TRABALHO FINAL
Fundamentos de Ecologia e de Modelagem Ambiental aplicados à conservação da Biodiversidade
CST-304
EFEITOS DO AUMENTO DA CONCENTRAÇÃO DE CO2 NA ATMOSFERA
SOBRE O CRESCIMENTO E O DESENVOLVIMENTO DAS PLANTAS
Professores: Dalton M. Valeriano, Silvana Amaral, Eduardo Arraut
Aluna: Juliana Paiva Nunes Kury ([email protected])
JUNHO, 2011
0
Efeitos do Aumento da Concentração de CO2 na Atmosfera sobre o Crescimento e o
Desenvolvimento das Plantas
Juliana Paiva Nunes Kury
1. INTRODUÇÃO
Conforme previsões do relatório do IPCC (Intergovernmental Panel of Climatic Change) em
2007, a concentração atmosférica global de dióxido de carbono (CO 2) aumentou de um valor préindustrial de aproximadamente 280 ppm para 379 ppm em 2005, e até meados deste século a
concentração pode chegar a 720 ppm. Além disso, o IPCC projeta um aumento da temperatura da
superfície terrestre da ordem de 1º a 3,5ºC até o ano de 2100.
Ainda que mudanças climáticas não seja uma influência nova na biosfera, a velocidade das
alterações provocadas pela intensificação do efeito estufa é maior que qualquer outra mudança que
tenha ocorrido nos últimos 10.000 anos. Essa alteração “brusca”, somada ao estresse provocado nos
últimos dois séculos pelas atividades antrópicas, pode ultrapassar a capacidade de regeneração dos
ecossistemas levando à perda significativa de espécies (Costa, 2004).
2. FOTOSSÍNTESE
Por meio da fotossíntese as plantas, algas e alguns tipos de bactérias convertem e energia física
da luz solar em energia química, onde as plantas fixam o CO 2 e o incorporam à sua biomassa
(Souza et al., 20008).
Conforme Martinez (2000), o processo fotossintético das plantas ocorre nos cloroplastos e
resulta na liberação de oxigênio molecular (O2) e na captura de CO2 da atmosfera, utilizado para
sintetizar carbono. Em geral, o processo é analisado em duas etapas: 1) fotoquímica, que resulta: a)
na produção do agente redutor (NADPH); b) liberação do O 2 como subproduto da dissociação da
molécula da água; e c) formação de ATP, por meio do complexo ATP-sintase, e 2) química na qual
o CO2 é fixado e reduzido até carboidratos, utilizando o NADPH e o ATP, produzidos na etapa
fotoquímica.
As plantas respiram, e por meio desse processo que ocorre no citoplasma e nas mitocôndrias, os
carboidratos e outros constituintes celulares são convertidos em CO 2 e água com a liberação de
energia. Assim, a fotossíntese e a respiração são processos importantes na regulação dos teores de
O2 e CO2 da atmosfera terrestre.
2.1. Sistemas Fotossintéticos
Segundo Martinez (2000), o ciclo de carbono descoberto por Melvin Calvin é denominado Ciclo
Fotossintético Redutivo de Carbono de Plantas C3, porque o primeiro composto estável formado é
um composto de 3 carbono (ácido fosfoglicérico). Após os trabalhos de Calvin, outros
pesquisadores (Hatch, Slack, Kortaschak) determinaram que algumas espécies de gramíneas (ex.
cana de açúcar e milho) são capazes de fixar CO2 em compostos de 4 carbono, além de executarem
o ciclo C3. Posteriormente, foi descoberto que em algumas espécies de plantas em regiões áridas
1
(ex. cactáceas) abrem seus estômatos somente à noite, quando fixam CO 2 pelo mecanismo C4, e
fecham durante o dia para evitar a perda excessiva de água, mas apresentam o ciclo C3. Estas
últimas são denominadas plantas CAM (metabolismo ácido crassuláceo).
