MEDICINA – UFPE – 119
ALUNO: JOSÉ RAMOS DA COSTA JÚNIOR
PROFª: ELIZABETH CHAVES
OBJETIVOS – GLICÓLISE
1.
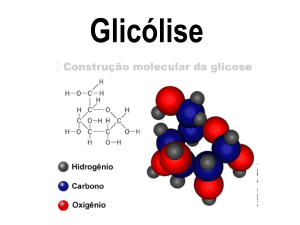
SABER QUE O TERMO GLICÓLISE REFERE-SE AO CATABOLISMO DA GLICOSE ATÉ A
FORMAÇÃO DO PIRUVATO.
“Na Glicólise, uma molécula de glicose é degradada em uma série de reações catalizadas por enzimas para liberar duas
moléculas de piruvato”-Lehninger. A rota glicolítica é empregada por todos os tecidos para a degradação da glicose para
fornecimento de energia e intermediários para outras rotas metabólicas.
2.
EXPLICAR PORQUE A FASE PREPARATÓRIA DA GLICÓLISE É CONSUMIDORA DE ENERGIA.
A fase preparatória da glicólise requer o investimento de duas moléculas de ATP e resulta na clivagem da cadeia da hexose
em duas trioses fosfato. O primeiro consumo de ATP ocorre na reação (irreversível) de transformação da Glicose em
Glicose-6-fosfato catalizada pela hexoquinase. Isso é um bom começo porque retém a glicose na forma de glicose-6-fosfato
no citosol da célula, onde todas as enzimas glicolíticas estão localizadas. As moléculas de açúcar fosforiladas não penetram
facilmente nas membranas célulares.A outra etapa da fase preparatória que gasta ATP é a reação (irreversível) de
transformação da frutose-6-fosfato em frutose-1,6-difosfato, catalizada pela enzima alostérica fosfofrutoquinase-1, sendo
esta etapa importantíssima na via glicolítica.
3.
ESCREVER A REAÇÃO COMPLETA DA TRANSFORMAÇÃO DE GLICOSE EM GLICOSE-6-FOSFATO
(GP6) E SABER COMO OCORRE A REGULAÇÃO DA ENTRADA DE GLICOSE NA CÉLULA.
No primeiro passo da via glicolítica, a glicose é ativada para as reações subsequentes pela sua fosforilação em C-6 para
liberar a glicose-6-fosfato. O doador de fosfato é o ATP.
Glicose + ATP glicose-6-fosfato + ADP
Para ser ativa a hexoquinase precisa de íons Mg 2+, pois o substrato da enzima é o complexo MgATP2-.
A hexoquinase é dependente da concentração de Gli6P, isso para qualquer célula. Então se a concentração de Gli6P estiver
elevada no interior da célula, pára a entrada de glicose e a hexoquinase é inibida. Entao isso é um processo que evita o
acúmulo de glicose dentro da célula se não houver necessidade de sua ultilização. Isoenzimas de hexoquinases são
fortemente inibidas pelo produto da reação. Isso é importante para que a hexoquinase não amarre todo o Pi de uma célula
na forma de hexoses fosforiladas.
hexoquinase
4.
IDENTIFICAR A REAÇÃO CATALIZADA PELA GLICOCINASE E EXPLICAR ONDE E QUANDO ESTA
ENZIMA ATUA.
