
147
1.5 EVOLUÇÃO
VOLUÇÃO DOS SISTEMAS REPRODUTIVOS DOS
METAZOA
Paulo Henrique
enrique Franco Lucinda
Doutor em Zoologia. Professor Adjunto IV
I do Departamento de Ciências Biológicas
da Universidade Federal do Tocantins - UFT, Porto Nacional, TO.
Introdução e Objetivos
Os organismos vivos mudam de forma no espaço e no tempo. Esta continuidade
temporal, ou seja, a manutenção da vida ao longo do tempo, somente é possível em
virtude do
o fenômeno reprodutivo. Neste capítulo fazemos a distinção entre
reprodução sexuada (sexual) e assexuada (assexual) e exploramos as razões pelas
quais, ao menos para animais, a reprodução sexuada parece oferecer vantagens
importantes sobre a assexuada. A seguir, discutimos a origem e a maturação
maturaçã das
células germinativas; o desenho dos sistemas reprodutivos; os padrões reprodutivos
dos animais; analisaremos a diversidade e o significado adaptativo dos padrões de
reprodução sexual e assexual e dos padrões de ciclo de vida
vida, incluindo as vantagens
seletivas
eletivas da própria reprodução sexual. Estas questões serão
s
discutidas em maiores
detalhes ao longo desta unidade.
Objetivos
Ao final desta unidade, o leitor deverá ser capaz de:
•
reconhecer
econhecer a importância da reprodução;
•
definir reprodução;
•
distinguir reprodução
dução assexuada de reprodução sexuada;
•
discorrer
iscorrer sobre a importância dos processos
pr
de reprodução assexuada e
reprodução sexuada, bem como enumerar as interações entre estes processos ao
longo da evolução dos Metazoa (animais pluricelulares);
•
enumerar e entender
ender os principais métodos de reprodução assexuada e
sexuada dos Metazoa;
identificar
dentificar os principais eventos, processos
proc
e mecanismos e suas inter•
relações relativas à reprodução sexuada dos Metazoa: padrões de sexualidade,
gametogênese, sistemas de determinação
determi
do sexo, desenvolvimento das gônadas e
gonodutos, fertilização, desenvolvimento direto e indireto, tipos de ovos, tipos de
larvas;
•
descrever
escrever os padrões básicos de ciclos de vida dos Metazoa;
•
descrever
escrever os principais mecanismos reprodutivos dos grandes grupos
de Metazoa.
Módulo V – Processos Reprodutivos
O Reino METAZOA ou
ANIMALIA
engloba
todos os seres vivos que
são
qualificados
tipicamente
como
animais.
148
O Processo Reprodutivo
DOIS MODELOS DE REPRODUÇÃO
DUÇÃO PODEM SER REC
RECONHECIDOS: ASSEXUADO E SEXUADO.
Na reprodução assexuada há apenas um progenitor e não há órgãos ou células
germinativas especializadas. Cada organismo é capaz de produzir cópias
geneticamente iguais de si mesmo, assim que se torna adulto.
Por outro lado, a reprodução sexuada, de maneira geral, envolve dois
progenitores, onde cada um contribui com células germinativas especializadas
(gametas), cuja fusão (fertilização) leva ao desenvolvimento de uma célula-ovo
(zigoto) que originará um novo indivíduo geneticamente diferente dos genitores. A
reprodução sexuada gera variabilidade fenotípica, o ingrediente básico para o
processo evolutivo.
Reprodução Assexuada
A reprodução assexuada é a produção de indivíduos sem envolver recombinação do
material genético. No caso dos metazoários, o que significa reprodução sem a
produção de gametas (óvulos ou espermatozóides).
esperma
A prole assim produzida terá
uma constituição genética idêntica
êntica aos organismos parentais, que inclui um grande
número de processos diferentes que serão descritos a seguir. Todos os descendentes
produzidos por meio da reprodução assexuada têm o mesmo genótipo e são
chamados de clones .
A reprodução assexuada é
também
chamada
de
reprodução clonal.
Há reprodução assexuada em muitos filos de metazoários. Nos filos animais
nos quais ocorre reprodução assexuada, a maior parte das espécies também emprega
em
a reprodução sexuada. Nesses
es grupos, a reprodução assexuada assegura o rápido
aumento de número, quando a diferenciação do organismo não atingiu o ponto de
formação de gametas. A reprodução assexuada pode ocorrer tanto pela subdivisão de
um corpo em duas ou mais partes multicelulares
multicelulares, como pela produção de ovos
diplóides.
As formas básicas de reprodução assexuada ou clonal dos metazoários são
brotamento, gemulação, fissão ou fragmentação (seguidas de regeneração) e
partenogênese, como se poderá ver a seguir:
O brotamento é a divisão desigual de um organismo. O novo indivíduo surge
como um botão (broto), crescendo a partir do progenitor, desenvolve órgãos iguais
aos dele e posteriormente se separa. O brotamento ocorre em diversos filos animais,
sendo especialmente importante em cnidários.
A gemulação é a formação de um novo indivíduo a partir de um agregado de
células envolvido por uma cápsula resistente, chamada gêmula. Em muitas esponjas
de água doce, as gêmulas desenvolvem-se
desenvolvem
no outono e sobrevivem durante o inverno
no corpo ressecado ou congelado
ado do progenitor. Na primavera, as células do
Módulo V – Processos Reprodutivos
Animais coloniais são
compostos por um grande
número de indivíduos
denominados zoóides. Os
zoóides se mantêm unidos
após
a
fissão
ou
brotamento. Este tipo de
reprodução
assexuada
(fissão ou brotamento
é
incompleto)
característico dos animais
coloniais e é responsável
pelo
crescimento das
colônias. Este tipo de
crescimento, denominado
crescimento modular, não
ocorre pelo aumento do
tamanho do corpo, mas
pela adição de novos
zoóides. Neste caso, a
149
agregado tornam-se
se ativas, emergindo da cápsula e crescendo para tornar-se
tornar
uma
nova esponja.
A fissão pode significar uma fissão binária (divisão em dois fragmentos),
sendo que cada um regenera as partes que faltam ou pode dar origem a fragmentos
múltiplos (processo chamado de fragmentação),
fragmentação sendo que cada um pode regenerar
um animal completo. A fissão é sempre seguida de regeneração e geralmente
combinada com a capacidade
dade para reprodução sexual em ciclos de vida complexos
com gerações assexual e sexual.
A partenogênese é uma forma de reprodução por meio de óvulos não
fertilizados. Há dois padrões básicos de partenogênese. No tipo denominado
partenogênese ameiótica não ocorre meiose, e o ovo é formado por divisão mitótica.
Esta é a forma assexual de partenogênese e ocorre, por exemplo, em algumas espécies
de platelmintos, rotíferos, crustáceos e insetos. Neste caso, os filhotes são clones dos
pais, já que, na ausência dee meiose,
meiose o complemento cromossômico do progenitor é
passado “intacto” para os descendentes. Ela envolve modificação da meiose de tal
modo que os óvulos são diplóides e não precisam fundir-se
fundir
com células germinativas
masculinas para dar origem a uma célula diplóide, a partir da qual pode prosseguir o
desenvolvimento. Partenogênese obrigatória (quando nunca ocorre reprodução
sexual) é rara, mas é encontrada, por exemplo, nos rotíferos bdelóideos, nos quais os
machos nunca foram observados.
os. A partenogênese ocorre
oc
frequentemente de modo
cíclico juntamente com episódios de reprodução sexual (o
( que será discutido mais
adiante). O segundo padrão básico, denominado partenogênese meiótica, pode ser
considerado reprodução assexuada ou sexuada, dependendo da participação
participa
ou não
dos machos neste processo. Este fenômeno será estudado mais adiante.
Reprodução Sexuada
Todos os seres vivos compartilham um mecanismo semiconservativo de replicação,
denominado código genético,, que inclui a replicação das moléculas de DNA. A
perpetuação dos organismos multicelulares requer não somente a replicação
semiconservativa das moléculas de DNA e das células, mas também a replicação dos
organismos, um processo que denominamos reprodução, e na grande maioria dos
organismos esse processo
ocesso também permite recombinação genética, isto é,
denominada reprodução sexual.. Entre os seres vivos, há diversos sistemas de
sexualidade, porém, eles compartilham a possibilidade de intercâmbio do material
genético entre indivíduos diferentes da mesma espécie.
Os animais adotaram um sistema de sexualidade que envolve
en
meiose e
fertilização. Estee sistema inclui organismos diplóides, ou seja, organismos que têm
dois conjuntos de cromossomos, cada um contendo informação genética codificada a
partir de cada um dos pais. Os indivíduos diplóides originarão gametas haplóides
(com um conjunto de cromossomos)
cromossomos), isto é, óvulos e espermatozóides, por um
processo chamado gametogênese (ver Fig. 1 a seguir).
Módulo V – Processos Reprodutivos
Felizmente, o processo
de
replicação
das
moléculas de DNA no
código genético não é
perfeito, pois são essas
falhas que permitem o
surgimento da variabilidade genotípica e
fenotípica.
150
Figura 1: Representação diagramática dos principais eventos celulares da gametogênese (Figura 1410, RCO** pág. 382).
Durante a gametogênese ocorre um tipo especial de divisão celular chamada
meiose. A meiose é uma divisão celular dupla. Os cromossomos separam-se
separam
uma
vez, mas as células se dividem em duas, produzindo quatro células, cada uma
contendo metade do número original
inal de cromossomos (número haplóide). A meiose
é seguida pela fertilização,, que é a fusão dos gametas masculino e feminino
formando o zigoto diplóide, cuja constituição genética difere daquela das células
diplóides que deram origem aos gametas haplóides. O zigoto combina padrões
genéticos dos dois genomas parentais em cada conju
conjunto de cromossomos. Além
disso, a fusão dos dois gametas haplóides restaura o número cromossômico normal
(diplóide) da espécie. O zigoto começa a dividir-se
dividir
por mitose para dar origem a um
indivíduo único, resultante da recombinação das características do pai e da mãe. A
recombinação genética é a grande vantagem da reprodução sexuada.
ATIVIDADE: Antes de aprofundarmos o estudo quanto à reprodução sexual
nos animais, responda às perguntas a seguir:
1.
como
omo se dá o processo reprodutivo?
2.
explique
xplique como se dá a reprodução
r
assexuada e a sexuada.
Módulo V – Processos Reprodutivos
151
Padrões de Sexualidade
A reprodução sexual nos animais sempre envolve a fusão de gametas
femininos grandes e imóveis com gametas masculinos pequenos e móveis. Este
fenômeno é denominado de anisogamia.
anisogamia Assim sendo, é possível reconhecer os
machos e as fêmeas. Porém, há outros padrões de sexualidade entre os metazoários.
metazoários
De fato, a reprodução sexuada nos metazoários segue três padrões básicos de
sexualidade, a saber: (1) gonocorismo (2) hermafroditismo e (3) partenogênese
meiótica.
1. Reprodução Bissexuada ou Gonocorismo
O fenômeno no qual os sexos são separados em indivíduos distintos é conhecido
como gonocorismo ou dioicia (ou ainda reprodução bissexuada). É a produção de
descendentes formados pela união de gametas originários de dois progenitores
(macho e fêmea) geneticamente
amente distintos. Como resultado, os descendentes possuirão
um genótipo novo, diferente do de ambos os progenitores. Os indivíduos
progenitores são de sexos diferentes, um macho e uma fêmea. Cada um deles tem seu
próprio sistema reprodutivo e produz apenas um tipo de célula germinativa,
espermatozóide ou óvulo, mas nunca ambos. Praticamente,
Praticamente todos os vertebrados e
muitos invertebrados têm sexos separados, uma condição denominada dióica ou
gonocórica.
O ESPERMATOZÓIDE é
uma
embalagem
simplificada que carrega
material
genético
altamente concentrado,
desenhado com o único
propósito de alcançar e
fertilizar um óvulo.
