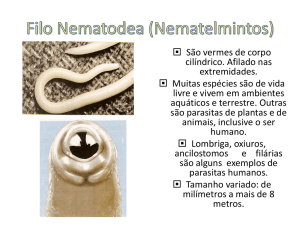
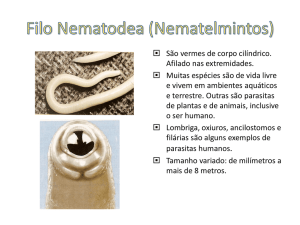
CURSO DE OCEANOLOGIA
DISCIPLINA DE BENTOLOGIA - 2009
ECOLOGIA DA REPRODUÇÃO DO ZOOBENTOS
O Investimento na Reprodução
A manutenção, crescimento e a reprodução competem por recursos. É necessário
energia para a maturação gonadal, a produção de ovos e a defesa do território e da postura
contra os predadores. A manutenção e proteção à desova consomem mais energia do que
lançar os ovos ou larvas livres no meio planctônico.
Um baixo crescimento somático e um alto investimento reprodutivo indicam que é
baixa a probabilidade de sobrevivência dos adultos e, que é mais vantajoso investir, o mais
rápido possível, na geração dos descendentes. Enquanto que um elevado investimento no
crescimento somático, em relação a reprodução, indica que o tamanho do organismo é
importante para suportar os esforços reprodutivos futuros e assim, obter um maior potencial
reprodutivo.
Quando há um grande risco de mortalidade dos indivíduos adultos a seleção favorece
a reprodução, num curto espaço de tempo numa única vez (semelparidade), exemplo é o
poliqueta estuarino Nereididae Laeonereis acuta. Quando os adultos apresentam um baixo
grau de mortalidade e uma fase larval de mais alto risco, então a iteroparidade é mais
vantajosa. Nestas condições, os adultos podem dedicar mais energia para interações
competitivas e para a sobrevivência de um modo geral, inclusive crescimento. Exemplo de
iteroparidade são as várias reproduções que ocorrem ao longo dos mais de uma década de
vida dos mexilhões do gênero Mytilus, que vivem em bancos em costões rochosos.
Espécies que vivem em ambientes onde a probabilidade de extinção é grande
apresentam um incremento no esforço reprodutivo e menor tempo entre as gerações. Ex.
Capitella capitata, que coloniza rapidamente locais com grande quantidade de matéria
orgânica, mas com baixos teores de oxigênio. A espécie combina uma alta fecundidade com
um tempo de geração de apenas 30 dias.
Os Tipos de Reprodução
Reprodução Assexual
Ocorre sem a participação de estruturas reprodutivas especiais. A maioria dos
organismos, com reprodução assexual, possui também reprodução sexual. A reprodução
sexual garante as vantagens da recombinação gênica e da dispersão. A reprodução
assexuada é comum em Cnidarios, Esponjas, Poliquetas e Nemertineos. Desconhecida em
Crinoides, Equinoides, Moluscos e Crustáceos.
Brotamento: é o tipo mais frequente. Ocorre, principalmente, em animais coloniais:
Esponjas, Cnidários, Briozoários, Tunicados (Ascidias). Uma larva (fase sexual) assenta no
substrato e ocupa o espaço formando uma colônia através do brotamento (assexual). É
vantajosa esta alternância entre reprodução sexual (recombinação gênica, busca de novas
áreas, dispersão) e assexual (garante o espaço disponível), principalmente em locais com
grande disputa pelo espaço disponível como é o caso dos costões rochosos.
Fragmentação: ocorre em Turbelarios, Nemertineos, Antozoários (Anemonas). A
anêmona Sagartia lacerata pode apresentar até 70 lacerações. A anêmona colonial
Anthopleura elegantissima no Pacífico, na costa oeste (USA), assenta como larva, forma um
pólipo que se divide em vários clones que asseguram a ocupação do substrato. Existe uma
variedade não colonial Anthopleura sola, que já é considerada uma nova espécie, que ocorre
de forma isolada.
