1
VISÃO
Vemos no meio ambiente um grande número de objetos tridimensionais. Esses objetos podem ser
descritos como estacionários ou móveis, ou apresentando uma disposição particular no espaço. Eles
podem variar quanto à cor, ao brilho, tamanho e à forma. Os objetos percebidos através da visão podem
ter um significado especial para nós ou podem ser destituídos de importância; podem despertar uma
emoção ou não desencadear nenhuma reação subjetiva. Os fisiologistas habituaram-se a dizer que
percepção visual ocorre porque a imagem do ambiente é projetada sobre a retina. Esta imagem
desencadeia certos processos de detecção de sinais e de elaboração nos receptores e das células nervosas
de ordem superior, resultando finalmente na percepção em nível “consciente”. Entretanto, todos nós
sabemos, através dos sonhos que a percepção visual pode ocorrer na ausência de imagens retinianas.
1. Receptores Visuais
Os receptores visuais, ou receptores eletromagnéticos ou fotorreceptores são os cones e os
bastonetes, localizados na retina. Ali se forma uma imagem menor e invertida, e a luz determina a
transmissão de impulsos nervosos através das vias ópticas até o córtex cerebral (área occipital), onde se
tem a sensação visual.
Os receptores visuais são sensíveis a somente uma reduzida porção do imenso espectro de
radiação eletromagnética que denominamos luz. Os comprimentos de onda capazes de estimular os
receptores visuais (espectro visível) estão entre 400 a 700 nm, e a luz de diferentes comprimentos de onda
nesta faixa está associada com diferentes sensações de cor. Ex: Azul: 450nm; verde: 535 nm;
vermelho:700nm
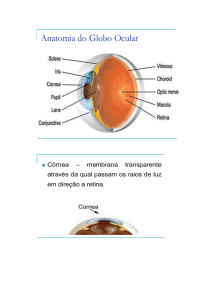
2. Estrutura do Olho (fig. 9.6)
Conjuntiva: de localização anterior, é um tecido conjuntivo transparente, infiltrado por pequenos
vasos sanguíneos e que funde com a córnea;
Córnea: estrutura convexa, transparente e avascular;
Íris: localizada atrás da córnea, é uma estrutura muscular pigmentada. Possui dois tipos de músculo
liso: radial e circular, cuja contração é controlada pelo sistema nervoso autônomo. A contração destes
músculos controla o diâmetro de um orifício no centro da íris a pupila, de acordo com a intensidade de
luz;
Câmara Anterior: entre a córnea e a íris, está cheia de líquido, o humor aquoso, incolor, formado de
plasma filtrado; serve como meio nutritivo aos tecidos avasculares do olho;
2
Cristalino (inglês- lens): localizado atrás da íris, é uma lente circular convexa, elástica, que fica
suspensa por seus bordos pelas fibras zonulares ligadas ao músculo ciliar;
Humor Vítreo: de consistência gelatinosa, preenche toda a região posterior do globo ocular. Este
líquido e o humor aquoso mantêm a pressão intraocular. O aumento na sua produção ou a deficiência
na sua drenagem provoca o glaucoma que consiste na elevação da pressão intraocular com perigo de
lesão dos fotorreceptores;
As três camadas que delimitam o globo ocular são, de fora para dentro:
Esclerótica: espessa, formada de tecido conjuntivo denso, tem a função de proteção e sustentação das
camadas internas;
Coróide: contém grandes quantidades de melanina (pigmento escuro) que impede a reflexão da luz por
todo o globo ocular;
Retina: é a camada onde estão os fotorreceptores e demais células nervosas, responsáveis pela
transdução e transmissão dos estímulos visuais, respectivamente.
Papila: também chamada de ponto cego. É a região da retina onde não existem fotorreceptores, pois é
o local de saída das fibras nervosas que compõem o nervo óptico. Se a imagem incidir sobre esse ponto
nós não a enxergaremos.
3. Alguns Princípios de Óptica
3.1. Refração
É o desvio dos raios de luz em uma superfície angulada.
a) Refração em Lente Convexa (fig. 49.2)
Lente convexa: bordos delgados e centro
espesso.
Quando os raios luminosos paralelos
penetram em uma lente convexa, aqueles raios
que passam pelo centro não sofrem refração;
porém, os raios luminosos que atingem as
bordas da lente inclinam-se mais e mais em
direção ao centro. Assim, raios paralelos
penetrando em lente convexa, dirigem-se para
um único foco puntiforme, a uma certa
distância da lente.
Ponto focal: ponto onde todos os raios que passam por uma lente se convergem.
Distância focal: é a distância entre uma lente convexa e o ponto focal. Quanto maior a curvatura
da lente convexa, menor é a distância focal.
b) Refração em Lente Côncava
Lente côncava: bordos espessos, curvandose para o centro delgado.
Os raios que penetram exatamente no
centro da lente não sofrem refração. Os raios que
passam pelos bordos da lente encontram uma
superfície progressivamente mais angulada,
penetrando primeiro na lente do que aqueles raios
que atravessam o centro. Isto fará os raios de luz
divergirem daqueles que passam pelo centro da
lente.
3
c) Medida do poder de refração de uma lente - Dioptria
Quanto mais uma lente desvia os raios luminosos, maior é o seu “poder de refração”. Este poder
de refração é medido em termos de dioptrias (D).