A distinção entre machos e fêmeas é baseada não em diferenças
dife
no tamanho ou
na aparência dos progenitores, mas no tamanho e na mobilidade dos gametas que
produzem (anisogamia). Os óvulos são produzidos pela fêmea e são grandes devido
ao acúmulo de nutrientes armazenados para manter o desenvolvimento inicial, não
n
possuem mobilidade e são produzidos em números relativamente pequenos. Os
espermatozóides são produzidos pelo macho. Espermatozóides são pequenos,
móveis e produzidos em enormes quantidades.
2. Hermafroditismo
O fenômeno no qual o mesmo indivíduo pode funcionar tanto como macho
m
e fêmea,
simultânea ou sequencialmente, é conhecido como hermafroditismo. Ao contrário do
estado dióico ou gonocórico, no qual os dois sexos são separados, os hermafroditas
são monóicos,, o que significa que tanto os órgãos femininos quanto os masculinos
estão presentes no mesmo organismo. Muitos invertebrados sésseis, subterrâneos
subterrâ
ou
endoparasitos (por exemplo, muitos platelmintos, alguns hidróides e anelídeos), e
uns poucos vertebrados (alguns peixes),
peixes) são hermafroditas. Alguns hermafroditas
fertilizam-se a si mesmos (hermafroditas
hermafroditas simultâneos).
simultâneos Este fenômeno é conhecido
como autofertilização,, entretanto,
entretanto a maioria das espécies hermafroditas não
apresenta os dois papéis sexuais ao mesmo tempo, mas um e em sequência o outro.
Estas espécies são hermafroditas sequenciais
sequ
(peixes de corais). Aquelas que iniciam
a vida como macho e se transformam mais tarde em fêmea são hermafroditas
protândricos; aquelas que iniciam a vida como fêmea e se transformam mais tarde
em macho são hermafroditas protóginos.
protóginos
Módulo V – Processos Reprodutivos
Geralmente o hermafroditismo sequencial é
determinado
geneticamente
(peixes
de
corais), ambientalmente,
ou hormonalmente. Por
exemplo: o gastrópodo
Crepidula
fornicata
se
agrupa em pilhas: o
indivíduo
inferior
é
sempre uma fêmea e os
indivíduos superiores são
machos, enquanto os
animais intermediários na
pilha
podem
ser
hermafroditas. Um mecanismo
de
feromônio
provavelmente
está
envolvido no mecanismo
de determinação do sexo
desses animais.
152
3. Partenogênese Ameiótica
A partenogênese é o desenvolvimento de um embrião a partir de um óvulo
não fertilizado, ou de um óvulo no qual os núcleos feminino e masculino deixam de
unir-se após a fertilização.
Como já vimos anteriormente, há dois tipos básicos de partenogênese:
ameiótica e meiótica.
Partenogênese ameiótica: é um fenômeno de reprodução assexuada e envolve a
formação mitótica de um óvulo diplóide sem a participação do macho.
Partenogênese meiótica: um ovo haplóide é formado por meiose, podendo ou
não ser ativado pela influência de um macho (podendo ser um fenômeno de
reprodução sexuada ou assexuada, respectivamente). Por exemplo, em algumas
espécies de peixes, a fêmea pode ser inseminada por um macho da mesma espécie,
ou de uma espécie relacionada, mas os espermatozóides servem apenas para ativar o
óvulo; o material genético do macho é rejeitado antes que possa penetrar no gameta
feminino. Em diversas espécies de platelmintos, rotíferos, anelídeos, ácaros e insetos,
o óvulo haplóide inicia seu desenvolvimento espontaneamente, não sendo
necessários machos para estimular a ativação de um ovo. A condição diplóide é
restaurada pela duplicação cromossômica.
A partenogênese é comum entre os animais. É uma simplificação dos passos
normalmente necessários à reprodução bissexuada. Pode ter evoluído para contornar
o problema, em alguns animais, de colocar juntos o macho e a fêmea no momento
certo para que a fertilização ocorra com sucesso.
A desvantagem da partenogênese é a capacidade limitada de gerar
variabilidade fenotípica e, consequentemente, baixa capacidade de adaptação frente a
novas condições que porventura venham a ocorrer.
Espécies bissexuadas, em virtude da recombinação gênica, têm uma chance
maior de produzir proles com variabilidade fenotípica geralmente suficientes para se
adaptar a novas condições ambientais.
Gônadas e Gonodutos
Os órgãos que produzem os gametas são chamados de gônadas. A gônada que
produz espermatozóides é o testículo, e a que forma óvulos é o ovário. As gônadas
representam os órgãos sexuais primários, os únicos órgãos sexuais encontrados em
determinados grupos de animais.
Os Poriferas (esponjas) e os Placozoa não possuem gônadas nem gonodutos (na
verdade, as esponjas e os placozoários não possuem nenhum órgão). No caso das
esponjas, os gametas são formados a partir de arqueócitos ou coanócitos
indiferenciados do meso-hilo (que dão origem aos óvulos e a partir de coanócitos -que formam espermatozóides). A reprodução sexuada dos Placozoa não é bem
compreendida e não se tem certeza se ela ocorre.
Módulo V – Processos Reprodutivos
Veja no endereço
abaixo
mais
informações sobre os
Porifera (esponjas) e
os Cnidaria :
http://www.portalb
rasil.net/educacao_s
eresvivos_invertebad
153
Entretanto, os Eumetazoa, isto é, os demais metazoários, possuem conjuntos de
células germinativas
nativas ectodérmicas, mesodérmicas ou endodérmicas responsáveis
pela produção dos gametas. Estes conjuntos de células germinativas são chamados de
gônada.
As gônadas podem ser simplesmente um aglomerado de células germinativas
ou podem ser mais complexas e revestidas por uma camada epitelial, formando
órgãos diferenciados e individualizados ger
geralmente pares (testículo ou ovário).
Nos Cnidaria, as gônadas são agregadas
agregad de células germinativas e se originam
na endoderme e, geralmente, crescem e se diferenciam na gastroderme, exceto em
alguns hidrozoários, nos quais elas migram para a epiderme. Os cnidários não
possuem gonodutos e os gametas são liberados dentro da cavidade do corpo e
expelidos posteriormente pela boca.
Os Ctenophora são os primeiros metazoários a apresentar um gonoduto e os
gametas saem das gônadas por canais curtos que conduzem a uma série de poros
chamados gonóporos, que se abrem para o exterior do corpo.
A maioria dos demais Eumetazoa, ou seja, a maioria dos Bilateria (animais com
simetria bilateral) possui gônadas geralmente individualizadas que se originam na
mesoderme.
As gônadas dos bilatérios apresentam três formas gerais de organização (ver
Fig. 2 a seguir)
Figura 2: Bilatéria: anatomia gonadal e oviposição. (A a D) Metade da seção transversal. (A)
Gônada na região do corpo com celoma e metanefrídio: gônada no mesotélio, gametas lançados dentro do
celoma, liberados pelo metanefrídio (Lophophorata, Trochozoa). (B e C) Gônada saculiforme com seu
próprio gonoduto em celomados que não possuem metanefrídios (B, deuterostômios) e em táxons com
hemocele ou corpo compacto
cto (acelomados) (C). (D) Gônada associada a tecido compacto, principalmente
em bilatérios de pequeno porte: gônadas ausentes, substituídas pela simples agregação de células
germinativas, gametas maduros saem através de gonodutos (não ilustrados) ou pela ruptura da parede do
corpo (Figura 10.47, RFB pág. 301).
1.
Se há celoma e metanefrídio, a gônada ocorre no mesotélio, libera
gametas no celoma e elimina-os
os através do metanefrídio (A).
2.
Se a região gônada está na hemocele, tecido celular compacto, ou
celoma sem um metanefrídio, a gônada está encerrada em sua própria cavidade e é
revestida por um mesotélio que contém células germinativas. Estas gônadas se abrem
para o exterior por meio de um gonoduto próprio (B, C).
3.
Animais muito pequenos sem cavidades internas (hemocele ou
celoma), isto é, preenchidos por tecido compacto, apresentam gônadas que são meras
agregações de células germinativas que não possuem um epitélio de revestimento.
Nestes,, os gametas são liberados através de um gonoduto ou pela ruptura da parede
do corpo (D).
Módulo V – Processos Reprodutivos
A mesoderme é o
folheto embrionário,
ou seja, uma das
camadas
que
iniciam
a
diferenciação
das
células no embrião,
dando origem, entre
outros órgãos, ao
coração.
154
Com poucas exceções, os bilatérios apresentam gonodutos. O gonoduto dos
bilatérios de fertilização interna é complexo e inclui uma série de órgãos acessórios
que estocam, nutrem, transportam e encapsulam os gametas.
Machos geralmente possuem glândulas secretoras para nutrição do esperma.
Algumas espécies possuem glândula espermatofórica que secreta uma cápsula
protetora, chamada espermatóforo, ao redor de um grupo de espermatozóides. O
espermatóforo pode ser fixado a um substrato e mais tarde coletado pela fêmea, ou
pode ser transferido diretamente para a fêmea. Outros órgãos acessórios masculinos
são a vesícula seminal, que armazena espermatozóides, e o pênis especializado, que
ajuda na transmissão de esperma para a fêmea.
Órgãos acessórios femininos comuns são a bursa copulatória e o receptáculo
seminal, que armazenam espermatozóides por curtos e longos períodos,
respectivamente. Bilatérios grandes de fertilização interna ou liberam o zigoto na
água, ou o incubam internamente (gestação), geralmente numa expansão distal do
oviduto chamada de útero, ou o incubam externamente, ou o envolvem numa
cápsula protetora. Os bilatérios muito pequenos geralmente são vivíparos ou
depositam seus zigotos no substrato ou em cápsulas.
Gametogênese
Nos animais, geralmente, dois tipos de gametas são produzidos. Estes são
derivados de células especializadas que foram segregadas precocemente no
desenvolvimento embrionário (gônadas), ou derivadas a partir de células somáticas
desdiferenciadas (Porifera). O processo de formação de gametas masculinos
(denominados espermatozóides) recebe o nome de espermatogênese, e a ovogênese
é o nome dado ao processo de formação de gametas femininos (denominados
óvulos).
A espermatogênese, na maioria dos metazoários, envolve divisões mitóticas
antes do início da meiose, para originar um grande número de células. Cada
espermatogônia, que se transforma em um espermatócito primário, dará origem a
quatro espermátides, as quais, por um processo de diferenciação, dão origem aos
espermatozóides (ver Fig. 1).
Para melhor visualização, apresentamos a seguir novamente a Figura 1.
Módulo V – Processos Reprodutivos
155
Figura 1: Representação diagramática dos principais eventos celulares da gametogênese (Figura 1410, RCO** pág. 382).
A presença de espermatozóides é um caráter único, uma novidade evolutiva
compartilhada (sinapomorfia) pelos metazoários (talvez a maneira mais fácil de
definir os animais é defini-los como seres que possuem espermatozóides).
Observe na Fig. 3 a seguir que muitos metazoários têm espermatozóides
semelhantes, com corpo celular arredondado e um flagelo posterior (a), mas outros
possuem tipos mais avançados (b-d). Alguns metazoários apresentam
espermatozóides sem flagelos que se movem por movimento amebóide
(Gnathostomulida, Nematoda, Nematomorpha, Crustacea, Diplopoda). Os flagelos
dos espermatozóides dos Rotifera e Acanthocephala são anteriores. Espermatozóides
mais avançados estão associados com ciclos de vida nos quais há armazenamento do
esperma ou fecundação interna, ou nos quais os ovos são muito bem protegidos.
Figura 3: (A) Espermatozóide do tipo
primitivo cuja cabeça é arredondada. (B- D)
Espermatozóides mais avançados (Figura 13.29,
RFB pág. 509).
