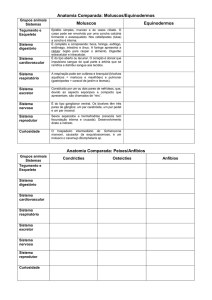
")
Visualização do documento
Cap+¡tulo 8 - Moluscos(1).doc
(97 KB) Baixar
P á g i n a | 63
Moluscos – Zoologia Aquática – Engenharia de Aquicultura
Capítulo 8
Moluscos
Os moluscos (do latim molluscus, mole) constituem um grande filo de animais invertebrados, marinhos, de água doce ou
terrestres, que compreende seres vivos como os caramujos, as ostras e as lulas. Tais animais têm um corpo mole e nãosegmentado, muitas vezes dividido em cabeça (com os órgãos dos sentidos), um pé muscular e um manto que protege uma
parte do corpo e que muitas vezes secreta uma concha. A maior parte dos moluscos são aquáticos, mas existem muitas
formas terrestres como os caracóis. A biologia dos moluscos é estudada pela malacologia, mas as conchas - ainda do ponto
de vista biológico, não do ponto de vista dos coleccionadores - são estudadas pelos concologistas. O filo Mollusca é o segundo
filo com a maior diversidade de espécies, depois dos Artrópodes (cerca de 50 000 espécies viventes e 35 000 espécies
fósseis) e inclui uma variedade de animais muito familiares. Essa popularidade deve-se, em grande parte, às conchas desses
animais que servem como peças para coleccionadores. O filo abrange formas tais como as ostras, as lulas, os polvos e os
caramujos. Os moluscos são variados e diversos, incluindo várias criaturas familiares conhecidas pelas suas conchas
decorativas ou como marisco. Variam desde os pequenos caracóis e amêijoas até ao polvo e à lula (que são considerados os
invertebrados mais inteligentes). A lula-gigante é possivelmente o maior invertebrado, e, exceptuando as suas larvas e, para
além de alguns espécimes jovens recentemente capturados, nunca foi observada viva. A lula-colossal poderá ser ainda
maior. Uma lula-colossal foi encontrada congelada por pescadores.O comprimento da lula chegou a aproximadamente
trinta metros.
GRUPOS DE MOLUSCOS DE INTERESSE NA AQUICULTURA
Existem dez classes de moluscos, oito que ainda vivem e duas que só são conhecidas através de fósseis, constituindo
250.000 espécies. Desses os mais importantes para a área de aquicultura correspondem ao moluscos gastrópodes, bivalves
e cefalópodes, por suas peculiaridades relacionadas ao processo de cultivo ou pelo fato de servirem como importante fonte
nutricional para o seres humanos. Assim podemos destacar:
(1) Bivalvia (do latim bi, duplicado + valva, porta de duas folhas, valva) anteriormente Pelecypoda e Lamellibranchia, é a
classe do filo Mollusca que inclui os animais aquáticos popularmente designados por bivalves, incluindo os familiares
mariscos, ostras e mexilhões. Estes organismos são comprimidos nas laterais e possuem uma concha carbonatada composta
de duas valvas, encaixadas em dobradiça dorsalmente, que envolvem todo o corpo e o protegem do meio externo. O pé
como o restante do corpo, é lateralmente comprimido daí a origem do nome Pelecypoda (pé-machadinha) e a cabeça e mal
desenvolvida. Dentro da concha ocorre uma grande cavidade delimitada pelo manto (cavidade do manto) que abriga
grandes brânquias com função de trocas gasosas e coleta de material, o pé muscular e o sifão inalante e exalante para a
entrada e saída da água, que traz oxigênio, depois absorvido por difusão direta pelas lâminas branquiais. Essa forma de
alimentação característica conhecida como filtração também aumenta o risco de intoxicações, pois este tipo de alimentação
é comum em animais fixos representando um grande risco caso a ostra e/ou marisco entre em contato com poluição. O
grupo surgiu no Câmbrico e é atualmente muito diversificado, com cerca de 15,000 espécies. A separação das diferentes
sub-classes faz-se pelo tipo e estrutura das guelras nos organismos vivos, e pelas características das valvas nos bivalves
fósseis. Três grupos principais podem ser identificados pela natureza de suas brânquias: Protobrânquios (mais primitivo