O poder de refração de uma lente convexa é D= ________1 m__________
distância focal
Ex.: uma lente que converge os raios para uma distância de 0,5 m tem um poder de refração= + 2
D, isto é 1 / 0,5 = 2 .
No olho normal, a distância entre a córnea e a retina é de 17 mm (0,17 cm), portanto, o poder de
refração do sistema óptico deverá ser de + 59 D, pois, 100 / 0,17 = 59.
O poder de refração de lentes côncavas não pode ser expresso em termos de sua distância focal,
porque os raios luminosos divergem, ao invés de se focalizarem num ponto. Assim, o poder de uma lente
côncava é formulado em termos de sua capacidade em divergir os raios luminosos em comparação com a
capacidade de lentes convexas em convergir-lhos. Isto é, se uma lente côncava diverge raios luminosos
com a mesma intensidade que uma lente convexa de + 1 D os converge, diz-se que a lente côncava tem
uma potência dióptrica de - 1 D.
Observe que as lentes côncavas podem “neutralizar” o poder de refração das lentes convexas.
Assim, a colocação de uma lente côncava de - 1 D imediatamente à frente de uma lente convexa de + 1D
resulta num sistema de lentes com poder de refração igual a zero.
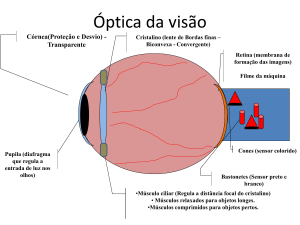
4. Óptica do Olho
4.1. Sistema Óptico do Olho
O sistema óptico do olho ou aparelho dióptrico consta de um sistema complicado de lentes
(córnea, cristalino), o qual não é perfeitamente centrado, e que projeta sobre a retina uma imagem
invertida do ambiente, de tamanho reduzido.
4.2. Processos de Adaptação do Aparelho Dióptrico
As formas da córnea e do cristalino e o comprimento do globo ocular determinam o ponto onde os
raios luminosos se reencontram. Portanto, os raios luminosos de objetos próximos ao olho atingem a
córnea em maiores ângulos (são mais divergentes) e devem ser mais desviados para convergirem sobre a
retina. Embora a córnea desempenhe quantitativamente a maior parte na focalização da imagem visual
sobre a retina, todos os ajustes para distância são feitos pelo cristalino. Estas alterações são denominadas
acomodação à distância.
4
Existem duas maneiras
para
conseguir
isto:
o
deslocamento do cristalino em
relação à retina ou o aumento
da capacidade de refração (o
grau
de
angulação)
do
cristalino. Anfíbios e répteis
utilizam a primeira maneira; já
o cristalino do olho humano
tem a capacidade de alterar o
seu grau de angulação,
podendo alterar a capacidade
de refração do olho de 59 para
70 D, numa criança. Para que
isto se realize, a forma do
cristalino é alterada de uma
lente moderadamente convexa
para uma lente muito convexa,
isto é possível devido à
natureza elástica do cristalino.
a) Reflexo de Acomodação à Distância
Na fig. acima, podemos observar que o cristalino está suspenso por um conjunto de fibras
denominadas de fibras zonulares que, por sua vez, estão ligadas ao músculo ciliar.
Durante o estado de acomodação para longe, onde o cristalino não precisa de um alto poder de
refração e o músculo ciliar encontra-se relaxado, afastando-se do cristalino, provocando com isso, uma
tensão nas fibras zonulares que, por sua vez, tracionam o cristalino, deixando-o menos angulado (ver
figura na próxima página).
Na acomodação para perto, onde o cristalino necessita estar com um alto poder de refração, o
músculo ciliar encontra-se contraído, aproximando-se do cristalino, deixando as fibras zonulares
frouxas, onde permite que as fibras elásticas do cristalino retornem à sua posição de repouso (cristalino
com maior curvatura). (ver figura na próxima página).
O processo de contração e relaxamento do músculo ciliar está sob o controle do sistema nervoso
autônomo, divisão parassimpática e simpática, respectivamente.
PRESBIOPIA: É a incapacidade de enxergar objetos próximos advinda com a idade. À medida que a
pessoa envelhece, o cristalino perde sua natureza elástica e torna-se uma massa relativamente rígida, em
parte devido à progressiva desnaturação de suas proteínas. Assim, a capacidade do cristalino em assumir
forma esférica diminui progressivamente, e o poder de acomodação diminui de cerca de 14 dioptrias na
criança pequenas, para menos de 2 dioptrias na idade de 45 a 50 anos. Daí em diante, o cristalino pode ser
considerado como sendo quase que totalmente não-acomodativo.
Uma vez que o indivíduo tenha alcançado o estado de perda da elasticidade máxima do cristalino,
cada olho permanece focalizado permanentemente numa distância quase constante; essa distância
depende das características físicas dos olhos de cada indivíduo. Obviamente, os olhos não podem mais se
acomodar, tanto para longe como para perto. Portanto, para o indivíduo idoso ter uma visão nítida, tanto
para longe quanto para perto, deve usar lentes bifocais, com o segmento superior normalmente focalizado
para visão à distância e o inferior para visão próxima.