A hexoquinase aparece em todos os nossos tecidos. Já a glicocinase é aquela que reage exclusivamente no fígado e
depende da concentração sanguínea de glicose. Se a concentração de glicose for muito alta, teremos a participação dessa
enzima ajudando o fígado a captar mais glicose e formar glicogênio, estocando essa glicose. Se a glicose voltar aos níveis
normais basais ela deixa de atuar. Entao ela é dependente dessa concentração no sangue, não sofrendo outro processo
regulatório. A glucocinase tem um alto Km, que contribui para a capacidade do fígado de “tamponar” os níveis de glicose
sanguínea. Qualquer aumento na concentração de glicose, leva a um aumento proporcional na velocidade de fosforilação
de glicose pela glicocinase. Da mesma forma, qualquer diminuição na concentração de glicose leva a uma diminuição
proporcional na velocidade de forforilação da glicose. Portanto, o fígado utiliza a glicose a uma velocidade significativa
apenas quanto os níveis de glicose sanguínea estão elevados. Por outro lado, uma isoenzima de hexoquinase e Km baixo
para a glicose, seria bom para tecidos como o cérebro, porque permite a fosforilação de glicose mesmo quando as
concentrações de glicose no sangue e no tecido são perigosamente baixas..
5.
RECONHECER QUE A TRANSFORMAÇÃO DE GP6 EM SEU ISÔMERO FRUTOSE-6-FOSFATO (F6P) É
UMA ETAPA PREPARATÓRIA PARA A QUEBRA DA HEXOSE EM DUAS TRIOSES.
Essa é uma etapa perfeitamente reversível, catalizada pela fosfoglicose isomerase (também necessita de íons Mg 2+). Essa
etapa não está sujeita a regulação, assim, ocorre tanto na glicólise como na gluconeogênese. Então, a G6P se transformando
em F6P, tem uma vantagem, pois é mais fácil quebrar a frutose em duas trioses, o que não ocorreria com a glicose.
6.
IDENTIFICAR A TRANSFORMAÇÃO DA F6P EM FRUTOSE-1,6-BIFOSFATO (F1,6bisP) COMO ETAPA
LIMITANTE DA VIA GLICOLÍTICA.
Nessa reação irreversível, nós temos a atuação da enzima fosfofrutoquinase-1, conhecida como “enzima marca-passo” da
via glicolítica. Ela representa o ponto principal de regulação da glicólise. Nos processos catabólicos, geralmente, a enzima
inicial é que sofre regulação. Só que no caso da hexoquinase e da glicocinase, essas enzimas não são exclusivas da via
glicolítica e apenas internalizam a glicose. E a glicose dentro da célula pode ser usada para outros fins além do processo
degradativo. Logo, não podemos trabalhar em cima do processo regulador da entrada da glicose. A hexoquinase regula a
entrada de glicose e a fosfofrutoquinase-1 regula a via glicolítica.
7.
IDENTIFICAR OS PRINCIPAIS MODULADORES POSITIVOS (ATIVADORES)
E NEGATIVOS
(INIBIDORES) DA FOSFOFRUTOQUINASE E EXPLICAR COMO ELES REGULAM A ENZIMA.
A fosfofrutoquinase é uma enzima alostérica, com os sítios de ligação para o inibidor, para o ativador, e para o prórprio
sítio ativo da enzima. Ela estará inibida toda vez que tivermos um excesso de energia na célula e estará ativada toda vez
que a energia da célula baixar.
Se tivermos muita molécula de ATP na célula, isso representa estoque de energia, isso quer dizer que eu não preciso
mais degradar glicose para transformar em energia. Logo, ATP é inibidor, e ADP e AMP são ativadores. Toda vez que
ADP e AMP estiver em alta, é sinal que o ATP baixou;
Citrato é inibidor: a glicose em excesso começa a degradar e formar Acetil-CoA e que em excesso chega a ácidos
graxos, pois a produção de acetil não acompanha a velocidade do ciclo de Krebs, pois a velocidade do ciclo é inferior à
velocidade de produção de acetilcoenzima-A. O figado pega essa acetilcoenzima, tira da mitocondria, passa para o
citoplasma e forma os acidos graxos. O acetil sai da mitocôndria na forma de citrato (acetil + oxalacetato). Ele
entrando no citoplasma vai indicar para que a rota da glicólise seja interrompida, que está ocorrendo
desnecessariamente, não esta acompanhando a velocidade do ciclo e precisa impedir que isso continue.
