DEMIAN WASHINGTON FONSECA
ANÁLISE DA ALTERAÇÃO LEUCOCITÁRIA PLASMÁTICA EM
RESPOSTA AO EXERCÍCIO AGUDO AERÓBIO.
CURlTIBA
2001
DEMIAN W ASBINGTON FONSECA
ANÁLISE DA ALTERAÇÃO LEUCOCITÁRIA PLASMÁTICA EM
RESPOSTA AO EXERCíCIO AGUDO AERÓBIO.
Monografia apresentada como Critério de A
vaUação parcial para Conclusão do Curso de
Fisioterapia do Setor de Ciências Biológicas e
da Saúde da Universidade Tuiuti do Paraná.
Orientado r: Pror. Luis Câmara
Co-orientadora: Dra. Regina CéU Mazepa
CURITIBA
2001
1
1
--- --------------- --
AGRADECIMENTOS ESPECIAIS
A meus pais, meus primeiros mestres na ciência de viver, que pela amizade, pela
dedicação e pelo diálogo, muito me ajudaram nos momentos difíceis.
Ao orientador Luiz Câmara pelo apoio fornecido no decorrer da pesquisa.
A Dra. Regina Céli Mazepa pela confiança, incentivo e orientações que foram de
extrema importância para a realização desta pesquisa.
Aos supervisores Shirley Vargas e Luiz Antonio Negrão pelo auxilio na
elaboração estrutural da pesquisa.
Aos mestres e aos colegas: aos funcionários da instituição, as pessoas que
formaram a população amostral ao Hematologista clínico Hemerson Bertossoni Alves, e
aos tantos amigos que aqui deixo minha homenagem e o tributo de minha gratidão.
1
1l
---
"Não importa qual seja seu sonho,
dedique a ele muito de seu tempo e de
seu pensamento. É preciso ter amor e
paixão pelo o que você está fazendo
para achar a energia a formação e
aplicação necessária para ter sucesso
em qualquer tipo de atividade. Se você
decide lutar por alguma coisa, tem que
dar o máximo afim de vencer..
Ayrton Senna da Silva
IV
-
--
SUMÁRIO
LISTA DE ABREViATURAS................................................................ VII
RESU MO. .....................................................................................................a ........... VIII
RESU M EN ........................................................................................ .IX
1.
INTRODUÇAO...........................................................................01
2.
REVISAO LITERARIA...............................................................02
2.1
CÉLULAS SANGuíNEAS...........................................................02
2.1.1
Glóbulos vermelhos ...................................................................02
2.1.2
Produção das hemácias.............................................................03
-,
2.1.2.1 áreas do organismo produtoras de hemácias .............................03
2.1.3
Gênese das células sanguíneas ................................................04
2.1.4
Estágios da diferenciação das hemácias ...................................05
2.1.5
Papel do rim na produção de eritropoetina ................................06
2.2
LEUCÓCITOS........................................................................... .06
2.2.1
Características gerais dos leucócitos..........................................06
2.2.2
Concentração dos diferentes glóbulos brancos no sangue ........07
2.2.3
Gênese dos leucócitos.................................................................07
2.2.4
Vida média do glóbulos brancos.................................................08
2.2.5
Propriedrades defensivas dos neutrófios e macrófagos .............. 09
2.2.6
Fagocitose .................................................................................1 O
2.2.7
Sistema monócito - macrófago ...................................................11
2.2.8
Eosinófilos...................................................................................12
2.2.9
Basófilos .................................................................................... .13
2.3
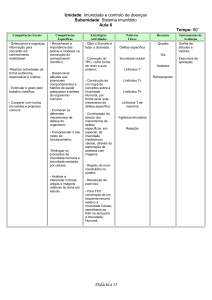
IMUNIDADE.............................................................................. ...14
2.3.1
Imunidade natural ...................................................................... .14
2.3.2
Imunidade adquirida....................................................................15
2.3.2.1 tipos básicos de imunidade adquirida............................................. ..15
v
--
2.3.2.2 os dois tipos de imunidade adquirida que são iniciadas pelos
antígenos....................................................................................16
2.3.2.3 os linfócitos como base na imunidade adquirida .........................17
2.3.2.4 pré-processamento dos linfócitos T e 8 .......................................18
2.4
FISIOLÓGIA DO EXERCíCIO ....................................................20
2.4.1
Sistema A TP - CP ................................................................... ...20
2.4.2
Metabolismo anaeróbio . ........................................................... .21
2.4.3
Metabolismo aeróbio ................................................................. .21
2.4.4
Utilização dos substratos energéticos no exercício ....................21
2.5
RESPOSTAS IMUNES AO EXERCícI0 ......................................22
3.
