancreática de bicarbonato ocorre durante a fase intestinal da
digestão e é regulada hormonalmente. O aumento da concentração de ácido vindo do
estômago estimula a produção de secretina pelo intestino delgado. Esta hormona entra em
Página 90
circulação e actua no pâncreas, onde pode ser potenciada pela presença de CCK promovendo
a secreção de bicarbonato. O aumento do fluxo deste ião para o intestino neutraliza o HCl, o
que inibe a secreção de secretina e, consequentemente, de HCO3-.
Secreção de bílis pelo fígado. Funções da
via biliar
O fígado desempenha funções muito importantes.
Pode-se destacar:
a sua função endócrina (activa e degrada
hormonas);
a formação, secreção e armazenamento da
bílis;
o seu papel na regulação do metabolismo de
nutrientes (glícidos, lípidos) e síntese
proteica;
As suas funções de coagulação (sintetiza
factores de coagulação e fibrinogénio);
A sua participação na desintoxicação do
sangue.
Os sais biliares (bílis) desempenham duas funções principais: emulsionam os lípidos ajudando à
sua digestão e participam na absorção de gorduras (ácidos gordos, colestrol, etc.) no intestino.
Mais de 90% desses sais são reabsorvidos. Por outro lado eleminam os produtos residuais do
sangue.
A circulação enterohepática tem início com a síntese de sais biliares pelo fígado. De seguida
estes são transportados através do ducto biliar até à vesícula biliar, onde ficam armazenados,
ou directamente para o intestino. Aí ajudam na digestão e absorção das gorduras e cerca de
5% dos sais são excretados nas feses. O restante é absorvido pelo epitélio intestinal e é levado
de volta ao fígado pela veia porta.
Regulação da entrada da bílis no duodeno
A presença de ácidos gordos no duodeno aumenta a secreção de CCK, que entra em
circulação. O aumento da sua concentração e a estimulação do SNE e SN parassimpático
(durante as fases gástrica e cefálica) provoca a contracção da vesícula biliar e o relaxamento do
esfíncter de Oddi provocando o aumento do fluxo de bílis para o duodeno.
Secreções do intestino delgado
No intestino, as chamadas glândulas de Brunner, segregam grandes quantidades de muco
alcalino (protecção contra pH ácido), como resposta a estímulos físicos, estimulação vagal, ou
secretina. Estas células são inibidas por estimulação simpática.
Secreção do suco intestinal
Página 91
O epitélio na mucosa intestinal contém células secretoras, que segregam fluido extracelular
quase puro e muco.
Regulação da secreção no intestino delgado
No intestino delgado, a secreção é regulada por reflexos nervosos entéricos locais,
especialmente por reflexos iniciados devido a estímulos físicos (irritação, por exemplo) por
parte do quimo no intestino.
Secreções no intestino grosso
A mucosa do intestino grosso tem como função principal segregar muco, que contém uma
quantidade considerável de iões bicarbonato. A taxa à qual este muco é segregado é
determinada principalmente por estímulos físicos. Este muco tem como função proteger o
intestino grosso contra agressões provocadas pelas fezes e contra bactérias.
Digestão e absorção no tracto gastrointestinal
A grande maioria dos alimentos que ingerimos são digeridos por acção de enzimas, num
processo denominado hidrólise, que requer o consumo de uma molécula de água.
Digestão e absorção dos hidratos de carbono
A digestão dos carbohidratos é maioritariamente químico, embora possa ser facilitada pela
digestão mecânica devido aos movimentos do tubo digestivo. A hidrólise de polissacarídeos
tem início na boca quando os alimentos se misturam com a amilase presente na saliva. Nesta
primeira fase dá origem a maltose e a pequenos polímeros de glucose. Quando o alimento
atinge o estômago, a acidez do meio bloqueia a acção da amilase salivar. A digestão dos
restantes glícidos dá-se já no intestinom para onde é secretada amilase pancreática. Os
monossacarídeos resultantes são absorvidos no intestino. A glucose e a galactose são
absorvidas por um mecanismo de co-transporte de sódio. A frutose á absorvida em parte por
difusão facilitada, e o restante é convertido em glicose e transportado como descrito acima.
