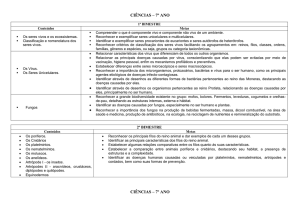
ORIGEM DA VIDA: Como surgiram os
primeiros animais?
por Carlos Daniel Perez
Pese a aparente estabilidade do mundo natural, as mudanças são constantes ao longo
da história evolutiva do nosso planeta. Estas mudanças podem ser observadas e medidas em
escalas de tempo muito variáveis e, independente disto, sabe-se hoje que cada característica
de vida atual é um produto do processo evolutivo. Por esta razão a evolução de todos os
organismos deve ser considerada como a pedra fundamental de todo o conhecimento
biológico.
Atualmente nossos conhecimentos avançaram em campos importantes como
reprodução, desenvolvimento e genética. Além disso, o desenvolvimento de equipamentos
e técnicas nos permite mergulhar de forma mais profunda e intensa na natureza dos seres
vivos. Este maior número de informações associada a interdisciplinaridade que comanda as
pesquisas atuais, permite um avanço significativo para responder a pergunta que inquieta a
humanidade desde suas origens: de onde viemos?
O primeiro indício de vida na Terra data de aproximadamente 3,5 bilhões de anos.
Estas primeiras células eram procarióticas, organismos semelhantes a bactérias. Após um
grande período de diversificação evolutiva dos procariotos, organismos eucariotos
unicelulares apareceram. Embora não possamos saber com certeza a origem dos eucariotos
unicelulares, ela claramente envolve um processo de simbiose. Certos tipos de bactérias
aeróbias devem ter sido englobadas por outras bactérias que eram incapazes de suportar o
aumento das concentrações de oxigênio na atmosfera. A bactéria aeróbia possuía as
enzimas necessárias para obter energia na presença de oxigênio, tendo sido, provavelmente,
a ancestral das mitocôndrias. A maioria dos genes da mitocôndria, mas nem todos, viriam a
residir no núcleo da célula hospedeira. Praticamente todos os eucariotos existentes hoje têm
mitocôndrias e são aeróbios.
Alguns eucariotos unicelulares ancestrais englobaram bactérias fotossintetizantes,
que evoluíram originando cloroplastos; desse modo, esses eucariotos foram capazes de
produzir suas próprias moléculas de alimento usando energia da luz solar. Os descendentes
de uma dessas linhagens, as algas verdes, possivelmente deram origem às plantas
multicelulares.
Em alguns eucariotos que não se tornaram “residência” para cloroplastos, e alguns
que se tornaram, evoluíram características semelhantes às dos animais, dando origem a uma
variedade de filos coletivamente chamados de protozoários.
Acredita-se que os verdadeiros animais tenham surgido como um grupo
monofilético a partir de um ancestral protista, há mais de 700 milhões de anos atrás. A
dúvida está em qual protista? Há três teorias sobre a possível origem:
1. Hipótese ciliada sincicial: (ancestral:protista ciliado)
É suportada pelas semelhanças entre os ciliados modernos e alguns turbelários
atuais. Foi consolidada em 1950 por J.Hadzi e E.D.Hanson.
Sugere que o metazoário ancestral foi um protista ciliado, multinucleado, com
simetria bilateral, que teria modo de vida bentônico (rastejando no fundo com sua abertura
oral dirigida para frente raspando o substrato). Em um passo evolutivo maior as superfícies
nucleares são divididas pela formação de membranas celulares, produzindo uma epiderme
celular contornando uma massa sincicial interna o que confinava a atividade de cada núcleo
a certas regiões do corpo e assim se tornaram internamente compartimentados. O resultado
disso e de outras mudanças foi um organismo acelomado semelhante a um turbelário (filo
Platyhelminthes).
Assim, esta teoria sugere que os primeiros ancestrais dos Metazoa eram acelomados
e bilaterais). Esta teoria é reforçada pelas grandes similaridades entre os ciliados modernos
e vermes achatados acelomado quanto a tamanho, forma, simetria, local da boca e cílios na
superfície. No entanto, alguns fatores contrariam esta idéia: não ocorre na embriologia dos
turbelários nenhum processo de celularização semelhante; a teoria não explica a presença
de espermatozóides flagelados nos metazoários; e, principalmente, implica que a simetria
radial de cnidários seja derivada de uma simetria bilateral primária, para o que não há
nenhuma evidência.
2. Hipótese flagelada colonial: (ancestral=protista flagelado)
Proposta por Ernst Haeckel em 1874 e bem difundida por Hyman (1940). É a teoria
mais aceita atualmente. Sugere que um protista colonial flagelado teria originado um
ancestral metazoário planulóide (plânula=tipo larval básico dos cnidários).