MATERIAIS E METODOS ........................................................25
4.
APRESENTAÇÃO DOS DADOS COLETADOS.........................26
4.1
DADOS ESTATíSTICOS DA PESQUISA...................................26
4.1.2
Medidas de dispersão.............................................................. ...27
4.2
REPRESENTAÇÕES GRÁFICAS DA PESQUISA.....................28
5.
ANALISE E DISCUSSÃO DOS DADOS .....................................31
6.
CONCLUSAO .. .........................................................................33
7.
REFERENCIAS BIBLíOGRAFICAS ................................................ 34
V
I
Lista de Abreviaturas
At (antes)
exercício.
-
Amplitude total da amostra, antes da realização do
At (depois) - Amplitude total da amostra, depois da realização do
exercício.
X (antes)
exercício.
Média aritmética, antes da realização do
X
exercício.
Média aritmética, depois da realização do
(depois)
X (diferença) - Média aritmética, da diferença entre os valores obtidos
antes e depois do exercício.
X difer.(%)
diferença entre os
exercício.
Média aritmética em percentual, valores
obtidos antes e depois
da
do
Di - Desvio do i-ézimo elemento, ou seja, desvio de cada elemento em
relação à média aritmética dos mesmos.
Dm - Desvio médio.
Dp - Desvio padrão.
P - Significância.
Fc = frequência cardíaca
NK = natural killer
V02 máx = Consumo Máximo de Oxigênio
vii
--
-
Resumo
Este trabalho tem como objetivo analisar a contagem de leucócitos, comparando
os valores obtidos antes e depois da realização do exercício agudo aeróbio e
cíclico. Foram selecionados para este estudo 10 pessoas, pelos seguintes
critérios de inclusão : peso corporal entre 70 a 82 Kg, idade entre 21 a 25 anos,
sadias, sexo masculino, não tabagistas, não etilistas, de raça branca, e com o
desenvolvimento prático desta pesquisa sendo no período da tarde. Para a
realização prática, as pessoas que formaram esta população amostral
submeteram-se a coleta sangüínea antes e depois do exercício na
cicloergométrica de intensidade moderada com duração de 30 minutos e com a
freqüência cardíaca de treinamento a 60% do V02 máx, foram então analisadas
as amostras sangüíneas obtidas e posteriormente efetuada uma análise
estatística dos dados comparados pareamente antes e depois do exercício,
onde observou-se que ocorreu uma significativa leucocitose em resposta ao
exercício. Consequentemente pode-se concluir que o exercício físico de
intensidade moderada em cicloergométrica, altera a composição percentual da
concentração celular sangüínea.
Palavras - Chave: Exercício físico, Contagem de leucócitos
viii
RESUMEN
Este trabajo tiene como objetivo analisar el contage de leucócitos comparando los
valores obtenidos antes y después de Ia realización dei ejercicio agudo aeróbico y
cíclico. Fueron seleccionados para esta investigación 10 personas por los siguientes
criterios de inclusión: peso corporal de 70 a 82 kg, edad de 21 a 25 anos, sexo
masculino, saludables, no tabagistas, no etilistas, de raza blanca, y com el desarrollo
práctico de esta investigación siendo por Ia tarde. Para Ia realización práctica, Ias
personas que formaron Ia población amostral se sometieron a una coleta sanguinea
antes y después dei ejercicío en el cicloergométrica de intensidad moderada con
duración de 30 minutos y con Ia frecuencia cardiaca de entrenamiento a 60% dei V02
máx, fueron entonces analizadas estas muestras sangüineas obtenidas y posteriormente
fue hecho un análisis estadístico de los datos comparados pareamente antes y después
dei ejercicio, dónde fue observado que ocurrió una leucocitose significante en Ia
respuesta ai ejercicios. Por consiguinte se puede concluir que el ejercicio físico de
intensidad moderada en el circloergométrica altera Ia composición dei porcentaje de Ia
concentración celular sangüinea.