Página 92
Digestão e absorção de proteínas
A digestão das proteínas é fundamentalmente química. No estômago a enzima pepsina inicia o
processo de digestão clivando algumas ligações peptídicas entre aminoácidos e dando origem
a proteases e polipéptidos. Já no intestino diversas enzimas presentes no suco pancreático dão
continuidade ao processo: tripsina e quimiotripsina quebram as proteínas em péptidos mais
pequenos; carboxipolipeptidase cliva aminoácidos dos terminais carboxilo das proteínas. Nas
microvilosidades são produzidas peptidases que actuam formando di/tripéptidos e alguns
aminoácidos livres que são finalmente absorvidos pela membrana luminar do epitélio
intestinal por um mecanismo semelhante ao transporte de glucose. Alguns aminoácidos são
também transportados por difusão facilitada.
Digestão e absorção de lípidos
A digestão de gorduras é fundamentalmente química. Na boca é produzida lipase o que
permite que alguns triglicéridos vão sendo degradados no estômago. No entanto, é no
intestino que se dá a maior parte da digestão. A bílis contém os sais biliares e lecticina (um
fosfolípido) que juntamente com a agitação mecânica proporcionada pelos movimentos do
tubo digestivo é fundamental para a emulsão de gorduras. A lipase secretada pelo pâncreas
actua então sobre as gorduras emulsionadas e degrada-as em ácidos gordos e gliceróis. Os sais
biliares ajudam também na digestão dos lípidos, através da formação de micelas de tamanho
reduzido, e que são solúveis em água.
Esta acção dos sais biliares facilita bastante a absorção dos lípidos digeridos. Depois, os ácidos
gordos e gliceróis saem da micela para o epitélio, pois também são solúveis na membrana do
epitélio, deixando a micela livre para actuar de novo. Deste modo, cerca de 97% dos lípidos
digeridos são absorvidos desta forma.
Página 93
Absorção de água e electrólitos
A absorção de água ao longo do tubo digestivo dá-se essencialmente por osmose, enquanto
que os electrólitos são absorvidos por outros processos. Quando o quimo está suficientemente
diluído os gradientes de concentração permitem a absorção de água por osmose na mucosa
intestinal. Um dos factores que contribui para a criação do gradiente osmótico que permite a
absorção de água é o transporte activo de iões do epitélio intestinal para os espaços
intercelulares. O sódio é absorvido por transporte activo, os iões cloreto por difusão, e os iões
bicarbonato combinam-se com hidrogénio formando ácido carbónico que depois se dissocia
em água e dióxido de carbono, sendo que este último é expulso pelos pulmões. Outros iões
são também absorvidos, como o cálcio, cuja absorção depende da quantidade necessária pelo
organismo; o ferro é também absorvido, também de acordo com as necessidades do
organismo. Para além destes, há também a absorção de potássio, magnésio, fosfato e outros.
Absorção no intestino grosso
A mucosa do intestino grosso tem uma elevada capacidade para absorver sódio e iões cloreto.
Esta mucosa segrega iões bicarbonato e absorve em igual número iões cloreto. Este
bicarbonato serve essencialmente para combater a acidez criada por acção bacteriana no
intestino grosso. O intestino grosso consegue absorver um máximo de 5 a 8 litros de água e
electrólitos por dia, sendo que o excesso aparecerá sob a forma de diarreia.
Página 94
Osso – Sistema Locomotor
Elevada dureza
Elevada resistência
Elevada vascularização
Elevada taxa metabólica
Grande actividade durante toda a vida
Grande capacidade de auto-regeneração
Constituição
Parte orgânica (30%) – flexibilidade
Parte inorgânica (70%) – rigidez
Tecido conjuntivo especializado
Células
Substância intersticial
Funções
Mecânicas
o Suporte para o corpo contra forças externas. Age como um sistema de
alavanca para transferir força
o Protecção para os órgãos internos
Metabólicas (na medula óssea formam-se células sanguíneas (hematopoiese) e
armazenamento de cálcio)
Estrutura microscópica
O estudo microscópico do tecido ósseo permite distinguir a substância óssea compacta e a
esponjosa. Embora os elementos constituintes sejam os mesmos nos dois tipos de substância
óssea, eles dispõem-se diferentemente conforme o tipo considerado e aí o seu aspecto
macroscópico também difere.
