Capitulo 6
Fisiologia da Motricidade
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
121
FISIOLOGIA MUSCULAR E SISTEMAS MOTORES
MOVIMENTO
Tendemos a pensar que movimento é um conceito mais associado ao deslocamento do
organismo de um lugar para o outro. Entretanto, os animais de vida séssil também realizam
movimentos isolados de partes do corpo, propelindo alimento ou fluidos através das suas partes
internas (como as esponjas e as ascídias).
Podemos reconhecer os seguintes padrões de movimentos:
a) Movimento amebóide. Corresponde a mudanças de forma da célula envolvendo o fluxo de
citoplasma e a formação de pseudópodes. Quando uma ameba se move, seu citoplasma flui entre os
prolongamentos celulares recém formados que se expandem e se avolumam gradualmente. Novos
pseudópodes são formados na direção do movimento. Durante o processo, o citosqueleto
ectoplasmático (formado de actina, miosina e outras proteínas) é fragmentado aumentando,
localmente, a pressão osmótica. Como conseqüência o endoplasma é arrastado a favor do gradiente
osmótico formando o pseudópode.
b) Movimento ciliar/flagelar: é o modo mais típico de locomoção dos protozoários ciliados ou
flagelados, no entanto, os animais metazoários também possuem células ciliadas nos epitélios que
movimentam mucos ou outros fluidos e células flageladas (espermatozóides).
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
122
Os cílios e flagelos possuem organização
estrutural semelhante, mas a diferença está no padrão
de batimento. O flagelo de espermatozóide p.e, bate
com uma ondulação simétrica ao longo de toda cauda.
Já um cílio bate de modo assimétrico com uma batida
rápida, como uma chicotada, em uma única direção
(linhas cheias), seguida por um movimento mais lento
de recuperação (linhas tracejadas). Por causa desse
O batimento flagelar propulsiona a água paralelamente
ao eixo principal do flagelo e o movimento ciliar
propulsiona a água paralelamente à superfície de
inserção.
movimento
o
fluido
em
volta
é
deslocado
paralelamente ao eixo do flagelo (seta); em relação ao
movimento ciliar, o fluido é movimentado perpendicularmete ao batimento ciliar (seta). Os
mecanismos causais dos movimentos ciliares são especulativos, mas a excitabilidade da membrana
celular e o fluxo de correntes de Ca++ são essenciais para determinar o sentido do batimento ciliar e
como conseqüência, a direção de movimento paramécio.
c) Movimento muscular: os animais maiores lançam mão de células especializadas, os miócitos
que possuem proteínas contráteis e, quando em atividade, geram tensão mecânica. Conforme o
arranjo dessas células os animais metazoários movimentam-se, estabilizam articulações ósseas, ou
então, propelem os fluidos corporais dentro de órgãos tubulares (intestino ou vasos). Conforme o
controle sobre a musculatura esquelética é possível a mais sutil expressão motora, atos complexos
como escrever, dançar, praticar esportes ao mais simples movimento como o ato de piscar. O
repertório motor de uma espécie depende do número de grupos musculares e dos mecanismos de
controle da motricidade. Todas as fibras musculares, sem exceção, são eletricamente excitáveis e
gastam energia metabólica para gerarem a tensão mecânica.
Os tipos de fibras musculares
No interior das fibras musculares, tanto de vertebrados como de invertebrados, há proteínas
que promovem a geração de tensão mecânica, ou seja, que promovem a contração. Baseado no
modo como as proteínas contráteis estão dispostas, as fibras musculares podem ser classificadas
conforme o padrão de estrias observadas ao microscópio óptico. Nos vertebrados reconhecemos
três tipos de fibras musculares: fibras musculares esqueléticas, cardíacas e lisas. Como será visto
mais tarde, as fibras musculares lisas e cardíacas (viscerais) estão sob controle distinto das fibras
musculares esqueléticas (somáticas) e possuem propriedades mecânicas peculiares.
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
123
Tamanho corpóreo e o método de propulsão
Organismos pequenos utilizam cílios (paramécio) e flagelos (coanoflagelado) para
realizarem movimentos natatórios e os maiores, fibras musculares (metazoários) que possuem
elementos contráteis. O método de propulsão depende de dois fatores:
1) Geometria do corpo. Organismos menores possuem a relação superfície/volume maior em
relação aos de porte maior. O movimento ciliar ou flagelar é suficiente para aproveitar a
viscosidade do meio e propelir o animal para frente contra a camada de água circundante. Mas a
relação superfície/volume de uma hipotética baleia “ciliada” seria tão pequena que o empuxo
necessário é insuficiente constituindo uma enorme desvantagem mecânica. Assim, o
mecanismo de propulsão alternativo para deslocar corpos maiores foi a contração muscular, ou
seja, um tecido especializado gerando força mecânica sobre o substrato e alavancando o corpo.
