MEDICINA – UFPE – 119
ALUNO: JOSÉ RAMOS DA COSTA JÚNIOR
PROFª: ELIZABETH CHAVES
OBJETIVOS – CICLO DE KREBS OU CICLO DOS ÁCIDOS TRICARBOXÍLICOS
1.
OBSERVAR QUE O CATABOLISMO DE CARBOIDRATOS, LIPÍDIOS E PROTEÍNAS GERA UM
INTERMEDIÁRIO COMUM, acetil-CoA.
A maioria das principais vias metabólicas geradoras de energia das células resulta na produção da unidade de dois
carbonos acetil coenzima A (CoA). A quebra metabólica, pela via glicolítica, de carboidratos ingeridos ou armazenados,
de ácidos graxos de cadeia longa na seqüenciada -oxidação ou de certos aminoácidos após a transaminação ou
desaminação e subseqüente oxidação, fornecem precursores para a formação de acetil-CoA.
2.
VERIFICAR QUE ESTE INTERMEDIÁRIO DE DOIS CARBONOS SERÁ OXIDADO A CO 2, NUM
PROCESSO CÍCLICO DENOMINADO CICLO DE KREBS OU CICLO DOS ÁCIDOS
TRICARBOXÍLICOS.
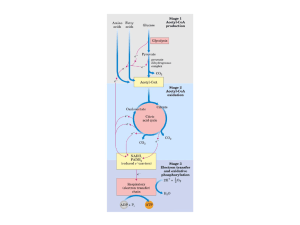
A função central do ciclo de Krebs é a oxidação total do acetil-CoA em CO2 e H2O. A principal função do Ciclo de
Krebs é gerar equivalentes de redução que são utilizados para gerar energia, isto é, ATP, na seqüência transporte de
elétrons-fosforilação oxidativa.
3.
ENTENDER QUE ESTE PROCESSO CÍCLICO ALÉM DE OXIDAR DOIS CARBONOS, DARÁ ORIGEM
A 4 PARES DE HIDROGÊNIO (EQUIVALENTES REDUTORES).
No ciclo de Krebs, dois átomos de carbono entram no ciclo como acetil-CoA e saem como CO2. Quatro pares de elétrons
são transferidos durante uma volta do ciclo: três pares de elétrons reduzindo NAD + a NADH + H+ nas reações 3, 4 e 8, e
um par reduzindo FAD a FADH2 na reação 6.
Reação geral:
Acetil-CoA + 3 NAD+ + FAD + GDP + Pi + 2 H2O 2 CO2 + 3 NADH + 3 H+ + FADH2 + GTP + CoA-SH
4.
SABER QUE OS ELÉTRONS DOS PARES DE HIDROGÊNIO SERÃO CONDUZIDOS ATÉ A CADEIA
TRANSPORTADORA DE ELÉTRONS.
Os quatro passos de oxidação do ciclo fornecem um grande fluxo de elétrons para a cadeia respiratória e esta leva à
formação de um grande número de moléculas de ATP durante a fosforilação oxidativa.
5.
ENTENDER QUE O TRANSPORTE DE ELÉTRONS PRODUZ ENERGIA SUFICIENTE PARA O
ANABOLISMO.
O rendimento líquido de ATP durante a completa oxidação da glicose a 6CO2 + 6 H2O é de 38 moléculas de ATP.
6.
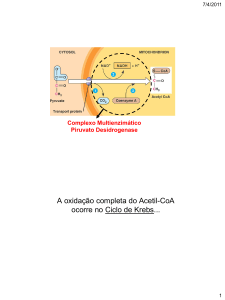
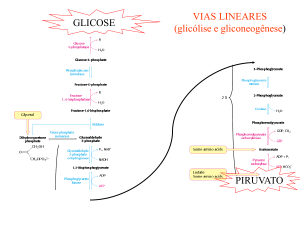
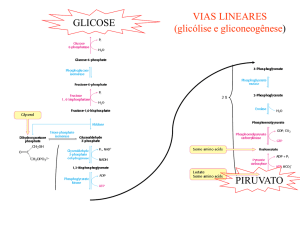
LEMBRAR QUE A GLICÓLISE PRODUZ PIRUVATO NO CITOSOL, O QUAL PENETRA NA
MITOCÔNDRIA EM CONDIÇÕES AERÓBICAS.
