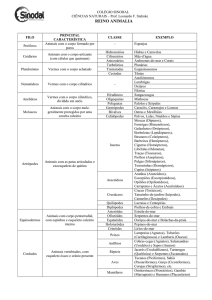
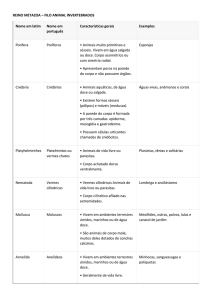
Visualização do documento
Cap+¡tulo 7 - Vermes.doc
(154 KB) Baixar
P á g i n a | 62
Vermes Platelmintes e Asquelmintes – Zoologia Aquática – Engenharia de Aquicultura
Capítulo 5
VERMES PLANOS: OS PLATELMINTES
A palavra “verme” é aplicada livremente a animais invertebrados compridos, bilaterais e sem apêndices. Entre eles
destacam-se os vermes planos (platelmintes) cuja organização corporal refletiu externamente um corpo alongado com
região cefálica distinta acompanhada por um extremo achatamento dorso-ventral. Os platyhelminthes (platy, achatado +
helminthes, vermes) ou vermes planos derivaram de um ancestral que provavelmente teve muitas características
semelhantes aos dos cnidários (aguas-vivas, hidras, anêmonas-do-mar, corais etc.), inclusive mesogléia gelatinosa. A
transformação para a forma vermiforme ocorreu inicialmente pela substituição da mesogléia na larva planular por um
parênquima (tecido de preenchimento) originado a partir da mesoderme, que forneceu a base para uma organização mais
complexa. Esse processo envolvendo os vermes planos explica sua natureza triblástica (organismos com três folhetos
embrionários: ectoderme, mesoderme e endoderme) com o aparecimento de um parênquima (tecido de preenchimento)
entre o tubo digestório e a pele, repleto de novas células e fibras mais especializadas do que aqueles presentes no grupo
das águas-vivas. Nesses organismos começam a aparecer determinados órgãos relacionados a excreção reprodução e ao
sistema nervoso, mas nenhum deles apresenta cavidades específicas, o que caracteriza esses animais como acelomados.
Outro aspecto bastante relevante para a evolução dos vermes foi a estabelecimento de uma simetria bilateral que permitiu
um novo tipo de vida, caracterizado pela exploração do ambiente o que não era possível na vida séssil dos animais radiais.
Para animais que passam suas vidas assentados e em espera, como no caso de alguns animais radiais como algumas espécies
de esponjas e cnidários, a simetria radial é a ideal porque um lado do animal e tão importante quanto o outro para a
percepção da presa que vem de qualquer direção. Mas se um animal é ativo em procurar sua presa, seu abrigo, locais para
viver, parceiros reprodutivos, ele requer um conjunto de estratégias diferentes em uma nova organização corpórea.
Movimentos ativos dirigidos demandam um alongamento da forma do corpo com extremidade cefálica (anterior) e
caudal (posterior). Ademais, um lado do corpo é mantido voltado para cima (dorsal) e o outro lado, especializado na
locomoção, mantido para baixo (ventral). O resultado disso é um animal bilateralmente simétrico no qual o corpo pode
ser dividido ao longo de um plano de simetria, resultando em duas metades iguais que são imagens especulares (espelho)
uma da outra. Essa nova configuração permitiu a acomodação de um grande número de células nervosas na região anterior
do corpo desenvolvendo uma estrutura ganglionar (cérebro primitivo), o que permitiu uma interpretação mais
refinada do ambiente como, por exemplo, formação de imagem, melhorias na recepção de olfato, tato e até mesmo
paladar.
ESTRUTURA CORPORAL E CLASSIFICAÇÃO DOS VERMES PLANOS
Os platelmintos variam em forma e tamanho desde um milímetro ou menos até algumas tênias de muitos metros de
comprimento. Seus corpos achatados podem ser afilados na forma de amplas folhas, ou longos e em forma de fita.