Vantagem: a reprodução assexuada é bastante eficiente na ocupação do espaço
disponível, o que é especialmente útil em costões rochosos. Desvantagens: a) A ausência de
recombinação gênica prejudica a variabilidade genética, diminui as chances de evolução e
determina que os organismos que terão menores possibilidades de sobrevivência caso
1
ocorram mudanças no ambiente b) a ausência de dispersão larval limita a distribuição da
espécie e colonização de novos hábitats.
Reprodução Sexual
Na reprodução sexual a união dos gametas dos dois sexos forma um zigoto, que após
irá originar um animal adulto. O maior benefício é o incremento da diversidade genética, que
possibilita a uma população viver numa maior variedade de hábitats. Descendentes com
combinações genéticas favoráveis, podem resistir com mais sucesso a modificações do
ambiente e enfrentar espécies invasoras ou viroses. Também a seleção sexual na qual
participam caracteres sexuais secundários (ex. tamanho da quela dos caranguejos do gênero
Uca), influencia na escolha dos parceiros e propicia a ocorrência de reprodução diferencial, a
qual determina modificações no pool de genes levando à evolução.
Os sexos podem ser separados ou um indivíduo pode apresentar os dois sexos
(hermafroditismo). A auto fecundação é rara entre hermafroditas. Os cirripédios toráxicos
(cracas) são hermafroditas, bem como espécies de Xylophaga que é um Teredinidae de
profundidade. Há também o hermafroditismo sequencial (ostras, poliquetas), neste caso
ocorre protandria (a espécie matura primeiro, como macho e depois como fêmea). Na
protoginia a espécie irá maturar primeiro, como fêmea e depois como macho. A protandria é
vantajosa quando a fecundidade das fêmeas aumenta com o tamanho e a idade (é o tipo
mais comum). Ex. ostra Crassostrea virginica. A protoginia é vantajosa quando a eficiência do
macho aumenta devido a disputas sexuais ou delimitação do território. É mais frequente em
peixes de arrecife de coral, onde ocorre uma intensa disputa por território.
Reprodução em Organismos com Sexos Separados
A ocorrência de sexos separados é o mais comum entre os invertebrados. Num
determinado momento do seu desenvolvimento os organismos passam pelas fases de
gametogênese, maturação, fecundação e postura.
Fecundação
A fecundação pode ser: a) externa com espermatozóides e óvulos lançados na coluna
d'água; b) com espermatozóides lançados na coluna d'água, mas fecundando os óvulos na
fêmea; c) e interna com a transferência do esperma através de um órgão copulatório ou
através do gonóporo do macho até a massa de óvulos da fêmea.
Fecundação externa: a) machos e fêmeas precisam estar próximos para ocorrer a
emissão dos gametas, sendo comum em Actineas, Gastrópodos Prosobranquios e
Equinoides; b) machos lançam primeiro seus gametas, estimulando a emissão pelas fêmeas,
sendo comum em Actineas, Ceriantharios, Equinodermas, Prosobranquios e nas ostras
Crassostera virginica e C. gigas; c) desova epidêmica, quando a população lança ao mesmo
tempo seus gametas, é comum em poliquetas, nemertineos e bivalvos (Mytilus edulis, Tivela
stultorum).
Os processos acima dão uma maior garantia de que os gametas estarão,
simultaneamente, presentes numa área bastante restrita, melhorando assim as chances de
fecundação dos óvulos na coluna d'água. Para que isto ocorra é importante, que o início da
maturação seja simultâneo na população. Deve haver uma interação entre os fatores
exógenos (gatilhos exógenos) e os endógenos (hormônios). Exemplos de gatilhos exógenos:
fotoperíodo, marés, fases da lua, "blooms" de alimento, elevação da temperatura. Foi
verificado que espécies de cirripédios apresentavam desova simultânea em laboratório.
Enquanto que no campo, Balanus balanoides mais superficial e mais exposto ao fotoperiodo
no intermareal, desovava um mês antes. O caranguejo Uca tem comportamento sexual entre
lua cheia e nova (coincide com as maiores marés), enquanto que o caranguejo do mangue
Cardisoma guanhumi tem picos reprodutivos entre junho e dezembro na lua cheia e nova.
Suas maiores desovas ocorriam um dia antes e um dia depois da lua cheia.
