11 - NUCLEÓTIDOS
Nucleótidos e nucleósidos: Estrutura, composição geral, ligação fosfodiéster, tipos mais
comuns; Funções específicas dos nucleótidos (com exemplos): informação, transporte,
cofactores/coenzimas, regulação metabólica)
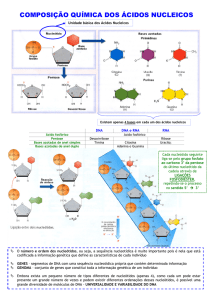
Cada nucleótido é constituído por 3 componentes: uma pentose, uma base azotada e
um grupo fosfato. Uma molécula que não tenha o grupo fosfato, isto é, que seja constituída
apenas pelo açúcar e pela base azotada, é chamada nucleósido.
As pentoses são açúcares com 5 carbonos, convencionalmente numerados de 1’ a 5’.
Há 2 tipos de pentoses: a desoxirribose (em que o carbono 2’ se encontra ligado a 2
átomos de hidrogénio) e a ribose (no mesmo carbono um dos átomos H é substituído por um
grupo hidroxilo). É a ocorrência destas que dá o nome ao nucleótido, respectivamente,
desoxirribonucleótido ou ribonucleótido. Ambos os tipos de pentose estão na sua forma βfuranose, um anel aromático resultante de 5 carbonos, fechado, em que o grupo hidroxilo da
ligação hemiacetal aponta para cima.
As bases azotadas estão organizadas em 2 grupos: as purinas, formadas por um anel
aromático duplo, e as pirimidinas, formadas por um anel simples. As purinas principais são a
adenina (A) e a guanina (G). E as pirimidinas mais conhecidas e frequentes são a citosina (C), a
timina (T) (exclusiva dos desoxirribonucleótidos) e o uracilo (U) (exclusivo dos ribonucleótidos).
Apesar disso, podem ocorrer ocasionalmente outras bases azotadas minoritárias ou mesmo
trocas entre desoxi e ribonucleótidos a nível da timina e do uracilo.
A ligação que se estabelece entre o açúcar e a base azotada chama-se ligação N-βglicosídica e é estabelecida pela remoção de um grupo hidroxilo da pentose e um hidrogénio da
base, libertando-se uma molécula de água e formando-se uma ligação covalente. Esta ligação
tem a designação de N-β-glicosídica, porque é a conexão de um átomo de azoto, N, com um
glícido na forma β. O grupo fosfato, por sua vez, está ligado ao carbono 5’ da pentose (por vezes
ao 3’) através de uma ligação O-glicósida (entre um átomo de oxigénio e o glícido).
Os nucleótidos podem, para além disso, estabelecer ligações entre si. Quando são até
cerca de 50 unidades básicas formam uma cadeia que se designa oligonucleótido. Quando a
cadeia é maior, polinucleótidos. As ligações fosfodiéster são as responsáveis pela formação de
tais cadeias, nas quais o grupo fosfato ligado ao carbono 5’ se liga ao grupo hidroxilo do carbono
3’ do nucleótido seguinte. A sequência tem sempre a direcção de crescimento 5’-3’. A cadeia
assim formada é dotada de uma polaridade específica e de 2 fins distintos: um 5’ e outro 3’.
Esta sequência nucleotídica constitui, então, uma cadeia cujo esqueleto é uma
alternância de grupos fosfatos e pentoses, ao qual se ligam grupos laterais, as bases azotadas.
Como o açúcar tem grupos hidroxilos livres, estes vão criar pontes de hidrogénio com moléculas
de água. Também o grupo fosfato, como se apresenta ionizado e negativamente carregado em
pH perto de 7, estabelece interacções iónicas com outras moléculas. Estes factos tornam o
esqueleto da cadeia hidrofílico.