Durante a ovogênese (Fig. 1),
reservas alimentares são depositadas
no óvulo em desenvolvimento. Em alguns metazoários os ovócitos (células
precursoras do óvulo) em desenvolvimento são células solitárias que ficam suspensas
livremente no líquido celomático. Os ovócitos se desenvolvem e elaboram seu
Módulo V – Processos Reprodutivos
Visite
o
site
Educação Pública do
Governo do RJ e
conheça mais sobre
caracteres compartilhados em:
http://www.educac
aopublica.rj.gov.br/
oficinas/ed_ciencias
/peixes/onde/comp
artilhando/caractere
s_e_homoplasias.ht
ml
156
próprio vitelo sem qualquer associação com outras células e flutuam livremente na
cavidade celomática. Tal padrão é descrito como ovogênese solitária.
Apenas para facilitar o estudo, segue abaixo novamente a Figura 1.
Figura 1: Representação diagramática dos principais eventos celulares da gametogênese (Figura 1410, RCO** pág. 382).
Nesses casos, as reservas alimentares do futuro óvulo são incorporadas pelo
próprio ovócito como precursores de baixo peso molecular e elaborados
posteriormente em moléculas complexas de alto peso molecular, denominadas
coletivamente de vitelo. Este processo é chamado autossíntese. O vitelo, portanto, é a
reserva alimentar do óvulo (a gema do ovo das galinhas, por exemplo, é formada
pelo vitelo). Esse tipo de ovogênese acontece em metazoários com amplas cavidades
celomáticas, por exemplo, nos Echiura, Sipuncula e em alguns poliquetos.
A maioria dos metazoários, contudo, apresenta uma ovogênese folicular ou
nutrimental, na qual os ovócitos em desenvolvimento estão intimamente associados
a células somáticas (denominadas células foliculares), ou a outras células
germinativas (chamadas células subsidiárias).
A maior parte das moléculas de alto peso molecular, depositada no citoplasma
do ovócito, é sintetizada não pelo próprio ovócito, mas pelas células somáticas ou
subsidiárias em outro lugar do corpo. Tais moléculas são então transportadas para os
ovócitos pelos líquidos do corpo ou pelo sistema circulatório. Este padrão da
ovogênese é descrito como heterossíntese e ocorre, por exemplo, em insetos,
crustáceos e moluscos.
Nos Turbellaria, o “ovócito” é frequentemente uma estrutura complexa,
composta por uns poucos ovócitos verdadeiros com pouco vitelo, associados a células
subsidiárias repletas de vitelo. Toda esta estrutura está contida num envoltório
Módulo V – Processos Reprodutivos
157
protéico esclerotizado. Este tipo de ovogênese, onde o vitelo não é armazenado no
ovócito, mas em células subsidiárias, é denominado nutrimental e ectolécito.
Os demais padrões (ovogênese solitária e folicular) são chamados de
endolécito, pois o vitelo é armazenado no citoplasma do ovócito.
Mecanismos de Determinação do Sexo
A determinação do sexo dos metazoários apresenta componentes materno, genético e
ambiental. No caso da determinação materna, o sexo dos filhos é determinado pela
mãe através da produção de diferentes tipos de ovócitos (Fig. 4).
Figura 4: Determinação materna do sexo e ciclo de vida de Dinophylus gyrociliatus (Annelida,
Polychaeta) -- (Figura do Quadro 14-3(1), RCO pág. 370).
Por exemplo, os ovários das fêmeas adultas de Dinophylus gyrociliatus
(Polychaeta) produzem dois tipos de ovócitos:
• ovócitos grandes tornam-se fêmeas; e,
• ovócitos menores tornam-se machos anões precocemente maduros.
Ocorre inseminação dos embriões femininos no casulo. Todos os adultos são
fêmeas inseminadas.
Além da determinação materna, existe um componente genético envolvido,
pois o macho de D. gyrociliatus produz espermatozóides masculino e feminino. Os
espermatozóides femininos (espermatozóides-X) têm um cromossomo que está
ausente nos espermatozóides masculinos (espermatozóides-0). Espermatozóides-X
selecionam ovócitos grandes (femininos) e espermatozóides-0 selecionam ovócitos
pequenos (masculinos).
Outro exemplo são as abelhas. A abelha rainha é capaz de controlar a
fecundação dos seus ovos (determinação materna); ovos diplóides fecundados
desenvolver-se-ão em condições normais em fêmeas operárias estéreis (determinação
genética), mas têm a capacidade em potencial de se tornarem fêmeas funcionais se
expostas às condições apropriadas durante o desenvolvimento (determinação
ambiental), ao passo que os ovos não fecundados darão origem, por partenogênese,
aos machos haplóides (determinação genética).
Módulo V – Processos Reprodutivos
158
O exemplo das abelhas ilustra um caso de determinação genética do sexo
chamado haplodiploidia,, pois o que determina o sexo é a condição haplóide
(machos) e diplóide (fêmea).
O exemplo dado do poliqueto (Dinophylus gyrociliatus), acima, ilustra um caso
de determinação genética do sexo chamado heterogamia, pois o que determina o
sexo é a fusão de gametas que diferem entre si quanto à sua constituição
cromossômica. Nestee exemplo, as fêmeas apresentam um cromossomo que é
chamado sistema XX-XO.
Em algumas outras espécies animais, machos e fêmeas apresentam d
diferentes
complementos cromossômicos. As fêmeas têm cromossomos sexuais idênticos,
idênticos
chamados cromossomos X, e os machos cromossomos diferentes,
diferentes um cromossomo X
e um cromossomo Y. O sexo do zigoto é determinado pelo complemento
cromossômico do espermatozóide fecundante. Neste caso, o gameta masculino
determina o sexo. O chamado sistema XX
XX-XY ocorre, por exemplo, em insetos e
craniados.
Outro sistema de determinação do sexo é o chamado sistema ZZ-ZW. É um
sistema semelhante ao XX-XY; porém,
porém o sexo é determinado pelo gameta feminino
que possui um cromossomo
mossomo Z e um cromossomo W. Este
Est sistema ocorre, por
exemplo, em lepidópteros, aves e peixes.
Poliquetos, do grego
polys (muitos) + chaeta
(pelo, espinho, cerda),
geralmente
de
vida
marinha ou costeira, é a
maior classe do filo
Anelida, Phylum, que
compreende os animais
que se caracterizam, em
especial,
por
apresentarem os seus
corpos
segmentados,
interna e externamente.
Consultado
no
site
http://www.algosobre.co
m.br/biologia/anelidios.h
tml em 23/10/09.
Uma consequência
ência da heterogamia é a manutenção da proporção sexual 1:1.
Tal manutenção é de extrema importância,
importância porque descendentes machos ou fêmeas
oferecem caminhos equivalentes para o sucesso reprodutivo em organismos
diplóides com gametas haplóides. Além disto,
dis a proporção sexual 1:1 garante maior
sucesso reprodutivo, pois o investimento nas funções de macho e de fêmea,
fême que estão
se reproduzindo sexualmente, será normalmente igual e mantém inalterada
ina
na
geração descendente.
O sexo de um indivíduo muitas vezes depende das condições ambientais que
são experimentadas pelo embrião ou larva em desenvolvimento.
Um dos exemplos
los melhor conhecidos é do verme equiúro Bonellia viridis, cujos
machos são anões e parasitas sobre as fêmeas. As fêmeas liberam uma substância
virilizante, na presença da qual todas as larvas tomam-se
tomam
machos. O macho anão vive
no saco genital da fêmea. O macho fertiliza os óvulos durante a ovoposição. Os ovos
dão origem a larvas trocóforas. Caso estas
es
larvas assentem-se no prostômio de uma
fêmea ou próximo dele, darão origem a machos anões; caso assentem-se
assentem
longe de
uma fêmea, então, darão origem a fêmeas. As larvas induzidas a se transformar em
machos desenvolvem um órgão adesivo pelo qual se fixam ao prostômio da fêmea.
Os Echiura são gonocóricos, mas o processo de sexualidade entre muitos
hermafroditas sequenciais
enciais é também profundamente influenciado pelas condições
ambientais ou hormonais.
Por exemplo, a Crepidula fornicata (Gastropoda) agrupa-se em pilhas: o
indivíduo inferior é sempre uma fêmea,
fêmea e os indivíduos superiores são machos,
enquanto os animais intermediários na pilha podem ser hermafroditas (ver Fig. 5).
Figura 5: Determinação do sexo em Crepidula fornicata (Mollusca, Gastropoda). (i) Formação inicial
de um par. (ii, iii) Associações mais tardias e complexas de machos, fêmeas e intersexuados (Figura 14-7,
RCO pág. 371).
Módulo V – Processos Reprodutivos
Um exemplo interes-sante
de
determinação
ambiental do sexo ocorre
nos
croco-dilianos,
quelônios, e lagartos.
Nestes animais o sexo da
prole é determinado pela
temperatura do ninho. Por
exemplo, os ovos de
jacarés
incubados
a
tempera-turas
mais
baixas, darão à luz a
fêmeas, enquanto que os
ovos em ninhos com
temperaturas mais altas
darão origem a machos.
159
Um mecanismo de feromônio provavelmente está envolvido no mecanismo de
determinação do sexo
xo desses animais.
Ciclos de Vida
O ciclo de vida dos Metazoa geralmente inclui um adulto multicelular que é
composto por muitas células diplóides. Adultos multicelulares dão origem a gametas
haplóides. A fusão determina um zigoto diplóide, que se origi
originará um indivíduo
adulto.
Em alguns casos, pode haver uma alternância bem marcada entre fases
diplóides que se reproduzem sexuadamente e assexuadamente
assex
por partenogênese.
Estes
es ciclos de vida incluem uma ou muitas gerações de indivíduos que se
reproduzem partenogeneticamente e são seguidas por uma geração de indivíduos
sexuados que, usualmente, dão origem a ovos resistentes. Nestes
Nest ciclos de vida, um
indivíduo diplóide, que emerge de ovo resistente, dá origem a um clone,
clone cujos
descendentes são geneticamente iguais.
Nessas circunstâncias, a taxa de crescimento da população está no máximo e o
ciclo de vida pode ser considerado como um meio para uma exploração máxima de
recursos alimentares,
limentares, temporariamente subaproveitados,
sub
e quando outros fatores
biológicos limitam o tamanho do corpo. Em determinado estágio, um novo tipo de
indivíduo surge na população e apresenta dois tipos de descendência, fêmeas e
machos sexualmente reprodutivos.
A mudança é uma resposta a modificações das
d condições ambientais, tais como
superpopulação, qualidade alimentar ou diminuição da duração dos dias (veja ao
lado o exemplo dos rotíferos Bdelloi
Bdelloidea).
O zigoto, ou célula-ovo
ovo de algumas espécies animais,
animais desenvolve-se
diretamente em jovens. Estee tipo de desenvolvimento é denominado
desenvolvimento direto.
Entretanto, muitas espécies de metazoários apresentam um estágio larval em
seu ciclo de vida.
O estágio larval é uma etapa de desenvolvimento temporária que vive
independentemente do estágio adulto. Difere deste em forma e ocupa um nicho
ecológico diferente. Animais com um ciclo de vida que inclui uma fase larval têm
desenvolvimento indireto.. Muitos metazoários têm em seu ciclo de vida um adulto
bentônico e uma larva planctônica.. Ao final do período larval, a larva instala-se
instala
em
um habitat satisfatório para o adulto e, então, sofre uma metamorfose e dá origem ao
estágio juvenil, que difere do adulto apenas por seu tamanho e imaturidade sexual.
Os Metazoa possuem dois tipos básicos de estágios larvais. Larvas com vida
curta geralmente alimentam-se pela absorção do vitelo do ovo e são chamadas
lecitotróficas.
A maioria das larvas é, inicialmente,
inicialmente lecitotrófica, mas logo que o intestino é
formado, a larva então passa a se alimentar de plâncton. Tais larvas são chamadas
planctotróficas.
Animais cujo ciclo de vida inclui uma fase larval planctotrófica geralmente
produzem muitos ovos microlécitos
icrolécitos (com pouco vitelo).
Animais cujo ciclo de vida inclui uma fase larval lecitotrófica geralmente
produzem um menor número de ovos ricos em vitelo (ovos macrolécitos). Como a
larva lecitotrófica alimenta-se apenas de vitelo, seu período de vida é relativamente
curto.
