SISTEMA IMUNE
DE INVERTEBRADOS
IMUNOLOGIA
Introdução
•
Sistemas imunes são geralmente caracterizados por sua habilidade
em distinguir entre células, tecidos ou moléculas próprias e nãopróprias, e eliminar as não-próprias (Janeway, 2001).
•
Os metazoários possuem mecanismos de reconhecimento entre
próprio e não próprio e são capazes de se proteger da invasão de
microorganismos, vírus, parasitas e células exógenas, porém, os
mecanismos utilizados não são necessariamente homólogos dentre
os filos (Du Pasquier, 2001).
Sistema Imune
O sistema imune pode ser dividido em 03 categorias (Cooper):
Barreiras Externas (Químicas e Físicas)
Resposta Celular
Imunidade Humoral
Barreiras Externas
Importantes para barrar a entrada de patógenos e impedir danos aos tecidos:
Testa de equinodermos
Concha de moluscos
Muco em celenterados, anelídeos, moluscos
- efetivos na primeira linha de defesa
- forma uma camada que pode impedir a entrada de microorganismos
(inclusive pelo batimento dos cílios)
- moléculas anti-microorganismos estão presentes no muco
Resposta Celular
Células do sangue (hemolinfa)
Reparo e coagulação
Fagocitose
Formação de nódulos
Encapsulação
Citotoxicidade
Rejeição de transplante
Resposta Celular
Fagocitose: ocorre em todos os filos, fatores quimiotáticos e de
reconhecimento envolvido
Células homeostáticas: coagulações de lesões físicas
Células de pigmento: sipuncula, echiura, celenterados,
lophophorados e equinodermos (podem ter propriedades
antibacterianas e antialgas)
Fagocitose
Primeira resposta celular contra vários patógenos e parasitas
Quimiotaxia em Lubricus, tecidos impróprios liberam
quimioatratores que atraem celomócitos; em Ostra, proteínas da
parede celular e envelope de bactérias são quimioatratores de
hemócitos
Receptores de C3 em celomócitos de anelídeos e equinodermos
Opsonização: substâncias sorologicamente relacionada à
fibronectina humana (opsonina) encontrada em tecido de
invertebrados (incluindo em hemócitos de caracóis), provavelmente
função similar às opsoninas
Killing factors nas células do sangue:
pH baixo em moluscos
Enzimas lisossômicas (peroxidades) em todos invertebrados
Lizosima em anelídeos, moluscos e equinodermos
Espécies reativas de oxigênio em moluscos
Formação de Nódulos
Alguns invertebrados são incapazes de realizar a fagocitose de
pequenas párticulas
Nódulos sequestram essas párticulas formando agregados
Microorganismos dentro do nódulo podem morrer por stress
fisiológico ou pelo efeito tóxico de quinonas e melaninas geradas
pela atividade da fenoloxidase
Ocorre em anelídeos, moluscos, crustáceos, insetos e
equinodermos
Encapsulação
Organismos externos muito grandes para serem fagocitados são
confinados em cápsulas multicelulares
Defesa adequada contra nematodes,fungos e bactérias
O processo de encapsulação é provavelmente iniciado pelo contato
randômico de hematócitos com o corpo exterior,fatores
quimiotáticos e componentes do sistema PpO ativado
Os parasitas encapsulados podem ser destruídos por moléculas
tóxicas como quinonas e também por peroxidases e formas reativas
do oxigênio presente em moluscos
Reações Citotóxicas
Além dos nódulos e da encapsulação as células sanguíneas de
invertebrados realizam reações citotóxicas visando invasores
Similar ao NK dos mamíferos
O contato entre a célula efetora e a célula alvo é necessário para
iniciar a reação citotóxica
A citotoxidade das células sanguíneas de anelídeos,moluscos e
equinodermos pode ser inibida por uma variedade de carboidratos
Imunidade Humoral
A resposta humoral dos invertebrados inclui lisinas, aglutininas e
fatores antibióticos.
Fatores parecidos com anticorpos e sistema complemento tem sido
descritos e tem importância para a filogenia.
Lisinas
Detectadas pela sua habilidade em lisar eritrócitos de vertebrados
Enzimas proteolíticas.
Imunidade Humoral
Aglutininas
Encontrados em bactérias fungos, invertebrados e mamíferos
Ficam frequentemente no fluído do celoma e na hemolinfa dos
invertebrados.
Muitos tipos de aglutininas para o mesmo organismo
Estimulam a produção de novas proteínas com função
imunológica
Função de reconhecimento vem sendo corroborada
Imunidade Humoral
Fatores antibióticos
Ambiente cheio de microorganismos perigosos
Proteção por meio desses fatores
Embora ocorra naturalmente e de forma induzida, pouco se sabe sobre a
natureza dos fatores
Considerações
Novos estudos indicam que invertebrados são capazes de uma resposta
antigênica análoga à dos vertebrados, isto é, resposta específica,
transferível.