5
b) Reflexo de Acomodação à Luz - Reflexo Pupilar (fig. na próxima página)
A pupila é uma pequena abertura localizada no centro da íris, cujo diâmetro é controlado pela
contração da musculatura lisa da íris. A pupila humana pode variar desde o diâmetro de 1,5 mm até o
máximo de 8 mm.
b.1.) Constrição da Pupila (Miose): quando a luz que penetra na pupila é intensa, ocorre um reflexo
através do sistema nervoso autônomo parassimpático que inerva as fibras musculares circulares da íris,
cuja contração diminui o diâmetro pupilar.
b.2.) Dilatação da Pupila (Midríase): quando a luz que penetra na pupila é pouco intensa, ocorre um
reflexo através do sistema simpático que inerva as fibras musculares radiais da íris, cuja contração
aumenta o diâmetro pupilar.
4.3. Erros de Refração
Emetropia (fig. 10.35a): olho normal. Quando o ponto focal incide exatamente sobre a retina
como o cristalino distendido, estando o indivíduo observando um objeto distante.
a) Hipermetropia (fig. 10.35b): ou visão para longe, ocorre normalmente devido a um globo ocular
demasiado pequeno ou a um sistema de lentes fraco demais quando o músculo ciliar está completamente
relaxado. O ponto focal localiza-se atrás da retina. Assim, o indivíduo hipermétrope, quando observa um
objeto distante, já se utiliza do mecanismo de acomodação do cristalino, contraindo os músculos ciliares.
Porém, com a aproximação do objeto, atinge-se um grau máximo de contração dos músculos ciliares.
Correção: uso de lentes convexas (positivas) (fig. 10.35c).
6
b) Miopia (fig. 10.35d): ou visão para perto, quando os músculos ciliares estão relaxados, o ponto focal
encontra-se na frente da retina. Isso se deve em geral, a um globo ocular muito alongado, porém pode,
ocasionalmente, resultar de um poder excessivo do sistema de lentes do olho. Não existe nenhum
mecanismo pelo qual o olho possa diminuir a potência do cristalino além daquela que existe quando o
músculo ciliar está relaxado, portanto, o indivíduo míope não tem nenhum mecanismo pelo qual possa
focalizar constantemente objetos distantes com nitidez em sua retina. Entretanto, quando um objeto lhe
aproxima do olho, torna-se finalmente próximo o bastante para que sua imagem seja focalizada sobre a
retina. Então, quando o objeto chega ainda mais perto, o indivíduo pode usar seu mecanismo de
acomodação para manter a imagem focalizada sobre a retina.
Correção: uso de lentes côncavas (negativas) (fig. 10.35e).
c) Astigmatismo (fig. ao lado): é um erro de refração causado,
geralmente, por córnea de forma oblonga(oval), ou em casos raros, de
cristalino oblongo. Uma lente de superfície oval, onde um lado possui
uma curvatura maior do que o outro, faz com que os raios de luz que
incidem nas partes periféricas da lente em um plano não são inclinados
quanto os raios que incidem nas partes periféricas do outro plano,
formando-se assim, vários pontos focais na retina, e não um único
ponto.
Correção: lentes cilíndricas.
7
Fig. 10.35: Principais erros de refração e suas
correções.
5. Função Receptora e Neural da Retina
A retina é a região fotossensível do olho que contém os cones, principais responsáveis pela visão
cromática (cores) e os bastonetes, principais responsáveis pela visão no escuro (fig.9.13). Quando cones e
bastonetes recebem um estímulo luminoso, os
sinais são transmitidos através de neurônios
sucessivos na própria retina e, finalmente, pelas
fibras do nervo óptico até o córtex cerebral (área
occipital).
5.1. Estrutura da Retina (fig. ao lado)
A retina é formada pelos fotorreceptores (cones
e bastonetes) e por mais quatro tipos diferentes
de células nervosas: bipolares, horizontais,
amácrinas e ganglionares.
A parte da retina que é sensível à luz é
formada pelos bastonetes (aproximadamente 120
milhões) e pelos cones (aproximadamente 6
milhões). Na fóvea, a retina é formada
exclusivamente por cones e, como veremos
8
posteriormente, é o local da retina de maior acuidade visual.
Cada fotorreceptor possui um segmento externo delicado, formado por cerca de mil discos
membranosos (bastonetes) ou invaginações da membrana (cones). As moléculas dos pigmentos visuais
encontram-se distribuídas em meio a estas estruturas membranosas.
Bastonetes e cones comunicam-se através de sinapses com as células bipolares e horizontais. As
células bipolares transmitem os sinais recebidos dos fotorreceptores à camada de células ganglionares e
amácrinas. As células horizontais e amácrinas destinam-se à transmissão “horizontal” dos sinais,
perpendicularmente à direção principal do fluxo de sinais (receptores células bipolares células
ganglionares SNC).
Note que a luz deve passar por todas essas camadas celulares para depois chegar nos
fotorreceptores. Porém, na fóvea (fig. abaixo) essas camadas mais internas estão ausentes, de modo que a
luz excita diretamente os fotorreceptores (cones), sendo este um dos motivos que faz a fóvea ser o local
de maior acuidade visual. O outro motivo para essa maior acuidade visual resulta do fato que na fóvea não
ocorre convergência de sinais neurais, ou seja, uma célula bipolar faz contato sináptico com apenas um
cone, e uma célula ganglionar é
estimulada por apenas uma célula bipolar
(ver fig. 10.37). Desta forma, cada célula
ganglionar recebe informação visual de
uma área da retina que corresponde ao
diâmetro de um cone (por volta de 2 m),
o que permite um detalhamento mais
apurado da imagem que se está
observando. Por outro lado, nas demais
regiões
da
retina,
ocorre
uma
convergência dos sinais, como podemos
observar na fig. 10.37, uma única célula
bipolar recebe informações de um grande
número de bastonetes, correspondendo a
uma área da retina de aproximadamente 1
mm2, o que dificulta o processamento mais detalhado da imagem observada.