Faria sentido que o lactato, como produto final da glicólise, inibisse a enzima marca-passo. Não inibe. Entretanto, íons
hidrogênio, outro produto final da via glicolítica, inibem a fosfofrutoquinase-1. Na verdade, a glicólise gera ácido
láctico e a célula deve dispor dele como tal, isso explica porque o excesso de glicólise no corpo baixa o pH do sangue
e leva a uma situação de emergencia chamada acidose láctica. Há um simporte nas células que liberam esses produtos
para a corrente sanguínea. Íons hidrogênio são capazes de “desligar” a glicólise, o processo responsável pela queda do
pH.
O glucagon é liberado nas células alfa do pancreas e circula no sangue até entrar em contato com os receptores de
glucagon, localizados na superfície externa da membrana plasmática das células hepáticas. A ligacao de glucagon a
esses receptores influencia a atividade da adenilil ciclase, uma enzima localizada na superficie interna da membrana
plasmatica, estimulando-a a converter ATP em cAMP. O cAMP dispara uma série de reações em cascata que resultam,
finalmente, na diminuição dos níveis de frutose-2,6-difosfato. Uma diminuição desse composto torna a
fosfofrutoquinase-1 menos efetiva, mas torna a 1,6-difosfatase mais efetiva, restringindo severamente, desta forma, o
fluxo de frutose 6-fosfato para frutose-1,6-difosfato na glicólise.
8.
OBSERVAR QUE AS FOSFORILAÇÕES DA GLICOSE E DA F1,6bisP SÃO REAÇÕES IRREVERSÍVEIS
DA GLICÓLISE E REQUEREM A PRESENÇA DE Mg++.
Muitas quinases requerem Mg2+. As reações: Glicose G6P, F6PF1,6bisP são reações irreversíveis.
9.
IDENTIFICAR AS DUAS TRIOSES FOSFORILADAS OBTIDAS DA QUEBRA DA F1,6bisP E A REAÇÃO
DE CONVERSÃO DAS TRIOSES.
A enzima frutose-1,6-difosfato aldolase, cataliza a reação de quebra da frutose-1,6-difosfato liberando duas trioses
diferentes, o gliceraldeído-3-fosfato e a diidroxicetona fosfato. Apenas o gliceraldeído resultante dessa reação pode ser
diretamente degradado nos passos seguintes. Entretanto, o outro produto, a diidroxicentona fosfato, é rapida e
reversivelmente convertida em gliceraldeído-3-fosfato pela triose fosfato isomerase.
10. SABER QUE A REAÇÃO DE OXIDAÇÃO DO GLICERALDEÍDO-3-FOSFATO REQUER COMO
COENZIMA ACEPTORA DE HIDROGÊNIOS, O NAD. E COMO AGENTE DE FOSFORILAÇÃO O
PRÓPRIO FOSFATO ORGÂNICO.
Bem, embora a Gli3P-desidrogenase não tenha um processo regulador, é dela que depende a continuidade ou não do
processo glicolítico. Vamos usar NAD+ que vai receber H2 do gliceraldeído, e este vai passar para a forma de ácido, o
glicerato, e além disso vai entrar outro grupo fosfato na posição 1, dando 1,3-difosfoglicerato. Diferente de outras
fosforilações, nós não vamos ter ATP. É o fosfato orgânico que fosforila. E depois nós estamos reduzindo o NAD. Existe
NAD presente nas mitocondrias e no citosol. A membrana mitocondrial é impermeável tanto a entrada quanto a saida de
NAD. Quando nós formamos o NAD reduzido, nós temos que regenerá-lo posteriormente na forma oxidativa para que essa
reação não pare. Para transformarmos o piruvato em lactato é preciso de H2 que vem do NAD reduzido daquela reação (de
gliceraldeido). Entao eu regenero o NAD oxidado. A única vantagem de se formar o lactato não é nenhuma vantagem
energértica. É, na realidade, uma forma de recuperarmos o NAD que foi perdido. Piruvato + NAD + H + Lactato +
NAD+. No eritrócito, assim como no fígado, quando se forma lactato, ele é mandado através da corrente sanguínea para o
fígado, que forma de novo glicose através de uma gliconeogênese. Assim, pode se formar a glicose, a qual pode ser
devolvida para o músculo.