2) Leis sobre a dinâmica dos fluidos. O meio liquido foi o ambiente primitivo onde as estratégias
de locomoção evoluíram. Nesse meio, a viscosidade determina a velocidade de deslocamento:
imagine-se nadando em uma piscina de xarope...e depois de água. A viscosidade de ambos é
diferente, portanto, oferecem resistências diferentes. Uma grande vantagem mecânica e
energética seria a redução das forças de atrito. A seleção natural favoreceu formas
hidrodinâmicas dos corpos, mucos, nadadeiras e músculos agindo sobre esqueleto. O que os
atletas de natação têm aprendido com a hidrodinâmica dos peixes?
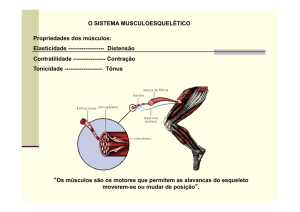
Músculo esquelético de vertebrado
O mesoderma origina todas as fibras musculares do corpo, sejam viscerais (lisas e
cardíacas) ou somáticas (esqueléticas). Nos vertebrados, a musculatura esquelética corresponde
aproximadamente a 40% do peso corporal e os demais tipos de tecido muscular a 10%. De um
modo geral, estão associados a ossos, mas fazem exceção os músculos que formam os esfíncteres
anais e vesicais.
Toda contração muscular envolvendo uma articulação móvel realizará movimento. O
movimento coordenado de vários músculos para a locomoção (deambulação, corridas, saltos, vôo,
etc), a exibição de comportamento sexual, lutas, escrever, bem como a manifestação de reflexos
posturais automáticos são orquestrados pelo SNC. Os comandos produzidos no SNC são enviados
para os músculos por meio de neurônios motores que determinam o tempo e a força de contração
das unidades musculares controladas.
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
124
Os músculos podem se prender diretamente a ossos por meio de tecido conjuntivo que os
envolve e os permeia. Mas também podem se prender por meio de tendões, cordões bastante
resistentes de ancoragem (os ligamentos ligam osso com osso). Alguns músculos não se prendem a
ossos: ao invés disso, distribuem a força sobre uma ampla superfície de lâminas de tecido
conjuntivo resistente chamada de aponeurose. O tecido conjuntivo frouxo que prende um músculo
ao outro e a pele ao músculo é denominada fáscia. Quando um músculo contrai, ele puxa as suas
extremidades com igual intensidade. A inserção do músculo corresponde à extremidade que se
encontra relativamente livre para mover-se e a origem, a parte mais fixa. Baseado no tipo de
movimento que resulta da contração, os músculos podem ser classificados em músculos flexores
(ao contrair, diminuem o ângulo articular) ou extensores (aumentam o ângulo). Se basearmos na
função os músculos são classificados em:
- Músculos adutores (aproximam) e abdutores (afastam) partes do corpo ou o eixo do apêndice
em direção ao plano sagital do corpo.
- Músculos elevadores (elevam) e os depressores (abaixam) partes do corpo como a mandíbula ou
o ombro.
- Músculos protadores (empurram) e retratores (puxam de volta) parte do corpo como a língua
ou apêndice. Os rotadores giram partes sobre um eixo longo (como a coluna ou apêndices).
- Músculos constritores: anéis de fibras musculares que circundam aberturas (esfíncter) como
anus, uretra ao se contraírem, reduzem o diâmetro da luz ou causam oclusão total.
- Músculos supinadores: giram a palma da mão ou a planta pé para cima e os pronadores giram
para baixo.
Durante a contração de um músculo, existe uma ação oposta ou de restauração promovida
pelos músculos antagonistas e, raramente, um músculo contrai sozinho; são suplementados por
outros músculos chamados de sinergistas (ou agonistas).
Outra classificação que auxilia muito quando estamos analisando o papel do SNC sobre o
controle da motricidade somática é a diferenciação entre músculos medial-lateral e proximaldistal. Os músculos axiais, da cintura pélvica e escapular, sejam extensores ou flexores, são
considerados mediais. Estes músculos estão associados com os movimentos corporais de todo o
membro ou da sua postura. Já os músculos distais dos membros e dos movimentos intrínsecos dos
dedos, não participam primariamente da postura, mas de atividades relacionadas com a
manipulação de objetos como no caso de primatas.
Fontes de energia da contração muscular esquelética
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
125
O suprimento energético imediato para a contração muscular é o ATP (adenosina
trifosfato). Quando hidrolisado, parte da energia liberada é utilizada para a geração de força
mecânica e o restante é perdido na forma de calor. Como a disponibilidade de ATP é pequena nas
fibras musculares, é necessária uma fonte adicional de energia como o fosfato de creatina que
transfere o grupo fosfórico para o difosfato de adenosina, restaurando rapidamente os níveis de
ATP. Mas é fonte muito limitada para um exercício muscular prolongado e o ATP precisa ser
obtido pela oxidação de carboidratos (glicose) e de ácidos graxos (gordura). A fonte de carboidrato
muscular está armazenada na forma de polímeros de glicose conhecidos como glicogênio (cerca de
0,5 a 2% do peso muscular fresco) e contribui com cerca de 100 vezes mais do que a fonte de
fosfato de creatina. Conforme as propriedades mecânicas das fibras musculares serão utilizadas ou
no exercício aeróbico ou anaeróbico.