Nas plantas C3, a fixação de carbono em nível de açucares ou outros compostos pode ser
considerada como ocorrendo nas fases: 1) carboxilação, catalisada pela enzima Rubisco; 2) redução,
onde se utiliza o NADPH e ATP; 3) regeneração do aspertor de CO 2; e 4) síntese de produtos.
Na carboxilação o CO2 é adicionado a um açúcar de 5 carbonos, a ribulose ,5-bifosfato (RuBP)
para formar duas moléculas de ácido fosfoglicérico (PGA) de três carbono. Essa reação é catalisada
pela Rubisco, proteína que atua como uma carboxilase e também apresenta uma atividade
oxigenase1. Em seguida, o PGA é convertido (reduzido) a um açúcar de 3 carbono (Triose-P), o
aspertor inicial de CO2 (RuBP) é regenerado para posteriores reações de fixação. Os produtos finais
da fotossíntese são considerados primariamente como açucares e outros carboidratos, mas gorduras,
ácidos graxos, aminoácidos e ácidos orgânicos tem sido também admitidos como sintetizados na
fixação fotossintética do carbono (Martinez, 2000).
As espécies de plantas C4 apresentam uma estrutura nas folhas, que se caracteriza por um feixe
vascular bastante desenvolvido, rodeado por células denominados “células da bainha do feixe
vascular”. A fixação inicial de CO2 ocorre nas células mesofílicas. No citossol dessas células, o CO 2
reage com o fosfoenolpiruvato, via enzima PEPcarboxilase para formar o oxalacelato (ácido com 4
átomos de carbono), que pode ser reduzido a malato com a utilização do NADPH ou pode ser
animado a aspartato. Posteriormente, os ácidos de 4 carbonos são transportados até as células da
bainha, onde são carboxilados, liberando CO2 e produzindo piruvato. A seguir, o CO2 é liberado e
refixado via C3. O Piruvato resultante de descarboxilação retorna as células mesofílicas, onde é
convertido em fosfoenolpiruvato, regenerando o aceptor inicial do CO 2. No mecanismo de fixação
das plantas C4, a alta atividade carboxilativa da PEPcarboxilase assegura uma alta concentração de
CO2 nas células da bainha, onde ocorre a refixação do CO2 via ciclo C3. Dessa forma, predomina
nas células da bainha, a atividade carboxilase da rubisco e uma menor taxa de fotorrespiração,
porque a alta concentração de CO2 compete melhor, como O2, pela enzima e pelo substrato (RuBP).
Por outro lado ao ocorre a fotorrespiração, o CO2 produzido não consegue sair das folhas, pois é
rapidamente refixado pela PEPcarboxilase nas células mesofílicas (Martinez, 2000).
Há várias consequências importantes da presença da bainha vascular. Uma delas é que as taxas
de absorção de CO2 são muito mais altas, pois o sistema pode armazenar mais carbono de forma
intermediária (no ácido C4) tornando a planta relativamente menos dependente de controlar a
abertura e fechamento de estômatos para prevenir a perda de água. O resultado é que quando se
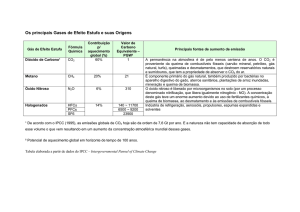
compara o rendimento quântico 2 de plantas C3 e C4 em diferentes temperaturas, nota-se que
enquanto as plantas C4 têm desempenho constante em temperaturas que variam entre 10 e 40ºC, as
C3 apresentam uma queda linear em desempenho quando se aumenta a temperatura. É possível
observar que plantas C3 levam vantagem até cerca de 25ºC, mas em função das C4 não
apresentarem queda de rendimento em temperaturas mais altas passam a ter maior eficiência
relativa acima de 25ºC (Figura 01) (Buckeridge et al., 2007).