9
5.2. Fotorreceptores (fig. 9.13)
A conversão da radiação eletromagnética em sinais neurais ocorre
nos 126 milhões de fotorreceptores na parte posterior da retina humana.
Cada fotorreceptor apresenta quatro regiões: um segmento externo e um
interno, um corpo celular e um terminal sináptico. O segmento externo
contém uma pilha de discos membranosos. Fotopigmentos sensíveis à luz,
localizados nas membranas desses discos, absorvem luz, determinando,
dessa forma, alterações no potencial de membrana do fotorreceptor (como
será discutido adiante). A fig. 9.13 mostra os dois tipos de fotorreceptores
encontrados na retina, os quais são facilmente distinguíveis pelo formato
de seus segmentos externos. Os fotorreceptores do tipo bastonete
apresentam um longo e cilíndrico segmento externo contendo muitos
discos. Os fotorreceptores do tipo cone apresentam um segmento externo
mais curto e que gradualmente diminui de espessura, contendo
relativamente poucos discos membranosos.
As diferenças estruturais entre bastonetes e cones estão relacionadas
a importantes diferenças funcionais. Por exemplo, o maior número de
discos e a maior concentração de fotopigmentos nos bastonetes os tornam
1.000 vezes mais sensíveis à luz que os cones. De fato, em um ambiente
com iluminação noturna, ou seja, em condições escotópicas, apenas os
bastonetes contribuem para a visão. Por outro lado, em ambientes com
iluminação diurna, ou seja, em condições fotópicas, os cones é que
realizam a maior parte do trabalho.
Bastonestes e cones diferem também em outros aspectos. Todos os bastonetes contêm o mesmo
fotopigmento, mas há três tipos de cones, cada qual com um diferente pigmento. Essas variações entre os
pigmentos fazem com que os diferentes cones sejam sensíveis a diferentes comprimentos de onda da luz.
Como veremos logo mais, os cones são os principais responsáveis por nossa capacidade de ver cores.
5.3. Fotoquímica da visão
Tanto cones como bastonetes contêm
substâncias fotossensíveis que se decompõem
pela exposição à luz. A substância química do
bastonete é a rodopsina e a substância química
dos cones é a iodopsina. Estudaremos a
fotoquímica da rodopsina, sabendo que devem
ser válidos os mesmos princípios para a
fotoquímica da iodopsina.
O segmento externo do bastonete
contém grandes quantidades de rodopsina.
Esta substância resulta da combinação de uma
proteína: opsina com o 11-cis-retinal (fig.
9.18). Quando a energia luminosa é absorvida
pela rodopsina, esta imediatamente começa a
se decompor. Isso acontece porque o 11-cisretinal transforma-se em todo-trans-retinal,
que tem uma estrutura diferente da forma cis (fig. 9.18). Como resultado, os sítios ativos do retinal não
mais se ajustam aos sítios da opsina, e o trans-retinal começa a se desligar da molécula de opsina,
formando combinações progressivamente mais frouxas destas duas moléculas até serem inteiramente
individualizadas.
Na ausência da luz, o 11-trans-retinal retorna à sua forma cis, isso na presença da enzima
isomerase, podendo combinar-se com a opsina, ressintetizando a rodopsina novamente.
A vitamina A é o maior precursor do 11-cis-retinal e está armazenada na camada pigmentar.
10
Carência de vitamina A cegueira noturna.
Assim, no escuro grande quantidade de rodopsina. No claro baixa quantidade de rodopsina.
5.4. Origem do Potencial Receptor
O segmento interno do fotorreceptor bombeia Na+ continuamente para fora da célula, criando
assim, um potencial negativo no
interior da mesma. Entretanto, na
condição de repouso (no escuro), a
membrana do segmento externo é
muito permeável ao Na+. Sendo
assim, o Na+ que é bombeado para
fora pelo segmento interno, através da
bomba de Na+/K+, flui continuamente
de volta para o interior da célula,
neutralizando a negatividade (fig.
9.17a). Em condições normais,
quando o bastonete não é excitado,
ocorre negatividade reduzida no
interior do bastonete, normalmente de
cerca de -30mV.
Porém, quando a rodopsina é
degradada pela luz, a permeabilidade
do segmento externo do bastonete ao
Na+ é reduzida. Isto diminui a entrada
de Na+, resultando num aumento da
eletronegatividade no interior da
célula, ou seja, na presença da luz o
fotorreceptor é hiperpolarizado (fig
9.17b). Em 1985, uma equipe de
cientistas russos descobriu que esses
canais de sódio têm sua abertura
estimulada pelo segundo mensageiro
intracelular
GMPc
(guanosina
monofosfato cíclico). Evidentemente,
o GMPc é produzido continuamente no fotorreceptor pela enzima gualinato ciclase, mantendo os canais
de Na+ abertos. A luz reduz a quantidade de GMPc, o que determina o fechamento dos canais de Na+, e
torna o potencial de membrana mais negativo (fig. 9.17b). Desta forma, os fotorreceptores são
hiperpolarizados em resposta à luz.