O protista ancestral seria uma colônia esférica de células flageladas ocas. As células
individuais dentro da colônia tornaram-se diferenciadas para papéis funcionais específicos
(células reprodutivas, nervosas, somáticas, etc), subordinando assim a independência
celular para o bem-estar da colônia. A forma ancestral colonial seria radialmente simétrica
no princípio, talvez semelhante às larvas plânulas livre-natantes dos cnidários. Esta larva é
radialmente simétrica e não possui boca. Os cnidários, com sua simetria radial, poderiam
ter evoluído a partir desta forma. A partir daí, desenvolveram algum grau de orientação
locomotora, antero-posterior, e algum nível de especialização de células em funções
somáticas e reprodutivas separadas (comum em protistas coloniais atuais), caracterizando a
chamada BLASTEA. Neste cenário, o primeiro metazoa surgiu pela invaginação da blastea
e o resultado seria um animal com dupla membrana, corpo como uma gástrula, denominado
GASTREA. A gastrea teria uma abertura como um blastóporo, parecendo a gástrula de
muitos invertebrados viventes. Ambos estágios são vistos durante a ontogenia de animais
modernos; a gastrea seria o precursor de Cnidaria. Dentro desta idéia, a simetria bilateral
seria derivada. A evolução de um eixo do corpo antero-posterior, o movimento
unidirecional e a cefalização, provavelmente, co-evoluíram em algum grau e devem
coincidir com a invasão dos ambientes bentônicos e o desenvolvimento da locomoção
rastejante. Parece ser que a origem da condição triploblástica surgiu após o aparecimento
das formas bilaterais.
Em 1985, Nielsen propôs uma teoria chamada Trochaea, uma ampliação da
apresentada por Haeckel. Na teoria trochaea a evolução inicial dos filos animais teria
comportado uma série de ancestrais holoplanctônicos, baseado no fato de que estruturas
ciliadas para alimentação estão presentes em larvas de muitos invertebrados e certos
animais adultos.
3. Hipótese da origem polifilética
Muitos zoólogos preferem crer que os metazoários tiveram uma origem polifilética,
tendo sido derivados independentemente de mais de um grupo de organismos unicelulares.
Assim sendo, as esponjas, cnidários, ctenóforos e eumetazoários restantes evoluíram de
forma independente não havendo um esquema único para a origem de todos eles. A
principal razão é por acreditar que as esponjas evoluíram a partir de um organismo
unicelular diferente dos metazoários superiores, assim metazoa seria um táxon polifilético.
O principal argumento é que apesar dos coanoflagelados
parecerem claramente
relacionados com as esponjas, não é claro o grau de relação das esponjas com os demais
Metazoa. É difícil de imaginar como um organismo com organização tão simples como
uma esponja poderia se desenvolver em organismos com uma estrutura corpórea complexa
e órgãos internos. No entanto, esta hipótese não vem sendo apoiada em estudos
moleculares.
As evidencias filogenéticas recentes baseadas na unidade menor do RNA
ribossômico e em similaridades de vias bioquímicas complexas corroboram a hipótese
flagelada colonial, que os metazoários formam uma unidade monofilética (seguindo o
esquema filogenético de Wainright et al. 1993) incluindo aos coanoflagelados. Esta
hipótese também é corroborada com dados morfológicos (Nielsen, 1995). O grupo-irmão
dos metazoários parece ser o dos fungos. A evidência molecular exclui a hipótese ciliada
sincicial porque os metazoários são aparentemente mais próximos das algas eucarióticas e
plantas do que o são dos ciliados.
Tudo que podemos afirmar é que algum tempo, durante a Era Proterozóica, um
protista (ou protozoários) desconhecido desenvolveu-se em uma forma colonial minúscula
que, eventualmente, veio a ser o ancestral dos Metazoa. A verdadeira natureza deste
organismo é desconhecida, já que tinha corpo mole e não deixou traços. No entanto, por
estudos fósseis, morfológicos e moleculares comparados, pode-se inferir sobre este
ancestral e buscar entender as relações filogenéticas entre os metazoários.
A conceituação de filos animais sofreu importantes mudanças desde suas origens
com as seis classes de Linné e as 4 ramificações de Cuvier. A medida que o conhecimento
sobre desenvolvimento e morfologia comparados dos metazoários foi se acumulando ao
longo dos anos, o número de táxons usualmente reconhecidos como filos viventes,
considerando que cada um representasse planos corporais derivados únicos, cresceu a cerca
de 35 filos. Se filo for definido como um grupo de animais que compartilham um plano
corporal devido a uma história comum, este deve ser monofilético, no sentido original do
termo (Haeckel, 1866), mas pode ser também parafilético ou holofilético. Formas
morfologicamente intermediárias que ligam filos a ancestrais comuns, não foram
encontradas. Na falta de tal elo análises pré-cladísticas, baseadas na morfologia dos filos
tiveram que se apoiar em eventos evolutivos hipotéticos para explicar a transição desde a
forma ancestral assumida até os planos corporais descendentes que vemos hoje. Em alguns
casos, especulações sobre esses eventos levaram a construção de cenários que
posteriormente serviram como suposições nas quais se baseavam interpretações de toda a
árvore de vida dos metazoários. No entanto, dados de estudos da morfologia comparada,
em qualquer estágio de vida, foram insuficientes para estabelecer uma filogenia de
Metazoa, amplamente aceita, em especial porque esses dados não são fortes o bastante para
falsificar/invalidar hipóteses alternativas. As técnicas cladísticas, embora forneceram uma
abordagem renovada para as filogenias em estudo, se provaram difíceis para aplicar aos
planos corporais já que homologias claramente identificáveis e informativas são raras.
Assim, diversos cenários concorrentes permaneceram viáveis, alguns por mais de um
século.