2
Fecundação interna: a) a cópula minimiza a perda de gametas, sendo comum em
gastrópodos, cefalópodos e crustáceos; b) a pseudo-cópula ocorre somente em algumas
espécies, como nos Nemertineos do gênero Lineus. No poliqueta Nereis diversicolor os
machos e a fêmeas ficam enrolados e o macho lança o esperma na massa de óvulos da
fêmea.
Modos de Desenvolvimento do Macrozoobentos
Desenvolvimento direto
Existe pouca relação entre o tipo de postura e o modo de desenvolvimento dos
invertebrados. Existe sim, relação entre o número ou o tamanho dos ovos e o tipo de
desenvolvimento.
No desenvolvimento direto os embriões originam-se de ovos com muito vitelo, dos
quais saem juvenis que irão colonizar áreas próximas a dos adultos. Esta estratégia é
interessante em hábitats com uma boa oferta de recursos, como espaço e alimento, onde não
é necessário que os juvenis fiquem afastados da população de adultos. O desenvolvimento
direto é comum em poliquetas, equinodermas, gastrópodos e crustáceos peracáridos
(possuem marsúpio).
Os crustáceos peracáridos, que são numericamente representativos em termos de
diversidade e abundância entre o macrozoobentos (isópodos, anfípodos, tanaidáceos,
cumáceos), caracterizam-se pela presença do marsúpio (bolsas que carregam massas de
ovos no abdome das fêmeas). No interior destes ovos desenvolvem-se os embriões, que
darão origem aos juvenis no momento da eclosão.
Como exemplos de desenvolvimento direto obtidos a partir de posturas aderidas ao
substrato, como ocorre entre os gastrópodos do gênero Stramonita (Thais). Thais lamellosa
(Pacífico Norte) possui sexos separados e fertilização por cópula. Durante a época de
reprodução formam-se grupos de 50 a 1000 gastrópodes. Cada fêmea põe cerca de 1000
ovos, em 40 a 60 cápsulas, que ficam aderidas ao substrato numa mesma área com a
postura de outras fêmeas do mesmo grupo. Destas posturas saem os juvenis do gastrópodo.
Exemplo de desenvolvimento que ocorre dentro da fêmea, até o ponto em que o
embrião sai como juvenil, é encontrado entre alguns gastrópodos prosobranquios, que retém
seus ovos no interior da concha. Em Littorina saxatilis o desenvolvimento dos embriões ocorre
dentro de ovos ricos em albumina, mantidos numa parte diferenciada do oviduto, que irá
conter embriões em vários estágios de desenvolvimento, dos quais sairão juvenis com o
desenvolvimento completo.
Crinóides Commatulidos na Antártida (a maioria tem larvas pelágicas em menores
latitudes) depositam seus ovos numa espécie de saco numa abertura da parede do corpo,
que apresenta uma abertura externa de onde saem os juvenis do crinóide, após completar
seu desenvolvimento. Este saco localiza-se na base das pínulas próximo das gônadas nos
braços do crinóide. A produção de ovos é menor em relação à de outros commatulidos com
desenvolvimento larval pelágico. Devido a produção de uma grande quantidade de vitelo,
existe um custo energético bastante elevado para a manutenção do embrião até seu completo
desenvolvimento.
Desenvolvimento Indireto
Os macroinvertebrados bentônicos podem apresentar desenvolvimento com larvas
lecitotróficas ou larvas planctotróficas pelágicas.
As larvas lecitotróficas se alimentam de ovos ricos em vitelo e apresentam uma fase
planctônica relativamente curta (de horas a poucos dias). O trato digestivo ainda não está
desenvolvido e as reservas de vitelo permitem a larva viver no plâncton por um curto período
de tempo. Como vantagens, estas larvas podem usar as correntes para a dispersão e não
dependem do alimento no plâncton. A desvantagem é o tempo relativamente curto para
dispersão e para achar um local propício para o assentamento. Este tipo de desenvolvimento
é encontrado em equinodermas, poliquetas, poríferos, anthozoarios, bivalvos, nemertineos,
3
gastrópodos, polyplacophoros. Ocorrem em cerca de 10% das espécies de invertebrados.
Este tipo de desenvolvimento é comum no bentos profundo.