Palabras - clave: Ejercicio físico, Contage de leucócitos
ix
1. INTRODUÇÃO
Em 1899, os médicos recomendavam a pacientes exercícios para
protegê-I os contra a trombose venosa profunda no pós-operatório. Com o
avanço dos estudos nas atividades físicas e esportivas, diferentes estudiosos
começaram a recomendar o exercício para o controle, tratamento e prevenção
de diversas doenças, aumentando a longevidade e melhorando a qualidade de
vida.
o
exercício físico provoca um desvio da situação de homeostase
induzindo uma reorganização das respostas funcionais dos diversos sistemas
orgânicos, incluindo a resposta inflamatória aos sistemas neuroendócrinos e
sistema imune.
A relação entre estes é hoje bem conhecidas e a interação por meio de
hormônios, neuropeptídeos e citocinas explica muitas das alterações que
ocorrem nestes sistemas após o exercício.
Este trabalho tem por objetivo analisar a contagem total de leucócitos
circulantes, comparando os valores obtidos antes e depois da realização do
exercício agudo aeróbio e cíclico (cicloergométrica) de intensidade moderada, e
despertar o interesse de fisioterapeutas para a realização de novas pesquisas
mais aprofundadas sobre os componentes do sistema imunológico
em função do exercício, abrindo um novo campo de atuação para a fisioterapia.
Este estudo compreende o embasamento teórico, os materiais e métodos
utilizados, apresentação dos dados coletados, análise e discussão dos dados,
conclusão, e ao final as referências bibliográficas.
2
2. REVISÃO LITERÁRIA
2.1. CÉLULAS SANGuíNEAS
2.1.1. Glóbulos vermelhos
A principal função dos glóbulos vermelhos, também conhecidos como
eritrócitos, é transportar a hemoglobina, a qual, por seu turno, transporta oxigênio dos
pulmões para os tecidos. Em alguns animais inferiores a hemoglobina circula como
proteína livre no plasma, não estando contida nos glóbulos vermelhos. Quando está
livre no plasma do ser humano, cerca de 3% atravessam a membrana capilar para os
espaços teciduais ou através da membrana glomerular renal para o
filtrado glomerular, cada vez que o sangue passa pelos capilares.
Em
consequência, para que a hemoglobina permaneça na corrente sanguínea, ela só
deve existir dentro dos glóbulos vermelhos.
Os glóbulos vermelhos tem outras funções além do transporte de hemoglobina.
Por exemplo, eles contêm uma grande quantidade de anidrase carbônica, que catalisa
a reação entre dióxido de carbono e água, aumentando a velocidade dessa reação
reversível por cerca de 1000 vezes. A rapidez dessa reação possibilita que a água no
sangue reaja com grandes quantidades de dióxido de carbono, transportando-o assim
dos tecidos para os pulmões na forma de íon bicarbonato. A hemoglobina das células
também é um excelente tampão ácidobásico (como o é a maioria das proteínas);
desse modo, os glóbulos vermelhos são responsáveis pela maior parte do poder
tampão de todo o sangue(GUYTON &
HALL, 1996).
Os glóbulos vermelhos normais, são discos bicôncavos com diâmetro de cerca
de 7,8f.lm3 ou menos no centro. O volume médio dos glóbulos vermelhos é de 90 a
95f.lm3.
A forma dos glóbulos vermelhos pode mudar extraordinariamente ao passarem
através dos capilares. Na realidade, o glóbulo vermelho é um "saco" que pode ser
deformado em quase qualquer forma. Além disso, como a célula normal possui um
excesso de membrana plasmática em relação ao seu conteúdo, a
--
-
3
deformação não distende muito a membrana e, consequentemente, não rompe a
célula, como aconteceria com muitas outras
No homem normal, o número médio de glóbulos vermelhos por milímetro
cúbico é de 5.200.000 ( :t: 300.000 ) e, na mulher 4.700.000 ( :t: 300.000 ). A altitude
que a pessoa vive afeta o número de glóbulos vermelhos que ela possui.
As hemácias possuem a capacidade de concentrar hemoglobina no líquido
citoplasmático até cerca de 34g/ml de células. A concentração nunca ultrapassa este
valor porque este é o limite metabólico do mecanismo celular de formação de
hemoglobina. Além disso, nas pessoas normais a percentagem de hemoglobina em
cada hemácia é sempre próxima do nível máximo. Todavia, quando a formação de
hemoglobina é deficiente na medula óssea, a percentagem de hemoglobina nas
hemácias pode cair consideravelmente abaixo desse valor, e o volume das células
pode diminuir devido a falta de hemoglobina para preencher a célula.