Substância óssea compacta:
as lâminas de tecido ósseo encontram-se fortemente unidas umas às outras pelas
suas faces, sem que haja espaço livre interposto.
é mais densa e rija
Substância óssea esponjosa:
as lâminas ósseas, mais irregulares em forma e tamanho, arranjam-se de forma a
deixar entre si espaços ou lacunas que se comunicam umas com as outras e que, à
semelhança do canal medular, contém medula.
Página 95
Nos ossos longos, a diáfise é composta por osso compacto externamente ao canal medular,
enquanto as epífises são compostas por osso esponjoso envolto por uma fina camada de osso
compacto.
Nos ossos planos, a substância esponjosa situa-se entre duas camadas de substância
compacta. Nos ossos da abóbada craniana, a substância esponjosa é chamada de díploe.
Os ossos curtos são formados por osso esponjoso revestido por osso compacto, como nas
epífises dos ossos longos.
O osso é formado por um tipo de tecido conjuntivo (tecido ósseo). É caracterizado por uma
substância intersticial (osteóide):
Matriz orgânica
o 95% fibras colagéneas (tipo 1)
o
5% fibras não colagéneas (ex: osteocalcina)
Mineral
o
cálcio (fosfato de cálcio –semelhante à hidroxiapatite)
o
outros minerais (magnésio, sódio, potássio, plutónio, chumbo, estrôncio)
É constituído por três tipos de células:
Osteócitos - estão dentro da matriz óssea; há comunicação entre os osteócitos por
onde passam pequenos iões, esta característica é essencial para a manutenção da
matriz, quando esta célula morre há reabsorção pela matriz.
Osteoblastos - são responsáveis pela produção da parte orgânica da matriz, ou seja,
colágeno tipo I, proteoglicanas e glicoproteínas. Concentram fosfato de cálcio e
participam da mineralização óssea. Quando forma a matriz a mesma ao redor do
osteoblasto e que não esta calcificada ainda chama-se osteóide.
Osteoclastos - são células gigantes, intensamente ramificadas. Elas secretam para
dentro da matriz óssea iões de hidrogénio, colagenases e hidrolases, digerindo a
matriz óssea e dissolvendo os cristais de sais de cálcio. A actividade desta célula é
comandada pela calcitonina e paratormona.
Por fim, podemos ainda distinguir os dois tipos de unidades funcionais que depende da
disposição lamelar. No osso compacto temos o sistema de Havers (osteon) enquanto que no
osso trabecular temos o sistema fragmentado por lamelas.
Formação óssea
Ossificação intramembranosa – o processo parte dos núcleos de
ossificação, que se expandem e se juntam ao longo do tempo.
o
Ossos planos (principalmente crânio) e diáfise de ossos longos
o
Crescimento em largura
Página 96
Ossificação endocondral – ocorre quendo as células mesodérmicas se transformam
em células produtoras de cartilagem, antes do início da formação do osso. Processo
mais lento que o anterior.
o
Ossos curtos e epífises de ossos longos
o
Crescimento em comprimento
Remodelação óssea
Objectivos
o
Renovação do tecido ósseo
o
Adaptação a novas solicitações mecânicas
o
Homeostasia do cálcio
Unidades de remodelação óssea
o
Reabsorção (osteoclastos) / formação (osteoblastos)
o
Frequência de activação
Unidade de remodelação óssea
1º -Activação dos osteoclastos
2º -reabsorção
3º -inversão (delimitação –linha cimentante)
4º -formação (osteoblastos)
5º -mineralização
Ciclo de remodelação óssea
Activação e Reabsorção
Os osteoclastos são recrutados para a superfície e reabsorvem uma
quantidade de mineral, criando uma cavidade - lacuna de Howship - no
osso trabecular. Essa fase dura em torno de duas semanas e é seguida por
um período de aparente inactividade no sítio da reabsorção.
Inversão
Durante essa fase, os osteoclastos desaparecem e são substituídos por macrófagos, cuja
função não está inteiramente elucidada, mas que parece ser a de depositar uma substância
que inicia a cimentação. Como esse processo ocorre entre a remoção do osso e a sua
subsequente substituição, é chamado de fase de reversão.