A resposta hiperpolarizante à luz ocorre porque a opsina, após se desligar da porção retinal, tornase ativa (fig.11.7). A opsina ativada, por sua vez, estimula uma proteína G que está a ela acoplada. Esta
proteína G, chamada de transducina, irá ativar outra enzima denominada fosfodiesterase do GMPc (PDE).
A PDE hidrolisa o GMPc que era o responsável em manter os canais de Na+ abertos durante o escuro.
Assim, na presença de luz, ocorre o fechamento dos canais de Na+, devido a redução de GMPc e o
fotorreceptor hiperpolariza.
Uma das características importantes dessa complexa cascata bioquímica iniciada pela captura de
um fóton é que ela permite uma considerável amplificação do sinal. Estima-se que uma única molécula de
rodopsina ativada pela luz possa ativar 800 moléculas de transducina. Embora cada molécula de
transducina ative apenas uma molécula de PDE, cada uma delas, por sua vez, é capaz de catalisar a
hidrólise de até seis moléculas de GMPc. Como resultado, a absorção de um único fóton por uma
molécula de rodopsina resulta no fechamento de aproximadamente 200 canais iônicos.
11
A hiperpolarização dos fotorreceptores faz com que estes deixem de secretar o neurotransmissor
que produz inibição nas células bipolares e horizontais da retina. As células bipolares, quando não são
inibidas pelo fotorreceptor, excitam as células ganglionares, cujos os axônios formam o nervo óptico que
leva a informação visual ao SNC.
As células horizontais inibem as células bipolares vizinhas às estimuladas. Esta inibição lateral
tem como finalidade dar maior nitidez e ajudar a detectar contrastes no cenário visual.
Fig. 11.7: Detalhes da fototransdução em receptores do tipo bastonete. O mesmo princípio vale para os cones, exceto
que a proteína opsina difere um pouco na sua composição de aminoácidos.
5.5. O Processamento na Retina - Campos Receptivos das Células Bipolares
Como vimos, os fotorreceptores estão despolarizados no escuro e se tornam hiperpolarizados com
a luz. Temos assim, uma situação contrária àquela que seria a mais esperada: na verdade, os
fotorreceptores liberam menos neurotransmissores na presença da luz do que no escuro. Podemos
conciliar esse aparente paradoxo, entretanto, se aceitarmos o ponto de vista de que é o escuro, e não a luz,
o estímulo preferido pelo fotorreceptor. Assim sendo, quando uma sombra passa sobre um fotorreceptor,
ele responde sofrendo uma despolarização e liberando glutamato.
Na camada plexiforme externa da retina, cada fotorreceptor efetua contato sináptico com dois
tipos de células retinianas: as células bipolares e as células horizontais. Lembre-se que as células bipolares
estabelecem a via direta, desde os fotorreceptores até as células ganglionares; células horizontais, por sua
vez, fornecem informações lateralmente para a camada plexiforme externa, influenciando a atividade de
células bipolares e fotorreceptores vizinhos.
As células bipolares podem ser divididas em duas classes, com base em suas respostas ao
glutamato liberado pelos fotorreceptores. Células bipolares do tipo OFF apresentam canais de cátions
ativados por glutamato, que medeiam um clássico potencial excitatório pós-sináptico (PEPS), o qual
despolariza a membrana da célula bipolar pelo influxo de íons Ca++. Já as células bipolares do tipo ON
apresentam receptores acoplados a proteínas G e responde ao glutamato com uma hiperpolarização.
Observe que os nomes OFF e ON referem-se ao comportamento dessas células em presença da luz,
estando desligada (off; quando não há glutamato para ativar a célula bipolar) ou acesa (on; quando não há
glutamato para inibí-la e ela pode gerar por si só seus PAs, pois é uma célula auto-excitável).
12
Cada célula bipolar recebe aferências sinápticas diretas de um grupo de fotorreceptores. O número
de fotorreceptores nesse grupo varia desde um, no centro da fóvea, até milhares, na retina periférica. Além
dessas conexões diretas com os fotorreceptores, as células bipolares estão conectadas, via células
horizontais, a um anel circunscrito de fotorreceptores que cerca aquele grupo. O campo receptivo de uma
célula bipolar (ou de qualquer outra célula do sistema visual) é a área da retina onde, em resposta à ação
da luz, ocorre uma alteração do potencial de membrana da célula. O campo receptivo de uma célula
bipolar é constituído por duas porções: uma área circular da retina que proporciona aferências diretas do
fotorreceptor, chamada de centro do campo receptivo, e uma área de retina adjacente, que proporciona
aferências via células horizontais, chamada de periferia do campo receptivo (fig. 9.22).
A resposta do potencial de membrana de uma célula bipolar à luz no centro do campo receptivo é
oposta àquela promovida pela luz da periferia. Por exemplo, se a iluminação do centro causa
despolarização da célula bipolar (uma resposta ON), então a iluminação da periferia causará uma
hiperpolarização antagônica. Da mesma forma, se a célula for despolarizada quando houver uma mudança
de luz para escuro no centro de seu campo receptivo (uma resposta OFF), ela será hiperpolarizada pelo
mesmo estímulo escuro quando esse for aplicado à periferia. Assim sendo, diz-se que essas células
apresentam campos receptivos organizados em centro-periferia, com centro e periferia antagônicos
entre si. Esse antagonismo entre centro e periferia parece vir de uma interação complexa nos contatos
sinápticos de células horizontais, fotorreceptores e células bipolares. A organização dos campos
receptivos em centro-periferia passa das células bipolares para as ganglionares por meio das sinapses na
camada plexiforme interna.