Módulo V – Processos Reprodutivos
Os rotíferos Bdelloidea
apresentam uma forma
extrema de diapausa;
seus ovos encistados
podem permanecer por
anos
no
que
é,
virtualmente, um estado
de animação suspensa,
até que eles sejam
carregados pelo vento a
lugares apropriados para
o
crescimento.
Esta
capacidade pode estar
relacionada
com
a
ausência da reprodução
sexuada;
evitando
condições desfavoráveis,
eles
vivem
para
usufruírem o que são, na
verdade,
permanentes
condições
ideais.
Provavelmente eles serão
menos
visados
por
parasitas
e
por
organismos que causam
doenças,
e
assim
escaparão das condições
que
se
acredita
favorecem a reprodução
sexual.
160
Por outro lado, a larva planctotrófica tem um período de vida mais longo,
longo pois
pode alimentar-se do plâncton e não depende do vitelo para sua sobrevivência
(exceto na fase muito inicial do seu desenvolvimento,
desenvolvimento quando o trato digestivo ainda
não se formou).
se que um ciclo de vida indireto com fertilização externa, adulto
Acredita-se
bentônico e larva planctotrófica, seja o padrão plesiomórfico dos metazoários, e que
os demais padrões de ciclos de vida apareceram mais tarde, ao longo da história
his
evolutiva dos Metazoa.
Ao longo da história dos Metazoa os padrões de ciclos de vida desenvolveramdesenvolveram
se como
omo respostas adaptativas face às pressões seletivas exercidas pelas condições
adversas exibidas por diferentes
rentes habitats. Deste modo, os ciclos de vida e as
estratégias reprodutivas são diferent
diferentes no ambiente marinho, dulciaquícola e
terrestre. Os ciclos de vida dos muitos metazoários marinhos podem ser classificados
em bentônico-pelágico,
pelágico, holopelágicos e holobentônicos, como detalhado a seguir.
Um estágio larval pelágico planctotrófico é característico do ciclo de vida de
muitos animais marinhos bentônicos.
De fato, a maioria dos filos de metazoários possui, ao menos, algumas espécies
com larvas pelágicas, o que é vantajoso para as espécies bentônicas,
bentônicas cujos adultos são
de locomoção lenta ou sésseis, porque larvas planctotróficas exibem grande
capacidade de dispersão. Estes
es animais exibem um ciclo de vida bentônico-pelágico.
Como vimos acima, estee é o padrão plesiomórfico
ples
dos metazoários. Um menor
número de espécies na maioria dos filos tem larvas pelágicas que não se alimentam e
que são supridas com vitelo pelo organismo parental.
Tais larvas apresentam desenvolvimento pelágico lecitotrófico. Esses animais
geralmente
ente exibem um ciclo de vida holopelágico. Outro grupo de espécies possui
larvas bentônicas, ou suprimiram completamente a fase larval,
larval e apresentam
desenvolvimento direto. Estes
es animais exibem um ciclo de vida holobentônico.
A opção por desenvolvimento planctotrófico
pl
ou lecitotrófico e
desenvolvimento direto, que é um aspecto do desenvolvimento no mar, não se
apresenta para os animais que habitam ambientes terrestres e de água doce. O
estresse osmótico e a luta contra a dessecação nesses habitats pressupõem
pressupõe uma série
de adaptações relacionadas à reprodução. A fecundação deve ser interna exigindo
acasalamento direto entre parceiros. Igualmente, os embriões em desenvolvimento
devem ser protegidos da perda da água ou do estresse osmótico, por exemplo, devem
estar
tar envoltos por uma cobertura impermeável
imper
ou por um casulo.
Consequentemente,
entemente, a fase larval é suprimida e o desenvolvimento é direto.
Muitas espécies não marinhas são vivíparas, apresentando incubação ou
cuidado à prole. Geralmente espécies de água doce apresentam fecundidade
reduzida (como resultado do maior investimento de recursos enérgicos na prole). Se
desconsiderarmos
rarmos os artrópodos, podemos dizer que o hermafroditismo é mais
frequente em água
a doce e em ambiente terrestre.
Os ambientes de água doce e terrestres são mais extremos em termos de
variáveis físicas do que os marinhos, e a maioria dos invertebrados que os habitam
tem
m a capacidade de entrar em um estágio de dormência fisiológica,
fisiológica conhecido como
diapausa.
Nessa condição, o organismo pode perder água e suportar condições extremas;
a exigência metabólica é diminuída e a necessidade de uma fonte energética externa
e
pode ser nenhuma, de tal modo a suportar a ausência periódica de alimento que
ocorre em regiões temperadas e polares. Algumas espécies longevas, tais como
alguns moluscos pulmonados e anelídeos clitelados, entrarão em diapausa como
adultos, mas muitas
as formas pequenas o fazem como ovos. Assim, muitas populações
Módulo V – Processos Reprodutivos
Hermafroditismo: os
ciclos de vida dos
platelmintos de água
doce,
anelídeos
clitelados e moluscos
pulmonados,
são
surpreendentemente
semelhantes. Eles são
hermafroditas
simultâneos,
com
comportamento sexual
complexo,
com
glândulas
acessórias
muito especializadas e
com a capacidade de
protegerem
seus
embriões. Seus ciclos
de
vida
requerem
longos períodos de
atividade de postura
161
de sanguessugas somente ocorrem como embriões no interior de casulos durante o
inverno.
O hábito de hibernar como ovos de resistência é particularmente característico
de formas diminutas, tais como rotíferos
rotífer e cladóceros. Estes organismos exibem um
modo de vida no qual há uma alternância entre gerações sexual e assexual; os ovos
de resistência, que surgem por reprodução sexual, eclodem na primavera para dar
origem a novas proles clonais que podem explorar os
o novos recursos.
Os rotíferos Bdelloidea apresentam uma forma extrema de diapausa; seus ovos
encistados podem permanecer por anos no que é, virtualmente, um estado de
animação suspensa, até que eles sejam carregados pelo vento até lugares apropriados
para o crescimento. Tal capacidade pode estar relacionada com a ausência da
reprodução sexuada; evitando condições desfavoráveis eles vivem para usufruírem o
que são, na verdade, permanentes condições ideais. Provavelmente,
Provavelmente eles serão menos
visados por parasitas
tas e por organismos que causam doenças ee, assim, escaparão das
condições que se acreditaa favorecer a reprodução sexual.
Uma hipótese para essa
diapausa dos rotíferos
Bdelloidea é para que,
provavelmente,
eles
sejam menos visados
por parasitas e por
organismos que causam
doenças
e
assim
escapem das condições,
que
se
acredita,
favorecem a reprodução
sexual.
Os artrópodos (com exceção dos crustáceos, xifosuros e picnogônidos) e os
craniados tetrápodos adaptaram-se
se de modo bem-sucedido
bem
aos ambientes terrestres
e, particularmente, em alguns casos,
casos adaptaram-se secundariamente ao ambiente
dulciaquícola como adultos ou como
mo formas larvais ou de ninfas.
A biologia reprodutiva desses grupos difere de um modo importante daquela
dos invertebrados de corpo mole e são muito raramente hermafroditas; a maioria das
espécies se reproduz sexualmente e os sexos são sempre separados. Os seus sucessos
devem-se, em grande parte, à presença de um revestimento
revestim
impermeável, mas
também é resultado das suas habilidades de colocarem ovos impermeáveis
resistentes à dessecação. Tais ovos devem ser fecundados antes da postura; por
consequência, estes grupos de animais geralmente apresentam
apre
fecundação interna,
frequentemente
entemente associada com comportamento copulador complexo. A prevalência
do gonocorisrno
ocorisrno parece ser uma consequência
consequ
de sua grande mobilidade.
ATIVIDADE:
1. Enumere as interações que ocorrem entre os processos de reprodução
assexuada ao longo da evolução dos Metazoa.
2. Quais os principais métodos de reprodução assexuada e sexuada dos
Metazoa? Explique cada um.
3. Descreva os principais eventos, processos e mecanismos que ocorrem na
reprodução sexuada dos Metazoa.
Mecanismos Reprodutivos dos Metazoa
Os mecanismos
mos e órgãos de reprodução nos metazoários são os mais diversos
possíveis. Tal fato
ato está diretamente associado à grande diversidade de forma que os
Metazoa abrigam, de sorte que esta seção não pretende detalhar todos os mecanismos
e órgãos reprodutivos dos animais, mas pretende apenas dar uma visão geral acerca
dos principais órgãos e mecanismos de reprodução nos diversos grupos metazoários.
1. Porifera: esponjas
sponjas reproduzem-se
reproduzem
assexuadamente por fragmentação,
brotamento e pela formação de propágulos de resistência,
re
chamados de gêmulas.
Módulo V – Processos Reprodutivos
Dulciaquícola: ser vivo
cujo hábitat é a água doce.
162
Esponjas, com poucas exceções, são hermafroditas. A fecundação é interna. Algumas
espécies são ovíparas e liberam zigotos na água, onde eles completam seu
desenvolvimento. A maioria das esponjas é vivípara, retendo os zigotos
zigoto no corpo
parental e liberando larva. Embriões e larvas são lecitotróficas. As larvas que se
desenvolvem de embriões também são diversas e descritas com os nomes de larvas
celoblástula, anfiblástula, parenquimela e triquimela (Fig. 6).
Figura 6: Porifera:
ra: reprodução sexuada, desenvolvimento larval e metamorfose. Em A, Clathrina,
uma blástula oca desenvolve-se no meso-hilo
hilo e é liberada na água. Depois da liberação, a ingressão celular
converte a celoblástula em estereoblástula, a fase de assentamento. Em B, Sycon, o óvulo é derivado de um
coanócito que perde seu flagelo e afunda no meso-hilo
meso
abaixo da coanoderme. Depois da fertilização, o ovo
(zigoto) divide-se
se para formar uma esfera flagelada interiormente, que se assemelha a uma câmara
coanocítica. Na liberação da esponja, a esfera se inverte para posicionar seus flagelos na superfície da larva.
Durante a metamorfose,
morfose, as células flageladas externas, depois de perder flagelos, retomam ao interior do
corpo e diferenciam-se
se em coanócitos. O jovem metamorfoseado de uma esponja calcária é chamado de
olinto. Em C, Haliclona, parenquimela diferenciada é liberada da esponja
es
no plâncton. Depois do
assentamento, ela sofre metamorfose complexa para formar um jovem, ou rágon (Figura 5.16, RFB pág.
107).
Todas as larvas de esponja são lecitotróficas. Depois de certo período de tempo,
as larvas rastejam sobre o fundo à procura
procu de um local satisfatório para fixação. Uma
vez que o local seja encontrado, a larva transforma-se
transforma
em uma esponja jovem que
difere um pouco para cada um dos tipos larvais Quando a esponja jovem fica
funcional, começa a alimentar-se,, e é um asconóide em miniatura
m
chamado de olinto
(Fig. 6). Quando a esponja jovem tem uma configuração inicial asconóide ou
siconóide, porém, com paredes espessas, antes de transformar-se
transformar
em uma esponja
leuconóide, esta fase juvenil é chamada de rágon (ver Fig. 6).
reproduzem
por fragmentação e por brotamento.
2. Placozoa: Placozoários reproduzem-se
Suspeita-se
se que haja reprodução sexuada, mas isto ainda é duvidoso.
3. Cnidaria: Os cnidários podem reproduzir-se
reproduzir
por fragmentação, fissão e
brotamento. Pólipos de antozoários exibem todas as formas de reprodução assexual
citadas acima; pólipos de cifozoários apresentam brotamento e fissão transversal e
pólipos de hidrozoários sofrem brotamento. Algumas medusas de Hydrozoa
reproduzem-se
se por fissão. Os adultos de cnidários geralmente são
s gonocóricos, mas
algumas espécies são hermafroditas. As células germinativas originam-se
originam
na
endoderme e, geralmente, crescem e diferenciam-se
diferenciam
na gastroderme, exceto em
Módulo V – Processos Reprodutivos
Os cnidários têm
uma história muito
antiga e o primeiro
fóssil encontrado em
Ediacara, no sul da
Austrália, data do
Pré-cambriano (ap.