Níveis de organização morfológica do músculo esquelético
Fibra muscular
O músculo esquelético é constituído de feixes de fibras musculares (fascículos) cujas
células possuem diâmetro que variam de 10 a 80 m e comprimentos de algumas a dezenas de mm.
Cada fibra muscular é envolvida por uma membrana denominada sarcolema, (nome dado à
membrana citoplasmática da fibra muscular) e por uma camada fina de polissacarídeos e fibrilas de
colágeno. As fibras musculares afunilam-se em suas extremidades fundindo-se a elementos fibrosos
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
126
e tendinosos que se ligam aos ossos. Entre os feixes de fibras musculares está o suprimento
sangüíneo, a inervação e o tecidos adiposo.
Estudando o interior da fibra muscular encontramos pacotes de miofibrilas (feixes
cilíndricos de proteínas) dispostos paralelamente, apresentando padrões repetitivos de estrias
transversais. Cada unidade repetitiva é denominada sarcômero que é a unidade contrátil da fibra
muscular. Além dos demais elementos subcelulares típicos de uma célula eucariótica, chama a
atenção o retículo sarcoplasmático que armazena íons Ca++ e possui canais de Ca voltagemdependentes. O retículo sarcoplasmático (RS) é uma estrutura volumosa (14% da fibra), formado de
túbulos que se anastomosam e acompanham as miofibrilas; possui extremidades que termina em
sacos (cisternas terminais). Neste ponto, são trespassados pelos túbulos transversais (túbulos T) que
se originam do sarcolema formando uma estrutura tríade. Os túbulos T dispõem-se
perpendicularmente ao RS e às miofibrilas, formando uma rede transversal e reticulada no interior
da fibra muscular. Funcionalmente comunicam o meio extracelular transversalmente ao longo da
fibra muscular. Esse arcabouço morfológico é fundamental para a condução da atividade elétrica na
fibra muscular e acionar a liberação de Ca++ de dentro do RS para o citoplasma.
Organização molecular do sarcômero
O sarcômero constitui a unidade contrátil da fibra muscular e está limitada pelas linhas Z.
No músculo de todos os vertebrados, o comprimento do sarcômero é semelhante sendo em torno de
3,5m no estado de repouso e de 1,5m quando contraído. Cada sarcômero apresenta uma
organização molecular constituída de filamentos finos e filamentos grossos. A partir da linha Z, os
filamentos finos estendem-se paralelamente para ambos os sentidos sendo intercalados pelos
filamentos grossos, apenas na região central do sarcômero. Tal organização produz um padrão de
bandas transversais que varia conforme o estado de contração muscular. Os filamentos grossos são
constituídos de macromoléculas protéicas de miosina e os filamentos finos, de actina,
tropomiosina e troponina.
A miosina corresponde a 55% da proteína miofibrilar e cada filamento grosso é formado
por 200 a 400 dessas moléculas; Cada molécula de miosina é formada por duas cadeias: uma leve e
outra pesada. Na cadeia pesada a cabeça possui dois sítios ativos: uma que possui alta afinidade
pela actina e uma outra catalítica que hidrolisa o ATP. As moléculas de miosina formam um feixe
em que as cabeças se destacam do eixo central e dispõem-se bem próximos aos filamentos de
actina. Apesar da elevada afinidade das moléculas de miosina para com a actina, quando o músculo
está em repouso (relaxado), a ligação entre elas está obstruída pelas moléculas de tropomiosina.
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
127
A actina constitui 20 a 25% da proteína miofibrilar e ocorre em duas formas (actina G e
actina F) que, quando polimerizadas, formam cordões duplos helicoidais. Cada molécula de actina
G liga-se com grande afinidade a um íon Ca++ e a uma molécula de ATP.
A tropomiosina é uma molécula formada por duas cadeias peptídicas separadas e que estão
enroladas entre si.
A troponina é uma proteína globular com função reguladora e possui três subunidades: C
que se liga a íons Ca++; I que é inibitória e T que se liga à tropomiosina. No músculo relaxado a
tropomiosina está obstruindo o sítio de ligação da actina com a miosina.
Como ocorre a contração muscular esquelética?
Veja http://www.blackwellscience.com/matthews/myosin.html
Quando o músculo não está sob estimulação nervosa, isto é, quando está relaxado, a [Ca++]
do mioplasma é insignificante em relação ao meio extracelular. Com a chegada dos impulsos
nervosos a placa motora responde com potenciais de placa e dispara PA no sarcolema. Graças aos
túbulos T a atividade elétrica propaga-se transversalmente e atinge as cisternas do RS abrindo os
canais de Ca++ voltagem dependentes. O cálcio armazenado no interior do RS flui passivamente
para o mioplasma a favor do seu elevado gradiente de concentração.