A partir da organização dos campos receptivos das células ganglionares, podemos inferir que
nosso sistema visual está especializado para a detecção de variações espaciais locais e não da magnitude
absoluta da luz que cai sobre a retina. Assim sendo, a percepção de claridade ou de escuridão não é
absoluta, mas relativa. Isso é demonstrado pelos dois quadros emoldurado na figura 9.25.
Esse sistema de contraste nos campos receptivos na retina também é utilizado para cores. Dois
tipos de oposição são observados, vermelho versus verde e azul versus amarelo (verde+vermelho). Assim,
a cor percebida depende da atividade relativa das células ganglionares, cujos centros de campos receptivos
recebam sinais de cones para o vermelho, para o verde e para o azul.
13
Desta forma, parece que as células ganglionares fornecem um fluxo de informação para o encéfalo
que está envolvido na comparação espacial de três processos oponentes: claro versus escuro, vermelho
versus verde e azul versus amarelo.
5.6. Adaptação ao Claro e ao Escuro
A permanência de um indivíduo em ambiente intensamente iluminado, durante longo período de
tempo, faz com que grandes proporções de pigmentos fotossensíveis, tantos dos cones como dos
bastonetes, sejam reduzidas a retineno e a opsinas. Além disso, grande parte do retineno é convertida em
vitamina A. Por causa destes dois efeitos, as concentrações de substâncias químicas fotossensíveis sofrem
considerável redução, e a sensibilidade do olho à luz diminui grandemente. A isso chama-se adaptação ao
claro.
Por outro lado, se a pessoa permanecer na escuridão por muito tempo, praticamente todo o
retineno e as opsinas dos fotorreceptores são convertidos em pigmentos fotossensíveis. Além disso,
grandes quantidades de vitamina A são convertidas em retineno, que então é transformado em pigmentos
fotossensíveis adicionais. Devido a esses dois efeitos, os receptores visuais tornam-se gradualmente tão
sensíveis que quantidades mínimas de luz produzem excitação. Isto se denomina adaptação ao escuro.
5.7. Teoria Tricromática da Visão
As substâncias fotossensíveis dos cones têm, praticamente, a mesma composição química da dos
bastonetes. Somente a proteína é uma fotopsina, a porção retineno é a mesma.
Existem três tipos de opsinas, portanto, três tipos de cones. Cada opsina possui pequenas
modificações na sua cadeia polipeptídica que a torna seletivamente sensível a uma faixa de comprimento
de onda. Assim, nós temos um cone que é extremamente sensível à cor azul; um cone com máximo de
sensibilidade à cor verde e um cone com sensibilidade máxima à cor vermelha.
Observando a figura 50.9, pode-se ver
que a luz monocromática laranja, com um
comprimento de onde de 580 nm, estimula os
cones vermelhos até aproximadamente 99%,
enquanto estimula os cones verdes com valor
aproximado de 42% e não estimula os cones
azuis. Assim, as proporções de estimulação dos
três diferentes cones, neste caso, são
99(vermelho) : 42(verde) : 0(azul). O sistema
Fig. 10.9: Absorção de luz dos pigmentos visuais
nervoso interpreta este conjunto de proporções
como sendo a cor laranja.
Proporção do azul (450 nm) 0 : 0 : 97;
Proporção do verde (535 nm) 31 : 67 : 36;
Proporção do amarelo 83 : 83 : 0
14
A estimulação aproximadamente igual de todos os cones vermelhos, verdes e azuis dá à pessoa a
sensação visual do branco.
Daltonismo (cegueira para cores): quando um único grupo de cones receptores de cor está
ausente do olho, o indivíduo é incapaz de distinguir certas cores de outras. Como pode ser observado na
fig. 50.9, se os cones vermelhos estão ausentes, a luz com comprimentos de onde de 525 a 675 nm pode
estimular apenas os cones sensíveis ao verde, de modo que a razão entre a estimulação dos diferentes
cones não se altera à medida que a cor muda do verde para o espectro do vermelho. Portanto, dentro dessa
faixa de comprimento de onda, todas as cores parecem ser a mesma para esse indivíduo “cego para cores”.
Por outro lado, se os cones sensíveis ao verde estão ausentes, as cores na faixa do verde ao
vermelho podem estimular apenas os cones sensíveis ao vermelho, e o indivíduo também percebe apenas
uma cor dentro desses limites. Portanto, quando a um indivíduo faltam os cones dos tipos vermelho ou
verde, diz-se que ele é cego para “vermelho-verde”.
O indivíduo com falta de cones vermelhos é denominado protanópico; seu espectro visual total
está apreciavelmente diminuído no extremo dos grandes comprimentos de onda, devido à falta dos cones
vermelhos. O indivíduo cego para cores que não tem os cones verdes é designado deuteranópico; esse
indivíduo tem um espectro visual de amplitude perfeitamente normal porque os cones verdes ausentes
operam na porção medial do espectro, onde os cones vermelhos ou azuis também operam.
Deficiência para o azul: ocasionalmente, um indivíduo tem “deficiência para o azul”, que resulta
da diminuição ou ausência de receptores azuis. Se observarmos novamente a fig. 50.9, veremos que os
cones azuis são sensíveis a uma série espectral quase inteiramente distinta daquela dos cones vermelhos e
verdes. Portanto, se os receptores azuis estão completamente ausentes, o indivíduo tem maior
preponderância de verde, amarelo, laranja e vermelho no seu espectro visual do que de azul, o que produz
esse tipo raramente observado de deficiência ou cegueira para cor. Neste caso, o indivíduo é denominado
de tritanópico.