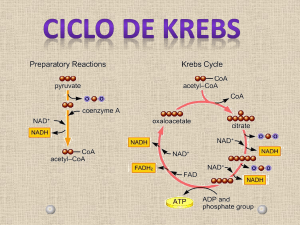
Formado durante a glicólise, o piruvato tem vários destinos metabólicos, dependendo do tecido e do estado metabólico
deste tecido. Pode ser descarboxilado oxidativamente em acetil-CoA pelo complexo multienzimático piruvato
desidrogenase localizado exclusivamente na matriz mitocondrial. Ou então:
* Através de uma reação de transaminação formar Alanina;
* Através de uma reação de carboxilação formar Oxaloacetato;
* Através de uma reação de redução formar o Lactato;
7.
SABER QUE A TRANSFORMAÇÃO DO PIRUVATO EM acetil-CoA REQUER UM COMPLEXO
ENZIMÁTICO FORMADO POR 3 ENZIMAS E 5 COENZIMAS.
O piruvato, derivado da glicose através da via glicolítica, é oxidado para liberar acetil-CoA e CO2 por um conjunto
estruturado de três enzimas, o complexo piruvato desidrogenase, localizado na mitocôndria das células eucarióticas e no
citosol das procarióticas. O complexo piruvato desidrogenase é o protótipo de dois outros importantes complexos
enzimáticos que encontraremos adiante: o da -cetoglutarato desidrogenase e o da desidrogenase dos -cetoácidos com
cadeia ramificada, envolvida na degradação oxidativa de vários aminoácidos. Provavelmente há uma origem
evolucionária em comum para os três.
As cinco coenzimas necessárias são: TTP – tiamina pirofosfato, FAD – flavina adenina dinucleotídeo, CoA - coenzima
A, NAD – nicotinamida adenina dinucleotídeo e lipoato. Podemos observar que quatro delas estão presentes na nossa
alimentação. NAD e FAD transportam elétrons, CoA com seu grupo reativo –SH transporta grupos acila e o lipoato pode
servir tanto como transportador de elétrons quanto como de grupos acil.
As três enzimas são: Piruvato desidrogenase, diidrolipoil transacetilase e diidrolipoil desidrogenase.
8.
VERIFICAR QUE A FORMAÇÃO DE acetil-CoA COINCIDE TAMBEM COM A PRIMEIRA REACAO
DE DESCARBOXILACAO DOS ATOMOS DE CARBONO DA GLICOSE.
9.
OBSERVAR QUE A AUSÊNCIA DE COFATORES OU MUTAÇÃO EM QUALQUER DAS ENZIMAS DO
COMPLEXO PIRUVATO DESIDROGENASE PODE ACUMULAR PIRUVATO, ALANINA E LACTATO
(DEVLIN, PÁG:196).
Uma série de doenças no metabolismo de piruvato já foram detectadas em crianças. Algumas envolvem deficiências nas
diferentes subunidades catalíticas ou regulatórias do complexo multienzimatico da piruvato desidrogenase. Crianças
diagnosticadas com deficiência de piruvato desidrogenase, normalmente, apresentam níveis séricos elevados de lactato,
piruvato e alanina, que produzem uma acidose láctica crônica. Tais pacientes, freqüentemente, exibem defeitos
neurológicos severos e, na maioria das situações, esse tipo de defeito enzimático resulta em morte. O diagnostico de
deficiência de piruvato desidrogenase é, normalmente, feito pelo ensaio do complexo enzimático, e/ou suas varias
subunidades enzimáticas, em cultura de fibroblastos de pele retirados do paciente. Em alguns casos, os paciente
respondem a terapia dietética na qual uma dieta cetogenica é administrada e os carboidratos são minimizados. Pacientes
em choque têm acidose láctica porque o decréscimo na distribuição de O2 para os tecidos inibe a piruvato desidrogenase
e aumenta o metabolismo anaeróbico. Essa situação tem sido tratada com dicloroacetato, um inibidor da piruvato
desidrogenase quinase e portanto, um ativador do complexo enzimático.