As bases azotadas constituintes dos nucleótidos são, também elas, dotadas de
propriedades químicas que consequentemente lhes conferem algumas características
especificas a nível estrutural e funcional. Como são hidrofóbicas a pH neutro, aumentando a
afinidade com a água em pH mais ácido ou básico, as bases de um nucleótido têm tendência a
estabelecer ligações de Van der Waals ou interacções dipolo-dipolo entre si, para se protegerem
da água do meio circundante. Para além disso, os átomos de azoto, bastante electronegativos,
formam entre bases, pontes de hidrogénio, estabelecendo-se uma grande complementaridade
entre certos pares de bases (C-G e A-T(U)).
São estas características dos nucleótidos que vão levar à formação dos ácidos nucleicos
como os conhecemos e lhes vão permitir desempenhas as funções a que estão destinados.
Funções específicas dos nucleótidos:
•
Subunidades dos ácidos nucleicos;
•
Informação genética;
•
Transporte de energia química;
•
Nucleótidos de adenina são constituintes de cofactores;
•
Alguns são reguladores metabólicos;
•
Informação genética.
Transporte de energia química:
Os nucleósidos podem ser mono-, di- ou trifosfato, de acordo com o número de grupos
fosfato que estão ligados. Cada um desses grupos são designados por α, β, e γ, começando no
fosfato que se encontra ligado à pentose.
UTP, GTP e CTP são fontes de energia para diversas reacções, mas o nucleótido mais utilizado
pela célula para fornecer energia química é o ATP. A fosfohidrólise do ATP tem um papel
muito importante para diversas biossínteses com variação da energia livre de Gibbs positiva,
uma vez que, como a hidrólise de ligações fosfoanidrido (ligações entre os grupos fosfato β, γ e
α, β) liberta cerca de 30 kJ/mol , o equilíbrio final dessas reacções é alterado, favorecendo a
formação dos produtos.
Nucleótidos de adenina são constituintes de cofactores:
Os nucleótidos de adenina fazem parte da estrutura de uma grande variedade de cofactores com
diversas funções e, embora não participem directamente na função primária do cofactor, a sua
remoção diminui consideravelmente a acção deste. Tendo como exemplo a acetoacetilCoA
(coenzima A derivada do acetoacetato), a remoção do nucleótido de adenina desta coenzima
reduz na ordem de 106 a reactividade dessa coenzima como substrato de uma enzima do
metabolismo dos lípidos, a β-cetoacil-CoA transferase. Outros exemplos são o NAD e FAD.
Regulação metabólica:
Os nucleótidos podem também ser produzidos como mensageiros secundários nas
células, provocando alterações no interior destas, em resposta a sinais químicos externos
(mensageiros primários). O mais comum é o AMPcíclico (cAMP), que tem funções reguladoras
em todas as células, excepto nas vegetais. Outros nucleótidos reguladores são: o cGMP e o
ppGpp (produzido em bactérias, inibe a síntese de tRNA e rRNA, como resposta a uma
diminuição de síntese proteica provocada pela falta de aminoácidos disponíveis).
Informação genética:
A
informação
genética
está
codificada
na
sequência
linear
de
quatro
desoxirribonucleótidos no ADN. A informação está agrupada em unidades codificantes (há
zonas não codificantes no ADN), denominadas de codões. Cada codão é uma sequência de três
nucleótidos que codifica um aminoácido. O RNA é o responsável pela expressão da informação
genética, actuando como intermediário, uma vez que usa a informação codificada no ADN de
forma a especificar a sequência de aminoácidos de uma determinada proteína. Portanto a
estrutura de cada proteína, e consequentemente de todas as biomoléculas e componentes
celulares, é produto da informação contida na sequência de nucleótidos dos ácidos nucleicos. A
capacidade de armazenar e transmitir a informação genética aos descendentes é uma condição
fundamental da vida.
1. Ácidos Nucleicos
DNA
Um importante trabalho para a descoberta da estrutura do DNA foi o de Erwin Chargaff (década de 40).
Descobriu que os 4 tipos de nucleótidos de DNA ocorrem em percentagens diferentes em moléculas de DNA
de organismos diferentes.
Através disto foi possível concluir que:
1) A composição em bases de DNA geralmente varia de uma espécie para outra.
2) Amostras de DNA isoladas de diferentes tecidos de um dado organismo têm a mesma composição
de nucleótidos.