1
Oxigenase: etapa inicial da fotorrespiração (processo que reduz a fixação de CO2 e o crescimento das plantas)
2
Rendimento quântico: equivale ao nº de moles de CO2 absorvido pelo nº de quanto é captado
2
Figura 01. Rendimento comparado de plantas C3 e C4
O fato de as plantas C4 lidarem melhor com temperaturas mais altas também permitem que seus
sistemas de captação de luz possam suportar intensidades luminosas muito maiores. Com isto,
enquanto as plantas C3 funcionam bem entre 400 a 500 μmoles , as plantas C4 podem funcionar em
intensidades maiores do que 2000 μmoles de fótons m-2.s-1 (Tabela 1).
Além disso, as plantas C4 são bem mais eficientes em lidar com a água, isto se deve à maior
eficiência em captar e armazenar o carbono oriundo do CO 2, isto é, a bomba de armazenamento do
ácido de 4 carbonos, o que permite às plantas C4 um gerenciamento melhor da abertura estomática,
que é um processo fundamental no controle da transpiração foliar (Buckeridge et al., 2007).
Tabela 1. Comparação geral entre os desempenhos dos sistemas fotossintéticos dos tipos C3 e C4
Temperatura ótima
Saturação de luz
Taxa transpiração
Fotorrespiração
Ponto Compensação CO2
Condutância estomática
Câmara Substomática (ppm)
C3
20 - 25
400 - 500
500 - 1000
SIM
20 - 100
0,5
210-280
C4
30 -45
> 2000
200 – 350
NÃO
0–5
0,2
120-150
Plantas C4 são mais eficientes para os níveis atuais de CO2, mas por outro lado são as plantas C3
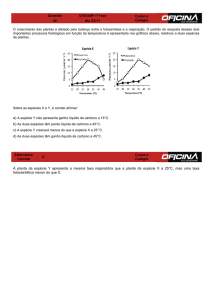
que respondem mais positivamente a um aumento de CO 2. A resposta da taxa fotossintética de
milho (C4) a concentrações crescentes de CO2 é de saturação a uma concentração de cerca de 450
ppm, enquanto a taxa fotossintética de trigo (C3) satura a uma concentração de 850 ppm (Figura 2).
Figura 2. Curvas de respostas da fotossíntese a incrementos na concentração de CO2 em função
do tipo de metabolismo C3 (trigo) ou C4 (milho)
3
Plantas com metabolismo fotossintético do tipo C3 aumentam sua biomassa em média 47%,
enquanto que em plantas C4 o aumento é de 11% quando submetidas ao dobro da concentração
atual de CO2 (Buckeridge et al., 2007).
3. RESPOSTAS DAS PLANTAS A ATMOSFERA ENRIQUECIDA DE CO2
Em geral, os efeitos provocados pelo aumento da concentração atmosférica de CO 2 sobre as
plantas podem ser classificados em: 1) diretos que referem-se aos incrementos na disponibilidade de
substrato para a fotossíntese e na produtividade primária das plantas pela ação do CO 2 como
“fertilizante atmosférico”, e 2) indiretos relacionados às alterações climáticas provocadas pelo
efeito estufa, principalmente no incremento da temperatura, que pode afetar a fisiologia e a
produtividade das plantas (Buckeridge et al., 2007).
Os efeitos sobre a fisiologia e crescimento das plantas são incluídos na: 1) fotossíntese líquida e
respiração; 2) condutância estomática e a eficiência do uso da água; 3) alocação de carbono e
crescimento; 4) estrutura da planta e fenologia; e 5) concentração de nutrientes na planta.
Sobre a fotossíntese os efeitos são: 1) aumento da atividade RUBISCO via estímulo da
carboxilação e inibição da oxigenação da RuBP; 2) redução da abertura estomática; 3) alteração da
respiração. Como consequências, ocorrem alterações no balanço de água e carbono nas plantas, com
efeitos secundários sobre o crescimento, partição de recursos e sínteses de composto de defesa
(Buckeridge et al., 2008).