700 m.a ). Em sua
longa
história
evolutiva
têm
desempenhado
importante
papel
ecológico, do mesmo
modo que os recifes
de corais o fazem na
atualidade.
Consultado
em:
163
alguns hidrozoários, nos quais elas migram para a epiderme. Adultos sexualmente
maduros
os geram seus gametas e, primitivamente, a fertilização é externa na água. O
desenvolvimento do zigoto resulta em uma larva planctônica conhecida como
plânula (Fig. 7).
Figura 7: Cnidaria: ciclo de vida primitivo, pólipo adulto – gametas - plânula - pólipo jovem. Após
certo período de tempo, a plânula se fixa ao fundo e se metamorfoseia em um jovem. O ciclo de vida
primitivo do cnidário ancestral consiste em um pólipo adulto, bent
bentônico e séssil e uma larva plânula
planctônica que se metamorfoseia em um pólipo jovem (Figura 7.14H, RFB pág. 145).
A plânula dos antozoários pode ser planctotrófica ou lecitotrófica, entretanto,
as larvas de Scyphozoa e Hydrozoa são lecitotróficas. A fase
fa medusóide fornece
ampla capacidade de dispersão, além de realizar a reprodução sexuada. O pólipo
passa a ser o responsável pela reprodução assexuada ou crescimento modular.
4. Ctenophora: a reprodução assexuada é rara nos ctenóforos, só ocorre em
algumas
mas espécies bentônicas, que se reproduzem por fragmentação. Quase todas as
espécies de ctenóforos são hermafroditas. Os gametas são liberados e os óvulos são
fertilizados na água do mar. A fertilização cruzada é provavelmente típica da maioria
dos ctenóforos,
ros, mas a autofertilização ocorre em algumas espécies. O
desenvolvimento embrionário é direto e origina uma forma jovem planctônica,
chamada cidipídeo.
5. Orthonectida: com raras exceções, ortonéctidos são gonocóricos e vivíparos.
A fertilização é interna após cópula. Os zigotos se desenvolvem em uma larva ciliada
simples formada de poucas células.
6. Dicyemida: Os diciemídeos são parasitas dos rins de cefalópodos. O estágio
adulto do parasita é denominado nematógeno.
nematógeno Nematógenos reproduzem-se clonal
ou sexuadamente. Uma vez que o parasita esteja estabelecido no rim do hospedeiro, a
reprodução assexuada aumenta rapidamente
pidamente o número de nematógenos através da
produção de nematógenos-filhos por partenogênese. Uma célula reprodutiva ou
axoblástica do nematógeno progenitor se desenvolve partenogeneticamente em um
nematógeno-filho.
filho. Quando a densidade aumenta muito, os nematógenos
ne
reproduzem-se sexuadamente e liberam larvas infectantes (larva
(
infusoriforme), que
são liberadas através da urina do hospedeiro. O restante do ciclo é desconhecido, pois
as larvas infusoriformes não infectam outros cefalópodos.
7. Platyhelminthes: muitos turbelários,
turbelários especialmente espécies de água doce,
Módulo V – Processos Reprodutivos
Os vermes achatados
PLATYHELMINTHES
164
reproduzem-se por regeneração, fissão, fragmentação, ou brotamento. A maioria dos
turbelários é hermafrodita. Ocorrem cópula e fertilização interna. Os sistemas
masculinos e femininos são complexos e muito variáveis. Impregnação hipodérmica
ocorre em algumas espécies. O pênis, que contém estiletes, é empurrado através da
parede do corpo do cônjuge, depositando espermatozóides dentro do parênquima.
Os espermatozóides, então, migram para os ovários. Turbelários possuem
desenvolvimento direto, exceto alguns policládidos que produzem uma larva
chamada larva de Goette com quatro braços, ou larva de Mueller, com oito.
A maioria dos trematódeos é hermafrodita com fertilização cruzada. Embora
variações no ciclo de vida ocorram entre espécies, o ciclo de vida generalizado dos
trematódeos (Fig. 8) consiste nos seguintes estágios: zigoto ~ larva miracídio ~
esporocisto ~ rédia ~ cercária ~ metacercária ~ adulto. O ciclo de vida inclui dois ou
mais hospedeiros e pelo menos dois estágios infectantes. O primeiro hospedeiro
intermediário (Fig. 8) é geralmente um molusco gastrópode (caramujo), se um
segundo hospedeiro intermediário ocorre, este é quase sempre um artrópode ou um
peixe; e um vertebrado é o hospedeiro definitivo ou final (Fig. 8).
Figura 8: Cnidaria, Scyphozoa: ciclo de vida (Figura 10.34, RFB pág. 289).
Os Monogenea são hermafroditas com fertilização cruzada ou autofecundação.
Os Cestoda apresentam um sistema reprodutivo completo dentro de cada
proglótide. Fertilização cruzada e autofertilização podem ocorrer entre duas
proglótides em um verme ou mesmo em uma proglótide. Em muitos casos, a
proglótide terminal madura cheia de “ovos” quebra-se do estróbilo. Os “ovos” são
liberados das proglótides pelas fezes do hospedeiro, ou no intestino do próximo
hospedeiro após ingerir uma proglótide. Tênias são endoparasitos do trato digestivo
de vertebrados. Seus ciclos de vida incluem geralmente dois ou mais hospedeiros
(artrópodos e vertebrados). O hospedeiro definitivo é sempre um vertebrado. O ciclo
de vida básico inclui zigoto ~ larva oncosfera (no hospedeiro intermediário ou
definitivo) ~ metacestóide (jovem extra-intestinal) ~ adulto intestinal (no hospedeiro
definitivo) (Fig 9).
Módulo V – Processos Reprodutivos
165
Figura 9: Cnidaria, Hydrozoa: anatomia dos hidróides e suas medusas (Figura 7.62, RFB pág. 189).
8. Nemertea: oss nemertinos reproduzem-se
reproduzem
assexuadamente por fragmentação
e regeneração. A maioria dos nemertinos é gonocórica. Os gametas desenvolvem-se
desenvolvem
a
partir de células-tronco
tronco que se agregam e tornam
tornam-se envolvidas por um epitélio,
formando uma gônada. Após a maturação dos gametas, um gonoduto curto cresce a
partir de cada uma das gônadas para o exterior,
exterio permitindo a saída dos gametas. A
fertilização é externa na maioria das espécies, e os ovos são liberados na água do mar
ou são depositados em depressões, em tubos ou cordões gelatinosos. Algumas
espécies são vivíparas. Os nemertinos têm desenvolvimento direto ou apresentam
larva lecitotrófica. No entanto, a maioria dos heteronemertinos apresenta larva
planctotrófica denominada pilídio.
9. Kamptozoa: oss camptozoários reproduzem-se
reproduzem
clonalmente por brotamento
e, desta maneira, promovem o crescimento das colônias (crescimento modular). As
espécies solitárias também se reproduzem por brotamento. A maior parte das
espécies de camptozoários é hermafrodita, mas alguns são gonocóricos. A fertilização
provavelmente é interna. A maioria das espécies é ovípara e apresenta
a
larvas
planctônicas. No entanto, algumas espécies retêm e incubam seus embriões no átrio.
O zigoto desenvolve-se
se numa larva ciliada livre
livre-natante chamada trocófora. Depois
de uma curta existência livre-natante,
natante, a larva se fixa e sofre metamorfose, gerando o
adulto. Em algumas espécies, a larva não se desenvolve diretamente em adulto, mas
produz brotos dos quais se originam os adultos.
10. Cycliophora: conforme mostra a Figura 10, os ciclióforos apresentam um
ciclo de vida muito complexo, que inclui fase sexual e assexual,
assexual ligados pelo
indivíduo-alimentar assexual séssil. O ciclo assexual inclui uma larva Pandora,
gerada por brotamento do indivíduo
indivíduo-alimentar assexuado. A larva Pandora gera
Módulo V – Processos Reprodutivos
Em 1995, um novo filo
veio somar-se aos 35 já
conhecidos no século XX.
Trata-se do Cycliophora,
nome dado em referência
à
coroa
de
cílios
166
outro indivíduo-alimentar. No indivíduo-alimentar, grupamentos de células-tronco
indiferenciadas podem desenvolver-se em três tipos de estágios livre-natantes: larva
Pandora (ciclo assexual), a larva Prometeu e a fêmea madura (ciclo sexual). A larva
Prometeu dá origem a um macho maduro. O macho e a fêmea maduros
desenvolvem-se fixados ao indivíduo-alimentar. Quando a fêmea está sexualmente
madura contém um único ovócito, se desprende do indivíduo-alimentar e, logo em
seguida, é fecundada internamente pelo macho maduro que estava à sua espera. O
zigoto formando dará origem a uma nova larva, chamada cordóide, que gerará um
novo indivíduo-alimentar. O novo indivíduo-alimentar gerado dará origem a novas
larvas Pandora e a novas larvas Prometeu (que formarão novos machos) e a novas
fêmeas.
Figura 10: Cnidaria, Hydrozoa: anatomia dos hidróides e suas medusas (Figura 2, Kristensen, 2002
***).
Módulo V – Processos Reprodutivos
167
11. Mollusca: o molusco generalizado é gonocórico, com fecundação externa na
água do mar. Gametas são liberados pelas gônadas para dentro do celoma. Eles
entram para os nefróstomas dos nefrídios, os quais servem como gonodutos, e saem
via nefridióporo, para dentro da câm
câmara exalante da cavidade do manto. Em alguns
moluscos, há gonodutos separados, não dependentes dos nefrídios. A fecundação é
interna em
m muitos moluscos, e alguns destes
dest
são hermafroditas. Ambos,
hermafroditismo e fecundação interna, requerem elaboração do sistema reprodutor.
O desenvolvimento do zigoto origina uma larva trocófora, que nos moluscos
primitivos é o estágio de eclosão. Em muitas espécies, entretanto, a trocófora é
suprimida. Alguns moluscos têm desenvolvimento direto. Trocóforas são geralmente
planctônicas e planctotróficas. A trocófora é sucedida no desenvolvimento de muitos
moluscos por um estágio mais avançado, a larva véliger, presente nos gastrópodos,
bivalves e escafópodos. O estágio de eclosão pode ser o de trocófora ou o de véliger.
Nestee caso, a fase de trocófora ocorre no ovo. Ao final da vida larval, a larva véliger
assenta sobre o substrato e metamorfoseia
metamorfoseia-se em um adulto bentônico. Alguns
moluscos, tais como os cefalópodes e muitos gastrópodes,
gastrópodes têm desenvolvimento
direto.
12. Sipuncula: a maioria das espécies de sipúnculos é gonocórica. A maturação
dos gametas ocorre no celoma do tronco. Quando maduros, os gametas são liberados
através dos funis nefridiais. A fertilização é externa na água do mar. O
desenvolvimento pode ser direto ou pode ser indireto, com uma larva trocófora ou
uma larva pelagosfera,, que se desenvolve a partir da trocófora.
13. Annelida
• Polychaeta: a reprodução clonal ocorre em alguns poliquetos por
brotamento, fissão, fragmentação e regeneração. Mas a maioria dos poliquetos
reproduz-se sexuadamente. A maioria das espécies é gonocórica. A fertilização e o
desenvolvimento do zigoto geralmente ocorrem externamente na água do mar.