dos bivalves existentes), Lamelibrânquios (a maioria das espécies de bivalves) e os Septobrânquios (altamente
especializados);
(2) Gastropoda (do grego gaster, estômago + pous, podos, pé) constituem uma grande classe de moluscos definida pela
primeira vez por Georges Cuvier em 1797, sendo a mais bem sucedida dentro do seu filo. A classe Gastropoda é a maior
classe de moluscos, habitam uma larga variedade de habitats e com certeza são os moluscos mais importantes em termos
evolutivos. As espécies marinhas adptaram-se à vida em todos os tipos de fundos, bem como uma existência pelágica
(animais que vivem nadando na coluna d’água). Eles invadiram a água doce, e os caramujos pulmonados e vários outros
grupos conquistaram a terra eliminando as brânquias e convertendo a cavidade do manto em um pulmão. Conta com cerca
de 60,000 a 75,000 espécies atuais que incluem os caracóis e lesmas terrestres (ca. 1/3 das espécies), bem como um grande
número de formas marinhas e de água doce (ca. 2/3). O registo fóssil dos gastrópodes é igualmente abundante. A grande
maioria dos gastrópodes tem o corpo protegido por uma concha, geralmente espiralada sobre o lado direito embora
algumas formas (como as lapas) tenham evoluído uma concha mais simples. Os gastrópodes apresentam uma cabeça bem
marcada, munida de dois ou quatro tentáculos sensoriais e uma boca com rádula. A cabeça encontra-se unida a um pé
ventral musculado em forma de pala. O seu desenvolvimento embrionário caracteriza-se por torção da massa visceral,
coberta pelo manto, que surge no adulto enrolada sobre um dos lados de forma a ser acomodada na concha. As lesmas têm
uma concha apenas vestigial e, em consequência, os efeitos deste enrolamento são diminutos sendo o corpo linear.
(3) Cephalopoda (do grego kephale, cabeça + pous, podos, pé) são a classe de moluscos marinhos a que pertencem os
polvos, as lulas, sépias, nautilus, argonauta e os chocos. Os cefalópodos (gr. Kephale, cabeça + podos, pé) são os moluscos
mais altamente desenvolvidos em estruturalmente diferentes. Em muitos aspectos, podem ser considerados como o grupo
mais avançado dentre os invertebrados viventes, graças ao cérebro desenvolvido e a capacidade de formação de imagens.
Em contraste com a figura estereotipada dos moluscos que apresentam locomoção lenta a base de mecanismos mucosos e
ciliares, o cefalópodos são predadores rápidos e principalmente pelágicos. A cabeça grande tem dois olhos conspícuos e
uma boca central que é ornada por um bico quitinoso, a qual é circundada por dez braços carnosos (lulas) e oito (polvo)
contendo ventosas em forma de taça, e no caso das lulas o quarto par de braços são tentáculos retráteis longos. Além do
bico, na boca que existe a característica mais marcante dos cefalópodos, a rádula, uma esspécie de lingua com dentículos
que funciona como uma lixa, para fragmentar o alimento. O corpo delgado, cônico, tem uma nadadeira triangular ao longo
de cada lado da extremidade afilada nas lulas e sépias, ou ainda, como no caso dos polvos uma grande massa cefálica visceral
globosa ligada aos braços. A cabeça e o corpo unem-se por um pescoço, ao redor do qual o bordo livre do manto forma um
colarinho frouxo. Abaixo do pescoço há um funil muscular o sifão. São animais extremamente rápidos, tendo desenvolvido
um sistema de propulsão na forma de trapezio (jato-propulsão), que é uma modificação do pé dos restantes moluscos. A
pele contém células pigmentadas, chamadas cromatóforos, que mudam de cor para efeitos de comunicação e camuflagem;
esta mudança de cores é dada por ações nervosas diretas. Os estatocistos são os orgãos de equilíbrio. A concha está ausente
nos polvos, é interna nos chocos e lulas e é externa no nautilus e no argonauta. As lulas e polvos apresentam a chamada
"glãndula de tinta". Quando o animal é atacado,ele elimina o conteúdo preto da glândula ,que o envolve em uma nuvem
escura e lhe permite fugir do inimigo.Esta tinta é o famoso nanquim legítimo dos pintores chineses.