As moléculas proveram um avanço operacional importante para esclarecer
problemas sobre a origem dos filos animais. Evidências moleculares relativas ao
desenvolvimento e comparação gênica ofereceram certo discernimento das bases genéticas
da evolução dos planos corporais. As evidências moleculares filogenéticas da topologia da
ramificação não estão relacionadas aos dados de desenvolvimento ou morfológicos,
fornecendo testes independentes de hipóteses prévias. As afinidades indicadas pelas árvores
moleculares são assunto de interpretação, e muitas perguntas sobre os relacionamentos
entre os filos ainda permanecem, porém as filogenias moleculares também foram
significantemente capazes de favorecer nosso conhecimento sobre os padrões de
ramificações dos descendentes entre muitos dos filos. Não obstante, moléculas só podem
ser obtidas de membros viventes dos filos. Filogenias moleculares indicam divergências
entre os últimos ancestrais comuns dos filos atuais, mas não indica suas morfologias.
Assim, os fósseis são uma importante fonte de dados para estudar a evolução inicial dos
animais. Em contraste com as moléculas, interpretações dos registros fósseis iniciais é
dependente da morfologia. Estudos fósseis revelaram inúmeros táxons importantes porém
extintos, que podem ser associados com filos viventes no campo morfológico, ou que são
tão distintos que não podem ser associados a nenhum grupo vivente. O que os fósseis ainda
não revelaram foram séries morfológicas intermediárias que liguem filos viventes aos seus
ancestrais comuns.
Com tudo que foi exposto, há uma nova gama de cenários evolutivos que tentam
explicar a origem e radiação dos filos metazoários. Alguns elementos desses novos cenários
guardam uma enigmática semelhança com aquelas teorias das escolas pré-cladísticas, com
filogenias baseadas em morfologia.
As classificações mais recentes costumam separar os Metazoa em Eumetazoa
(considerando a presença de tecidos verdadeiros) de outros grupos como as esponjas e, as
vezes, também Placozoa, os quais são denominados Parazoa. Esta separação implica a
parafilia dos organismos diploblásticos. Diversos estudos foram realizados, e dados
moleculares com mecanismos para reduzir a atração dos ramos longos, assim como estudos
morfológicos, defendem a monofilia de Metazoa e a parafilia dos diploblásticos. Por esta
hipótese, conhecida como hipótese Eumetazoa, Placozoa estaria agrupado com os
Eumetazoa (Cnidaria, Ctenophora e Bilateria), deixando apenas Porífera em outro clado.
Morfologicamente esta hipótese é mais parcimoniosa que a monofilia de Diploblásticos, já
que esta implicaria em diversas aquisições independentes entre, por exemplo, Cnidaria +
Ctenophora e Bilateria, ou, como outra alternativa, que a simplicidade das esponjas fosse
derivada e não primitiva. Não obstante, análises moleculares recentes (Ender &
Schienwater, 2003) não concordam com a proximidade entre cnidários e placozoários.
Dessa forma, Porífera aparece como um grupo de suma importância sendo
considerado o primeiro animal multicelular a surgir no planeta. Pela idéia de que um plano
corporal corresponde a um filo (Valentine, 1992) as esponjas sempre foram consideradas
um único filo. A sinapomorfia apresentada por este grupo seria a presença de um sistema de
circulação de água através de seu corpo do qual dependem praticamente todas as funções
fisiológicas destes animais. Não obstante, devido a grande diversidade de seus
representantes, a monofilia de Porifera foi várias vezes questionada, desde o ponto de vista
molecular e também morfológico. A confirmação de que Coanoflagelados seria um grupo
irmão de Metazoa, leva a crer que o ancestral dos metazoários seria um organismo similar a
uma esponja, o que indica que vários caracteres de Porífera seriam plesiomórficos entre
Metazoa e assim as esponjas seriam provavelmente parafiléticas. De fato, estudos recentes
demonstram que Porifera é uma reunião parafilética de, pelo menos, duas linhagens basais
de Metazoa (Borchiellini et al., 2001). Isto também reforça a idéia dos coanoflagelados
como grupo irmão de Metazoa o que representa que a similaridade entre este grupo e os
coanócitos das esponjas representariam homologias. Assim, metazoários não espongiários
derivariam de uma esponja ancestral que perdeu seus atributos, em especial coanócitos e
sistema aqüífero. A descoberta de demosponjas da família Cladorhizidae, altamente
derivada, reforça esta idéia. Este grupo mudou seu plano corporal para adequar-se ao
oceano profundo, perdendo o sistema aqüífero e desenvolvendo um eficiente sistema de
captura de presas. Algo similar poderia ter ocorrido na origem dos Metazoa. De fato, foram
descobertos alguns fósseis pré-cambrianos similares a esponjas, porém sem nada parecido a
câmaras coanocitárias.
Dentro de Porífera, a confirmação da parafilia, trouxe consigo mais informações
sobre as relações entre as tradicionais classes. A classe Hexactinellida aparece como a mais
basal em estudos paleontológicos e moleculares. Esta classe exibe alguns caracteres
morfológicos bem particulares como, por exemplo, a organização sincicial, o que levou a
alguns autores proporem a separação desta classe das demais. No entanto, estes caracteres
são autopomórficos, não dando indicações sobre a posição filogenética de Hexactinellida.