Entretanto, algumas espécies
écies retêm e incubam seus ovos. O desenvolvimento é
indireto. O zigoto desenvolve-se
se em uma larva trocófora. As gônadas são
segmentares e são grupamentos de células germinativas presentes no revestimento
celômico. Os gametas são liberados e armazenados nas
na cavidades celômicas. Em
indivíduos sexualmente maduros, as cavidades celômicas encontram-se
encontram
completamente preenchidas por gametas. Os gametas são liberados por
metanefrídios ou por ruptura da parede do corpo. A ruptura da parede do corpo é
comum nos poliquetos
quetos que sofrem epitoquia. A epitoquia é a formação de um
indivíduo reprodutor pelágico (epítoco) a partir de um indivíduo não reprodutor
bentônico (átoco). A epitoquia coincide com a produção de gametas e maturidade
sexual. Um ou mais epítocos podem se originar a partir da forma átoca, seja por
metamorfose de todo o animal, seja por diferenciação e separação (brotamento) do
epítoco a partir da metade posterior do corpo do átoco (Figs. 11 e 12). A epitoquia é
uma estratégia de poliquetos para sincronizar a maturação sexual e garantir a
fertilização dos óvulos. Esta sincronização é chamada enxameamento. Durante o
enxameamento, vários epítocos nadam em busca de parceiros para reprodução.
Módulo V – Processos Reprodutivos
Polychaetas
Poliquetos
são
animais
marinhos
com muitas cerdas,
inseridas
em
projeções laterais ao
corpo
chamadas
parapódios.
O
primeiro anel do
corpo destes animais
forma a cabeça, que é
destacada do resto
do corpo do animal.
Raramente
são
hermafroditas;
a
maioria possui sexos
separados.
Exemplos: nereida,
sérpula.
168
Figura 11: Cnidaria, Hydrozoa: ciclos de vida. (A) Obelia (B) Tubularia (Figura 7.64, RFB pág. 190).
Módulo V – Processos Reprodutivos
169
Figura 12: Figura 10.34 - Digenea: ciclo de vida. O trematódeo chinês do fígado Opisthorchis sinensis.
(A) Vista dorsal do verme adulto. (B) Ciclo de vida (Figura 13.30, RFB 13.30 pág. 510).
• Clitellata: oss clitelados (Oligochaeta + Hirudínea) possuem uma estrutura
chamada clitelo.. O clitelo é um conjunto
conjun de segmentos encerrados por uma espessa
epiderme glandular, que engloba os gonóporos femininos ou se localiza
posteriormente a eles. O clitelo secreta muco para a cópula, albumina para a nutrição
dos ovos e um casulo no qual os ovos e a albumina são depositados.
dep
Os clitelados são
hermafroditas e realizam cópula. As gônadas de clitelados estão sempre restritas a
alguns poucos segmentos genitais e os testículos são anteriores aos ovários. Os
zigotos se desenvolvem no casulo e eclodem como juvenis. O desenvolvimento
desenv
é
direto.
Muitos oligoquetos podem reproduzir-se
reproduzir
assexuadamente por fissão ou
fragmentação, sendo que indivíduos sexuados são raros ou nunca foram observados
em algumas espécies que se reproduzem assexuadamente. Por haver alternância de
gerações assexuais e sexuais em algumas espécies de oligoquetos. Os hirudíneos
sanguessugas por sua vez não se reproduzem assexuadamente.
Os oligoquetos são hermafroditas e durante a cópula ocorre transferência
mútua de espermatozóides. Os vermes mantêm contato ventral
v
entre as extremidades
anteriores que se encontram voltados para direções opostas (Fig. 13A). Na maioria
dos oligoquetos, os gonóporos masculinos de um animal se justapõem diretamente às
aberturas dos receptáculos seminais do outro (Fig. 13 B,C). As cerdas genitais que
prendem um verme ao outro e um invólucro mucoso secretado pelos clitelos mantêm
os dois vermes na posição correta (Fig. 13B)
13B). Os espermatozóides são liberados dos
gonóporos masculinos de cada animal; atingem o interior do outro animal e
penetram seus receptáculos seminais. Depois da cópula, cada verme secreta um tubo
mucoso e uma camada quitinosos formando um casulo ao redor do clitelo (Fig. 13D).
À medida que o casulo desliza para frente, recebe os óvulos do oviduto, albumina de
células epidérmicas e os espermatozóides do parceiro armazenados nos receptáculos
seminais. A fertilização ocorre dentro do casulo. Quando o casulo se desprende do
animal, e se fecha nas extremidades, tomando um formato mais ou menos esférico
Módulo V – Processos Reprodutivos
Charles Darwin, em
1881, estudou as
minhocas
e
as
relacionou com os
processos
de
formação
das
características gerais
do
solo,
sendo
considerado por ele
um dos grupos mais
importantes para o
estabelecimento de
outras espécies no
ambiente terrestre.
Consultado
em
http://web2.sbg.org.
br/congress/sbg200
8/pdfs2009/GA37729569.pdf
170
(Fig. 13 D-F). Os oligoquetos possuem desenvolvimento direto. Depois de algum
tempo após a cópula, os jovens oligoquetos eclodem do casulo.
Figura 13: Cestoda: Anatomia e ciclo de vida da tênia de carne bovina, Taenia saginata (Figura 13.68,
RFB 13.68 pág. 542).
Módulo V – Processos Reprodutivos
171
Os hirudíneos são hermafroditas protândricos de fertilização interna. A cópula
nos hirudinídeos é semelhante àquela dos oligoquetos, porém a transferência de
espermatozóides pode ocorrer através da penetração de um pênis no gonóporo
feminino do parceiro
ceiro ou por impregnação hipodérmica (nas espécies sem pênis).
Durante a impregnação hipodérmica um espermatóforo é expelido de um animal e
penetra no tegumento do outro. O espermatóforo,
espermatóforo então, libera os espermatozóides
que migram para os ovissacos. Após a cópula ocorre a formação do casulo como nos
oligoquetos. O desenvolvimento é direto.
14. Echiura: oss equiúros são gonocóricos. A fertilização é externa. O
desenvolvimento é indireto com uma larva trocófora planctotrófica. A reprodução de
Bonellia difere daquela de todos os outros equiúros (Para
(
relembrar, leia o item
Mecanismos de Determinação do Sexo tratado a partir da página 11).
nicóforos são gonocóricos com fertilização interna e
15. Onycophora: onicóforos
transferência indireta de espermatozóides por meio de espermatóforos. Alguns
onicóforos são ovíparos, mas a maioria é vivípara (matrotróficos ou lecitotróficos).
lecitot
Nas espécies vivíparas, os embriões desenvolvem-se
desenvolvem
no útero. Nas espécies vivíparas
matrotróficas, os embriões são nutridos por secreções do epitélio uterino. Pode haver
um tecido placentário para o transporte destas
des
secreções. Na ausência de placenta, as
secreções são liberadas no lúmen do útero e absorvidas pelo embrião.
16. Tardigrada: a maioria das espécies de tardígrados é dióica, mas algumas
são hermafroditas. Muitas são partenogenéticas e os machos são desconhecidos em
muitas espécies. A fertilização pode ser externa ou interna. A cópula e oviposição
ocorrem em sincronia com a ecdise da fêmea (Fig. 14). Em algumas espécies, o macho
deposita os espermatozóides na exúvia recém-liberada
recém
da fêmea, que contém os
óvulos (Fig. 14B). Nestee caso, a fertilização é externa. Em outras espécies, o macho
copula e fertiliza os óvulos da fêmea antes que
q a ecdise desta seja completada (Fig.
14A). Nestee caso, a fertilização é interna. Alguns tardígrados podem produzir ovos
resistentes
ntes quando as condições são adversas. O desenvolvimento é direto.
Módulo V – Processos Reprodutivos
Hermafrodita protândrico
- estado de um ser
hermafrodita
que
se
desenvolve
primeiro
como masculino e depois
como feminino.
172
Figura 14: Cycliophora, ciclo de vida de Symbion Pandora mostrando que o macho primário
desenvolve dois machos anões secundários dentro do broto. É o macho secundário (17) quem fertiliza a
fêmea (18). A parte sexual do ciclo de vida inclui o indivíduo-alimentar séssil, que dá origem à larva
Pandora (3–11). A parte sexual do ciclo de vida inclui dois tipos de indivíduos-alimentares, cada um forma
um macho primário e uma fêmea, o macho secundário e a larva cordóide (12-2) (Figura 15.8, RFB pág. 596).
Módulo V – Processos Reprodutivos
173
17. Arthropoda: a maioria dos artrópodes é dióica, mas alguns crustáceos são
hermafroditas e alguns hexápodos são partenogenéticos. A fertilização pode ser
externa (nos xifosuros, alguns crustáceos, picnogonídeos) ou interna (aracnídeos,
hexápodos, crustáceos).. A transferência de espermatozóides pode ser direta,
direta
geralmente por meio de um pênis
pênis, ou indireta, por meio de espermatóforos
(hexápodos e aracnídeos, miriápodos) ou apêndices especializados denominados
gonópodes (crustáceos e miriápodos). Entretanto,
Entretanto as aranhas apresentam
transferência indireta sem espermatóforos através de um apêndice modificado dos
machos. Geralmente há cópula. As fêmeas guardam espermatozóides em um
receptáculo seminal e, depois, usam-nos
usam
para fertilizar os ovos. Muitos aracnídeos e
miriápodos
riápodos exibem um elaborado comportamento de ritual de corte que envolvem
sinais visuais, táteis e químicos. A maioria dos artrópodos é ovípara, mas a maioria
dos crustáceos incuba seus ovos, e alguns grupos como os escorpiões e alguns ácaros
e alguns insetos
etos são vivíparos. As gônadas dos artrópodes se desenvolvem em
espaços celômicos embrionários (gonocele) e conectam-se
conectam
com o exterior por
gonodutos pares derivados de celomodutos. As gônadas são geralmente tubulares e
dorsais. O duto deferente do macho po
pode apresentar uma produtora de
espermatóforos. O oviduto da fêmea geralmente apresenta um receptáculo seminal
especializado no armazenamento de espermatozóides. O desenvolvimento pode ser
direto, como na maioria dos artrópodos,
artrópodos ou indireto (como nos xifosuros,
picnogônidos, crustáceos, paurópodos, sínfilos, diplópodos e um grande número de
quilópodos). Os crustáceos apresentam vários estágios larvais e pós-larvais,
pós
que
recebem nomes diferentes, dependendo do grupo taxonômico, mas
ma a larva básica dos
crustáceos é o náuplio. São exemplos de estágios larvais e pós-larvais
pós
dos crustáceos:
o metanáuplio, a cípris, o copepodito, a protozoe, a zoe, a mísis e a megalopa. Por sua
vez, os xifosuros possuem uma larva trilobita e os picnogônidos
picnogôni
apresentam um
estágio larval denominado protoninfa
protoninfa.
18. Chaetognatha: oss quetognatos são hermafroditas. A fertilização é interna. A
transferência de espermatozóides é indireta,
indireta por meio de espermatóforos, e envolve
cópula. O desenvolvimento é direto.
19. Gnathifera: os gnatostomúlidos são hermafroditas. A cópula ainda não foi
observada, mas a transferência de espermatozóides uniflagelados ou aflagelados
provavelmente ocorra por impregnação hipodérmica da parede co
corpórea, por fixação
do gonóporo masculino ao tegumento do parceiro ou por injeção para dentro da
vagina. Um único ovo é posto em cada oviposição. O ovo rompe a parede corpórea e
adere ao substrato. O desenvolvimento é direto.
Pouco se sabe sobre a reproduç
reprodução dos Micrognathozoa. Nenhum macho foi
descoberto e Limnognathia provavelmente se reproduza por partenogênese. As
fêmeas possuem ovários pares. Aparentemente não há oviduto ou gonóporo.
gonópo
O
desenvolvimento é direto.
Os rotíferos são gonocóricos ou partenogenéticos. Os machos, quando
presentes, são anões e degenerados,
degenerados basicamente contendo apenas o aparato
reprodutor. Os bdelóideos não possuem machos e a reprodução é exclusivamente
partenogenética. Nos monogonontes, os machos têm existência breve em períodos
restritos e a maior parte do ciclo reprodutivo é partenogenético. A maioria dos
rotíferos é ovípara, mas algumas poucas espécies são vivíparas.