As larvas planctotróficas pelágicas podem permanecer no plâncton de 2 a 4 semanas
no verão e 1 a 3 meses no inverno. Originam-se de ovos pequenos, pobres em vitelo. Estas
larvas apresentam estruturas para alimentação e digestão, pois precisam consumir desde
bactérias até detritos, fito ou microzooplâncton para completar seu desenvolvimento. Algumas
larvas tropicais sobrevivem vários meses no plâncton e podem percorrer distâncias que
permitem atravessar os oceanos (larvas teleplânicas). A falta de um substrato adequado
resulta num retardamento da metamorfose, que pode resultar na degeneração e na morte da
larva. A longa permanência no plâncton está geneticamente regulada através de uma série de
estágios larvais. Existe uma grande variedade morfológica entre as larvas, mas um grande
número de espécies apresenta bandas ciliadas. Estas bandas auxiliam no posicionamento da
larva e servem, principalmente, para a captura de partículas ou organismos em suspensão.
As larvas de grupos numericamente representativos do macrozoobentos irão ocorrer
em abundância na coluna d´água, como integrantes do meroplâncton. Entre essas, as fases
Nauplius, Zoé e Megalopa dos crustáceos decápodos, as fases nauplius e cypris dos
cirripédios, as etapas de veliger (moluscos) e pediveliger dos bivalvos, trocófora de poliquetas
e as larvas pluteus dos ouriços irregulares, as equinopluteus dos ouriços regulares, as
auricularia das holoturias, as ophiopluteus dos ofiuróides e as braquiolaria das estrelas, entre
outras.
Entre as vantagens, o desenvolvimento planctotrófico pelágico possibilita que as
fêmeas já no primeiro ano de vida possam colocar em abundância ovos pequenos, pobres em
vitelo. Também a longa permanência no plâncton permite as larvas uma ampla dispersão e a
ocupação de um amplo espectro de hábitats, de acordo com as exigências e tolerâncias de
cada uma das espécies. Estas larvas são as mais capacitadas para invadir novas áreas.
Entre as desvantagens está a dependência das condições encontradas no plâncton,
que podem determinar uma mortalidade elevada e a ocorrência dos denominados “anos bons
e maus de recrutamento”. As flutuações no recrutamento são maiores entre as larvas de
longa permanência, em relação às curta vida no plâncton. O modo de desenvolvimento
planctotrófico pelágico é encontrado em cerca de 70% dos macroinvertebrados bentônicos.
Qual a vantagem da dispersão? As populações, que permanecem num mesmo local
sem dispersão, sofrem uma maior probabilidade de serem extintas devido a catástrofes locais.
A função da dispersão não é apenas a de afastar do banco de adultos (menos competição por
espaço e alimento), mas também colonizar hábitats potenciais a grandes distâncias. A
dispersão para hábitats variados, em longas distâncias, protege as populações dos riscos de
catástrofes que eliminam a espécie. Paleontologistas descobriram que espécies de moluscos
com larvas pelágicas de longa duração tiveram um menor grau de extinção. A ampla
dispersão e a possibilidade de ocupar hábitats em distintas áreas, de um modo geral,
asseguram boas chances de sobrevivência para as espécies frente a catástrofes locais.
Também a dispersão e a chance de ocupar uma série de ambientes em algum deles pode
produzir um bom recrutamento.
O recrutamento é a combinação da seleção do hábitat com a sobrevivência durante o
desenvolvimento antes, durante e após o assentamento. O que se observa recrutado num
determinado local resulta das larvas que sobreviveram à mortalidade na coluna d’água,
conseguiram selecionar o hábitat, escaparam da predação no momento do assentamento
pelos suspensívoros e após o assentamento pelos comedores de depósito, raspadores e
predadores da macro e meiofauna. Atualmente o termo assentamento é mais utilizado para
os eventos relacionados com a passagem da vida pelágica para a vida bentônica. Enquanto
que o recrutamento, implica num lapso de tempo posterior ao do assentamento. Os recrutas
seriam então, os novos indivíduos recém assentados, que teriam sobrevivido a mortalidade
logo após o assentamento (predação pela macro e meiofauna e consumo pelos comedores
de depósito e raspadores) tendo já atingido um determinado grau de desenvolvimento no
meio bentônico.