Nenhum estudo conseguiu até o momento assegurar a parafilia das esponjas silicosas
(Hexactinellida e Demospongiae). Não obstante, a posição da Classe Calcarea foi
claramente definida por Borchiellini et al (2001). Este estudo demonstrou que os
representantes da Classe Calcarea estão mais proximamente relacionados com Eumetazoa
que com as demais esponjas. Com as novas descobertas, a filogenia leva a necessidade de
definir classificações mais modernas. Neste contexto, segundo os autores, o táxon Porífera,
parafilético, não deverá mais ser usado. A antiga Classe Calcarea deverá ascender ao status
de Filo (Filo Calcispongia, reunindo as classes Calcinea e Calcaronea).
Uma vez esclarecida a posição basal dos poríferos entre os metazoários, nos resta
analisar agora as relações entre os eumetazoários. Na base deste grupo se encontram
organismos que, assim como algumas esponjas, apresentam simetria radial. A separação
dos grados segundo a simetria parecia ser pouco discutida já que simetrias distintas
implicariam em organizações corporais bem diferentes. Além do mais, a posição mais basal
dos radiados reflete uma das teorias mais aceitas sobre a origem dos metazoários, que
implica na simetria radial precedendo organismos bilaterais, como já foi discutido. Os
radiados pareciam ter derivado de um ancestral comum que não estaria na nossa linha direta
de descendência. Todos os radiata possuem tecidos diferenciados, mas normalmente não
apresentam mais que um tecido interno (endoderma) e outro externo (ectoderma) com certo
nível de especialização. Desde o ponto de vista embriológico eles não sofrem gastrulação e
por isso não apresentam mesoderma, um tecido que caracteriza todos os demais animais. A
maioria é radialmente simétrico. Seu plano corporal está baseado em um cilindro aberto em
uma extremidade, com esta abertura servindo tanto para a alimentação como para a
eliminação de partículas.
A transição para um corpo vermiforme é um dos eventos mais importantes na
evolução animal, levando à impressionante radiação dos Bilateria. No entanto, havia grande
dificuldade em identificar o grupo irmão de Bilateria, com várias hipóteses propostas.
Wallberg et al. (2004) utilizando um grande número de táxons reavalia estas hipóteses e
parece ter colocado fim a discussão. Eles analisam as 3 hipóteses que propunham a relação
entre os táxons Ctenophora, Cnidaria e Bilateria (denominados por eles como
Epitheliozoa). A primeira hipótese, mais antiga, considerava a simetria radial na base dos
eumetazoários e une os chamados Coelenterata (Cnidaria + Ctenophora). Vale salientar
que aqui, não cabe mais a terminologia radiata, pois no seu sentido original, este clado
incluiria também as esponjas, as quais já estão claramente definidas na base de Metazoa. A
hipótese Coelenterata era a hipótese de Haeckel amplamente difundida por Hyman. A
segunda hipótese, com base em análises cladísticas de caracteres morfológicos, apontava os
ctenóforos como grupo irmão dos bilaterais. Este clado, denominado Acrosomata
compartilhava diversos caracteres morfológicos, incluindo a presença de acrosoma nos
espermatozóides, o que deu nome ao clado. A terceira hipótese, baseada nas análises da
unidade menor do RNA ribossômico apontava os ctenóforos como grupo irmão do clado
Cnidaria + Bilateria. Esta idéia, denominada pelos autores como Planulozoa foi confirmada
com evidências bem robustas no trabalho. Esta hipótese dos Ctenóforos ocupando uma
posição mais basal entre os eumetazoários já era defendida por outros autores (CavalierSmith et al. 1996; Kim et al. 1999 e Borchiellini et al. 2001) e implica que vários caracteres
considerados sinapomórficos entre Ctenophora e Bilateria devem ser revistos. A
semelhança superficial entre eles deve ser devida a homoplasias. De fato, já foi visto
acrossoma em espermatozóides de Porífera e Cnidaria. Por estas análises, Epitheliozoa e
Planulozoa constituem agrupamentos monofiléticos.
Estabelecer as relações filogenéticas não é tarefa fácil. Trabalhos que levam em
conta a morfologia apenas, as vezes não apresentam argumentos forte o bastante para
refutar hipóteses contrárias as que eles próprios tentam comprovar. Por outro lado, análises
moleculares também têm a desvantagem de não poder incluir grupos fósseis que podem
representar a chave para as homologias, além de apresentar algumas dificuldades
metodológicas. Uma das maiores dificuldades encontradas nos estudos mais recentes de
filogenia é evitar os artefatos criados nas árvores pelo fenômeno de atração das ramas
longas. Este é um dos maiores problemas para entender a filogenia na base de Metazoa. O
clado composto pelos organismos bilaterais forma uma rama longa, podendo a sua posição
representar um artefato em algumas análises. Alguns autores chegam a admitir e alertar
para este problema questionando os próprios resultados (por exemplo: Collins & Valentine,
2001) e outros buscam usar mecanismos e técnicas que diminuam o problema (por
exemplo: Borchiellini et al., 2001).
Desta forma, considerando os últimos estudos realizados, Cnidaria estaria
mais relacionado com os Bilateria que com Ctenophora. No entanto, definir qual dos
grupos de cnidários é mais basal é uma tarefa que ainda parece estar longe de ser concluída.