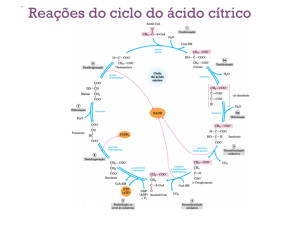
10. SABER QUE O CICLO DE KREBS APRESENTA AO TODO OITO REAÇÕES , SENDO A PRIMEIRA
DELAS A CONDENSAÇÃO DO acetil-CoA COM O OXALOACETATO DANDO CITRATO.
Para iniciar uma volta no ciclo, o acetil-CoA transfere o seu grupo acetila para um composto com quatro átomos de
carbono, o oxaloacetato, para formar o citrato, um composto com seis átomos de carbono.
As reações do ciclo de Krebs são:
citratosintase
citrato
acetil-CoA + oxaloacetato
cis-aconitato
isocitrato
citrato
aconitase
aconitase
istocitrato desidrogenase
isocitrato -cetoglutarato + CO2
complexo cetoglutarato desidrogenase
-cetoglutarato succinil-CoA +CO2
succinilCoAsin tetase
succinil-CoA succinato
succinato desidrogenase
succinato fumarato
malato
fumarato
fumarase
malato desidrogenase
oxaloacetato
malato
11. RECONHECER AS DEMAIS REACOES DO CICLO DE KREBS PARTICULARMENTE, AS QUE
FORMAM NADH + H+ E FADH2; AS DE DESCARBOXILAÇÃO E A DE FOSFORILAÇÃO AO NÍVEL
DO SUBSTRATO.
citratosintase
citrato
acetil-CoA + oxaloacetato
Esta reação é altamente exergônica no sentido do produto. A variação de energia livre negativa nesta reação é essencial
para a operação do ciclo porque normalmente a quantidade de oxaloacetato presente é muito baixa. A CoA deixada nesta
reação é reciclada e pode vir a participar das varias reações em que atua. A citrato sintase está na matriz mitocondrial.
cis-aconitato
isocitrato
citrato
A enzima aconitase catalisa a transformação reversível do citrato em isocitrato através da formação intermediária do cisaconitato, um ácido tricarboxílico que, normalmente, não se dissocia do sítio ativo. Embora a mistura contenha menos de
10% de isocitrato na célula, a reação é descolada para a direita porque o isocitrato é rapidamente consumido no passo
subseqüente do ciclo, diminuindo a concentração de equilíbrio estacionário. Embora essa reação não necessite de cofatores, ela requer Fe2+ no seu mecanismo catalítico.
aconitase
aconitase
istocitrato desidrogenase
isocitrato -cetoglutarato + CO2
No passo seguinte a isocitrato desidrogenase catalisa a descarboxilacao oxidativa do isocitrato para formar o cetoglutarato. Formam-se o primeiro CO2 (de dois) e os primeiros NADH + H+. O equilíbrio desta reação é para a
formação dos produtos. Existem duas “versões” desta enzima, uma que emprega o NAD+ e outra que emprega o
NADP+. A enzima dependente de NAD é encontrada na matriz mitocondrial e participa do ciclo de Krebs. A outra
enzima está tanto na matriz quanto no citosol e sua função é gerar NADPH.