11. MOSTRAR QUE AS DUAS ETAPAS DE FORMAÇÃO DE ATP NA GLICÓLISE OCORREM QUANDO O
1,3-DIFOSFOGLICERATO SE TRANSFORMA EM 3-FOSFOGLICERATO E O FOSFOENOLPIRUVATO
FORMA O PIRUVATO.
A enzima fosfogliceratoquinase transfere o grupo fosfato de alta energia do grupo carboxila do 1,3-difosfafatoglicerato
para ADP, formando ATP e 3-fosfoglicerato.
Reação: gliceraldeído-3-fosfato + ADP + Pi + NAD+ 3-fosfoglicerato + ATP + NADH + H+
O último passo da glicólise é a transferência de um grupo fosfato do fosfoenolpiruvato para o ADP, catalisado pela
piruvato quinase. A reação da piruvato quinase requer K+ e Mg2+ ou Mn2+. Ela é um importante sítio de regulação.
12. OBSERVAR QUE TAMBÉM ESTAS REAÇÕES DE FOSFORILAÇÃO DO ATP REQUEREM Mg ++, DAÍ A
IMPORTÂNCIA DESTE ÍON NO METABOLISMO PRODUTOR DE ENERGIA.
OK.
13. DEMONSTRAR O DESTINO DO PIRUVATO EM CONDIÇÕES DE ANAEROBIOSE E AEROBIOSE.
Em condições aeróbicas o piruvato é oxidado a acetato, o qual entra no cíclo de Krebs e é oxidado até CO 2 e H2O.
Entretanto, sob condições anaeróbicas, o NADH gerado pela glicólise não pode ser reoxidado pelo O 2. A incapacidade de
regenerar o NADH em NAD+ deixaria a célula sem receptor de elétrons para a oxidação do gliceraldeído-3-fosfato e a
glicólise cessaria. A quantidade de NAD+ no citosol é pequena.
AEROBIOSE na mitocondria, através da piruvato desidrogenase, é transformado em ACETIL CoA que vai participar
do ciclo de Krebs ou então sai na forma de citrato, e vai formar ácidos graxos no citosol.
ANAEROBIOSE é reduzido a LACTATO, sendo importante para recuperar o NAD+ perdido na glicólise,
ETANOL+CO2 .
14. EXPLICAR PORQUE O PIRUVATO É CONVERTIDO A LACTATO NO MÚSCULO EM EXERCÍCIO E
NO ERITRÓCITO, E O DESTINO DO MESMO.
O músculo se contraindo rapidamente e intensamente, está formando piruvato, mas não tem onde colocar pois a
mitocondria não aceita mais. Porem tem que ser mantida a movimentacao, porque mesmo sendo de baixo rendimento
energetico, o processo anaerobico tem um saldo de 2 ATP. Porém, durante contração intensa, já está provado que a
glicólise ocorre de maneira muito mais rápida, entao você compensa a baixa produção pela velocidade de degradação.
Então o piruvato se reduz, produzindo lactato: piruvato + NAD+ + H+ lactato + NAD+. Após sua formação o lactato sai,
pois o musculo não tem condicoes de fazer reacao inversa. No eritrocito, assim como no musculo, quando se forma o
lactato, ele é mandado atraves da corrente sanguinea para o figado, que forma de novo glicose através de uma
gliconeogenese. Assim, pode-se formar glicose, que pode ser devolvida para o músculo. Esse é o ciclo de Cori. Ele é
realizado para tirar lactato, que em grandes concetrações, provoca dores e caimbras.