ESTRUTURA CORPORAL
A epiderme é denominada de palio ou manto e é ricamente ornamentada com células mucosas que se encarregam de
secretar os materiais a base de carbonato de cálcio que irão compor a concha do animal. Na região da concha há uma
proeminência que cria uma câmara chamada de cavidade do manto, onde estão alojados os elementos respiratórios
(brânquias e pulmão). O sistema esquelético é representado pela concha ou valva, constituindo normalmente um
exoesqueleto calcário ou endoesqueleto e algumas vezes ausente. A concha é formada por três camadas: O perióstraco
externo é composto de um material protéico córneo e bronzeado com quinona, chamado conchina. As camadas mais
internas da concha consistem de carbonato de cálcio. A camada calcária média é geralmente prismática, ou seja, o mineral
deposita-se como cristas verticais cada um deles circundado por uma matriz protéica fina. As camadas mais internas se
depositam como lâminas perpendiculares em cima de uma matriz orgânica e recebem a denominação de lamela. Algumas
vezes as lâminas são paralelas à superfície e têm um aspecto liso e lustroso, este tipo de lâmina corresponde a nacarada
ou madrepérola. As conchas dos gastrópodes exibem uma grande variedade de formas, cores e padrões, em alguns casos
a concha só é espiralada em estágios juvenis desaparecendo logo depois com o crescimento. As lapas por exemplo
apresentam a concha como se fosse um chapéu chinês. Em alguns gastrópodos também ocorreu a redução do manto e a
perda da concha (lesmas). Provavelmente a grande redução da concha fez com que ela se enterrasse dentro dos tecidos do
manto.
Nos gastrópodes a concha é uma estrutura espiral composta de várias voltas tubulares e contendo a massa visceral do
animal. No vértice do cone estão as espirais menores e na base esta a maior que termina em uma abertura a partir do qual
a cabeça e o pé se protraem. As espirais acima da volta do corpo são denominadas de espiral. A maneira como as espirais
se orientam na concha determinam o formato da mesma que pode variar muito de acordo com as espécies. Apesar do grande
número de formas e denominações as conchas mais comuns são aquelas denominadas de: (1) Espirais planas são aquelas
que possuem as voltas uma em cima da outra dessa forma aparentam ser achatadas lateralmente; (2) Espirais cônicas são
aquelas que apresentam as voltas uma fora da outra e se distingue normalmente um vértice pontiagudo dando a concha a
aparência de um cone; (3) Espirais helicoidais são aquelas que ocorrem na família Helicidae do caramujo de jardim como
o Helix aspersa, onde as voltas se distribuem semelhante a uma hélice. O vértice da concha é baixo e não alto como nos
caramujos com espirais cônicas.
As conchas espiraladas evoluíram de quatro alterações principais que ocorreram no corpo dos gastrópodes: 1.
Desenvolvimento da cabeça; 2. Alongamento dorso-ventral do corpo; 3. Conversão da concha de um escudo para
um abrigo protetor profundo; 4. Torção. Primitivamente a cabeça possui um par de tentáculos, co m um olho em cada
base tentacular. A concha na maioria é uma espiral que funciona como um abrigo portátil, formada a partir do aumento da
altura e redução na abertura, transformando o escudo em um cone. Entretanto o cone impedia a exploração de fendas e
buracos menores, a partir disto a evolução dos gastrópodes constituiu a espiralização da concha acima da cabeça, que
resolveu este problema. Com esta evolução houve uma redução da cavidade do manto o que provavelmente ocasionou a
redução das brânquias, músculos retratores e rins a um par único. A torção foi a modificação mais distintiva observada nos
gastrópodes, alguns pesquisadores acreditam que a torção tem como vantagens a proteção da cabeça ou a utilização por
parte das brânquias ou órgãos sensoriais da corrente de água que chega na cavidade do manto. As primeiras conchas eram
do tipo plano espiral, ou seja, cada volta repousava fora da volta precedente e o diâmetro da podia se tornar relativamente
grande e frágil. Para resolver esse problema houve o enrolamento assimétrico, no qual as voltas se situam uma sobre a
outra, mas em torno de um eixo central denominado de columela. Tal concha é mais compacta e mais forte que uma concha
plano espiral. A nova concha formada inclinou-se para o lado direito com a abertura para o lado esquerdo. Para recolher os
pés e a cabeça desenvolveu-se um músculo retrator denominado de músculo columelar, que surge no pé e se insere na
columela. Alguns gastrópodes também desenvolveram um disco córneo que fecha a abertura da concha, que corresponde
ao opérculo.