Os íons Ca do mioplasma ligam-se à troponina C, modificando a sua organização espacial
e resultando no deslocamento da tropomiosina que libera o sitio de ligação da actina com a
miosina. Agora com o sitio livre a miosina liga-se à actina e forma-se a ponte cruzada entre o
filamento fino e o grosso. Como a miosina tem um sítio catalítico para a hidrólise de ATP, esta
união torna a energia química disponível para o dobramento da cabeça de miosina. O dobramento
faz com que os filamentos finos sofram um ciclo de arraste em direção ao centro, isto é, os
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
128
filamentos finos deslizam-se sobre os grossos e cada sarcômero encurta-se e a fibra muscular
como um todo contrai. Com a hidrólise do ATP, a molécula de miosina perde a afinidade pela
actina, desliga-se e reassume a sua posição original. Mas se continua a existir ATP e Ca ++ no
mioplasma, o ciclo das pontes cruzadas é restabelecido novamente e o deslizamento progride,
encurtando o sarcômero a cada ciclo. Cada ciclo move o filamento fino cerca de uns 10nm e para
cada molécula de troponina ativa, sete sítios fixadores de miosina são descobertos.
Denominamos de acoplamento excitação-contração (ou acoplamento eletro-mecânico),
o processo em que a atividade elétrica da fibra muscular (potenciais de ação) é transformada em
atividade mecânica (encurtamento do sarcômero), ou seja, a transdução de atividade elétrica em
mecânica.
Relaxamento muscular
Se impulso nervoso cessar, não haverá mais PA no sarcolema e a contração será encerrada:
os canais de Ca++ se fecham e o cátion será recaptado ativamente de volta para dentro das cisternas
do RS por ATPases Ca/Mg dependentes. Como a afinidade dessa bomba pelo Ca++ é maior do que
do íon pela troponina, o recolhimento ativo predomina. Com a cessação dos PA no sarcoplasma, o
músculo encontrar-se-á no estado de repouso.
Rigor mortis
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
129
Com a morte o sistema de transporte ativo entra em falência e, mesmo na ausência de
impulsos nervosos, o Ca++ difundirá passivamente para o mioplasma. Como conseqüência, inicia-se
a formação de pontes cruzadas sem relaxamento resultando na contração muscular generaliza-se até
o estado de que denominamos de rigor mortis. O grau de rigidez dependerá da reserva de
glicogênio do músculo disponível e da temperatura.
Unidade motora
Agora já sabemos como a fibra muscular contrai e que para isso depende de controle
nervoso. Mas como a força muscular é gerada e controlada?
Denominamos de unidade motora o motoneurônio e as fibras musculares por ele
inervadas. A fibra muscular esquelética dos vertebrados só recebe a inervação de um único
motoneurônio. Num mesmo individuo, a inervação da musculatura varia: há unidades motoras cuja
relação de inervação é baixa, (o motoneurônio inerva 1.700 fibras musculares) ou alta (o
motoneurônio inerva algumas dezenas de fibras musculares). Os músculos de grande precisão
motora como os músculos oculares e dos dedos possuem uma relação de inervação de
1motoneuronio para 10 fibras.
MECÂNICA DA CONTRAÇÃO MUSCULAR
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
130
O sistema contrátil depende da concentração de Ca++ disponível no mioplasma e, como
conseqüência, da freqüência com que as fibras musculares da unidade motora estão sendo
estimuladas. Quando o motoneurônio de uma unidade motora dispara um PA haverá uma resposta
mecânica correspondente denominada abalo (espasmo muscular) que é uma breve contração
seguida de relaxamento. A resposta mecânica manifesta-se após um breve período de latência, uns
2ms após o inicio da despolarização e a sua duração varia com tipo de fibra muscular (se são de
contração rápida ou lenta).
O abalo mecânico é uma resposta do tipo tudo-ou-nada e, se um segundo estimulo for
aplicado antes do relaxamento, haverá uma somação ou superposição das respostas mecânicas
resultando em tensões progressivamente maiores e observamos o “fenômeno da escada”. A
aplicação de estímulos cada vez mais freqüente produzirá, finalmente, contrações sustentadas e
uniformes denominadas contrações tetânicas. A força desenvolvida num tétano perfeito é de 4
vezes maior em relação a um abalo isolado.
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
131
Um músculo é formado de várias unidades motoras. Assim, o desempenho mecânico do
músculo é aumentado recrutando-se a atividade de todas as unidades motoras. É por causa dessa
organização morfo-funcional que os vertebrados podem graduar precisamente a força muscular. Se
todas as fibras musculares de um músculo fossem inervadas pôr um único motoneurônio, esse
músculo contrairia sempre como um todo. O recrutamento é realizado em uma determinada ordem
onde as unidades motoras mais fracas (em termos de potencia mecânica) são recrutadas antes e, só
depois, as mais potentes.
Contrações isométricas e isotônicas
A contração muscular é o estado de atividade mecânica. O músculo comporta-se como um
sistema de dois componentes onde a parte contrátil (sarcômeros) está em série com o componente
elástico (tendões e tecido conjuntivo).
Quando os sarcômeros encurtam-se a parte
elástica sofre tração e se uma das
extremidades do músculo for móvel,
causará movimento e se ambas estão fixas,
causará apenas tensão (ou estresse). Nas
contrações isométricas a força está sendo
exercida nos pontos de fixação muscular
sem o encurtamento significativo das fibras
Contração isotônica
Contração isométrica
musculares. Nesse tipo de contração, as
pontes
cruzadas
se
estabelecem
aumentando a tensão ou a rigidez do músculo (na palpação você sentirá o músculo em atividade
duro). O desenvolvimento deste tipo de força ocorre tipicamente nos músculos posturais ou quando
um músculo atua contra uma força oposta. Apesar de não ocorrer trabalho externo, o músculo
realizou o que chamamos de trabalho interno.