Formação e mineralização
Por um sinal desconhecido, os osteoblastos - células que sintetizam a nova matriz - aderem-se
à superfície da cavidade. Essas células sintetizam colagénio e outras proteínas não
colagenosas, que são secretadas dentro da cavidade para formar o osteóide, uma matriz não
mineralizada, que o será mais tarde, formando-se osso novo. Essa fase de formação pode levar
vários meses para se estabelecer.
Página 97
Portanto, unidade de remodelação óssea é uma porção de osso continuamente reabsorvido
(pelos osteoclastos) e depositado (pelos osteoblastos) a um ritmo (frequência de activação)
que permita não só a renovação do tecido ósseo mas também a sua adaptação a novas
solicitações mecânicas e a manutenção dos níveis sanguíneos de cálcio adequados.
Pico de massa óssea é a altura na vida de um indivíduo em que a sua densidade óssea é maior.
Ocorre por volta dos 30 anos.
O aumento dos níveis de remodelação óssea leva ao aparecimento de zonas de perfuração e
zonas de fragilidade, podendo levar à formação de neuropatologias.
Fisiologia do Cálcio
Coagulação do sangue
Actividade muscular – papel importante na contracção dos músculos cardíaco, liso e
esquelético.
Condução nervosa – importante para a excitabilidade normal da célula e condução do
impulso nervoso
Integridade da membrana celular – manutenção da permeabilidade da membrana de
todas as células vivas.
Normalmente o nível plasmático de cálcio é mantido dentro de limites estreitos: 2.10 –2.60
mmol / Lou 8.5 –10.5 mg / 100 ml. Mas quando este reduz abaixo dos limites gera-se
hipocalcemia, que se pode repercutir em:
Excitabilidade aumentada do SNC –por aumento da permeabilidade aos iões sódio,
podendo levar a despolarizações espontâneas
– Fenómenos de tetania: início para valores de 6,5mg/dl e letais para 4mg/dl
– Sinais de Chvostek e Trousseau
– Espasmo laríngeo
Dilatação e insuficiência cardíaca
Alterações da coagulação
Por outro lado se esse nível exceder o limite superior gera-se hipercalcemia, que se repercute
em:
Depressão do SNC (12 mg/dl – 15mg/dl)
Diminuição do intervalo QT
Obstipação
Precipitação de cristais de fosfato de cálcio (17 mg/dl)
Absorção
O cálcio é absorvido parcialmente no intestino (cerca de 35% - dependente da
quantidade ingerida).
A absorção intestinal do cálcio ocorre fundamentalmente através de um mecanismo
de transporte activo (dependente da Vit D).
Ocorre excreção de cálcio através das secreções intestinais e da descamação da
mucosa, pelo que cerca de 90% se elimina nas fezes.
Eliminação
Cerca de 10% do cálcio ingerido é excretado na urina.
Página 98
Cerca de 99% do filtrado glomerular é reabsorvido. Aproximadamente 90% nos
túbulos proximais e na ansa de Henle e os restantes 10% nos túbulos distais e nos
ductos colectores de forma selectiva, dependente da concentração plasmática de
cálcio.
O factor mais importante no controle da excreção renal é a PTH.
Fisiologia do fósforo
Síntese de nucleótidos – fundamental para a síntese de DNA e RNA
Metabolismo energético – formação de fosfocratina, ADP e ATP
Mensageiros celulares – síntese de segundos mensageiros como o AMPc e o GMPc.
A absorção de fósforo é feita de forma quase total por difusão passiva (excepto para o fosfato
eliminado nas fezes em combinação com o cálcio não absorvido), sendo praticamente todo
absorvido.
A eliminação de fosfato é praticamente totalmente renal, dependente da concentração
plasmática e da taxa de filtração.
Hormonas
A parathorma é uma hormona polipeptídica (84 a.a.) produzida nas paratiróides que é
estimulada por hipocalcemia, desencadeando uma resposta hipercalcemiante e
hipofosforemiante. Atinge o osso (aumento da remodelação óssea), rim (aumento da
reabsorção do cálcio e diminuição da reabsorção do fósforo) e o intestino (aumento da
absorção do cálcio).