3) A composição de bases do DNA numa espécie não muda com a idade, estado nutricional ou
ambiente de um dado organismo.
4) Em todas as moléculas de DNA celular, independentemente da espécie, o número de resíduos de
adenina é igual ao número de resíduos de timina e o nº de resíduos de citosina é igual ao número de resíduos
de guanina. Assim, o número de resíduos de purinas é igual ao número de resíduos de pirimidinas: A + T = G
+ C.
Estas relações quantitativas, também chamadas as leis de Chargaff, foram a chave para o
estabelecimento da estrutura tridimensional do DNA e abriram portas para o conhecimento de como a
informação genética está codificada no DNA (e como é processado essa informação).
Rosalind Franklin e Maurice Wilkins utilizaram difracção de raios-X para analisar fibras de DNA. Eles
mostraram (década de 50) que o DNA produz um padrão característico de difracção de raios-X. A partir deste
padrão supôs-se que as moléculas de DNA seriam helicoidais com duas periodicidades ao longo do seu eixo,
uma primária de 3,4 Å e uma secundária de 34 Å.
Em 1953 Watson e Crick propuseram um modelo tridimensional da estrutura do DNA, que tinha em
conta todos os dados recolhidos até à altura (relação A-T e C-G). Este modelo consistia numa dupla hélice, de
mão direita, de cadeias pulinucleotídicas. As partes hidrofílicas das desoxirriboses e grupos fosfato estão
voltados para fora, enquanto que as bases azotadas encontram-se no interior da hélice, com os seus anéis
hidrofóbicos de estrutura quase planar, numa posição perpendicular ao eixo. É de referir que as desoxirriboses
estão na conformação endo C-2’. Cada base do nucleótido emparelha no mesmo plano com a base
complementar na cadeia oposta, estabelecendo ligações de hidrogénio: dupla entre adenina e timina e tripla
entre citosina e guanina, daí que seja tanto mais difícil separar cadeias de DNA, quanto maior for a %GC.
Trabalhos com DNA polimerases mostraram que as duas cadeias são anti-paralelas, isto é, têm
orientações opostas.
Para ter em conta as periodicidades encontradas nos padrões de difracção de fibras de DNA, Watson e
Crick observaram que as bases “empilhadas” verticalmente no interior da dupla hélice estão a 3,4 Å de
distância. A periodicidade secundária encontrada anteriormente (de 34 Å) foi explicada pela presença de cerca
de 10 pares de bases em cada volta completa da dupla hélice. Nota: em solução aquosa, a estrutura altera-se
ligeiramente, havendo 10,5 pares de bases, por cada volta da hélice, aumentando a distância para 36 Å.
A dupla hélice é mantida por dois tipos de forças: as ligações de hidrogénio, entre bases
complementares, e interacções hidrofóbicas entre bases sobrepostas. A complementaridade entre as cadeias de
DNA atribui-se à ligação de hidrogénio entre os pares de bases. As outras interacções, muito menos
específicas do que as ligações de hidrogénio, fazem a maior contribuição para a estabilidade da molécula.
É uma molécula bastante flexível, podendo haver rotação considerável à volta de algumas ligações
pentose-fosfato. Podem ocorrer flutuações térmicas que causam dobras, estiramentos e desemparelhamento
das cadeias complementares.
Ao nível do DNA celular encontramos alguns desvios em relação ao modelo de Watson e Crick, que
desempenham importantes funções no metabolismo do DNA. Estas variações estruturais geralmente não
afectam as propriedades chave definidas por Watson e Crick: complementaridade entre cadeias antiparalelas e
as requeridas ligações A-T e G-C.
As variações na estrutura do DNA são o reflexo de três factores:
1) Diferentes conformações possíveis da desoxiribose.
2) Rotação livre em volta da ligação C-1’- N – glicosídica.
3) Rotações nas ligações da desoxirribose com o grupo fosfato.