complexo cetoglutarato desidrogenase
-cetoglutarato succinil-CoA +CO2
Nessa descarboxilacao oxidativa, o -cetoglutarato é convertido em succinil-CoA e CO2 pela ação do complexo cetoglutarato desidrogenase; o NAD serve como aceptor de elétrons. De novo, TTP, lipoato, CoASH, FAD e NAD+
participam do mecanismo catalítico. As duas reações são virtualmente idênticas. Envolvem a oxidação de um cetoácido com a perda do grupo carboxila na forma de CO2 e a energia de oxidação do -cetoglutarato é conservada na
forma de tio-éster do succinil-CoA.
succinilCoAsin tetase
succinil-CoA succinato
Como o acetil-CoA o succinil-CoA tem uma energia livre de hidrólise de sua ligação tioéster forte e negativa. No
próximo passo do ciclo do ácido cítrico a energia liberada no rompimento dessa ligação é empregada para dirigir a
síntese de uma ligação de anidrido fosfórico no ATP ou no GTP, e, finalmente, formam-se succinato livre. A enzima
desta reação é succinil-CoA-sintetase é fosforilada na posição 3 de um resíduo de histidina. Essa síntese de GTP é um
exemplo de fosforilação ao nível do substrato. ATP e GDP são energeticamente equivalentes.
succinato desidrogenase
succinato fumarato
O succinato formado a partir do succinil-CoA é oxidado a fumarato pela enzima fumarato desidrogenase, que é
fortemente ligada à membrana mitocondrial interna (a única). Os elétrons tirados do succinato passam através do FAD e
dos centros ferro-enxofre antes de entrarem na cadeia transportadora de elétrons na membrana mitocondrial interna. O
fluxo de elétrons do succinato até o O2, o receptor final de elétrons, através desses transportadores, está acoplado à
síntese de duas moléculas de ATP por par de elétrons.
malato
fumarato
A hidratação reversível do fumarato em malato é catalisada pela fumarase. Esta enzima é altamente estereoespecifica;
ela catalisa a hidratação da dupla ligação trans do fumarato, porém não é capaz de agir no isomero-cis do fumarato.
fumarase
malato desidrogenase
oxaloacetato
malato
Na última reação do ciclo de Krebs, a malato desidrogenase, ligada ao NAD, catalisa a oxidação do malato em
oxaloacetato. O equilíbrio desta reação esta muito deslocado para a esquerda sob condições termodinâmicas padrão.
Entretanto, nas células intactas, o oxaloacetato é continuamente removido pela reação da citrato sintase que é altamente
exergônica. Isto conserva a concentração do oxaloacetato na célula em valores extremamente pequenos, deslocando a
reação da malato desidrogenase para direita, ou seja, a formação do oxaloacetato.
12. CALCULAR O NÚMERO DE MOLÉCULAS DE ATP QUE PODEM SER OBTIDAS POR CADA
MOLÉCULA DE GLICOSE DEGRADADA ATÉ CO2 E H2O.
Durante a completa oxidação da glicose até CO2 e H2O, são formadas:
2 moléculas de ATP por glicose, na conversão de glicose a 2 moléculas de piruvato;
6 moléculas de ATP por glicose como resultado da translocação e subseqüente oxidação na matriz mitocondrial
de 2 moléculas de NADH + H+, formadas na reação do gliceraldeido-3-fosfato desidrogenase na glicólise;
30 moléculas de ATP por glicose, a partir da oxidação das 2 moléculas de piruvato na reação da piruvato
desidrogenase e a subseqüente conversão de 2 moléculas de acetil-CoA e CO2 e H2O, no ciclo dos ácidos
tricarboxílico. Assim, o rendimento líquido de ATP durante a completa oxidação da glicose a 6CO 2 + 6H2O é de
38 moléculas de ATP.