3.1. Respostas das Plantas Brasileiras a uma Elevação na Concentração Atmosférica de CO 2
No Brasil, estudos pioneiros em linhas de pesquisa experimental consolidadas com respostas de
plantas às mudanças climáticas foram iniciados pelos grupos de pesquisa dos professores Marcos
Buckeridge (Instituto de Botânica de São Paulo) e Carlos Martinez (Universidade Federal de
Viçosa). Resultados iniciais são encontrados em Aidar et al. (2002) e Costa (2004) examinando
inicialmente as respostas da espécie Hymenaea courbaril e posteriormente em Buckeridge et al.
(2007); Godoy (2007) com Sesbania virgata, Schizolobium parahyba, Piptadenia gonoacantha,
Dalbergia nigra e Hymenaea courbaril,; e Marabesi (2007) com Senna alata.
Inicialmente o estudo de Aidar et al. (2002) sobre o comportamento de plântulas de Hymenaea
courbaril, uma espécie secundária tardia, mostrou que a fotossíntese foi significativamente
aumentada em atmosfera de CO2 elevado. Nos experimentos, sementes recém geminadas foram
cultivadas em câmaras de topo aberto com concentrações alteradas de CO 2. A comparação entre
plântulas crescendo em atmosfera de CO2 a 360 e 720 ppm, mostraram aumento significativo em
área foliar apenas nos metáfilos de plântulas crescendo nas concentrações mais altas. Contudo, de
acordo com resultados, parece que as plantas crescidas sob concentrações enriquecidas de CO 2 não
aclimataram3 e, portanto, sob as condições climáticas previstas com base nos níveis atmosféricos
atuais, plântulas de H. courbaril deverão estabelecer mais rapidamente em seu ambiente natural e
podem servir como um mecanismo eficiente de sequestro de carbono pela floresta.
Em experimentos similares foi observado que além de aumentar a fotossíntese, há também um
grande aumento nos teores de sacarose, amido e celulose nas folhas e caules de H. courbaril. Esse
3
caracterizado por redução da capacidade fotossintética da planta para responder ao incremento do CO2 atmosférico
4
aumento no teor de amido das folhas dos jatobás confirma a hipótese de que o fluxo de carbono é
maior quando a planta cresce em CO2 elevado Além disso, foi observado em plantas que cresceram
em atmosfera de CO2 a 720 ppm, apresentaram uma diminuição significativa no índice estomático.
Com base nesses resultados, folhas de espécies de herbário cuja coleta havia sido feita no início do
século XX, foram analisadas e os resultados indicaram que folhas de 1919 e 1959 apresentaram o
dobro do índice estomático em comparação com as folhas de plantas atuais e incubadas em alto CO 2
(Costa, 2004). Estes resultados mostraram um sinal de aclimatação da planta, que vem diminuindo
sua capacidade total de entrada de CO2 nas folhas ao longo de um período em que a concentração
atmosférica de CO2 aumentou de 280 para 380 ppm. Como a diminuição do número de estômatos
ocorreu simultaneamente com um aumento no fluxo de carbono, é provável que os dois fenômenos
estejam relacionados (Buckeridge et al., 2008).
Posteriormente, Marabesi (2007) visando estudar o efeito do CO2 sobre o crescimento e
desenvolvimento de plantas da espécie Senna alata realizou um experimento, onde metade das
plantas foi submetida à concentração atmosférica atual de CO 2 (em torno de 380 ppm) e a outra
metade a uma atmosfera com 720 ppm. No final do experimento concluiu-se que as plantas
crescidas em alta concentração de CO2 possuíram menor altura (o que coincidiu com o menor
investimento no caule), maior número de folhas (relacionado à maior retenção das folhas antigas menor taxa de queda de folhas - e não devido a uma maior produção de folhas novas) e maior área
foliar total. Além disso, as plantas apresentaram maior taxa de assimilação de CO2 saturada de luz,
sem modificações na atividade da rubisco, o que sugere inibição da fotorrespiração. Contudo, ao
final do experimento as plantas tiveram um aumento de biomassa na ordem de 60% o que torna esta
espécie indicada para programas de sequestro de carbono em curto prazo, uma vez que estas plantas
possuem um ciclo de vida curto.