PLATELMINTES DE VIDA LIVRE:
Os platemintos incluem formas de vida livre e parasitárias, mas seus representantes livre são encontrados somente na
Classe Turbellaria (turbilhão). Alguns turbelários são comensais ou parasitas, mas a maioria é adaptada para a vida como
habitante de fundo de água doce ou marinho ou ainda vivem em lugares úmidos no ambiente terrestre. Muitos,
especialmente de espécies maiores são encontrados no lado inferior de rochas e outros objetos duros em fluxos de água
doce ou nas zonas litorâneas do oceano. Variam de formas ovais a alongadas achatadas dorso-ventralmente, podendo em
algumas espécies existirem projeções na cabeça em forma de tentáculos ou laterais chamadas de aurículas (orelha).
Apresentam coloração do corpo variando entre o marrom, preto e cinza. São aquáticos de vida livre e a maioria é marinha,
mas ocorre um grande número de espécies do ambiente de água doce. As planárias e alguns outros turbelários freqüentam
riachos montanhosos ou formados na primavera, bem como poças formadas temporariamente. Algumas espécies mais
resistentes podem ocorrer em gêiseres quentes, ou até mesmo no ambiente terrestre debaixo de rochas e troncos. A
principal característica do grupo dos turbelários é o corpo recoberto por uma epiderme ciliada simples na qual cada célula
porta muita cílios que geram os movimentos de turbilhamento na coluna d’água característico desses vermes planos de
onde deriva o nome Turbellaria. Os cílios podem estar distribuídos por todo o corpo incluindo a superfície dorsal ou pode
ser restrito a uma região corporal como no caso dos Tricládios (três ramos internos) e Policládios (mais de três ramos
internos) cujos cílios se confinam a superfície ventral.
PLATELMINTES PARASITAS:
Todos os membros das outras classes que incluem os Monogenea, Trematoda (fasciolas e xistossomas) e Cestoda (tênias)
são parasitos. Esse conjunto de vermes planos tem uma relativa importância para o seres humanos, pelo fato de muitos dos
espécimes relacionados as classes de Trematoda e Cestoda causarem danos consideráveis a saúde do homem. O parasitismo
é uma forma de associação entre dois animais, na qual existe unilateralidade de benefícios, ou seja, o hospedeiro é espoliado
pelo parasito, pois fornece alimento e abrigo para este. De modo geral essa associação tende para o equilíbrio, pois a morte
do hospedeiro é prejudicial. Assim , nas espécies em que essa associação vem sendo mantida há milhares de anos, raramente
o parasito leva o hospedeiro à morte, dessa forma ocorre uma espoliação constante, mas insuficiente para lesar gravemente
o hospedeiro. Como surgiram os vermes ou demais organismos parasitos? A maioria dos seres vivos possui algum tipo
de inter-relacionamento que varia entre a colaboração mútua, o predatismo e canibalismo. O parasitismo, seguramente
ocorreu quando na evolução de uma dessas associações um organismo menor se sentiu beneficiado, quer pela proteção,
que pela obtenção do alimento. Como conseqüência dessa associação e com o decorrer de milhares de anos houve uma
evolução para o melhor relacionamento com o hospedeiro. Essa evolução, feita as custas de adaptações, tornou o invasor
mais e mais dependente do outro ser vivo, surgindo então os organismos parasitos. Quais foram as adaptações que
explicam o aparecimento de organismos parasitos? Entre as adaptações encontram aquelas relacionadas a morfologia
do parasito, ou seja, muitos organismos parasitas perderam a capacidade de se locomover, e o aparato digestório o que
implica em pegar emprestado essas atividade desenvolvidas pelo hospedeiro. A perda da locomoção torna a vida externa
muito difícil para o espécime a medida que predadores podem ameaçar a vida mais facilmente. Esse processo favorecu mais
ainda a sobrevivência dentro de um hospedeiro, uma fortaleza segura contra predadores e levou a hipertrofia de algumas
estruturas nos parasitos especialmente os órgãos de fixação, resistência e proteção. Assim alguns helmintos possuem
órgãos de fixação muitos fortes como lábios, ventosas, acúleos. Obviamente é de se esperar que o hospedeiro tenha alguma
reação para expulsar o intruso, contudo os parasitos desenvolveram a antiquinase uma enzima que neutraliza a ação de
sucos digestivos, anticorpos e imunossupressão. Outro aspecto relevante foi o aumento da taxa reprodutiva para suplantar
as dificuldades de atingir novo hospedeiro e escaparem da predação externa, acompanhado de um aumento acentuado do
ovário e útero para armazenar os ovos. O próprio hermafroditismo, a partenogênese, a poliembrionia e a esquizogonia
foram métodos desenvolvidos pelos helmintos e outros parasitos que asseguram uma reprodução mais segura.