A estrutura de Watson e Crick é também chamada DNA forma B (ou B-DNA) e é a forma mais estável
existente para a molécula de DNA. Existem no entanto duas variantes bem caracterizadas em estruturas
cristalinas, denominadas formas A e Z. A forma A ainda tem a conformação de hélice de mão direita, mas é
mais larga e tem maior número de pares de bases por volta da hélice do que da forma B e as bases azotadas
emparelham num plano com cerca de 20º de inclinação. A forma Z tem já a conformação de hélice de mão
esquerda, sendo mais estreita do que a forma B e tendo maior número de pares de bases por volta da hélice.
Não se sabe ao certo se a forma A ocorre ou não nas células, mas existem provas da existência da forma Z em
tanto procariotas como eucariotas, e pensa-se que pode ter um papel importante na regulação da expressão de
alguns genes, ou na recombinação genética.
Já foram também detectadas variações de estrutura dependentes da sequência nucleotídica, que podem
afectar a função e o metabolismo de segmentos de DNA na sua vizinhança. Por exemplo, ocorrem dobras na
hélice de DNA, sempre que 4 ou mais resíduos adenosina aparecem seguidos numa cadeia.
Outro tipo de sequências são as sequências palindrómicas. Palindroma é, basicamente, uma palavra ou
frase que se lê da mesma maneira da esquerda para a direita ou vice-versa. O termo aplica-se a regiões no
DNA com sequências repetidas na ordem inversa na cadeia complementar. Desta forma, surgem sequências
auto-complementares, dentro de cada cadeia, e portanto têm a capacidade de gerar formas de gancho de
cabelo (quando o DNA está sob a forma de cadeia simples) ou forma cruciforme. Quando a repetição
invertida ocorre na mm cadeia individual de DNA, a sequência tem o nome de repetição espelhada, não dando
origem a nenhuma conformação especifica.
Outro aspecto a focar é que o DNA pode ocorrer em cadeias triplas ou quádruplas. Quanto a cadeias
triplas, refere-se que nucleótidos ligados segundo uma ligação normal de Watson e Crick, podem formar
ligações de hidrogénio adicionais, sendo que os átomos que participam nessas ligações adicionais têm o nome
de posições de Hoogsteen. O par adicional é chamado emparelhamento de Hoogsteen. Assim, forma-se um
triplex de DNA. Este tipo de estrutura é mais estáveis a pH baixo, pois envolve muitas vezes a ligação de
resíduos de nucleótidos protonados. Quanto a cadeias quádruplas, ocorrem apenas para moléculas de DNA
com uma percentagem muito elevada de nucleótidos de guanina.
Uma estrutura rara que ocorre no DNA é chamada a H-DNA e é encontrada apenas em longas
sequências polipúricas ou polipirimidicas, causando uma dobra acentuada na estrutura, fazendo com que uma
porção da cadeia exista sob cadeia tripla, em que duas das três cadeias contêm apenas purinas (ou pirimidinas)
e a outra contem apenas pirimidinas (ou purinas).
RNA
A molécula de RNA tem uma muito maior variedade de funções e conformações do que o DNA,
fazendo com que não exista uma estrutura secundária que sirva de base, tal como a dupla hélice era para o
DNA. Dos diferentes tipos de RNA destacamos: RNA mensageiro, que transcreve a informação genética do
DNA e a transporta para o citoplasma; RNA ribossomal, que é parte integrante do esqueleto dos ribossomas,
que são o local de síntese proteica; RNA de transferência, que funciona como descodificador do código
genético, funcionando como ponte entre informação genética e síntese proteica; RNAs especiais tais como as
ribozimas, catalizando reacções.
O RNA ao ser transcrito existe sempre em cadeia simples, adquirindo uma estrutura helicoidal de mão
direita, sempre que não existem sequências auto-complementares ao longo da cadeia polinucleotídica. No
caso de existirem essas sequências, ocorre a formação dos tais ganchos de cabelo falados anteriormente, e no
caso de haver falhas na complementaridade de base, podem ocorrer arcos internos, protuberâncias. É de
referir que as estruturas ganchos de cabelo não são estruturas planares, adquirindo também uma estrutura
helicoidal de mão direita.