Estes dados são importantes para estudos filogenéticos e evolutivos.
Imunidade Humoral
Propriedade de opsonização mediada por lectinas.
Lectinas também ativam o sistema complemento PpO.
Ausência de resposta antecipada
A maioria apresenta células análogas a leucócitos.
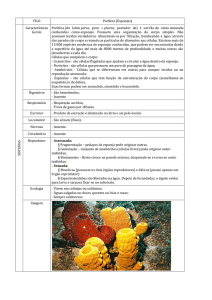
Porifera
Sistema imune em nível celular – modelo muito utilizado para o
sistema imune humano
Células das esponjas quando separadas em solução de água
salgada, elas brevemente se juntam, formando novas esponjas, a
não ser que sejam de tipos diferente.
Metchnikoff (1892) descobriu a atividade fagocitária exercida pelas
células das esponjas (arqueócitos), como um mecanismo para
eliminar as partículas não-próprias e, de um modo mais avançado
encapsular o material estranho dentro de agregado de células de
esponjas para uma posterior eliminação. Contexto de inflamação.
Experimentos de transplante para estudar sistema imune em
esponjas.
Porifera
Formação de barreiras para separar
os tecidos não-próprios
Axinella verrucosa
esponja marinha Axinella
verrucosa (Buscema and van de
Vyver, 1983)
esponja de água doce Ephydatia
muelleri (Mukai, 1992)
Reação por fatores citotóxicos que
destróem o transplante
Ephydatia muelleri
as esponjas marinhas
Callyspongia diffusa (Hildemann
et al., 1979) e Geodia cydonium
(Pfeifer et al., 1992)
Geodia cydonium
Porifera
Defesa contra microorganismos/parasitas
Esponjas marinhas contém bactérias (simbióticas, comensais e
parasitas)
Mecanismo molecular discriminam entre bactérias simbióticas ou
comensais das parasitas
Forte mecanismo de eliminar microorganismos – digestão
intracelular - arqueócitos (macrófagos das esponjas)
Domínio SRCR contém moléculas de membrana que estão
envolvidas no reconhecimento de bactérias. As bactérias parasitas
ingeridas são mortas por um mecanismo oxidativo ou não-oxidativo
(enzimático)
Porifera
Respostas de Histo(in)compatibilidade em Esponjas
Sistema “primmorph”, sistema de agregados tridimensional que
oferece a proliferação e diferenciação de células.
Reações de células de esponjas misturadas (MSCR)
Células do mesmo indivíduo (autogenético MSCR)
Células de indivíduos diferentes (alogenético MSCR)
Agregados de diferentes organismos formam, depois de um
pequeno período de contato, diferentes indivíduos de agregados
específicos (Fig. 3D)
FK506 causa immunosupressão (como em humanos) (Fig. 3C)
Usando modelos de transplantes em G. cydonium e S. domuncula
foi estabelecido que citocinas derivadas de macrófago são ativadas
durante a rejeição “allograft” (enxerto externo).
Porifera
Função de citocina AIF-1 e trancrição do fator
TCF durante a rejeição “allograft” (nível tecidual)
e reação alogenética das células de esponjas
(MSCR, nível celular)
O reconhecimento de tecido/célula alogenético
faz com que ocorra um aumento nas expressões
de gene AIF-1 e TCF. Este efeito é inibido pela
incubação com o imunosupressor FK506. Desde
que o polipetídeo recombinante AIF-1 causa in
vitro expressão do gene de TCF; assim ocorre um
efeito seqüencial de AIF-1 em TCF. Esse aumento
das expressões de AIF-1 e TCF resultam em uam
rejeição por histoincompatibilidade, enquanto a
mediada por FK506, geram uma baixa regulação
desses dois fatores e uma fusãodos
tecidos/células alogenéticos. Em humanos a
expressão do gene de AIF-1 induz uma resposta
inflamatória e uma rejeição do transplante
Porifera
Porifera
Similaridades entre G. cydonium e humanos:
MAP quinases são componentes essencias para o via mediada por LPS.
Estudos recentes revelaram que a quinase KRS que é envolvida na
fosforilação de MAP quinases, também é presente em Demonspongiae.
Via conservada entre esponjas e humanos.
Seqüências isoladas na esponja contém domínios de imunoglobulinas,
receptores quinas (RTK) e moléculas de adesão de esponjas (SAM).