Hyman (1940) sugeriu que Hydrozoa seria o ancestral do grupo. Por várias décadas,
pesquisadores concordaram com esta hipótese baseados na morfologia e características de
desenvolvimento. Por outro lado, considerando algumas teorias para a origem dos
metazoários, como a de um ancestral colonial, somado a estudos do genoma, a idéia de
Anthozoa como grupo mais basal passou a ser defendida por alguns autores. O fato
preponderante é a presença em Anthozoa de genoma mitocôndrial circular semelhante a
outros animais, enquanto que Hydrozoa e os outros grupos com medusa apresentam o
genoma mitocôndrial linear. Análises da subunidade menor do RNA ribossomal (SSU)
coloca a Hydrozoa na base, porém não refuta a outra hipótese. Assim, o subfilo Anthozoa é
alternativamente considerado o grupo mais basal ou o mais derivado. A primeira hipótese
posiciona aos pólipos como forma original e a medusa (e a metagênese) como forma
derivada (Fig. 1A). A segunda teoria indica que no típico ciclo de vida metagênico, a
medusa é gametogênica e constitui o estádio definitivo ou adulto com um pólipo que atua
como uma larva persistente. Assim é razoável que o pólipo evoluiu secundariamente e
perdeu a forma original do corpo, a medusa, localizando aos antozoários como o táxon mais
derivado (Fig. 1B). Willmer (1991) sugeriu que os antozoários e sua condição bilateral
representariam o estado primitivo do filo Cnidaria, baseado em evidencias dos organismos
viventes e fosseis. Analisando os registros fosseis é impossível predizer quem é mais
primitivo se o pólipo ou a medusa, ambas formas foram encontradas em depósitos précambrianos (Glaessner, 1984).
Definir a filogenia dentro de cnidaria é fundamental para entender a evolução dos
metazoários. Segundo a teoria de origem de Metazoa, o grupo mais basal em Cnidaria seria
diferente e a origem dos Bilateria seria bem estabelecida. A maior parte dos autores atuais
aceita a condição radial como primitiva, colocando assim os pólipos, e com isso a classe
Anthozoa na base do filo. Isto é corroborado por estudos morfológicos e moleculares. No
entanto, a ultima teoria da possível origem dos cnidários foi publicada, faz horas apenas,
por Seipel & Schmidt (2005) baseada em estudos moleculares da musculatura e na
ontogenia da formação da medusa. O trabalho conclui que o ancestral dos cnidários seria
um organismo triploblástico móvel. Em conseqüência, a condição diploblástica teria
evoluído secundariamente nas larvas e pólipos dos cnidários, qual era suportado pela Teoria
Sincicial de Hadzi em 1953.
Antes de seguir discutindo e inferindo sobre a evolução dos metazoários devemos
considerar um grupo de organismos bem particulares, os myxozoários. Estes endoparasitas
microscópicos de peixes e invertebrados com corpo amorfo e plasmodial sempre foram
considerados protozoários, ate que em 1970 ganharam a condição de filo pela descoberta da
multicelularidade de seus esporos infectantes. Devido a grande similaridade dos
nematocistos com as cápsulas polares dos myxozoários Sidall et al, 1995, baseados em
caracteres morfológicos e moleculares, propuseram que Myxozoa pertencia a Cnidaria.
Trabalhos posteriores não suportaram esta nova posição filogenética (Kim et al, 1999).
Somado a isto, Anderson et al, 1998 e Ferrier & Holland, 2001 demonstraram que os
myxozoarios possuíam Hox genes, um tipo de genes totalmente ausentes dentro dos
diploblásticos. Esta discussão seguiu até que Okamura & Canning (2003) apresentaram um
trabalho que confirmava que os myxozoarios não são cnidários, sugerindo que algumas
espécies de myxozoarios possuíam duas formas morfológicas alternativas, uma vermiforme
e outra plasmodial. A forma vermiforme descrita anteriormente como Buddenbronckia e
não alocada em nenhum filo, apresenta clara simetria bilateral. Desta maneira, o DNA
ribossômico, os Hox genes e os dados estruturais considerados em seu conjunto, indicariam
que os myxozoários não são cnidários e que formariam parte da linhagem dos Bilateria no
reino animal. Dessa forma, algumas hipóteses bem robustas colocam Myxozoa e alguns
vermes acelomados como os primeiros clados bilaterais, assegurando a posição dos
Myxozoários na base de Bilateria (por ex. Telford et al. 2003).
Já dentro dos Bilateria, tradicionalmente os filos são divididos em Protostômios e
Deuterostomios. Esta divisão não implica apenas a diferença na origem da abertura oral,
mas também uma série de distinções na embriologia. Organismos caracterizados como
protostômios normalmente são associados ao conceito de que o blastóporo, ou a região
próxima a ele, irá originar a boca do organismo. No entanto, diversas outras características
estão implícitas neste conceito. A clivagem do tipo espiral, principalmente determinada, a
formação do celoma por esquizocelia, ou seja, a partir de bandas mesodérmicas, e a origem
do endomesoderma a partir de um blastômero especial (4d) são outras características do
desenvolvimento dos protostômios. Por outro lado, organismos deuterostomios apresentam
a boca sendo formada de uma região diferente da do blastóporo, a clivagem sendo
principalmente radial e geralmente indeterminada, o celoma formado a partir da fusão de
bolsas enterocélicas (exceto nos cordados) e o endomesoderma a partir de bolsas
enterocélicas (exceto nos cordados). Considerando este conjunto de características, nem
sempre é fácil determinar a posição de alguns táxons que possuem características dos dois
tipos de desenvolvimento misturadas (por exemplo, os filos conhecidos como lofoforados).