Quando o hematócrito (a percentagem do sangue que é constituída por células
-
40 a 45 % normalmente) e a quantidade de hemoglobina em cada hemácia são normais, o
sangue total dos homens contêm uma média de 16g de hemoglobina por 100 mi, e o
das mulheres, uma média de 14g por 100 ml. Portanto, no homem normal, mais do
que 21 ml de oxigênio podem ser transportados em combinação com a hemoglobina
de cada 100ml de sangue e, na mulher normal 19m1 de oxigênio podem ser
transportados.
2.1.2. Produção das hemácias
2.1.2.1 Áreas do organismo produtoras de hemácias
Nas primeiras semanas da vida embrionária, hemácias nucleadas primitivas
são produzidas no saco vitelino. Durante o segundo trimestre da gestação, o fígado é
o principal orgão de produção de hemácias, embora moderada quantidade também
seja produzida pelo baço e linfonódos. Em seguida, durante o último mês da gestação
e após o nascimento, as hemácias são produzidas exclusivamente pela medula
óssea. Fundamentalmente a medula óssea de todos os ossos produz hemácias até
os cinco anos de idade ; mas a medula dos ossos longos, com
4
exceção da porção proximal do úmero e tíbia, toma-se gordurosa e não produz mais
hemácias depois dos vinte anos. Acima dessa idade as células vermelhas são
produzidas na medula dos ossos membranosos, tais como as vértebras, o estemo, as
costelas e o ilíaco. Mesmo nesses ossos, a medula toma-se menos produtiva com o
avançar da idade. (GUYTON & HALL, 1996)
2.1.3. Gênese das células sanguíneas
Na medula óssea estão as células denominadas células-tronco hemopoiéticas
pluripotenciais, a partir das quais derivam todas as células do sangue circulante. As
divisões sucessivas das células pluripotenciais para formar diferentes células
sanguíneas periféricas. À medida que essas células se reproduzem, continuando
através da vida do indivíduo uma parte delas permanece exatamente como a célula
pluripotencial original, sendo retida na medula óssea para manter um suprimento de
células, embora sua quantidade diminua com a idade. A progênie precoce ainda não
pode ser identificada como diferente das células - tronco- pluripotenciais, embora se
tenham tomado comprometidas com uma linhagem celular particular e sejam
chamadas de células - tronco comprometidas.
As diferentes células - tronco comprometidas , quando crescem em cultura
produzirão colônias de tipos específicos de células sanguíneas. Uma célula-tronco
comprometida que se produz eritrócitos é chamado de unidade formadora de colônias
de eritrócitos, e abreviação CFU- E é usada para designar esse tipo de célula - tronco.
Do mesmo modo, unidades formadoras de colônia que formam granulócitos e
monócitos têm designação de CFU- GM e assim por diante.
O crescimento e a reprodução das células-tronco são controlados por múltiplas
proteínas denominadas indutores de crescimento. Quatro indutores de crescimento
principais foram descritos, cada um com características diferentes. Um deles,
interleuquina- 3, promove o crescimento e a reprodução de todos os diferentes tipos
de células-tronco, enquanto os outros induzem o crescimento de tipos específicos de
células- tronco comprometidas.
Os indutores de crescimento promove
o crescimento,
mas não a
diferenciação das células. Essa é a função de outro conjunto de proteínas,
5
denominado indutores de diferenciação. Cada um desses indutores determina a
diferenciação de um tipo de célula-tronco em uma ou mais etapas em direção ao tipo
final de célula sanguínea adulta (GUYTON & HALL, 1996).
A produção de indutores de crescimento e diferenciação é contolada por
fatores situados fora da medula óssea. Por exemplo, no caso das hemácias, a
exposição do organismo à oxigênio insuficiente durante um longo período resulta na
redução de crescimento, diferenciação e produção de grande quantidade de
eritrócitos. No caso de alguns leucócitos as doenças infecciosas causam crescimento,
diferenciação e eventual formação de tipos específicos de leucócitos necessários para
combater a infecção.