Nos bivalves a concha típica consiste de duas valvas ovais e geralmente convexas, que se prendem e se articulam
dorsalmente. Uma protuberância dorsal chamada de umbo, que surge acima da linha da articulação. As duas conchas se
prendem por meio de uma faixa protéica elástica não calcificada, chamada de ligamento de dobradiça. O fechamento da
concha é condicionado pela musculatura adutora (adução, fechar) contraída. Quando essa musculatura relaxa, a
elasticidade natural do ligamento faz com que as valvas se abram. Alguns bivalves como a vieiras (Pecten sp.) possuem um
condróforo, estrutura em forma de aba na dobradiça que proporciona uma grande área superficial para junção do
ligamento da dobradiça interna. O deslizamento lateral das conchas durante o fechamento em algumas espécies é evitado
por meio de dentes ou cristas localizado na linha de dobradiça. A concha é formada externamente pelo periostráco formado
por duas a quatro camadas calcárias que é geralmente espesso e protege a concha contra a dissolução do carbonato de
cálcio subjascente mantendo a integridade da concha. A camada prismática é formada por deposições de aragonita ou
aragonita mais calcita em prismas que se encerram na parte mais interna da concha que é a camada nacarada. O manto
que se acomoda debaixo das valvas, e na sua borda verifica-se a formação de três pregas. A prega interna contém músculos
radias e circulares, a média tem função sensorial e a externa secreta a concha. O manto se prende a concha for fibras
musculares que descrevem uma linha semicircular. A linha de junção do manto imprime-se na parte interna da concha
como uma cicatriz conhecida como linha palial.
Nos cefalópodes uma concha externa só encontrada em forma fósseis como o Nautilus, nas lulas e sépias é reduzida e nos
polvos se encontra completamente ausente. A concha do Nautilus é planoespiral bilateralmente simétrica e divide-se
internamente em septos transversais (câmaras), de tal forma que somente a última é ocupada pelo animal vivo. À medida
que o animal cresce, ele se move periodicamente para a frente, e a parte posterior do manto secreta um novo septo. Cada
septo é perfurado no meio, e se conecta com um tubo calcário que abriga um cordão de tecido delgado (denominado de
sifúnculo) que se estende ao longo dos septos. O sifúnculo secreta um gás através da parede porosa do tubo no interior
das câmaras vazias, fazendo com que a concha flutue, permitindo que o animal nade. A concha é composta de uma camada
externa semelhante a uma porcelana, que contém prismas de carbonato de cálcio em uma matriz orgânica, e de uma camada
nacarada interna. Exceto quanto ao Nautilus, todos os cefalópodos vivos pertencem à subclasse Coleoidea, na qual a concha
encontra-se reduzida e interna ou absolutamente ausente. Acredita-se que esses cefalópodos tenham evoluído a partir de
algum nautilóide de concha reta inicial, cuja concha tornou-se completamente envolvida pelo manto. Na linhagem evolutiva
que levou às lulas a concha reduziu-se a uma longa placa quitinosa achatada, chamada de pena ou gládio. A pena exerce a
mesma função do sifúnculo, ou seja, funciona como um bolsão de gás que controla a flutuação do corpo na água.
PRODUÇÃO DE PÉROLAS
O processo de formação de pérolas ocorre nos moluscos conhecidos como Bivalves especificamente no grupo de ostras
perlíferas e alguns mariscos. Nessas espécies objetos ocasionalmente alojados entre o manto e a concha inicia-se o
processo de formação da pérola que se constitui num processo de defesa do molusco contra corpos estranhos que
invadem o seu organismo. Um nematoide ou grão de areia, instalado entre a camada epitelial e a concha, provoca uma
irritação que estimula a secreção de certas células mucosas, produzindo várias camadas envolventes de nácar sobre o
corpo estranho, formando a pérola. Se o objeto intruso se torna um núcleo ao redor do qual depositam camadas
concêntricas de concha nacarada e se mover de um lado para o outro durante a secreção, a pérola torna-se esférica. No
entanto é muito comum a pérola se aderir a concha se incrustando completamente a ela. As pérolas mais valiosas são
produzidas pelas ostras perlíferas como a Pinctada margaritifera e Pinctada mertensi. Sementes de pérolas (glóbulos
microscópicos) podem ser produzidas por mariscos de água doce e transplantadas para estas duas espécies que produzem
um nácar de melhor qualidade. Esse procedimento é realizado porque a introdução de sementes pode ser direcionada para
as pregas do manto, impedindo que a pérola se ligue ao o nácar da camada interna da concha das ostras perlíferas. Algumas
pérolas formam-se em bivalves de água doce, mas os tipos mais valiosos provêm de ostras perlíferas marinhas do golfo
pérsico e do oceano Índico. Os japoneses desenvolveram a técnica de produção artificial de pérolas introduzindo e
costurando pequenas esferas, ou objetos de metal com formatos diversos (flor, animais, símbolo religiosos etc.) no manto.