Os domínios de imunoglobulinas e de RTK, mostram uam grande
similaridade com o domínio V dos domínios de imunoglobulinas de
mamíferos.
Ao nível de aminoácido o domínio de Ig, de RTK e de SAM, são 94%
parecidos com o dos mamíferos.
São mais parecidos com os humanos (tanto em relação aos domínios,
quanto às moléculas envolvidas no sistema imune) do que com outros
invertebrados (como a D. melanogaster e C. elegans).
Cnidaria
Anos 70: imunologista Burnet traz à luz a
evolução do sistema imune.
Colônias sésseis capazes de distingüir tecidos
próprios de não próprios.
O hidróide Hydractinia passa a ser muito
usada em estudos de imunologia de cnidarios.
Tecidos isogenéticos e alogenéticos
histocompatíveis são capazes de se fundir em
Hydractinia.
Tecidos incompatíveis não se fundem e geram
uma rejeição peculiar.
Reação efetuada por um tipo especial de
nematocisto: mastigóforo microbásico.
Estas células ocorrem exclusivamente no
compartimento das ligações vasculares de
uma colônia.
Cnidaria
Os mastigóforos microbásicos migram para a região do contato
entre os tecidos e se acumulam.
Os nematocistos reunidos ajustam seu cnidocílios em direção ao
tecido exógeno.
Todos nematocistos atacam o tecido exógenos quase
sincronizadamente, descorregando suas toxinas.
Os nematocistos que chegam posteriormente, descarregam
imediatamente sobre o tecido exógeno.
Apenas o contato entre tecidos incompatíveis causa a descarga dos
nematocistos.
Cnidaria
Esta reação ocorre até que um dos dois indivíduos morra.
Após a interrupção do contato, o número de mastigóforos
microbásicos é reduzido ao nível normal por fagocitose.
A fusão dos tecidos compatíveis ocorre de 2 a 4 horas após o
contato, criando grandes ligações vasculares entre as colônias.
Há também a fusão transitória
Os tecidos se fundem nas primeiras 12 ou 24 horas.
Após esse período ocorre necrose local do tecido que se espalha.
De 1 a 3 dias depois do aparecimento da necrose, rejeição passiva
e separação das colônias.
Molusco
A maioria dos moluscos apresentam
fagócitos, células quimiotáticas e citotóxicas
Fatores humorais: lectinas, hormônios
(ACTH, POMC), etc.
Discriminação entre próprio e não-próprio
Resposta inflamatória inicial seguida de
encapsulamento do tecido estranho
Apresentam resposta por stress
Resposta por stress não requer intervenção
de inúmeras células ou orgãos
Célula imuno-neuroendócrina realiza funções
imunológicas fundamentais
ACTH e POMC são os principais hormônios
mediadores de diversas funções no
organismo
Vermes
Estudos recentes demonstraram uma nova
visão na pesquisa do sistema imune inato,
pois, normalmente, são utilizados modelos
de insetos para o estudo.
Foi a partir da observação de que o
nematóide “Caernorhabditis elegans”
poderia ser morto pelo patógeno
“pseudomonas aeruginosa”, que iniciou-se
uma verificação de quando a resposta
imune inata de humanos e vermes
poderiam ser similares.
Dois genes foram identificados – sek1, que
codifica a MAP quinase quinase, e nsy1,
que codifica a MAP quinase quinase que
está associada com a susceptibilidade de
vermes a patógenos. Estes genes foram
identificados como pertencentes ao mesmo
padrão de sinalização envolvido na
resposta imune inata humana.
Referências Bibliográficas
Werner E. G. Müller And Isabel M. Müller. 2003. Origin of the Metazoan
Immune System: Identification of the Molecules and Their Functions in
Sponges. INTEGR. COMP. BIOL., 43:281–292
Louis Du Pasquier. 2001. The immune system of invertebrates and
vertebrates. Comparative Biochemistry and Physiology Part B., 129:1-15
Luis F. Cadavid. 2004. Self-discrimination in colonial invertebrates: genetic
control of allorecognition in the hydroid Hydractinia. Development and
Comparative Immunology., 28: 871-879.
http://www.unicamp.br/cipoi/lic/overview.htm F. Aspectos evolutivos do
sistema imune.
R. J. Turner. 1994. Imunology: a comparative approach. John Wiley & Sons.
222 p.
E. Ottaviani. 2004. The mollusc as a suitable model for mammalian
immune-neuroendochrine investigations. ISJ, 1: 2-4.
Componentes do grupo
César Rafael Ferreira
Daniel Justi
Diego Morais
Fábio Góis
Gedsney Müller
Guilherme Galera
Heirie Murakami
Fernanda Saturni
Natália Ravanelli
Nathalia Machado