Quando as fibras musculares encurtam-se e uma carga externa é movida de um lugar para
outro temos a realização de trabalho externo e dizemos que houve contração isotônica. No
exemplo acima, quando o bíceps ergue a carga e realiza movimento “trabalha” isotonicamente, mas
quando se tenta puxar o cordão que está fixo, não há movimento, mas o músculo está “trabalhando”
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
132
isometricamente. Na prática, uma contração muscular passa de isométrica a isotônica e vice-eversa, o tempo todo.
Relação entre a força máxima e o comprimento do músculo
A força máxima desenvolvida
no tétano depende do comprimento
inicial do músculo. O músculo possui
elementos elásticos em série e em
paralelo (todos os elementos situados
entre
as
pontes
cruzadas
e
as
extremidades dos tendões). Quando um
músculo é passivamente estirado ele
tende a retornar ao seu comprimento de
repouso.
Trata-se
da
propriedade
elástica do músculo que pode ser
descrita
numa
curva
de
tensão
passiva. Quando um músculo executa
uma contração isométrica, a tensão ativa gerada pelas pontes cruzadas é adicionada à tensão
passiva. Na figura, a curva em tracejado é a curva de tensão ativa em função do comprimento e
mostra que a faixa operacional do comprimento do músculo se encontra dentro de uma pequena
porcentagem do comprimento em repouso. A tensão ativa é máxima quando a contração começa do
comprimento em repouso; neste ponto a superposição dos filamentos finos e grossos é tal que todas
as cabeças miosínicas podem formar pontes cruzadas. Se houver uma distensão passiva, o grau de
superposição entre os filamentos diminui e o número de pontes cruzadas também, reduzindo
drasticamente a geração de força ativa. O mesmo verifica-se quando o músculo tenta se contrair a
partir de um grau de sobreposição já existente entre os filamentos. Em suma, os músculos
esqueléticos estão adaptados para gerarem a sua força máxima a partir do comprimento de repouso
e dentro de uma faixa muito estreita em torno desse comprimento. Essa faixa de desempenho ótimo
é denominada faixa operacional real do músculo.
Sistema de alavancas
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
133
As alavancas são constituídas de três partes: o fulcro
(ou pivô), a carga (resistência), o esforço e alavanca em si.
Suponha que você queira levantar uma carga usando uma
alavanca, com o pivô situado bem no meio. Nesse caso você se
esforça, mas o máximo que conseguirá é equilibrar a carga na
outra extremidade. Afastando-se o fulcro do ponto onde o
esforço será realizado, o objeto poderá ser levantado com um
esforço bem menor (ainda que tenha que fazer um percurso
maior para baixo). No cotidiano, várias ferramentas são
utilizadas usando esse principio. Não é diferente para a
realização de movimento de partes ou de todo nosso corpo:
como vimos o custo da contração muscular é grande (consumo
de ATP) e é uma grande vantagem, o uso de alavancas para a
economia de energia mecânica.
Quando um músculo isolado é submetido a contrair suportando uma carga de 7 Kg, ele
gerará uma força equivalente e “puxará” o objeto com um deslocamento de 1cm (que corresponde
ao encurtamento muscular). Mas, a associação do músculo com o esqueleto produz um sistema de
alavanca que amplifica a força gerada pela contração muscular. O bíceps tem a sua inserção além
da articulação móvel do cotovelo. Assim quando ele se encurtar, moverá o antebraço, fletindo-o.
Suponha que a mesma carga de 7Kg seja movida pelo antebraço. Nesse caso, com menos
força a mesma carga poderá ser movida facilmente.
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
134
Exercícios Físicos: Energia e Potencia Mecânica
A gravidade é uma força inexorável. Os peixes mantem o corpo em equilíbrio postural,
contraindo ritmicamente a musculatura que move as nadadeiras, pois do contrário, cairiam para o
lado. Além de estabilizar o corpo contra a ação da gravidade e da correnteza, a musculatura axial e
apendicular do peixe deslocam o seu corpo para frente através de movimentos natatórios. Com base
nas características temporais os movimentos natatórios dos peixes podem ser classificados em:
1) Natação de cruzeiro (sustentado ou estável): movimentos repetitivos e cíclicos com pequena
amplitude de curvatura do corpo. Esse tipo de natação é usada quando longas distâncias são
percorridas ou para a natação de velocidade constante.
2) Movimentos em surto (ou instável): inicio rápido com ampla curvatura do corpo
correspondente a manobras de evasão ou de captura de presa.
Asafa Powel (Jamaica)
Carlos Lopes (Portugal)
Se analisarmos a musculatura axial do peixe, reconhecemos dois tipos de fibras:
“vermelhas” e “brancas". Na musculatura de uma espécie filogeneticamente distante, como o ser
humano, também encontramos padrão de organização semelhante. Que significado biológico teria
tais variações em relação aos tipos de fibras musculares?