Alguns autores acreditam que podem, através da análise da forma de gastrulação de
organismos basais em cada divisão (por exemplo, poliquetos entre protostômios) poderia
levar a inferir sobre o tipo de gastrulação do ancestral do grupo. Comparações usando
marcadores moleculares entre a parte oral e o blastóporo em protostômios basais e
deuterostomios basais sugeriram que o ancestral destes grupos (comumente chamado de
Urbilateria) usava o mesmo set de genes para formar a boca e o ânus desde lados opostos
ao blastóporo (Lartillot et al. 2002). Assim, a divisão entre protostômios e deuterostômios
não é marcada pela forma independente e diferente de formar a boca (como conceituado
por Grobben, 1908). Em realidade Deuterostomia seria um modo derivado de gastrulação,
que facilmente teria evoluído desde a Protostomia. Estas considerações foram analisadas
em um estudo de Technau & Scholz (2003) avaliando a origem e evolução do endoderma e
mesoderma. Além de proporem que a invaginação/epibolia deve ser a forma de gastrulação
ancestral, os autores, considerando a Anthozoa como o grupo mais basal entre os
diploblásticos supõe que o processo de dobras epiteliais que dá origem aos mesentérios dos
antozoários deve ter ocorrido de forma similar na origem do mesoderma a partir de
evaginações no Urbilateria. Eles sugerem que os genes que especificam o endoderma e
mesoderma em Bilateria deve ter evoluído suas funções a partir de papéis ancestrais na
regulação básica celular como mobilidade celular, adesão e proliferação.
A divisão dos Protostômios segundo a presença de uma cavidade corpórea
celomática também vendo sendo discutida sob alguns aspectos. De forma geral se considera
três possibilidades:
1. Acelomados (sem cavidade celomática)-A mesoderme forma uma massa de tecido mais
ou menos sólido (Mesênquima), às vezes com espaços (lacunas), entre o tubo digestivo e a
parede do corpo.
2. Celomados ou eucelomados- Há o desenvolvimento de uma cavidade (celoma)
preenchida de líquido entre a parede do corpo e o tubo digestivo. Tal cavidade está dentro
da própria mesoderme e está completamente envolvida por um tecido fino, o peritônio
(derivado da mesoderme), de forma que os órgãos do corpo não são livres dentro do espaço
celômico, mas separados dele pelo peritônio.
3. Pseudocelomados ou blastocelomados- Possuem cavidades pequenas ou grandes do
corpo, que nem sempre são formadas a partir da mesoderme e não são envolvidas por
peritônio ou outra forma de tecido mesodermal. É assim chamada de falsa cavidade. Os
órgãos ficam livres na cavidade do corpo mergulhados diretamente no seu fluido. Em
muitos casos o espaço representa remanescentes persistentes da blastocele embrionária e aí
não há nada de falso. Por esta razão, alguns autores preferem usar o termo blastoceloma.
Acredita-se que a origem do celoma esteja relacionada com o modo de vida
bentônico assumido por alguns organismos bilaterais, pequenos. Nesta condição, um
esqueleto fluido (hidrostático) seria essencial para certos tipos de movimento. A evolução
de uma cavidade do corpo cheia de fluido, contra o qual os músculos poderiam operar,
ofereceria uma grande vantagem locomotora adicional proporcionando uma circulação
média e espaço para o desenvolvimento de órgãos. Há teorias que sugerem o aparecimento
do celoma de uma só vez, outras em duas vezes.
1. Teoria enterocélica
O celoma teria originado pelo estreitamento e isolamento do divertículo estomacal
embrionário.
2. Teoria gonocelomica
Um primeiro espaço celômico surgiu pela parede das cavidades gonadais derivada
mesodermicamente e que persistiu subseqüentemente pela liberação dos gametas.
3. Teoria nefrocelomica
Protonefridio dos vermes achatados expandiriam para as cavidades celômicas, argüindo
que o celoma derivou de estruturas ectodérmicas derivadas. Certamente as cavidades
celômicas de muitos animais estão relacionadas a funções excretoras.
Não obstante, alguns estudos recentes consideram que a divisão dos animais
bilaterais segundo a cavidade corporal é arbitrária já que resultaria em grupos que não são
monofiléticos. Assim, estudos filogenéticos moleculares recentes (Aguinaldo et al., 1997;
Balavoine & Adoutte, 1998) têm proposto uma hipótese filogenética diferente para os
Bilateria. Os resultados transferem a quatro filos considerados tradicionalmente como
Protostomia para Deuterostomia (Brachiopoda, Chaetognatha, Ectoprocto e Phoronida).
Além disso, os grandes agrupamentos dos protostômios em acelomados, pseudocelomados
e eucelomados parecem, realmente, não ser monofiléticos. As evidências sugerem que
algum tempo depois que o ancestral deuterostômio divergiu do ancestral protostômio, no
Pré-Cambriano, os protostômios dividiram-se novamente em dois grandes grupos
monofiléticos (ou superfilos): os Ecdysozoa, contendo os filos que passam por uma série de
mudas durante seu desenvolvimento, e os Lophotrochozoa, incluindo os filos de
lofoforados e os filos cujas larvas são assemelhadas as trocóforas.
Dessa forma, três clados principais nos Bilateria aparecem desde cedo nos registros
fósseis do Cambriano: Deuterostomia, Lophotrochozoa e Ecdysozoa. De todas as hipóteses
filogenéticas recentes, a divisão dos protostômios parece ser uma das mais importantes. A
evidência do clado Ecdysozoa, caracterizado pelo hábito de muda, foi comprovado
morfologicamente (Barnes et al., 1993), pelo RNA ribossômico (Aguinaldo et al., 1997;
que deu nome ao clado) e por características no Hox genes (de Rosa et al., 1999).