4
1)
2)
3)
4)
5)
6)
As vantagens do desenvolvimento pelágico podem ser sumarizadas por:
Recrutamento de um amplo espectro de hábitats
A expansão da distribuição geográfica
A colonização de novos hábitats
A exploração dos blooms de fitoplâncton.
Como forma de minimizar a competição e a predação próximo ao estoque parental.
A ampliação da distribuição diminui os riscos de mortalidades catastróficas, que ocorram
numa escala regional.
As Exigências para o Sucesso do Desenvolvimento.
Será dada ênfase ao desenvolvimento larval planctrófico pelágico, que atinge cerca de
70% dos macroinvertebrados bentônicos e apresenta a maior mortalidade.
a) Condições Alimentares - a maioria das larvas necessita ingerir fitoplâncton com tamanho
entre 2 a 30u, sendo a exigência do alimento, proporcionalmente, de 5 a 10x maior do que a
dos adultos. Certos bivalvos, por exemplo, só liberam suas larvas após um bloom de
fitoplâncton. Devem ser distinguidas as condições alimentares: 1- que matam as larvas pela
fome; 2- que permitem as larvas sobreviver, porém prejudicam seu desenvolvimento; 3- que
estimulam o desenvolvimento larval. A disponibilidade do alimento influencia o sucesso das
larvas de forma direta (causando mortalidade) ou indireta (atrasando o desenvolvimento). Um
trabalho clássico, desenvolvido por Bousfield que acompanhou a relação entre o fitoplâncton e
o sucesso das larvas do cirripédio Balanus balanoides, mostrou que a sobrevivência larval foi
diretamente relacionada com a quantidade de alimento no meio ambiente.
b) Temperatura - os limites de tolerância são mais estreitos nas fases iniciais de
desenvolvimento. Para os caranguejos Crangon crangon, foi observado que em invernos
muito frios morreram os ovos nos pleópodos, mas os adultos sobreviveram. Muitas espécies
necessitam migrar para reproduzir a fim de que as larvas tenham maiores chances d
sobrevivência. Sabe-se que a temperatura pode prolongar ou acelerar o desenvolvimento
larval. Mudanças repentinas de temperatura podem matar os invertebrados, mas estas são
pouco comuns no meio marinho. Deve ser considerado, que em geral existe uma temperatura
para maturar, outra para emitir os gametas e que esta última é normalmente a exigida para o
desenvolvimento das larvas. A temperatura é um forte condicionante da distribuição das
espécies, dos seus ciclos reprodutivos e um importante fator indireto da mortalidade larval.
3- As características do Substrato – a maioria das larvas tem uma série de exigências para o
assentamento. Espécies de poliquetas podem retardar a metamorfose em busca de substrato.
Em algumas espécies a trocófora pode diferenciar o sedimento no momento de assentar,
influindo o tamanho do grão e a presença de microorganismos (bactérias). O poliqueta
Ophelia bicornis exige determinadas bactérias para assentar. O poliqueta Hydroides, que vive
em colônias de pequenos tubos calcáreos, tem dois tipos de larvas, uma que assenta junto ao
banco de adultos e uma outra pioneira, que busca espaços vazios no substrato. Ostras e
cirripédios também são atraídos pelos adultos no fundo. Entre as vantagens do gregarismo
estaria a proteção oferecida pelos microhábitats formados pelo banco de adultos, enquanto
que a competição e a predação pelos organismos já assentados seria uma desvantagem. A
atração para o assentamento pode ser: química, pela artropodina em cirripédios ou um
peptideo solúvel (aminoácido) em ostras; mecânica (macroalgas), como para os mexilhões
em costões ou poliquetas que selecionam pela granulometria do sedimento. A veliger de
Mytilus edulis busca algas vermelhas filamentosas para assentar, mas também assenta sobre
cabos fibrosos com estrutura similar.