Como os peixes, outros animais também realizam exercícios musculares semelhantes.
Várias espécies de aves e de mamíferos herbívoros fazem longas jornadas migratórias exigindo
atividades musculares continuas, ou seja, devem praticar exercícios de resistência. Por outro lado,
os animais precisam realizar ações musculares que exigem muita rapidez e potencia mecânica para
uma caçada ou manobras evasivas anti-predatórias. Será que os tipos de fibras “vermelhas e
brancas” teriam alguma relação?
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
135
Analisemos duas modalidades de atletismo: a figura acima mostra o campeão olímpico da
prova de 100m (velocidade) e da maratona de 42 km (resistência). É evidente que a primeira é uma
prova de curtíssimo prazo (recorde: 9,77s) e a outra de longo prazo (recorde: 2h:09min). Em ambos
os casos, os atletas usam os mesmos grupos de músculos para a corrida. Mas será que exatamente
do mesmo modo? Note que o velocista aparenta maior massa muscular (parece ser mais forte) e o
fundista, franzino. Se prestar bem a atenção durante os dois tipos de provas, o velocista nem respira,
mas o maratonista mantém uma respiração rítmica e uniforme. A corrida de 100m exige velocidade,
e, portanto, grande potência mecânica (força), para otimizar já na saída o atleta assume uma postura
que a partir dele consegue um lançamento eficaz do corpo.
Examinando a musculatura da perna humana verificamos que há basicamente dois tipos de
fibras, como no peixe: fibras vermelhas e brancas. Esse padrão está associado às propriedades
metabólicas e mecânicas das fibras musculares esqueléticas que são classificadas em: fibras do tipo
I: relacionados à resistência e fibras do tipo II: à força e velocidade. As fibras do tipo II podem ser
subdividas em A (metabolismo anaeróbico) e B (aeróbico).
PROPRIEDADES
Cor
Suprimento sanguíneo
No. de mitocôndrias
Grânulos de Glicogênio
Quantidade de mioglobina
Metabolismo
Velocidade de contração
Tempo de contração
Força contrátil
Resistência a fadiga
Tipo I
(resistência )
Vermelha
Rico
Alto
Raros
Alta
Aeróbico
Lenta
Longo
Baixa
Grande
Tipo IIA
(Força e velocidade)
Branca
Pobre
Baixo
Numerosos
Baixa
Anaeróbico
Rápida
Curto
Alta
Grande
Tipo IIB
(Força e velocidade)
Intermediaria
Intermediario
Intermediário
Intermediários
Media
Aeróbico
Rápida
Intermediário
Média
Pequena
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
136
A tabela compara as propriedades dos três tipos de fibras e nos permite concluir que os
músculos posturais que estão o tempo todo em atividade (atividade tônica), como as que estabilizam
as articulações (como os músculos anti-gravitacionais) são fibras musculares aeróbicas resistentes à
fadiga. Como exemplo podemos citar a musculatura do atum que possui uma massa muscular
vermelho-escura (por causa da grande concentração de mioglobinas), situado lateralmente e se
entende à coluna vertebral e são recrutadas durante as jornadas constantes em velocidade de
cruzeiro. Quando atividades fásicas (contração e relaxamento) que exigem força e velocidade são
necessárias, ainda que por pouco tempo, entram em ação as fibras do tipo IIA, cujo metabolismo é
anaeróbico. O atum possui também uma grande massa muscular de fibras brancas que são
recrutadas quando a velocidade é exigida. O músculo sartório da rã extende-se ao longo da sua
coxa é responsável pelo salto e é constituída inteiramente de fibras fásicas, inervadas por
motoneurônios cujas fibras são de grosso calibre. Quando esse músculo é recrutado as suas fibras
musculares respondem manifestando respostas mecânicas máximas.
Mas também há fibras musculares (IIB) que realizam atividades intermediárias: não exigem
grande desempenho mecânico, são mais resistentes à fadiga e apresentam metabolismo aeróbico.
Assim os peixes, aves e mamíferos que migram longas distâncias estão preparados para tais
exigências e, ao mesmo tempo, se durante a navegação de cruzeiro forem atacados por um predador
podem recrutar fibras musculares anaeróbicas e realizar manobras de evasão que são ações rápidas
de força e velocidade.
Esse conhecimento é aplicado no treinamento de atletas. O treinamento do atleta fundista e
do velocista devem ser específicos, ou seja, o primeiro deverá recrutar predominantemente fibras
aeróbicas em seus exercícios enquanto o segundo, exercícios anaeróbicos. É por essa razão que os
velocistas não respiram durante a corrida, mas ficam com débito de oxigênio no final da corrida.
Músculos Sônicos
Alguns animais produzem sons e esses sinais acústicos proporcionam comunicação a
distância entre indivíduos da mesma espécie. Quando uma coluna de ar vibra ou quando se
movimenta ar por uma membrana vibrátil (cordas vocais) é possível gerar sons. Um peixe-sapo
macho emite sons monotônicos vibrando a coluna de ar que fica dentro da bexiga natatória e a
cascavel, chacoalha os guizos caudais para advertir seus inimigos naturais. Para que se produza som
os músculos “sônicos” devem realizar ciclos de contração e relaxamento extremamente rápidos, na
mesma freqüência do som produzido.