Esse processo ocorre quando as ostras são retiradas de engradados e durante alguns minutos abrem a concha. Nesse
momento hastes de madeira são colocadas entre as valvas para que a concha permaneça aberta. Os objetos são então
suturados ao manto, e a haste retirada, a concha se fecha e o molusco e devolvido ao engradado. Os bivalves são mantidos
nos engradados durante alguns meses até que as pérolas “cultivadas” sejam formadas. As ostras mais comumente
comercializadas no mercado são: (1) Pérolas Cultivadas de Água Doce: As pérolas de água doce são acessíveis e oriundas
de lagos e rios da China. Possuem formato assimétrico, brilho médio a alto, e medem entre 3,5 mm a 7,0 mm. Essas pérolas
são na maioria brancas, mas podem ter um sombreado rosa ou vermelho claro; (2) Pérolas do Taiti: As pérolas do Taiti
têm uma coloração única, negra a verde-acinzentada. As maiores, de 11 mm a 13 mm, são obtidas nos atóis vulcânicos e
recifes do Taiti. São redondas na maioria das vezes e possuem bastante brilho. São mais caras devido à escassez e ao visual
distinto; (3) Pérolas Akoya: As pérolas Akoya são pérolas de água salgada da China e do Japão. Possuem formato
arredondado e simétrico com poucas manchas e medem entre 6 mm a 8,5 mm. As pérolas Akoya possuem mais brilho que
as de água doce, preços médios e uma cor clara que varia de creme a branca/rosa; (3) Pérolas dos Mares do Sul: As
enormes pérolas dos Mares do Sul (de até 13 mm), obtidas na Austrália, na Indonésia, e nas Filipinas, variam em cor de
branco até o preto. Podem ser perfeitamente redondas ou ligeiramente assimétricas e brilho médio a alto. Os preços variam
com o brilho.
LOCOMOÇÃO
Os gastrópodes podem apresentar um pé com a sola ciliada (atrito) com muitas glândulas (aderência), ou no caso dos
gastrópodes pulmonados uma glândula podal. Tanto os cílios como as glândulas facilitam a locomoção. A locomoção,
entretanto é promovida pela contração dos músculos longitudinal e dorso-ventral que formam as ondas diretas de trás
para frente do corpo do animal, e retrógradas que é o oposto e envolvem a contração dos músculos transversais.
Na maioria dos bivalves o pé ficou comprimido, tornando-se uma estrutura laminar que se direcionou anteriormente como
uma adaptação à escavação. O movimento podal é efetuado por uma combinação de pressão sanguínea e das ações
musculares dos protratores e dos retratores, homólogos aos retratores podais dos outros moluscos. O sangue é bombeado
para o interior do pé, e em combinação com a contração da musculatura protratora estende-o para dentro da areia ou do
lodo e depois os músculos retratores puxam a concha em direção ao pé.
A maioria dos cefalópodes nada por meio de propulsão a jato, expelindo rapidamente a água da cavidade do manto. O
manto que é um envoltório que circunda a vísceras contém fibras musculares radiais e circulares. Durante a fase inalante
da circulação hídrica, as fibras circulares relaxam e os músculos radiais contraem-se. Essa ação aumenta o volume da
cavidade do manto, e a água entra rapidamente na parte lateral entre a margem anterior do manto e a extremidade
posterior da cabeça. Durante a fase exalante, a contração dos músculos circulares não só aumenta a pressão da água dentro
da cavidade, como também fecha as bordas do manto firmemente ao redor da cabeça. As valvas protuberantes selam
ventralmente a cavidade do manto, e a água é, em conseqüência, forçada a sair através do funil tubular ventral. Dessa forma,
as contrações e expansões alternadas fazem com que a água seja acumulada e expelida para fora do manto. “Para nadar
com a cauda para a frente”, o sifão é dirigido para o lado dos braços, “para locomover-se com a cabeça para a frente”,
a ponta do sifão para forçar a água na direção da cauda. As nadadeiras funcionam como lemes estabilizadores e
direcionadores da natação. No caso das lulas os músculos radiais são mais ativos durante a natação de fuga e fazem com
que a cavidade do manto tenha uma expansão acima da capacidade normal (hiperinflam). Na natação lenta, a cavidade do
manto expande-se por meio de enrolamento elástico das fibras colágenas do manto, e as ondulações das nadadeiras
proporcionam certo poder de propulsão Os polvos também são capazes de se locomover através de jatos d’água, entretanto,
os órgãos mais importantes para a locomoção são tentáculos (Braços longos) que dispõem de discos com sucção
(ventosas), utilizados para puxar o animal ou ancorá-lo ao substrato. É muito comum a observação de várias espécies de
polvo rastejando-se sobre o substrato rochoso dos recifes coralinos.