A Classe Monogenea são vermes planos parasitos externos de vertebrados aquáticos especialmente peixes, que possuem
um corpo dividido em cabeça, tronco e o haptor. O haptor é o principal órgão de ligação posterior, portando ganchos,
acúleos e ventosas permitindo que ele se fixe em um hospedeiro, especialmente nas brânquias e tegumento de animais
aquáticos. A cabeça pode apresentar ventosas, mas isso não é freqüente em todas as espécies, contudo, um grande número
de glândulas adesivas invariavelmente é encontrado. O nome Monogenea que significa “apenas uma geração” deriva do
ciclo de vida desses vermes que não possui hospedeiro intermediário, pois não utilizam um invertebrado para mudar a fase
do estágio larval.
Ao contrário do monogenéticos todos os trematódes e céstodes são parasitas internos (endoparasitas) e seus ciclos sempre
envolve dois hospedeiros, um abriga temporariamente o verme e o outro abriga definitivamente. A Classe Trematoda é
representada por duas sub-classes de vermes planos parasitas, intimamente relacionados que incluem os Digenea
(medicamente e economicamente importantes) como exemplos a Fasciola hepatica e o Schistossoma mansoni (responsável
pela doença esquistossomose) e os Aspidogastreos. Os Digeneos são endoparasitas comuns de todas as classes de
vertebrados (peixes, anfíbios, répteis, aves e mamíferos) podendo causar doenças debilitantes em animais domésticos e
humanos. O ciclo de vida inclui pelo menos dois estágios infectivos de parasitismo, por isso o nome Digenea que significa
duas gerações. O primeiro estágio envolve um hospedeiro intermediário ou temporário que geralmente é um molusco (e
caso haja um segundo sempre é um Artropodo) e o hospedeiro definitivo é sempre um vertebrado. Os vermes planos
digenéticos são via de regra pequenos, mas alguns podem atingir dimensões bem visíveis a olho nu como o caso da fasciolas.
A característica distintiva do grupo é a presença das ventosa oral circundando a boca, e a ventosa ventral que se
assemelha a um acento podendo também receber a denominação de acetábulo ventral. Embora a distância entre as
ventosas possa variar conforme a espécie, invariavelmente estão sempre próximas e exercem primordialmente a função de
ligação com o órgão parasitado, impedindo o desalojamento do verme, e especificamente no caso da ventosa oral auxiliando
também na condução do alimento para a boca. Os aspidogastreos correspondem a um pequeno grupo de trematódes que
parasitam as cavidades intestinais de peixes e répteis e a cavidade pericárdica e renal de moluscos bivalves. A característica
distintiva do grupo é o órgão adesivo, que consiste de uma ventosa septada única na superfície ventral ou de uma fileira
longitudinal de ventosas.