No estudo de Godoy (2007) foi avaliado, o estabelecimento de cinco espécies de Leguminosae
arbóreas da Mata Atlântica, pertencentes a diferentes grupos funcionais. As espécies estudadas
foram: S. virgata (feijão-do-mato) - pioneira; S. parahyba (guapuruvú), P. gonoacantha (paujacaré), e D. nigra (jacarandá-da-bahia) - secundárias iniciais; e H. courbaril (jatobá) - secundária
tardia. Após a germinação, as plântulas foram cultivadas por cerca de dois meses em câmaras de
topo aberto sob dois tratamentos: atmosfera ambiente (370 ppm) e enriquecida com CO 2 industrial
(720 ppm). Sob CO2 elevado as respostas dependeram da espécie, do parâmetro e da fase do
experimento analisada. No geral, as espécies mostraram maiores valores para altura, área foliar,
taxa de crescimento relativo inicial, assimilação líquida de CO 2, eficiência de uso da água e
capacidade fotossintética. Porém, uma análise mais detalhada revelou que as espécies mais iniciais
na sucessão (S. virgata, S. parahyba) apresentaram maiores valores para as taxas iniciais de
crescimento relativo, para o número de folhas em torno dos 40 dias após o plantio, para a
assimilação líquida de CO2, para o incremento percentual na capacidade fotossintética e no acúmulo
de biomassa quando submetidas a uma atmosfera enriquecida com CO 2. Essas espécies mostraram
também os menores valores para os incrementos percentuais na assimilação líquida de CO 2, na
eficiência intrínseca de uso da água e na redução da taxa de carboxilação máxima da Rubisco. Esses
resultados evidenciaram que os ciclos de vida mais curtos, as maiores densidades populacionais, as
madeiras de menor densidade e o quadro oposto apresentado pelas espécies secundárias tardias que
os diferentes grupos funcionais podem, no longo prazo, garantir maior estabilidade ao sequestro de
carbono em relação às espécies isoladamente, uma vez que sequestro de carbono é maior e mais
consistente quando se executa o processo de sucessão em contraposição ao plantio de uma única
5
espécie. Segundo Buckeridge (2007) isto porque com o passar do tempo o carbono deixa de ser
assimilado e transformado em biomassa pelas plantas pioneiras com maior rapidez e intensidade e
passa a ser captado de forma mais lenta, porém mais duradoura e com maior potencial de resposta
às mudanças climáticas que estão por vir, pelas espécies secundárias tardias.
4. CONCLUSÃO
De maneira geral, pode-se concluir que as espécies pioneiras S. alata e S. virgata, que iniciam a
formação da floresta no processo de sucessão ecológica, crescem rápido e sequestram grandes
quantidades de carbono rapidamente, enquanto as espécies de crescimento mais lento sequestram
menos carbono no mesmo período, porém vivem mais. Como as espécies que iniciam o processo (S.
alata e S. virgata) vivem entre 5 e 10 anos, as intermediárias (S. parahyba e P. gonoacantha) entre
25 a 30 anos, e as finais (D. nigra e H. courbaril) mais de 100 anos, uma resposta das pesquisas é
que uma floresta tropical em regeneração sequestra carbono continuamente por um longo período.
Além disso, os dados obtidos permitiu calcular o desempenho fisiológico em alto CO 2 de forma
comparada, e os dados sugerem que as espécies intermediárias, principalmente a P. gonoacantha,
têm um desempenho fisiológico melhor em relação às demais espécies. Isto significa que essas
espécies poderão ter maior probabilidade de se adaptarem as condições de alto CO 2 e pouca água
que se espera no futuro (Buckeridge et al., 2008).