2.1.4. Estágios da diferenciação das hemácias
A primeira célula que pode ser identificada como da série vermelha é o próeritroblasto. Sob estimulação apropriada grandes quantidades dessa célula são
formadas a partir da célula-tronco CFU-E.
Logo que o pró-eritroblasto é formado, ele se divide várias vezes e,
consequentemente, forma muitas hemácias maduras. As primeiras linhagens
celulares são chamadas de eritoblastos basófilos, porque se coram com corantes
básicos; nesse momento, a célula acumulou pouca hemoglobina. Nas etapas
sucessivas as células ficam preenchidas com hemoglobina a uma concentração de
aproximadamente 34%, o núcleo condensa para um tamanho pequeno e os seus
restos são expulsos da célula. A célula neste estágio é chamada de reticulócito
porque ainda contêm uma pequena quantidade de material basófilo que consiste em
restos
do
aparelho
de
Golgi,
mitocôndrias
e
algumas
outras
organelas
citoplasmáticas. Durante o estágio de reticulócito, as células passam da medula óssea
para os capilares por diapedese ( passando através dos poros da membrana capilar).
O material basófilo remanescente no reticulócito desaparece normalmente
dentro de um ou dois dias, e a célula é , então, o eritrócito maduro. Devido a vida
curta dos reticulócitos, sua concentração entre as hemácias é normalmente menor
que 1%. (GUYTON & HALL, 1996)
--
6
2.1.5. Papel do rim na produção de eritropoetina
Na pessoa normal, aproximadamente 90% do total de eritropoetina são
produzidos no rim; o restante é formado principalmente no fígado. Não se sabe
exatamente onde a eritropoetina é produzida nos rins. Uma possibilidade é que as
células epiteliais dos túbulos renais secretam eritropoetina, porque o sangue anêmico
dos capilares peritubulares, sendo incapaz de transferir oxigênio suficiente para as
células tubulares consumidoras de oxigênio, consequentemente estimula a produção
de eritropoetina.
Às vezes, a hipóxia em outras partes do corpo, mas não no rim, também
estimula a produção de eritropoetina, sugerindo que deve existir um sensor não renal
que envia uma sinalização adicional para que os rins produzam esse hormônio. Em
particular tanto a norepinefrina quanto a epinefrina e várias prostaglandinas estimulam
a produção de eritropoetina.
2.2. LEUCÓCITOS (GLÓBULOS BRANCOS)
Os leucócitos são as unidades móveis do sistema de proteção do organismo.
São formados em parte na medula óssea (os granulócitos, monócitos e alguns
linfócitos) e, em parte, no tecido linfóide (Iinfócitos e plasmócitos). Após sua formação,
são transportados pelo o sangue para as diferentes partes do corpo onde serão
utilizados. O valor real dos glóbulos refere-se ao transporte específico da maioria
deles para as áreas de inflamação e infecção grave, fornecendo uma defesa rápida e
potente contra qualquer agente infeccioso que possa estar presente.
2.2.1. Características gerais dos leucócitos
Seis tipos de glóbulos brancos são normalmente encontrados no sangue. São
os
neutrófilos
polimorfunucleares,eosinófilos
polimorfunucleares,
basófilos
polimorfunucleados, monócitos, linfócitos, e ocasionalmente, os plasmócitos. Além
desses, há um grande número de plaquetas, que são fragmentos de um sétimo tipo
-
7
de glóbulo branco encontrado na medula óssea : os megacariócitos. Os primeiros três
tipos, os polimorfunucleares, têm uma aparência granulosa, razão pela qual são
chamados de granulócitos, ou, na terminologia clínica, "poli", devido ao núcleo
multilobado.
Os granulócitos e monócitos protegem o organismo contra os organismos
invasores, principalmente ingerindo-os isto é, por fagocitose. Os linfócitos e
plasmócitos funcionam juntamente com o sistema imune. Finalmente, a função das
plaquetas é principalmente ativar o mecanismo de coagulação sanguínea. (GUYTON
& HALL, 1996)
2.2.2. Concentração dos diferentes glóbulos brancos no sangue
O adulto possui aproximadamente 7.000 leucócitos por microlitro de sangue.
As
percentagens
normais
dos
diferentes
tipos
de
glóbulos
brancos
são
aproximadamente as seguintes.