5
4- Transporte por Correntes - Para as espécies que vivem no intermareal o transporte por
correntes para o mar aberto pode ser um importante fator de mortalidade. Correntes paralelas
ou obliquas a linha de costa, em regiões costeiras, favorecem a dispersão e ampliam a
distribuição das espécies. A heterogeneidade da linha de costa pode cortar a distribuição e/ou
promover o acúmulo de larvas, com reflexos no recrutamento das espécies costeiras. Na
europa o vento predominante no verão é o Oeste, que transporta as larvas para lugares
costeiros. Na Guiné africana predomina o vento Nordeste que leva as larvas para longe da
costa. Nesta região, cerca de 68% das espécies de gastrópodos costeiros, estudados pelo
pesquisador dinamarquês G. Thorson, apresentavam desenvolvimento direto .
5- Mortalidade por predação – As larvas de invertebrados são pequenas, em geral menores
do que 250u e sofrem o ataque de muitos predadores. A predação na coluna d'água é
provavelmente o maior fator de mortalidade para as larvas do macrozoobentos. Na coluna
d'água as larvas sofrem o ataque de peixes zooplantófagos (p.ex. Clupeideos); do
zooplâncton predador como Medusas, Ctenophoros, Chaetognathus, e do meroplâncton
(larvas de crustáceos decápodos e de algumas espécies de poliquetas).
A captura no momento do assentamento é outra importante fonte de mortalidade das
larvas do macrozoobentos. G. Thorson considerou que suspensívoros como Mytilus edulis,
que filtram 1,5 litros de água por hora, podem capturar até 100.000 larvas de bivalvos em 24
horas no Kattegat (Dinamarca). Em fundos moles, S. Woodin observou que a infauna não
conseguia manter densidades elevadas, devido a fltração de suas larvas, em locais ocupados
por bancos de bivalvos suspensívoros.
Após o assentamento em fundos duros, as larvas sofrem a ação dos raspadores
(gastrópodos pateliformes, ouriços, quitons). Em fundos moles os exemplares recém
assentados são consumidos por comedores de depósito e macrófagos predadores, estrelas,
ofiuros, caranguejos, camarões, entre outros. Também exemplares da meiofauna, Turbellaria,
Nematodos, Copépodos Harpaticoides, predam sobre os recrutamentos do macrozoobentos.
Dinâmica da Colonização do Substrato
As associações de macroinvertebrados de fundos moles, na maioria das vezes, se
distribuem em forma de manchas, que apresentam flutuações temporais na composição e
abundância. Estas associações são mantidas através dos recrutamentos via assentamento
larval ou do desenvolvimento direto. A capacidade de certas larvas de retardar a
metamorfose e poder escolher o substrato para o assentamento indica que o recrutamento,
geralmente, não é aleatório.
A distribuição em manchas e sua variabilidade temporal são influenciadas, além do
recrutamento, pela ocorrência de continuas perturbações provocadas por fatores físicos
(p.e. a hidrodinâmica) e biológicos (predação-bioperturbação), que eliminam os organismos
em áreas restritas. Estas áreas são ocupadas por novos indivíduos no processo de
recolonização, o qual varia temporalmente em decorrência de complexas interações
biológicas envolvendo as distintas estratégias de vida dos organismos (espécies
oportunistas x em equilíbrio), a sobrevivência dos mesmos durante suas fases larvais e
durante o assentamento, até o efetivo recrutamento. Destas complexas interações resultam
que diferentes áreas apresentam associações com composição e abundância distinta, em
função do tempo decorrido desde a última perturbação, da época do ano e do estilo de vida
dos organismos envolvidos.
Literatura Complementar
Knox, G. A 2001. The Ecology of Seashores. CRC Press, Boca Raton, 557p.
Lalli, C.M. & Parsons, T.R. 1993. Biological Oceanography: an Introduction. Butterworth &
Heinemann, Oxford, 301p.
Levinton, J.S. 1995. Marine Biology. Function, Biodiversity, Ecology. Oxford University
Press, New York, 420p.
6
Pereira, R.C. & Soares-Gomes, A. 2009. Biologia Marinha. Segunda Edição. Editora
Interciência, Rio de Janeiro, 631p.
Ruppert, E.E.; Fox, R.S. & Barnes, R.D. 2005. Zoologia dos Invertebrados, Sétima Edição,
Editora Roca Ltda., São Paulo, Brasil, 1145p.
7