5.8. Tipos de Células Ganglionares
A maior parte das células ganglionares na retina apresenta campo receptivo organizado como
centro-periferia com seu centro ON ou OFF. Essas células podem ainda ser divididas em função de
seu aspecto, de suas conexões ou de suas propriedades eletrofisiológicas. Na retina do macaco, assim
como na retina humana, distinguem-se dois tipos principais de células ganglionares: células
ganglionares do tipo M, que são grandes, e células ganglionares do tipo P, menores (M significa
magno, grande; P significando parvo, pequeno).
Células M(magno): campos receptivos maiores; condução rápida do PA; mais sensíveis a
estímulos com baixo contraste. Importantes para detecção de movimento e profundidade (5% do
total das céls. ganglionares).
Células P(parvo): campos receptivos menores; condução mais lenta do PA; sensíveis à diferença
de luminosidade. Importantes para detecção da forma da imagem e a detalhes finos. Responsáveis
pela detecção da cor (90% do total das céls. ganglionares).
6. Vias Ópticas (fig. 10.3 e 10.5)
Após deixarem a retina, os impulsos são transmitidos posteriormente pelo nervo óptico, que é
formado pelos axônios das células ganglionares. Os nervos ópticos provenientes dos dois olhos formam o
quiasma óptico na base do cérebro. Neste local, há o cruzamento das fibras provenientes das duas metades
nasais da retina para o lado oposto, e as fibras das metades temporais permanecem do mesmo lado,
formando o feixe ou trato óptico. As fibras do trato óptico se dirigem ao tálamo (corpo geniculado
lateral). Do corpo geniculado lateral, partem fibras que se dirigem para o córtex visual primário, no lobo
occipital.
Para entender a importância da decussação parcial da projeção retinofugal no quiasma óptico,
vamos entender o conceito de campo visual. O campo visual completo é toda a região do espaço que pode
ser vista com ambos os olhos olhando diretamente para frente. Fixe seu olhar em um ponto bem a sua
frente. Agora imagine uma linha vertical passando através de ponto de fixação, dividindo o campo visual
15
em duas metades, direita e esquerda. Por definição, objetos que aparecem à esquerda dessa linha média
estão no hemicampo visual esquerdo, e objetos que aparecem à direita da linha média estão no
hemicampo visual direito (fig. 10.3).
Olhando diretamente à frente com ambos os olhos abertos e, então, fechando alternadamente um
olho e depois o outro, você verá que a porção medial de ambos hemicampos visuais é vista por ambas as
retinas. Essa região do espaço é, portanto, chamada de campo visual binocular. Observe que objetos na
região binocular do hemicampo visual esquerdo serão visualizados na retina nasal do olho esquerdo e na
retina temporal (lateral) do olho direito. Uma vez que as fibras da porção nasal da retina esquerda cruzam
para o lado direito no quiasma óptico, toda a informação acerca do hemicampo visual esquerdo é dirigida
para o lado direito do encéfalo. Lembre-se desta regra: fibras do nervo óptico cruzam no quiasma óptico
de forma que o hemicampo visual esquerdo é “visualizado” pelo hemisfério direito, e o hemicampo
visual direito é “visualizado” pelo hemisfério esquerdo.
6.1. Alvos do Trato Óptico (fig. 10.3)
Um pequeno número de axônios do tracto óptico desprende-se do conjunto para estabelecer
conexões sinápticas com células no hipotálamo, e cerca de 10% restante continua após atravessar o
tálamo, indo inervar o mesencéfalo. A maior parte deles, entretanto, inervará o núcleo (ou corpo)
geniculado lateral do tálamo dorsal (NGL). Os neurônios do NGL originam axônios que se projetam para
o córtex visual primário.
A partir do que sabemos acerca de como o mundo visual está representado na projeção retinofugal,
podemos predizer os tipos de déficits perceptuais que resultariam de sua destruição em diferentes níveis,
como poderia ser o caso de um traumatismo craniano, um tumor ou uma interrupção no fornecimento de
sangue. Como mostrado na figura 10.5, enquanto uma transecção do nervo óptico esquerdo tornaria uma
pessoa cega apenas do olho esquerdo, uma transecção do tracto óptico esquerdo causaria cegueira no
campo visual direito, visto por ambos os olhos. Uma transecção na linha média do quiasma óptico afetaria
apenas as fibras que cruzam a linha média. Uma vez que essas fibras originam-se nas porções nasais de
ambas as retinas, teríamos uma resultante cegueira apenas para as regiões do campo visual enxergadas
pelas retinas nasais, ou seja, a periferia do campo visual em ambos os lados. Tendo em vista que déficits
únicos são o resultado de lesões em diferentes locais, neurologistas e neuroftalmologistas podem localizar
os locais das lesões, verificando os déficits no campo visual.