Neutrófilos polimorfunucleados
62,0%
Eosinófilos plimorfunucleados
2,3
Basófilos polimorfunucleados
0,4 %
Monócitos
5,3 %
Linfócitos
30,0%
%
O número de plaquetas, as quais são somente fragmentos celulares, é
normalmente de cerca de 300.000 por microlitro de sangue.
2.2.3. Gênese dos leucócitos
Os granulócitos e monócitos são formados somente na medula óssea.
Linfócitos e plasmócitos são produzidos, principalmente, nos vários orgãos linfóides
incluindo os ganglios linfáticos, baço, timo, tonsila e diversas concentrações de
8
tecido linfóide em outras áreas do corpo, especialmente na medula óssea e nas
placas de Peyer abaixo do epitélio na parede intestinal.
Os glóbulos brancos
formados na medula óssea, especialmente os
granulócitos, são armazenados na medula até serem necessários no sistema
circulatório. Então, quando surge uma emergência, vários fatores, induzem a
liberação dos leucócitos. Normalmente, cerca de três vezes o equivalente dos
granulócitos que circulam no sangue total são armazenados na medula óssea . Isto
representa aproximadamente seis dias de suprimento de granulócitos.
Os linfócitos são, na sua maioria, armazenados em várias áreas de tecido
linfóide, com exceção de um pequeno número que está sendo transportado no
sangue temporariamente.
Os megacariócitos também são formados na medula óssea e fazem parte do
grupo mielógeno das células da medula óssea. Estes megacariócitos fragmentam-se
na medula óssea; os pequenos fragmentos conhecidos como plaquetas ou
trombócitos passam para o sangue.
2.2.4. Vida média dos glóbulos brancos
A razão principal da presença dos glóbulos brancos no sangue é a de serem
transportados da medula óssea para o sangue ou do tecido linfóide para as áreas do
corpo onde são necessários. Uma vez liberados da medula óssea, a vida dos
granulócitos é de 4 a 8 horas circulando no sangue e quatro a cinco dias em outros
tecidos. Durante uma infecção grave, este período de vida diminui para poucas horas
somente,
porque
os
granulócitos
atuam
rapidamente
na
área
infectada,
desempenham suas funções e são destruidos no processo.
Os monócitos também têm uma vida curta, de 10 a 20 horas no sangue, antes
de atravessarem as membranas dos capilares para os tecidos. Uma vez nos tecidos,
eles se avolumam e diferenciam-se em macrófagos teciduais e nessa configuração,
podem viver durante meses ou mesmo anos, a não ser que sejam destruidos
realizando a função de fagocitose. Estes macrófagos teciduais constituem a base do
sistema dos macrófagos dos tecidos, responsáveis por uma defesa continua nos
tecidos contra infecções. (GUYTON & HALL, 1996).
9
Os linfócitos entram no sistema circulatório continuamente, junto com a linfa
drenada dos linfonodos e outros tecidos linfóides. Em seguida, após algumas horas,
passam para os tecidos por diapedese, depois retomam à linfa e voltam para o tecido
linfóide ou para o sangue várias vezes; consequentemente, existe uma circulação
contínua de linfócitos pelo organismo. Os linfócitos têm uma vida média de semanas,
meses ou mesmo anos, dependendo das necessidades do organismo em relação a
essas células.
As plaquetas no sangue são substituídas a cada 10 dias; em outras palavras,
cerca de 30.000 plaquetas são formadas por dia para cada microlitro de sangue.
2.2.5. Propriedades defensivas dos neutrófilos e macrófagos
Essencialmente são os neutrófilos e macrófagos que atacam e destroem as
bactérias, vírus e outros agentes infecciosos. Os neutrófilos são células maduras que
podem atacar e destruir bactérias e vírus no sangue circulante. Por outro lado, os
macrófagos começam a vida como monócitos no sangue, que são células imaturas,
enquanto estão no sangue, e tem pouca habilidade para combater agentes
infecciosos. Entretanto, após entrarem nos tecidos, começam a se avo lu mar,
algumas vezes aumentam seu diâmetro até cinco vezes alcançando 80 ~m, tamanho
suficiente para serem vistos a olho nu. Também desenvolvem um número grande de
lisossomos no citoplasma, dando-Ihes uma aparência de um saco cheio de grânulos.
Estas células são chamadas de macrófagos e são extremamente
capazes de combater agentes infecciosos.
Os glóbulos brancos entram nos espaços teciduais por diapedese .