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
137
Fadiga muscular
A fadiga é uma condição temporária em que a força muscular não pode mais ser gerada
pelo músculo por problemas metabólicos.
PECULIARIDADES DA MUSCULATURA DOS INVERTEBRADOS
Os invertebrados possuem músculos estriados que geram tensão mecânica sobre um
endosqueleto hidrostático ou exosqueleto quitinoso. Possuem músculos de contração rápida e lenta
como os vertebrados, mas há várias diferenças em relação ao sistema de controle da motricidade.
Para os predadores que caçam presas vivas a rapidez é essencial. As lulas, por exemplo,
capturam suas presas eficazmente graças às fibras de contração excepcionalmente rápidas dos seus
tentáculos. O padrão de estrias das fibras musculares dos tentáculos não é transversal como nas
fibras dos braços, que são oblíquas. Os moluscos bivalves fecham as suas conchas através de
ligamentos e músculos adutores que podem ser um ou dois, conforme a espécie. Esses músculos
lisos quando fecham as conchas podem mantê-lo tonicamente contraído por horas a dias.
Músculos assincrônicos do Vôo (músculos fibrilares)
As asas mais primitivas dos insetos são
as da libélula que são mantidas sempre
esticadas. O surgimento de escleritos foi uma
inovação que possibilitou o dobramento das
asas sobre o abdômen quando em repouso
(como fazem as moscas). A asa sobe e desce e
funciona como se fosse uma gangorra com o
fulcro fora de centro. Mas, não basta as asas
subirem e descerem: é necessário movimentos
para frente e para trás e, assim, num ciclo
completo de batimento observamos a descrição
de uma elipse ou de um oito conforme a
espécie.
A libélula possui músculos para o vôo chamados músculos sincrônicos, pois as fibras
musculares contraem uniformemente em resposta ao estimulo nervoso. O mecanismo básico
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
138
consiste na contração alternada de dois pares de músculos dorso-ventrais. Por meio de alavancas
mecânicas, os músculos elevadores sobem as asas e os depressores, abaixam-na. Uma outra
alternativa para o vôo é o praticado pela maioria de himenópteros (abelhas e vespas), dípteros
(mosca), coleópteros (besouros) e hemípteros (percevejos) que usam a musculatura assincrônica.
A freqüência de batimento é tão alta (de 100 a mais de 1000 batimento por minuto) que devem
possuir mecanismos alternativos para alcançar tal velocidade: de fato, há uma disposição diferente
dos músculos torácicos que estão acoplados a parede interna do tórax. Quando os músculos dorsoventrais contraem, esses puxam o teto do tórax para baixo e a asa se eleva. Ao mesmo tempo
quando o tórax é deformado, isso causa o estiramento da musculatura antero-posterior provocando
uma contração reflexa. Essa contração deforma o tórax para cima, abaixando as asas e servido
estímulo para a contração reflexa da musculatura dorso-ventral, e assim por diante. O sistema
funciona como se fosse um oscilador mecânico: um único impulso nervoso (inicio do vôo) começa
a oscilação mecânica repetitiva e depois, outros impulsos nervosos são transmitidos a intervalos
regulares para a manutenção do batimento alar.
COMPARAÇÃO ENTRE AS FIBRAS MUSCULARES ESQUELÉTICAS, LISAS E
CARDÍACAS DE VERTEBRADOS
Fibras musculares cardíacas
O coração bate ritmicamente de modo que durante a sístole há contração e durante a
diástole, relaxamento. As fibras musculares cardíacas são miogênicas e entre elas há
“células marcapasso” que assumem uma freqüência natural de descargas. As células
marcapasso despolarizam-se e repolarizam-se espontaneamente (não precisam de estímulos
nervosos) e propagam a atividade elétrica para as células vizinhas com as quais estão
acopladas eletricamente. De fato, as fibras musculares cardíacas aderem-se umas as outras
por meio de intedigitações (discos intercalados) onde estão os desmosssomos e as junções
abertas. Possuem canais iônicos que se abrem e de fecham espontaneamente, gerando PA
do tipo tudo-ou-nada, porém com uma configuração diferente das fibras esqueléticas: a
despolarização é bem rápida, mas uma repolarização é tão lenta que forma até um platô.
Com uma duração maior do PA mais Ca fica disponível para o ciclo das pontes cruzadas, e
os abalos mecânicos são considerados fortes.
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
139
Os túbulos T são curtos e não há formação de tríades. A resposta mecânica
praticamente coincide com a duração do PA e como conseqüência é impossível ocorrer
somações mecânicas, ou seja, uma contração tetânica. Para se aumentar o volume de
sangue ejetado na unidade de tempo, a alternativa é a de aumentar a freqüência de
batimento do coração.
Apesar das propriedades miogênicas, o sistema nervoso visceral regula a
ritmicidade dessas células marcapasso, acelerando ou retardando o ritmo cardíaco,
conforme as exigências do organismo.