16
Tálamo - núcleo
geniculado lateral
Córtex Visual
• Percepção
visual consciente
Hipotálamo – n.
supraquiasmático
• Ritmos biológicos
Mesencéfalo
(área pré-tectal):
• Diâmetro Pupilar
• Acomodação cristalino
• Controle dos músculos
extrínsecos dos olhos
6.2. Alvos não-talâmicos do tracto óptico (fig. 10.3)
Como dissemos, algumas células ganglionares da retina enviam axônios para inervar outras
estruturas que não o NGL. Projeções diretas a uma parte do hipotálamo têm um papel importante na
sincronia de uma variedade de ritmos biológicos - incluindo sono e vigília – com o ciclo diário de claroescuro. Projeções diretas para uma parte do mesencéfalo chamada de área (ou núcleo) pré-tectal,
controlam o tamanho da pupila e certos tipos de movimento oculares, além do controle do cristalino,
acomodando-o para uma visão próxima ou distante.
6.3. O Núcleo Geniculado Lateral (NGL) (fig. 10.9)
Os NGL direito e esquerdo, localizados no tálamo dorsal, são os principais alvos dos dois tractos
ópticos. Vistos em uma secção transversal, cada NGL parece estar arranjado em seis camadas distintas de
células. Por convenção, as camadas são numeradas de 1 a 6. A separação dos neurônios no NGL em
camadas sugere que vários tipos de informações oriundos da retina são mantidos separados nesse relé
sináptico, e, na verdade, é exatamente esse o caso: axônios provenientes de células ganglionares do tipo
M terminam nas camadas 1 e 2; e os das células ganglionares do tipo P terminam nas camadas 3 a 6.
Além dessa separação, os sinais oriundos dos dois olhos também são mantidos separados no NGL. No
NGL direito, os axônios do olho direito (ipsilateral) estabelecem sinapses nas camadas 2, 3 e 5, enquanto
que aqueles provenientes do olho esquerdo (contralateral) estabelecem sinapses nas camadas 1, 4 e 6.
17
7.Córtex Visual Primário
Localizado no córtex occipital, encontra-se principalmente ao longo da fissura calcarina (área 17). É
responsável pela detecção de formas, sombras, contrastes, bem como pela orientação espacial e de bordas.
A estimulação desta área faz com que o indivíduo tenha alucinações simples, tais como: lampejos
de luz, manchas, cores ou formas simples.
A destruição bilateral do lobo occipital leva à cegueira. A lesão unilateral faz com que o indivíduo
perca as metades contralaterais dos campos visuais. Por exemplo, a lesão do lobo occipital esquerdo faz
com que o indivíduo perca a visão das metades direitas dos campos visuais dos dois olhos (luz que incide
sobre a retina nasal direita e sobre a retina temporal esquerda).
8. Áreas Visuais Secundárias
Localizadas ao redor da área visual primária, estão as áreas de associação visual. A lesão destas
áreas faz com que o indivíduo deixe de reconhecer os objetos. Também ocorre a dislexia ou cegueira da
palavra, isto é, o indivíduo tem dificuldade para entender o significado das palavras que lê.
É nesta área que estão armazenados os dados de experiências prévias, ou seja, a memória visual.
9. Comentário Final
O que significa ver? Uma resposta é que a visão é o processo de descobrir, a partir de imagens, o
que está presente no mundo visual e onde está presente.
Nós sabemos hoje que o sistema visual do cérebro executa essa tarefa por meio de processamento
distribuído em muitas regiões corticais (aproximadamente 50% do cérebro estão de, alguma forma,
envolvidos com processamento visual). Estudos clínicos e experimentos com animais levaram à
conclusão de que cada área é responsável por um aspecto particular da visão, como profundidade, forma,
movimento e cor.
Apenas recentemente ficou claro que essas características são processadas mais em paralelo do que
em série. Duas vias (das células ganglionares P e das células ganglionares M) originam-se na retina e
continuam em duas vias de processamento cortical, que se dirigem aos córtices parietal posterior e
temporal inferior (fig 25.12). A via temporal inferior está mais envolvida na determinação do o que da
visão, enquanto a via parietal posterior mais com o onde da vida.
18
A descoberta dessas vias paralelas produziu um novo problema para o estudo da percepção visual.
A integração em uma via em série é realizada progressivamente, pela transformação da informação
conduzida de uma área à próxima. Em um sistema de vias paralelas, cada uma com sua própria função, a
integração pode apenas ser feita interativamente.
Como e onde essa interação ocorre no sistema visual? Nesse ponto, nos deparamos com um
importante fato anatômico: não há uma área cortical única com a qual todas as outras áreas corticais se
comunicam exclusivamente, nem no sistema visual e nem em qualquer outro sistema. Em resumo, o
córtex deve usar uma estratégia diferente para gerar a imagem visual integrada. A percepção visual é
provavelmente um processo de estágios múltiplos. Outros centros do cérebro que fazem conexões com o
sistema visual e que são conhecidos por influenciar a atenção visual, tais como o córtex pré-frontal, o
claustro ou o pulvinar podem permitir que os mecanismos de atenção se correlacionem com os fluxos de
informação visual em uma percepção coerente.
Bibliografia:
1- BEAR, MF et al. Neurociências, 3a edição, Ed. Artmed, 2008.
2- GUYTON, A e HALL, JE. Tratado de Fisiologia Médica, 11a ed., Ed. Elsevier, 2006.
3- KANDEL, ER et al. Princípios de Neurociência, 4a ed., Ed. Mc Graw-Hill, 2002.
4- LENT, R. Cem Bilhões de Neurônios, 2a ed., Livraria Atheneu, 2010.
5- SILVERTHORN, DU. Fisiologia Humana, 2a ed., Ed. Manole, 2003.