O ultimo grupo de vermes planos é a Classe Cestoda (Cestóides) que inclui endoparasitas conhecidos como solitárias que
possuem adaptações tegumentares especiais associadas ao consumo de nutrientes, pois não possuem trato digestório. O
corpo é formado por cabeça anterior denominada de escólice adaptada para aderir ao hospedeiro provido em quatro lados
de ventosas e ganchos para a ligação com a parede do intestino. Após o escólice encontra-se o colo estreito (semelhante a
um pescoço), que se prolonga dando origem ao corpo chamado de estróbilo. O estróbilo é constituído por seções
segmentares, chamadas de proglótides. O tegumento é importante no transporte ativo de carboidratos e aminoácidos e na
evasão da resposta imune do hospedeiro.
PAREDE CORPORAL
A exceção dos monogenéticos a epiderme dos vermes planos é desprovida de uma cutícula protetora sendo que no grupo
dos turbelários observa-se distribuição de cílios em todo o tegumento ou restritos a região ventral. Em alguns digenéticos
e nas cestódes a camada epidérmica forma um sincício, ou seja, não pode estabelecer limites entre uma célula epidérmica
e outra de tal forma que os núcleos ficam isolados através do citoplasma somente. Essa configuração parece estar
relacionada a permeabilidade em relação a água e a absorção de nutrientes pela pele. Apesar da epiderme não exibir limites
bem definidos observa-se embaixo dela uma lâmina basal, com uma série de fibras intracelulares para sustentar a parede
corporal. O esqueleto intracelular que dá certa consistência ao corpo desses invertebrados é representado por uma teia de
filamentos de actina (proteína estrutural) dentro da epiderme, podendo haver a ocorrência de espículas calcárias nos
marinhos para auxiliar a sustentação. Uma característica importante e a presença de células glandulares denominadas de
rabdomeros penetrando na epiderme e abaixo das camadas musculares. As glândulas mucosas rabdomeras podem
promover adesão e secreção de muco (existem glândulas ciliares, glândulas ventosas, glândula dupla, glândula frontal),
através de pequenos bastões protéicos, os rabditos que quando expelidos entram em contato com a água do meio externo
e se desmancham formando uma camada mucosa protetora sobre a epiderme. A maioria das ordens de turbelários tem
órgãos adesivos duoglandulares na epiderme que incluem três sistemas celulares, as células viscosas e liberadoras e
as células de ancoragem. As secreções das células glandulares viscosas aparentemente firmam os microvilos (formam a
base dos cílios) das células de ancoragem ao substrato, e secreções da glândula liberadora ativam rapidamente o
mecanismo químico de liberação de muco.
Na parede do corpo abaixo da membrana basal dos platelmintos, existem camadas de fibras musculares que se organizam
circularmente, longitudinalmente, diagonalmente e dorso ventralmente. Os músculos circulares se encarregam de
aumentar ou diminuir o diâmetro corporal enquanto os músculos longitudinais auxiliam no alongamento e encurtamento
do verme. A contração alternada desses músculos cria os peristaltismos de movimentos ondulatórios característicos, que
são mais importantes ainda nos turbelários, porque auxiliam a movimentação dos cílios junto a uma sola ventral
especializada. Os músculos dorso ventrais promovem o dobramento ou enrolamento do corpo e os diagonais servem
especialmente para o retorcimento e para uma postura vertical. O peristaltismo pode exercer em algumas espécies um
papel mais importante para a mistura de do conteúdo do intestino do que previamente a locomoção, porque na maioria das
espécies de vermes planos a musculatura intestinal inexiste ou fracamente desenvolvida.
Entre a musculatura corporal e o intestino encontramos um parênquima constituído de células mesênquimais ou
parênquimais com uma matriz extracelular fibrosa. Basicamente ocorrem quatro tipos de células parenquimais: (1)
Células parenquimais fixas que são ramificadas e fazem contato com outras células; (2) Células de reposição
epidérmica que migram do parênquima para a superfície corporal e servem para reposição de células epidérmicas; (3)
Neoblastos que servem para cicatrização e regeneração dos ferimentos; (4) Cromatóforos que serve para produção de
pigmentos escuros e conferem a coloração ao animal.