A cópula geralmente ocorre por impregnação hipodérmica, ou seja, o macho
espeta qualquer
alquer parte do corpo da fêmea com seu pênis. Os espermatozóides, que
possuem flagelo anterior, se dirigem até o ovário para fertilizar um ovócito. Os
rotíferos bdelóideos reproduzem-se
se partenogeneticamente. As fêmeas produzem
Módulo V – Processos Reprodutivos
O filo dos artrópodes
arthros
=
(grego:
articulado + poda = pé)
contém a maioria dos
animais conhecidos (mais
de 3 em cada 4 espécies
animais), mais de 1
milhão
de
espécies,
muitas
das
quais
extremamente abundantes
em
número
de
indivíduos.
Limnognathia maerski
é uma espécie de
animal microscópico,
único representante
do
filo
Micrognathozoa. É
um
animal
extremófilo,
tendo
sido descoberto em
nascentes de água
fria na Groelândia,
no ano de 1994,
descrita em 2000 .
Possui
mandíbula
complexa
se
alimentando
de
bactérias,
algas
verdes, diatomáceas
e
algas-azuis
e
reproduz-se
por
meio
de
partenogênese. Seu
comprimento pode
chegar
a
150
micrometros.
Consultado
em:
http://www.melhor
dawiki.com.br/wiki
/Micrognathozoa
Rotíferos são animais
aquáticos microscópicos
que constituem o filo
Rotifera. O seu nome
deriva do latim para
“roda”, com referência à
coroa de cílios que rodeia
a boca destes animais e
que
se
movem
rapidamente, para captar
as partículas de alimento,
parecendo uma roda a
girar.
174
ovos diplóides por mitose e oss ovos eclodem como mais fêmeas diplóides.
Nos rotíferos monogonontes o ciclo de vida é mais complexo, sendo que a
reprodução pode ser gonocórica ou partenogenética.
No ciclo partenogenético ou amíctico uma fêmea amíctica diplóide (Fig. 15)
produz um ovo amíctico de casca fina. Este
Es ovo é diplóide, não pode ser fertilizado e
se desenvolve numa fêmea amíctica geneticamente idêntica à mãe. Os ovos amícticos
desenvolvem-se e eclodem dando origem a novas fêmeas diplóides. Um segundo
tipo de ovo haplóide de casca
sca fina, chamado de ovo míctico, é produto de meiose.
Quando fertilizados, eles se desenvolvem em machos haplóides. Os ovos mícticos
fertilizados secretam cascas pesadas e resistentes. Estes ovos fertilizados são
chamados de ovos dormentes (ovos em repouso
repous ou “de inverno”). Os ovos
dormentes são diplóides e eventualmente irão eclodir como fêmeas diplóides. Os
ovos dormentes contêm um embrião em diapausa, capaz de resistir à dessecação,
temperaturas extremas e outras condições adversas,
adversas e pode não eclodir por vários
meses, ou mesmo anos. Ovos dormentes eclodem quando as condições ambientais
são favoráveis.
Figura 15: Epitoquia e poliquetos epítocos. (A) Palola viridis, com região posterior epítoca. (B) Parte
posterior de Trypanosyllis, que mostra um agregado de epítocos brotando. (C) Brotamento de epítocos em
Myrianida pachycera. (D) Poliquetos silídeos durante o enxameamento. O macho nada ao redor de fêmea
liberando espermatozóides (Figura
Figura 23.21, RFB pág. 932).
932)
Módulo V – Processos Reprodutivos
175
Os acantocéfalos são gonocóricos. A fertilização é interna, com cópula. O
sistema masculino inclui dois testículos, uma bursa copulatória, pênis protraível e
glândulas acessórias. Os espermatozóides possuem um flagelo anterior. O
desenvolvimento do zigoto se dá na hemocele e resulta na formação da larva acântor,
que é encapsulada (“ovo”). Os “ovos” passam da hemocele para o funil uterino, a
qual se abre no útero muscular. Um esfíncter muscular do útero controla a postura
dos “ovos”. Os “ovos” são liberados do gonóporo para o intestino do hospedeiro
definitivo (geralmente um vertebrado) e são eliminados nas suas fezes. Se a cápsula
for engolida por um hospedeiro intermediário (por exemplo, um inseto ou crustáceo),
a larva acântor emergirá da cápsula, perfurará a parede do intestino e se alojará na
hemocele do hospedeiro intermediário. No hospedeiro intermediário, a acântor
passará por mais dois estágios larvais chamados acantela e cistacanto (que também
são encistados). Quando o hospedeiro intermediário for ingerido por um vertebrado,
o cistacanto sai do cisto e a larva jovem se fixa à parede do intestino do vertebrado
completando o ciclo (Fig. 16).
Figura 16: Macho epítoco de Nereis irrorata. Os segmentos
reprodutores posteriores portam os espermatozóides (Figura 23.25, RFB pág.
938).
20. Cycloneuralia: gastrótricos são hermafroditas, mas algumas espécies
apresentam sistema reprodutor masculino rudimentar e se reproduzem
partenogeneticamente. Os gastrótricos possuem ovos de casca fina (ovos normais) e
ovos de casca grossa (ovos de resistência). A transferência de esperma é indireta por
espermatóforos e envolve cópula. O desenvolvimento é direto.
A maioria dos nematódeos é gonocórica, mas algumas espécies são
hermafroditas ou partenogenéticas. A fertilização é interna, com cópula. Muitas
espécies apresentam dimorfismo sexual. Durante a cópula, o macho se enrola e se
prende à fêmea com auxílio das espículas copuladoras, que são inseridas no
gonóporo feminino aberto durante a transmissão dos espermatozóides para a vagina.
Os espermatozóides são aflagelados e se movem por movimento amebóide. Os
zigotos são envolvidos por uma casca secretada em parte pelo próprio zigoto, e em
parte pelo epitélio do útero. A maioria das espécies é ovípara e os ovos dos
nematódeos podem ficar armazenados no útero ate serem depositados e,
frequentemente, o desenvolvimento embrionário começa enquanto os ovos ainda
estão na fêmea. Muitas espécies são vivíparas. O ciclo de vida inclui o ovo, três
estágios juvenis e o adulto. O desenvolvimento é direto.
Módulo V – Processos Reprodutivos
176
Nematomorfos são gonocóricos. Durante a cópula, o macho enrola firmemente
sua extremidade posterior ao redor da fêmea e transfere um espermatóforo ou
espermatozóides na cloaca da fêmea. Os espermatozóides, que não têm flagelo,
flagelo
migram até o receptáculo seminal, onde são
sã armazenados. A fertilização ocorre no
útero. As fêmeas depositam um fio de ovos na água ou entre as raízes das plantas nas
margens. O ciclo de vida inclui ovo, uma ou duas fases larvais parasitas e o adulto de
vida livre. Os hospedeiros podem ser insetos,
insetos miriápodos, aracnídeos, gastrópodos e
sanguessugas. O ciclo de vida pode conter um ou mais hospedeiros.
Priapúlideos são gonocóricos e a fertilização é externa ou interna, dependendo
da espécie. A maioria das espéciess tem desenvolvimento indireto.
Loricíferos são gonocóricos. Órgãos reprodutores e excretores são combinados
num sistema urogenital. Gametas e urina são liberados por um duto urogenital
comum, o qual se abre com ou ao lado do ânus. Suspeita-se
Suspeita
que haja fertilização
interna. O desenvolvimento é indireto,
indireto com fase larval chamada larva de Higgins.
Pouco se sabe sobre a biologia reprodutiva dos quinorrincos. Eles são
gonocóricos. A fertilização é provavelmente interna por meio de espermatóforos. O
desenvolvimento é direto.
oronídeos reproduzem-se assexuadamente por brotamento
21. Lophophorata: foronídeos
e fissão promovendo o crescimento m
modular de suas colônias. Além disto, foronídeos
têm grande capacidade de regeneração. A maior parte dos foronídeos é hermafrodita.
Os espermatozóides armazenados em espermatóforos são liberados na água. Os
espermatóforos são capturados pelo lofóforo dos outros
ou
indivíduos e, em seguida, os
espermatozóides tornam-se
se amebóides e atravessam a parede corporal para adentrar
a metacele. A fertilização é interna. Os zigotos são liberados pelas aberturas dos
metanefrídios e podem ser incubados no lofóforo ou liberados
liberad na água. Quase todos
os foronídeos apresentam desenvolvimento indireto e possuem uma larva chamada
actinotroca.. Após uma existência planctotrófica e livre-natante,
livre
a actinotroca sofre
uma metamorfose para gerar o adulto bentônico.
Quase todos os braquiópodos
ópodos são gonocóricos e apresentam fertilização
externa na água do mar. Algumas espécies são incubadoras ou vivíparas. O
desenvolvimento pode ser direto ou indireto.
Os nematódeos ou
nemátodos
(Nemathelminthes) -também chamados de
vermes cilíndricos-- são
considerados o grupo
de metazoários mais
abundante na biosfera,
com
estimativa de
constituírem até 80%
de
todos
os
metazoários, com mais
de 20.000 espécies já
descritas,
de
um
número estimado em
mais de 1 milhão de
espécies atuais, que
incluem muitas formas
parasitas de plantas e
animais. Apenas os
Arthropoda
apresentam
maior
diversidade. O nome
vem da palavra grega
nema, que significa fio.
Consultado
em
http://www.tiosam.co
m/?q=Nematelmintos
Foronídeos:
Os
foronídeos
são
animais
marinhos,
de
corpo
vermiforme,
cilíndrico,
não
segmentado;
cada
indivíduo vive em
tubo
membranoso
secretado por ele
mesmo.
A
extremidade anterior
contém
tentáculos
ciliados
e
um
lofóforo (conjunto de
tentáculos) em forma
de
ferradura.
Consultado
em
http://www.biomania.c
om.br/bio/conteudo.asp
?cod=1765
A maioria dos briozoários é hermafrodita. Briozoários possuem um ou dois
ovários e um até vários testículos. Os ovários localizam-se
localizam
na extremidade distal do
zoóide e os testículos sobre o funículo, na extremidade basal. Os gonodutos estão
ausentes, sendo os gametas liberados por ruptura das gônadas. A maior parte dos
Bryozoa é de hermafroditas protândricos, mas alguns poucos são hermafroditas
simultâneos. Assim, uma colônia pode ser composta de zoóides funcionalmente
masculinos e femininos em diferentes estágios de protandria. A ffertilização pode
ocorrer entre zoóides de uma mesma colônia ou entre colônias diferentes e pode ser
interna ou externa. A maioria dos briozoários incuba seus ovos, algumas poucas
espécies são vivíparas. Alguns briozoários exibem poliembrionia, ou seja, o embrião
original produz, assexuadamente, numerosos embriões secundários que, por sua vez,
podem produzir embriões terciários, e assim por diante. O desenvolvimento é
indireto, e após uma breve existência planctônica, a larva se fixa e sofre a
metamorfose para
ra dar origem ao primeiro zoóide (ancéstrula),
(
que dará origem a
uma nova colônia. Embora uma colônia contenha milhares de zoóides, todos são
descendentes assexuais da ancéstrula. Por meio de brotamento, a ancéstrula dá
origem a uma série de novos zoóides que, mesmo sendo geneticamente idênticos,
podem ser polimórficos, exibindo diferentes formas e tamanhos. A reprodução
Conheça ...
assexual contínua aumenta o tamanho da colônia. Espécies de água doce também se
reproduzem por meio de gemulação, produzindo estatoblastos.
estatoblastos Estatoblastos são
Módulo V – Processos Reprodutivos
gêmulas resistentes às condições ambientais desfavoráveis, como seca e frio, e podem
permanecer dormentes enquanto durarem estes períodos. Quando as condições
ambientais tornam-se
se favoráveis, um zoóide se desenvolve
desen
a partir do estatoblasto.