NUTRIÇÃO E DIGESTÃO
Os moluscos gastrópodes aquáticos alimentam-se principalmente de algas e outros organismos que crescem nas rochas. Já
os moluscos terrestre atacam progressivamente a superfície de folhas de diversas espécies de plantas, especialmente as
leguminosas. O sistema digestivo é do tipo completo, onde a faringe ou a cavidade bucal apresenta a rádula, que constituise o órgão ralador do substrato para obtenção do alimento. A cavidade bucal apresenta-se revestida por uma cutícula, e o
seu piso é bastante espesso, cartilaginoso e muscular e é denominado de odontóforo. Na boca existem também fileiras de
dentes quitinosos transversais que se ordenam em uma série longitudinal formam a rádula. A rádula surge de uma bolsa
profunda, chamada saco radular, proveniente da parede posterior da cavidade bucal. Tanto o odontóforo como a rádula
podem se projetar para fora em direção ao substrato, por ação de seus músculos específicos. O odontóforo cartilaginoso é
munido por dois conjuntos de músculos, os protratores que projetam a língua radular para fora em direção ao substrato
a ser consumido e os retratores que recolhem a língua radular para dentro da boca trazendo os fragmentado para dentro
da boca afim de formar o bolo alimentar com a saliva. A rádula também apresenta esse conjunto de músculos sendo os
protratores para movimentar a rádula para frente e o retratores para movimentar a rádula para trás, criando o
movimento de serra.
A observação da estrutura e do número de dentes da rádula nos permite determinar o hábito alimentar dos gastrópodes.
Nos carnívoros eles se apresentam com formato pontiagudo, sendo maiores em tamanho, mas em menor número na
fileira. Os carnívoros apresentam ao redor da boca uma tromba ou probóscide extensível, que permite ao animal alcançar
e penetrar em áreas vulneráveis da presa. Na alimentação, a probóscide projeta-se para fora por meio de pressão
sanguínea, e por estímulos químicos de proteínas específicas liberadas pela presa ou pela carniça. A dieta dos carnívoros é
baseada em moluscos bivalves, outros gastrópodos, ouriços-do-mar, estrelas-do-mar, poliquetos, crustáceos e até peixes.
Alguns imobilizam as presas utilizando secreções salivares com ácido sulfúrico e outras toxinas (dentes radulares tóxicos)
e outros podem utilizar o pé para asfixiar suas vítimas. Em moluscos perfuradores de conchas uma substância ácida é
produzida por uma glândula eversível localizada na parte anterior da sola do pé, que reduz a estrutura e desmineraliza a
concha, permitindo a ação da rádula para a abertura de um pequeno orifício.
Os gastrópodes herbívoros apresentam a rádula reduzida em tamanho com cúspides do tipo serra e uma fileira
longitudinal única de dentes utilizada para fender células de algas ou superfícies vegetais. Apesar de serem menores quando
comparados ao dos carnívoros, as o número de dentes na fileira tende a ser numerosos podendo chegar a 12 dentes numa
única fileira, alguns deles com impregnações de minerais que os tornam altamente resistentes ao atrito com superfícies
duras de rochas. Alguns moluscos da família Elysidae incorporam cloroplastos a suas células glandulares digestivas, onde
a fotossíntese então ocorre. Em muitas espécies herbívoras, o esôfago ou a parte anterior do estômago encontra-se
modificada em papo e moela, revestida de cutícula ou conter grãos de areia. As amilases...
Arquivo da conta:
evaldofarteski
Outros arquivos desta pasta:
IGSg(1).dll (1361 KB)
libEGL.dll (58 KB)
libGLESv2.dll (507 KB)
IGOptExtension.dll (142 KB)
alchemyext.dll (15 KB)
Outros arquivos desta conta:
Documentos
Galeria
Privada
Relatar se os regulamentos foram violados
Página inicial
Contacta-nos
Ajuda
Opções
Termos e condições
Política de privacidade
Reportar abuso
Copyright © 2012 Minhateca.com.br
")