No entanto, é importante ressaltar que esses resultados ainda possuem incertezas em relação aos
efeitos futuros sobre o crescimento e desenvolvimento das plantas, uma vez que o único efeito
avaliado nos estudos foi o da concentração atmosférica de CO2. Seria necessário saber também,
quais os efeitos em relação ao aumento da temperatura e as variações na disponibilidade hídrica.
Contudo, a compreensão do processo de sucessão ecológica é importante para a conservação,
manejo, monitoramento de áreas naturais e modelagem de biomas com elevada biodiversidade
como a floresta tropical, uma vez que este ecossistema pode desempenhar um papel fundamental na
mitigação dos impactos causados pelas crescentes concentrações de CO 2 na atmosfera, relacionadas
às alterações do clima global, captando carbono através da fotossíntese e transformando-o em
biomassa.
6
REFERÊNCIAS
Aidar, M.P.M., Martinez, C.A., Costa, A.C., Costa, P.M.F., Dietrich, S.M.C. & Buckeridge, M.S.
(2002). Effect of atmospheric CO2 enrichment on the establishment of seedlings of jatobá
(Hymenaea courbaril L. Leguminosae-Caesalpinioideae). Biota Neotropica v2 (n1).
Buckeridge, M. S., Mortari, L. C. & Machado, M. R. (2007). Respostas fisiológicas de plantas às
mudanças climáticas: alterações no balanço de carbono nas plantas podem afetar o ecossistema? In:
Rego, G. M., Negrelle. R. R. B & Morellato, L. P. C. Fenologia - Ferramenta para conservação e
manejo de recursos vegetais arbóreos. - Colombo, PR: Embrapa Florestas. Cap. 12: 213-230
Buckeridge M. S., Aidar M. P. M., Martinez C. A., Silva E. A. (2008). Respostas de Plantas às
Mudanças Climáticas Globais. In: Buckeridge, M.S. (Org). Biologia & Mudanças Climáticas no
Brasil. São Carlos: Rima Editora. 316p. Cap 04.
Costa, P.M.F.(2004). Efeitos da alta concentração de CO2 sobre o metabolismo de reservas e
estabelecimento de plântulas de jatobá de mata - Hymenaea courbaril L. var. stilbocarpa (Heyne)
Lee & Langenheim - Caesalpinioideae. Dissertação de mestrado, Instituto de Biologia, UNICAMP,
Campinas.
IPCC (2007). Climate Change 2007: Synthesis Report. Summary for Policymakers. An
Assessment of the Intergovernmental Panel on Climate Change. Acesso em:
<http://www.ipcc.ch/pdf/assessment-report/ar4/syr/ar4_syr_spm.pdf>. Acesso em: 30 mai. 2011.
Godoy, J.R.L. (2007). Ecofisiologia do estabelecimento de leguminosas arbóreas da Mata Atlântica,
pertencentes a diferentes grupos funcionais, sob atmosfera enriquecida com CO2: uma abordagem
sucessional. Tese de doutorado, Instituto de Botânica, Secretaria de Estado do Meio Ambiente, São
Paulo. 113 p.
Marabesi, M.A. (2007). Efeito do alto CO2 no crescimento inicial e na fisiologia da fotossíntese em
plântulas de Senna alata L. (Roxb.). Dissertação de mestrado apresentada ao Curso de Pósgraduação em Biodiversidade e Meio Ambiente do Instituto de Botânica de São Paulo. 70 p.
Martinez, C. A. (2000). Fotossíntese. Departamento de Biologia Vegetal. Apostila de graduação do
curso de Engenharia Florestal da Universidade Federal de Viçosa. Brasil. 63 p.
Souza A. P., Gaspar M., Tiné M. A., Buckeridge M. S. (2008). Ajustando os Botões: Como as
Plantas Lidam com o Aumento do CO2 Atmosférico?. In: Buckeridge, M.S. (Org). Biologia &
Mudanças Climáticas no Brasil. São Carlos: Rima Editora. 316p. Cap 06.
7