NUTRIÇÃO E TRATO DIGESTÓRIO
Com exceção dos céstodes que não possuem sistema digestório, dos vermes planos envolve via de regra uma boca, uma
faringe e um intestino, que caracteriza um trato digestório incompleto pela ausência de um ânus. Os turbelários são
principalmente carnívoros e alimentam-se em grande parte de pequenos crustáceos, nematódes, rotíferos e insetos. Elas
podem detectar sua comida a alguma distância por meio de quimiorreceptores distribuídos na margem região cefálica. A
captura é realizada envolvendo a presa com secreções de muco de suas glândulas mucosas e rabdomeros. Uma planária,
por exemplo, apreende sua presa com sua extremidade anterior, enrola seu corpo ao redor da presa, estende sua
probóscide e suga seu alimento em pequenas quantidades. A probóscide é representada por uma faringe eversível que
se protai através de uma boca localizada no meio ventral atingindo os tecidos corporais da presa que são fragmentados pela
ação de enzimas proteolíticas produzidas por glândulas faringianas internas localizadas na ponta da probóscide ou por
ganchos de preensão. Nem todos os turbelários dispõem de probóscide eversível, algumas espécies como o Microstomum
caudatum de água doce dilatam surpreendentemente sua boca em dimensões suficientemente grande para engolfar presas
maiores do que ele mesmo, especialmente minhocas aquáticas a base de sua dieta.
O conteúdo parcialmente digerido é liqüefeito e então bombeado para o interior do intestino por meio de ação peristáltica
(movimento de contração do sistema digestivo). O intestino poder ser um saco cego único bem desenvolvido como no caso
dos Rhabdocoela, Acoela e Macrostomida, ou dividido em ramos como nos caso dos Tricladida (três ramos intestinais) e
Policladidas (vários ramos intestinais). De qualquer forma a digestão extracelular iniciada na região faringiana é finalizada
no intestino onde ocorre a absorção do conteúdo alimentar liquefeito, e onde os fragmentos alimentares resultantes são
engolfados por células fagocitárias, no qual a digestão por parte de endopeptidases continua em um meio ácido. Cerca de 8
a 12h após a ingestão o vacúolo fagocitários se torna alcalino, o que assinala exopeptidases, das lípases e das carboidrases
necessárias para completar a digestão. Durante o curso da digestão intracelular, o vacúolo afunda mais profundamente no
interior da célula fagocitária e finalmente desaparece. Os resíduos alimentares originados a partir do processo de digestão
são eliminados via oral.
Ao contrário dos turbelários a boca dos trematódes e monogeneos se abre em uma faringe muscular não extensível,
localizada na extremidade anterior de seus corpos e são posicionadas adequadamente para alimentação através de uma
ventosa oral. Em posição mais posterior seus esôfagos se abrem em um intestino de fundo cego, o qual é geralmente em
forma de “Y”, mas pode ser altamente ramificados ou sem ramificações dependendo da espécie. Como esses vermes se
alimentam de sangue ou fluídos sanguíneos algumas adaptações interessantes são observadas. A faringe nesse caso exerce
papéis vitais na alimentação, pois funciona como uma bomba de sucção mediante forças dos movimentos peristálticos e
sua glândula produz muco anticoagulante que impedem que o sangue se torne viscoso entupindo o tubo digestório. ...
Arquivo da conta:
evaldofarteski
Outros arquivos desta pasta:
IGSg(1).dll (1361 KB)
libEGL.dll (58 KB)
libGLESv2.dll (507 KB)
IGOptExtension.dll (142 KB)
alchemyext.dll (15 KB)
Outros arquivos desta conta:
Documentos
Galeria
Privada
Relatar se os regulamentos foram violados
Página inicial
Contacta-nos
Ajuda
Opções
Termos e condições
Política de privacidade
Reportar abuso
Copyright © 2012 Minhateca.com.br