A vitamina D é uma hormona esteróide (liposdolúvel) que se forma na pele sob a acção dos
U.V (colecalciferol). É hidroxilada no fígado (25 –hidrocolecalciferol) e no rim (1,25 dehidrocolecalciferol). Pode ser ingerida na dieta e efectua uma resposta hipercalcemiante e
hiperfosforemiante. A ausência desta causa osteomalácea e raquitismo. Atinge o osso
(aumento da reabsorção óssea e mineralização da substância osteóide), rim (aumento da
reabsorção do cálcio e do fósforo) e o intestino (aumento da absorção do cálcio e do fósforo).
A calcitonina é uma hormona polipeptídica (32 a.a.) produzida na tiróide (células C) que é
estimulada por hipercalcemia desencadeando uma resposta hipocalcemiante e
hipofosforemiante. Atinge o osso (diminuição da reabsorção óssea), rim (diminuição da
reabsorção do cálcio e do fósforo) e o intestino (pensa-se que diminui a absorção de cálcio).
Existem ainda outras hormonas como os estrogénios, androgénios, hormonas tiroideias,
corticoesteróides e a hormona do crescimento que afectam a remodelação óssea.
Biomecânica
Do ponto de vista biomecânico, o tecido ósseo tem um comportamento:
Bifásico – apresentando resistência e leveza devido à presença de compostos
orgânicos (fibras de colagénio tipo I -30%) e compostos inorgânicos (matriz de cristais
de Ca – 70%). Se o osso só tivesse cristais de cálcio comportava-se como o aço mas
como tem colagénio também este confere-lhe ductibilidade e capacidade de absorção
de tensões
Não – linear – pois depende de diversos factores como a idade, o envelhecimento, a
regulação hormonal, entre outros.
Elástico – Plástico. A resposta elástica ocorre no início da solicitação do osso, onde o
comprimento de mesmo volta ao comprimento inicial após a retirada da carga. Esta
Página 99
não excede 3% do comprimento total do osso. A resposta plástica é uma deformação
permanente do osso (mesmo após a retirada da carga) decorrente de microfracturas e
desestruturação trabecular. Antecede a fractura macroscópica do osso.
Anisotrópico – o tecido ósseo define-se por diferentes propriedades mecânicas em
diferentes direcções. A grande maioria dos biomateriais é isotrópico, ou seja, define as
suas propriedades apenas numa direcção que é igual às outras
Resistência óssea - reflexo da quantidade de osso e da qualidade óssea.
Quantidade óssea
o Massa óssea
o Densidade mineral óssea (g/cm2)
o Tamanho ósseo
Qualidade óssea
o Macroarquitectura (geometria óssea)
o Microarquitectura
Espessura e número de trabéculas e conectividade trabecular
Espessura e porosidade cortical
o Remodelação ou turnover ósseo : formação e reabsorção
o Propriedades dos materiais
Mineralização
Microfracturas
Pontes de colagéneo e tipos de colagéneo
Osteoporose – doença sistémica do esqueleto (e silenciosa) caracterizada por uma diminuição
da resistência óssea, predispondo a um aumento do risco de fractura.
Características:
Densidade mineral óssea baixa (DMO) (≥ -2,5, quando a normal é > -1,0)
Deterioração da micro-estrutura óssea
Fragilidade óssea
Risco aumentado de fractura
É a mais comum das doenças ósseas metabólicas
Não é exclusiva mas está ligada ao envelhecimento
Fracturas osteoporóticas: coluna vertebral, colo do fémur, extremidade distal do
punho (Colles). A fractura do colo do fémur está associada à elevada mortalidade da
doença
Página 100
Agradecimentos:
Sofia Faria (autora do capítulo Sentidos Químicos, Sangue, Sistema Digestivo e Sistema
Locomotor)
Inês Amorim (resumos deixados escritos à mão de algumas matérias)
Vasco Duarte (Sistema Renal, Sistema Respiratório, Sistema Cardiovascular – uma parte
Ana Mafalda Valente (Sistema Nervoso, Cardiovascular)
Dude que deixou os outros resumos de fisiologia (muito obrigado)
Guyton (um bacano)
Página 101