Ecdysozoa apresenta uma grande diversidade morfológica, incluindo desde vermes
pseudocelomados (Priapulida) ou com pseudocele reduzida (Nematoda) até filos superiores
e complexos como Arthropoda, que é basicamente hemocélico, porém com pequenos
espaços intramesodermais. Mesmo com esta disparidade, diversos estudos recentes
suportam a monofilia do clado. Considerando que a grande maioria dos animais viventes é
Ecdysozoa, é fundamental compreender as condições e oportunidades que se combinaram
para originar os fatores principais deste clado.
Acredita-se que tanto o ancestral dos Protostômios e Deuterostômios como o
ancestral comum entre Ecdysozoa e Lophotrochozoa, eram organismos simples, e
provavelmente paracelomados. Este último termo se refere a organismos pseudocelomados
e acelomados (Valentine & Collins, 2000). O fóssil mais antigo é da Idade Neoproterozóica
(cerca de 570 Milhões de anos), são rastros pequenos com características paracelomadas.
Representa a evidência mais antiga de locomoção animal, parecendo ter sido criado pela
ação de músculos pedais ou das paredes de organismos bilaterais. Também poderiam ser
rastros deixados pelo sistema de locomoção mucociliado comum em invertebrados para a
alimentação e limpeza de superfícies e para a locomoção em pequenos animais, como os
usados até hoje em alguns cnidários, Lophotrochozoa e Deuterostômios.
Animais que apresentam mudas não tem cílios ectodérmicos móveis e, em
conseqüência não podem usar sistemas mucociliares ectodérmicos. O clado Ezdysozoa
evoluiu a partir de animais que se ancoravam com a cutícula e se movia por ondas
peristálticas e/ou pela introverte. Eram provavelmente rastejantes ou enterradores. Dessa
forma, a muda deve ter evoluído entre o ancestral Ecdysozoa/Lophotrochozoa ou no último
ancestral comum em Ecdysozoa. A função hidrostática do Euceloma no tronco de vários
protostômios (ex: anelídeos) e Deuterostômios (ex: cephalopoda) é reduzido em Ecdysozoa
a fluidos no compartimento da blastocele, como pseudoceloma ou hemoceles. Da mesma
forma, a função hidrostática na luz dos tentáculos de alimentação de alguns protostômios
(ex: Phoronida) e deuterostômios (Ex: Pterobranchia) não aparece em Ecdysozoa. A
evolução de tentáculos ciliados neste clado foi impedida pela ausência de tratos
mucociliados. É possível que a ausência de um compartimento celômico no tronco em
Ecdysozoa também esteja associado ao hábito de muda. Uma vez que a muda permite não
apenas aumentar estruturas cuticulares como espinhos ou apêndices que funcionam como
estruturas de locomoção auxiliares, mas também promovem um fortalecimento da cutícula.
Os espaços celômicos blastocélicos provêm resistência e apoio para a ação dos músculos da
parede do corpo, reforçados pela cutícula que os envolve. A presença deste sistema deve
haver inibido a evolução de uma compartimentalização adicional para dar suporte as
atividades de Ecdysozoa.
A origem e conseqüência da muda são assim, fundamentais para entender a história
do clado. Em Ecdysozoa, apresentar muda significa uma obstrução das estruturas ciliares
ectodérmicas. Segundo evidências filogenéticas e fósseis, os primeiros membros de
Bilatéria seriam pequenos, vermiformes e, provavelmente, tinha desenvolvimento direto.
Em geral organismos pequenos apresentam baixo esforço reprodutivo e a presença de larva
aumentaria a mortalidade e a reprodução seria inviável. Dessa forma, o desenvolvimento de
larvas planctônicas ocorreu de forma independente mais de uma vez (ao menos 3) dentro de
Bilatéria. Clados sem muda evoluíram larvas que nadam e se alimento por tufos de cílios ou
bandas ciliadas, incorporando essas formas aos seus sistemas de desenvolvimento muito
cedo. Por outro lado, dentro de Ecdysozoa, larvas alimentares não tem tratos ciliados
alimentares e evoluíram pela modificação, durante estágios iniciais, empregando setas ou
apêndices para gerar correntes de alimentação. É provável que a muda tenha evoluído como
uma adaptação que permitisse o aumento, durante o crescimento do animal, de estruturas
secretadas pela cutícula como espinhos e outras, utilizadas para a locomoção auxiliar. Entre
os Ecdysozoa ocorrem táxons com desenvolvimento direto (como Nematoda) e outros com
estágios bentônicos prematuros com introverte que lembra um pouco o adulto (como
Loricifera). Este último pode ser considerado larva, porém é mais provável que se trate de
elaborações de juvenis produzidos em um desenvolvimento direto ancestral. De todos
modos, nada que se assemelhe a larva planctônica primária ciliada. Os cílios em
Ecdysozoa, quando presentes, são exclusivamente sensitivos (sensilla).
Se as larvas planctônicas, de fato, surgiram mais de uma vez entre os Bilatéria,
então hipóteses evolutivas que se apóiam no fato de larvas planctônicas serem
plesiomórficas para estes clados, tanto presente no clado ancestral como larva (por ex.