13. SABER AS ESTAPAS DE REGULAÇÃO DO CICLO DE KREBS.
O fluxo de átomos de carbono do piruvato pára e através do ciclo de Krebs é estreitamente regulado em dois níveis: a
conversão de piruvato em acetil-CoA (reação do complexo piruvato desidrogenase), o material inicial do ciclo, e a
entrada de acetil-CoA no ciclo (reação da citrato sintase). O ciclo também é regulado na altura da reação da isocitrato
desidrogenase e na reação do complexo -cetoglutarato desidrogenase.
Piruvato desidrogenase: é regulado alostericamente e através de modificação covalente. O complexo é fortemente inibido
por ATP e por acetil-CoA, produtos da reação. Quando muito pouco acetato flui para o ciclo de Krebs, acumulam-se
AMP, CoA e NAD+, todos eles ativam alostericamente o complexo da piruvato desidrogenase. Assim, a atividade dessa
enzima é desligada quando as substancias combustíveis estão disponíveis de maneira ampla na forma de ácidos graxos e
acetil-CoA e quando a concentração de ATP celular e a relação NADH/NAD+ estão altas.
Há de se dizer ainda que este complexo existe sob duas formas: (1) uma ativa, complexo desfosforilado, e uma (2)
inativa, fosforilada. A inativação deste complexo é realizada por uma proteína quinase Mg2+-ATP- dependente, que está
firmemente ligada ao complexo enzimático. A reativação é realizada por uma proteína fosfatase, que desfosforila o
complexo em uma reação Mg2+ e Ca2+ dependente. Essa é uma regulação diferencial é a chave do controle.
Três fatores regulam a velocidade do fluxo através do ciclo: disponibilidade de substratos, inibição por acúmulos de
produtos e inibição alosterica reativa das primeiras enzimas da via pelos últimos intermediários.
O malonato, um análogo do succinato, é um potente inibidor competitivo da succinato desidrogenase e por este
motivo é bloqueador do ciclo do ácido cítrico.
No tecido muscular, Ca2+ sinaliza o início da contração e estimula o metabolismo liberador de energia, de maneira a
repor o ATP consumido pela atividade muscular.
14. ENTENDER O PAPEL DO CICLO DOS ÁCIDOS TRICARBOXÍLICOS NO ANABOLISMO E AS
REAÇÕES ANAPLERÓTICAS DO MESMO.
Nos organismos aeróbicos, o ciclo de Krebs é uma via anfibolica, isto é, serve tanto para processos anabólicos quanto
para catabolicos. Ela não funciona apenas no catabolismo oxidativo de carboidratos, ácidos graxos e aminoácidos, mas,
como progenitores anaeróbicos, também fornece precursores para muitas vias biossiteticas. Certos intermediários do
ciclo, particularmente o -cetoglutarato e oxaloacetato, podem ser removidos do mesmo para servirem como precursores
de aminoácidos.
Quando os intermediários são retirados para servirem de precursores biossinteticos, o resultante decréscimo na
concentração desses intermediários deveria provocar um rebaixamento no fluxo, entretanto, esses intermediários podem
ser fornecidos novamente por meio das reações anapleróticas. Uma reação anaplerótica importante nos tecidos animais é
a carboxilação reversível do piruvato por CO2, para formar oxaloacetato, catalisada pela piruvato carboxilase. Quando o
ciclo de Krebs está deficiente em oxaloacetato ou qualquer um de seus intermediários, o piruvato é descarboxilado para
produzir mais oxaloacetato.
Reações anapleróticas:
piruvato carboxilase
oxaloacetato + ADP + Pi
Piruvato + HCO-3 + ATP
Fígado, rim
Fosfoenolpiruvato + CO2 + GDP oxaloacetato + GTP
Coração, musc.esqul
PEPcarboxiquinase
Fosfoenolpiruvato + HCO 3 oxaloacetato + Pi
-
PEPcarboxilase
enzimamálica
Piruvato + HCO 3 + NADPH malato + NADP
-
+
Vegetais, bactérias
Largamente distribuída nos euc/proc