Fibras musculares lisas
As fibras musculares lisas são pequenas e fusiformes; possuem actina e miosina, mas não
apresentam padrões de estriações transversais. Não possuem moléculas de troponina e os filamentos
ficam ancorados nos corpos densos. As células musculares lisas podem contrair até 80% de seu
comprimento – em contraste aos menos de 30% do músculo esquelético. Há dois tipos de fibras
musculares lisas:
a)
Fibras unitárias (ou viscerais): funcionam como sincício funcional e estão eletricamente
acopladas entre si e possuem propriedades miogênicas.
b)
Fibras multiunitárias: parecem-se as fibras musculares esqueléticas e dependem de controle
nervoso.
Nas fibras unitárias, a atividade elétrica gerada numa célula propaga-se para as outras como
se fosse uma onda. Durante a fase de despolarização há abertura de canais de Ca e a sua entrada
promove o deslizamento dos miofilamentos. Como exemplo, citaremos a motilidade intestinal. A
parte muscular do trato gastrointestinal de vertebrados é composta por fibras musculares lisas
circulares e longitudinais. A contração dessas fibras promove a propulsão ou a mistura do seu
conteúdo. O peristaltismo intestinal resulta da combinação de dois mecanismos distintos:
a) controle instrísenco ou miogênico: As fibras musculares geram espontaneamente ondas de
despolarização e repolarização produzindo um ritmo ondas lentas (REB). O Ca que flui para
dentro da célula liga-se a calmodulina (a musculatura lisa não possui troponina) e promove a
contração. Note-se que a tensão mecânica pode ser criada sem a geração de PA no sarcolema. A
presença das ondas REB garante ao intestino o seu tônus de repouso.
b) Controle extrínseco (neural e humoral): por outro lado os REBs podem ser modulados por
mediadores químicos (hormônios, NTs do SNA e outros mediadores do próprio plexo entérico).
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
140
Outra característica da pequena fibra muscular lisa é o fato de dispensarem os túbulos T e
possuir um RS bastante discreto. A fonte de Ca necessária para a contração vem principalmente
de fora.
As fibras musculares multiunitárias podem ser exemplificadas pelas fibras do músculo da
íris, da parede das artérias, e aquelas que eriçam os pelos. Essas fibras não possuem acoplamento
elétrico e dependem totalmente do controle nervoso para contrair. O sarcolema responde
eletricamente ao estimulo neural sem gerar PA. A despolarização abre os canais de Ca necessários
para ativar os mecanismos moleculares da contração.
Propriedades/
Componentes
Bandas Visíveis
Filamentos Grossos e
Finos
Tropomiosina e troponina
Túbulos transversais
Reticulo Sarcoplasmático
Mecanismo de contração
Inervação
Inicio da Contração
Fonte de Cálcio p/
ativação
Junções abertas
Velocidade de contração
Relação nítida entre
tensão e comprimento
Músculo Estriado
Músculo Estriado
Músculo Liso
Músculos
associados
a
esqueletos e são responsáveis
pela postura e movimento do
corpo
Musculatura que formam o
coração cujo ciclo de contração e
relaxamento propulsionam o
sangue através dos vasos.
Musculatura que formam os demais órgãos
cavitários do corpo (vasos sanguíneos, bexiga,
trato gastrointestinal, útero, etc.) além de outras
estruturas (esfíncteres, músculo ciliar, etc.)
Músculo Estriado
Músculo Liso
Esquelético
Cardíaco
Multiunitário
Uma só Unidade
sim
sim
Não
Não
sim
sim
sim
sim
sim
sim
Muito desenvolvido
Deslizamento dos filamentos
finos sobre os grossos
Sistema nervoso somático
Neurogênica
sim
sim
Muito desenvolvido
Não
Não
Pouco
sim
Não
Pouco
Idem
Idem
Idem
SNA
Miogênica
SNA
Neurogênica
SNA
Miogênica
RS
LEC e RS
LEC e RS
LEC e RS
Não
Rápida ou Lenta, dependendo
da fibra
Sim
Não
Sim
Lenta
Muito Lenta
Muito Lenta
Sim
Não
Não
Sim
Curso de Ciências Biológicas 2007 Disciplina de Fisiologia Comparada Ciclo de Neurofisiologia
Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida
141
Comparando os controles neurais de vertebrados e invertebrados
A força de contração das fibras musculares esqueléticas dos vertebrados depende da
freqüência dos PA na unidade motora e no número de unidades recrutadas. No caso dos
invertebrados, raramente, as fibras musculares geram PA. A força de contração depende da
amplitude das ondas de despolarizações e isso acontece porque as terminações nervosas ocorrem ao
longo de toda fibra muscular, “dispensando” a geração e a propagação dos PA. O fato de não ter
que gerar um evento elétrico do tipo tudo-ou-nada, possibilita a graduação de força na própria fibra
muscular. Assim, a quantidade de Ca liberada é proporcional a amplitude dos potenciais pósjuncionais.
Finalmente, outro diferencial é que os invertebrados possuem inervação inibitória nas fibras
musculares assim como mais de uma inervação excitatória (inervação multineuronal) sendo
possível a somação algébrica de sinais no próprio sarcolema.