Nielsen, 1995) ou representando elas mesmas os ancestrais (Por ex. Davidson et al., 1995 e
Petterson et al., 1997) devem ser revistas. Apesar de toda esta discussão, as considerações
sobre a origem independente de larvas planctônicas dentro de Bilatéria ainda não são
conclusivas (Valentine & Collins, 2000).
Dentro de Ecdysozoa a filogenia ainda não está bem definida. Nielsen et al., 1996,
usando caracteres morfológicos une Arthropoda + Tardigrada + Onychophora em um único
clado, também apoiado por evidências moleculares. O filo Nematoda apresenta uma rama
muito longa, sendo difícil a utilização do 18S RNA ribossomal. No entanto, Aguinaldo et
al., 1997 conseguiu ramas mais curtas e uniu este filo aos Ecdysozoa. Alguns estudos
colocam este filo junto com Nematomorpha (Nielsen et al., 1996). Os filos Priapulida e
Kinorhyncha conformariam um clado irmão aos demais Ecdysozoa (Aguinaldo et al.,
1997). Já o filo Loricifera ainda não apresenta amostras do gene 18S, sendo duvidosa sua
localização. Não obstante, por evidências morfológicas deve estar entre os Ecdysozoa,
aliado, provavelmente a Priapulida + Kinorhyncha. O filo Cahetognatha também forma
rama longa e é difícil definir sua posição filogenética. Alguns estudos o consideram dentro
de Ecdysozoa, porém sua posição é ainda instável e duvidosa.
Concluindo, Ecdysozoa seria grupo irmão de Lophotrochozoa (Anelídeos, moluscos
e lofoforados). Estes dois clados formam o agrupamento Protostomia, o qual é irmão de
Deuterostomia (Aguinaldo et al., 1997 e Littlewood et al., 1998). Protostomia e
Deuterostomia formam Bilateria. Este, juntamente com Cnidaria forma um clado, no qual
também estaria inserido Placozoa, segundo algumas classificações, o qual unido a
Ctenophora, cuja posição é mais basal forma o grupo Eumetazoa. Dentro de Metazoa
Porífera forma um agrupamento parafilético na base deste grupo.
Como foi visto até aqui, entender a origem e as relações filogenéticas dos animais
não é tarefa fácil. Mesmo considerando apenas os invertebrados marinhos a tarefa não se
torna menos árdua. A primeira e mais óbvia dificuldade que surge é a de tratar da origem e
evolução em um agrupamento totalmente artificial que, ao contrário do que se esperaria em
uma boa filogenia, reúne táxons que possuem em comum uma ausência. Pois bem, a
ausência de coluna vertebral define os invertebrados. Proposto por Lamarck em 1801, este
termo, apesar de não constituir um clado ou um táxon nem representar qualquer categoria
hierárquica reconhecida no sistema de Linneaus, segue em uso na grande maioria dos livros
didáticos e nos programas de cursos de graduação e pós-graduação. Nenhuma característica
positiva é capaz de abranger todos os seus representantes. A diversidade de formas,
habitats, hábitos é tão grande que pode ser exemplificada quando consideramos seus
representantes. Os chamados Invertebrados são formados por cerca de 35 filos e mais dois
subfilos do Filo Chordata, o mesmo que contém os seres humanos e demais vertebrados. É
assustador! Mais ainda se considerarmos que os invertebrados constituem cerca de 97% dos
organismos viventes! Daí a grande dificuldade de estabelecer teorias e classificações que
reflitam a filogenia do grupo. É quase como discutir a origem e evolução do reino animal.
Nos oceanos e mares há uma enorme variedade de formas de vida, e um número de
táxons superiores muito maior do que no ambiente terrestre. Segundo as estimativas, o mar
apresenta apenas 20% de todas as espécies animais, mas estas representam 90% ou mais de
todas as classes ou filos (MAY, 1988). Basta lembrar que a maioria dos filos conhecidos
são exclusivamente ou principalmente marinhos. Dos 35 filos de invertebrados não
cordados, 16 são exclusivamente marinhos e apenas 2 não apresentam representantes
marinhos (Pentastomida, parasita de vias respiratórias aéreas de vertebrados, e
Onychophora, exclusivamente terrestre).
É lógico que, depois de tudo que foi visto, estamos longe de por um ponto final no
tema. Se nas últimas 3 décadas a grande quantidade de artigos publicados sobre a evolução
dos metazoários nos levou a discutir, refletir e reconsiderar antigos conceitos, certamente,
no mínimo mais 3 décadas serão necessárias para esclarecer os pontos ainda não
conclusivos e a posição filogenética de alguns grupos na grande árvore dos metazoários.
Certamente todos este progresso na compreensão da origem e evolução dos animais, em
especial dos invertebrados marinhos, base para todo o resto, leva a inúmeras modificações
nos sistemas de classificação em uso. É de se esperar que todo este conjunto de
informações leve um tempo para ser incorporado aos sistemas em uso, em especial devido a
inúmeras divergências que surgem entre trabalhos com metodologias e/ou dados diferentes.
Apenas aquelas hipóteses amplamente testadas e corroboradas por mais de um tipo de dado
devem ser incorporadas de forma definitiva nas classificações. Há muita coisa por fazer, e
ainda estamos longe de desvendar os mistérios da origem dos animais e, com isso, entender
a nossa própria origem.