22. Chordata: os Craniata apresentam gônadas pares que se desenvolvem no
mesotélio e geralmente são retroperitoniais (exceto nalguns mamíferos cujos
testículos são localizados dentro da bolsa escrotal). Os ovários contêm células sexuais
primárias chamadas folículos, que nutrem o óvulo
ó
em desenvolvimento, estimulando
a formação de vitelo no ovócito. Quando os óvulos amadurecem, os folículos se
rompem liberando os óvulos completos (ovulação). A ovulação pode ser quase
contínua, sazonal ou única. Os testículos são compostos por túbulos
túbulo seminíferos
interconectados nos quais os espermatozóides se desenvolvem. Nestes
Nes
túbulos os
espermatozóides são sustentados e nutridos por células de suporte. Nos craniados
basais com os peixes-bruxa
bruxa e as lampréias não existem gonodutos e os gametas
movem-se
se através do celoma até gonóporos que se abrem para o exterior. Entretanto,
nos gnatostomados há gonodutos para transporte dos gametas. Nos machos, com
exceção dos peixes actinopterígios, os gonodutos são formados a partir do ducto do
metanefrídio (ducto arquinéfrico) que drenava originalmente o rim. Nas fêmeas, os
óvulos ainda são liberados para o celoma, mas recolhidos pelo oviduto. Os ovidutos
produzem, através de glândulas especializadas, envoltórios mucosos para os
o óvulos
dos anfíbios. Produzem ainda as cascas dos ovos de tubarões e répteis e mamíferos
monotremados, e albumina e vitelo nos amniotos. Os ovidutos podem expandir
expandir-se e
fusionar para formar um útero ou úteros pares para armazenamento dos ovos ou do
embrião. Os craniados com poucas exceções são gonocóricos, mas alguns peixesbruxa, peixes ósseos e alguns anfíbios são hermafroditas e alguns répteis são
partenogenéticos. A fertilização pode ser externa (maioria dos peixes e anfíbios), ou
interna (peixes cartilaginosos, alguns peixes ósseos, répteis
rép
e mamíferos). Alguns
craniados com fertilização interna desenvolveram órgãos copulatórios como o pênis
dos mamíferos, os hemipênis dos lagartos e cobras, os clásperes dos peixes
cartilaginosos e os gonopódios de alguns poucos grupos actinopterígios. Os
O urodelos
(salamandras) apresentam transferência indireta de espermatozóides por meio de
espermatóforos. Muitos craniados apresentam elaborados rituais de corte,
corte bem como
prolongados períodos de cuidado com a prole (o exemplo dos mamíferos humanos é
um caso
so extremo!). Os craniados podem ser ovíparos lecitotróficos, ovíparos
matrotróficos, ou vivíparos. O desenvolvimento geralmente é direto, exceto nas
lampreias, que passam por um estágio larval denominado amocetes e nos anfíbios
modernos, que apresentam um
uma fase larval conhecida como girino. O
desenvolvimento do embrião pode ocorrer na água, no ambiente terrestre ou dentro
do corpo da mãe.
Os cefalocordados são animais dióicos com fecundação externa e ciclo de vida
complexo que envolve um adulto bentônico e uma larva planctotrófica. As gônadas
são pares e numerosas. Os gametas maduros rompem as gônadas, caem no átrio e são
carregados ao exterior na corrente de água expelida pelo atrióporo.
Os ascidiáceos são hermafroditas e geralmente têm fertilização cruzada.
Ascídias solitárias são ovíparas. Os gametas são expelidos através do sifão exalante e
a fertilização ocorre na água do mar. A maioria das espécies coloniais é vivípara. O
embrião desenvolve-se como uma larva girinóide. A larva fixa-se e sofre
metamorfose para gerar o adulto. O crescimento modular das ascídias coloniais
ocorre por brotamento. As colônias de taliáceos crescem por adição de novos zoóides
que se originam por brotamento. A reprodução sexuada, com fecundação interna,
inicia-se
se quando o óvulo único, presente em cada zoóide, é fecundado.
fec
O
desenvolvimento pode ser direto ou indireto, ou seja, uma larva girinóide pode estar
ou não presente. O primeiro zoóide (oozoóide),
(
formado a partir da metamorfose da
larva girinóide,, fundará nova colônia. Alguns grupos apresentam ciclos de vida que
Módulo V – Processos Reprodutivos
estruturas
rochosas.
Existem a cerca de 520
milhões de anos. Os
indivíduos
possuem
tentáculos ciliados ao
177
redor da boca. Vivem em
água salgada ou doce.
Consultado
em
http://www.rc.unesp.br/
igce/aplicada/MUSEU/b
riozoarios.html
CRANIATA. Grupo que
inclui todos os animais
possuidores de crânio:
Esqueleto
da
cabeça
distinta, incorporando a
porção
anterior
da
notocorda, cérebro, órgãos
olfatórios, olhos e ouvido
interno. Consultado em
http://www.dse.ufpb.br/
lapec/arquivos/Craniata.
ppt
Os
cordados
(Chordata, do latim
chorda,
corda)
constituem um filo
dentro
do
reino
Animalia que inclui
os vertebrados, os
anfioxos
e
os
tunicados.
São
caracterizados pela
presença de uma
notocorda, um tubo
nervoso
dorsal,
fendas branquiais e
uma cauda pós-anal,
em pelo menos uma
fase de sua vida. O
grupo
abrange
animais adaptados
para a vida na água,
na terra e no ar.
Consultado
em
http://pt.wikipedia.
org/wiki/Cordados
Hemichordata (do grego
hemi, metade + chorda,
corda) é um filo de
animais
marinhos,
considerados como as
formas mais primitivas
dos cordados. É um filo
de animais vermiformes
que
mostram
semelhanças tanto com
os cordados como com
os
equinodermos,
e
geralmente
são
considerados como um
grupo muito próximo
dos equinodermos. O
grupo
surgiu
no
Câmbrico
e
conta
actualmente com cerca
178
alternam fases assexuadas e sexuadas, formas solitárias e coloniais. Os larváceos
apresentam reprodução sexuada e desenvolvimento direto. Todos os larváceos são
hermafroditas protândricos, exceto Oikopleura dioica, que é gonocórico.
23. Hemichordata: enteropneustos
nteropneustos podem se reproduzir assexuadamente por
fragmentação e regeneração. Os zoóides de pterobrânquios originam-se
originam
assexuadamente por brotamento durante o crescimento da colônia. Além disto,
dis um
disco adesivo de Cephalodiscus é capaz de fragmentação ou fissão para formar
colônias novas. Enteropneustos são gonocóricos. A fertilização é externa na água do
mar. O desenvolvimento é indireto e inclui fase larval lecitotrófica ou planctotrófica
chamada tornária.
Colônias de pterobrânquios
quios são hermafroditas, mas os zoóides que as
compõem geralmente são masculinos, femininos ou neutros. Os detalhes da
fertilização não são inteiramente conhecidos e geralmente ocorre incubação. Os
zigotos desenvolvem-se em larvas lecitotróficas. Após uma breve existência pelágica,
a larva se fixa e se encapsula num casulo antes de sofrer a metamorfose gradual,
gradual que
originará num único zoóide que, por sua vez, fundará uma nova colônia.
24. Echinodermata: crinóides,
rinóides, asteróides e ofiuróides reproduzem-se
reproduzem
clonalmente
nalmente por fissão e fragmentação seguida de regeneração. A maioria dos
equinodermados são dióicos com fertilização e desenvolvimento embrionário
externos na água do mar.
Algumas espécies são incubadoras e apresentam desenvolvimento direto, mas
a maioria dos Echinodermata apresenta estágio larval planctotrófico, que difere
ligeiramente entre si nas diferentes classes e,
e portanto, recebem denominações
distintas: bipinária e
braquiolária (Asteroidea), ofioplúteo (Ophiouroidea),
equinoplúteo (Echinoidea), auriculária
uriculária e doliolária (Holothroidea) e doliolária
(Crinoidea). Após uma existência livre-natante,
livre
a larva sofre metamorfose que resulta
na formação de uma miniatura do adulto.
O estágio larva de alguns Echinodermata pode passar por estágio de fixação ao
a
substrato anterior à metamorfose.
ATIVIDADE:
Metazoa assim como os
1. Descreva os padrões básicos de ciclos de vida dos Metazoa,
principais mecanismos reprodutivos dos grandes grupos de Metazoa.
Módulo V – Processos Reprodutivos
O filo Echinodermata
echinos,
(do
grego
espinho + derma, pele +
ata)
são
animais
marinhos, de vida livre
(estrela-do-mar,
holotúria e ouriço-domar). Os espinhos estão
presentes em diversos
formatos e atuam com a
função de proteger o
animal
e
para
a
locomoção. Podem ser
recobertos
por
substâncias de caráter
tóxico.
Muitos
têm
notável
poder
de
regeneração:
uma
estrela-do-mar cortada
radialmente em várias
partes vai, depois de
alguns meses, regenerar
em
tantas
estrelas
viáveis quantas foram as
partes
separadas.
Consultado
em
http://pt.wikipedia.org
/wiki/Echinodermata
179
Bibliografia
BARNES, R.S.K.; P. CALOW; P.J.W. OLIVE. Os invertebrados: uma nova síntese.
Atheneu, São Paulo. 526 pp. 1995.
BRUSCA, R.C., BRUSCA, J.G. Invertebrates. 2ª edition. Sunderland: Sinauer
Associates, 936 pp. 2003.
GRASSÉ, P.P. Traité de Zoologie. Paris, Masson et Cie Editeurs, 27 vol. 1948-1994.
HICKMAN, C.P.; L.S. ROBERTS; A. LARSON. Princípios Integrados de Zoologia.
11ª edição. Guanabara Koogan. Rio de Janeiro. 846 pp. 2004.
HILDEBRAND, M.; G. GOSLOW. Análise da Estrutura dos Vertebrados. 2ª edição.
Atheneu Editora. São Paulo. 641 pp. 2008.
HYMAN, L.H. The Invertebrates: Platyhelminthes and Rhynchocoely. Vol. 2
McGraw Hill. New York. pp. 52-219. 1951.
HYMAN, L.H. The Invertebrates: Protozoa Through Ctenophora. Vol. 1. McGraw
Hill New York. pp. 365-696. 1940.
KRISTENSEN, R.M. An Introduction to Loricifera, Cycliophora, and Micrognathozoa.
Integrative and Comparative Biology, v. 42, n. 3, p. 641–651 . 2002.
POUGH, F.H.; C.M. JANIS; J.B. HEISER. A Vida dos Vertebrados. 4ª. Edição.
Atheneu Editora. São Paulo. 750 pp. 2008.
ROMER, A. S. & T. S. PARSONS. Anatomia Comparada dos Vertebrados. Atheneu
Editora. São Paulo. 559 pp. 1985.
RUPPERT, E.E., R.S. FOX; R.D. BARNES. Zoologia dos Invertebrados. 7ª edição. São
Paulo. Editora Roca. 1145 pp. 2005.
RUSSEL-HUNTER, W.D. Uma Biologia dos Invertebrados Inferiores. Edusp e
Editora Polígono. São Paulo. 236 pp. 1969.
STORER, T.I; R.L. USINGER. Zoologia Geral. Companhia Editora Nacional e Edusp.
São Paulo. 757 pp. 1983.
* RUPPERT, E.E., R.S. FOX; R.D. BARNES. Zoologia dos Invertebrados. 7ª edição.
São Paulo. Editora Roca. 1145 pp. 2005.
** BARNES, R.S.K.; P. CALOW; P.J.W. OLIVE. Os invertebrados: uma nova síntese.
Atheneu, São Paulo. 526 pp. 1995.
*** KRISTENSEN, R.M. An Introduction to Loricifera, Cycliophora, and
Micrognathozoa. Integrative and Comparative Biology, v. 42, n. 3, p. 641–651 . 2002.
(em anexo).
Módulo V – Processos Reprodutivos

![aula1repro2[1]](http://s1.studylibpt.com/store/data/001690094_1-fa21ae7b4289ef06a76492482fda6a4b-300x300.png)


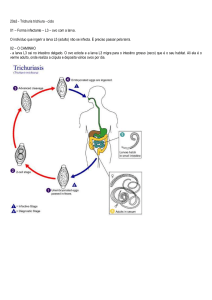
![Nematelmintos [Somente leitura]](http://s1.studylibpt.com/store/data/000747447_1-2661e0ded4244b6613a86a9aa535a4ae-300x300.png)