Neutróflos e monócitos
podem passar através dos poros dos vasos
sanguíneos por diapedese. Isto é, embora o poro seja muito menor que o tamanho da
célula, uma pequena parte da célula desliza através do poro, e esta porção que
desliza é comprimida até o tamanho do poro.
Glóbulos brancos movem-se através dos espaços teciduais por movimento
amebóide.
-
.._~,.......... _ __~__L1~~~_ ____________ __ __1__ .&.__: _______________ : ____ &.____a...;..:...._
Algumas dessas células podem mover-se por uma velocidade de 40 I-Im/min, isto é,
várias vezes seu próprio comprimento a cada minuto.
Os glóbulos brancos são atraídos para as áreas de tecidos inflamados por
quimiotaxia.
Diversas substâncias químicas do tecido induzem a movimentação dos
neutrófilos e macrófagos na direção da origem da substância. Este fenômeno é
conhecido como quimiotaxia . Quando um tecido se torna inflamado, são formados pelo
menos 12 produtos diferentes que podem induzir a quimiotaxia na direção da área
inflamada. (GUYTON & HALL, 1996).
A quimiotaxia
depende do gradiente de concentração da substância
quimiotática. A concentração é maior perto da origem, o que induz o movimento
direcional dos glóbulos brancos. A quimiotaxia é eficiente até 100l-lm além do
tecido inflamado; como quase nenhum tecido está distante por mais de 50 I-Im
de um capilar, o sinal quimiotáxico pode movimentar grande quantidade de
glóbulos brancos do capilares até a área inflamada.
2.2.6. Fagocitose
A função mais importante dos neutrófilos e macrófagos é a fagocitose, que
significa ingestão celular do agente agressor.
Fagócitos devem ser seletivos em relação ao material que é fagocitado; ou, de
outro modo, algumas células e estruturas normais do corpo seriam ingeridas. Se a
fagocitose ocorrerá ou não, isso depende especialmente de três processos seletivos.
Primeiro, a maioria das estruturas naturais nos tecidos tem superfícies lisas
que resistem a fagocitose. Mas, se a superfície é áspera, a probabilidade de fagocitose
é aumentada.
Segundo, a maioria das substâncias do corpo tem coberturas protéicas defensivas
que repelem os fagócitos. Por outro lado, tecidos necrosados e a maioria das
partículas estranhas com frequência não possuem coberturas protetoras, o que se
torna sujeitas a fagocitose.
-----
~-~,--_.
11
Terceiro, o organismo tem um método específico de reconhecer certos
materiais estranhos. Essa é uma função do sistema imune, onde produzem anticorpos
contra agentes infecciosos como as bactérias. Os anticorpos aderem-se à membrana
bacteriana e desse modo tornam a bactéria especialmente suscetível a fagocitose.
Feito isto, a molécula de anticorpo também se combina com o produto C3 da cascata
complemento, que é um componente adicional do sistema imune. As moléculas C3
ligam-se a receptores na membrana do fagócito, iniciando, assim, a fagocitose. O
processo total é chamado de opsonização.
2.2.7. Sistema monócito-macrófago (sitema retículoendotelial)
Uma grande parte dos monócitos, ao penetrarem nos tecidos e após se
diferenciarem em macrófagos, tornam-se fixos e permanecem fixos nos tecidos
durante meses, ou mesmo anos, até serem convocados para desempenhar funções
protetoras específicas. Eles tem a mesma capacidade, como os macrófagos móveis,
de fagocitar grandes quantidades de bactérias, vírus, tecido necrosado ou outras
partículas estranhas no tecido. Quando apropriamente estimulados, podem romper
suas conexões, tornando-se outra vez macrófagos móveis que respondem à
quimiotaxia e a outros estímulos relativos ao processo inflamatório. Dessa maneira, o
organismo possui um "sistema monócito- macrófago" difuso virtualmente em todas as
áreas.
A reunião de monócitos,macrófagos móveis, macrófagos teciduais fixos e
algumas células endoteliais especializadas na medula óssea, baço e linfonodos é
também denominada de sistema retículoendotelial . Contudo, todas ou quase todas
estas células se originam de células
-
tronco monocíticas ; daí o sistema
reticuloendotelial é quase sinônimo do sistema reticuloendotelial. Embora o termo
sistema "reticuloendotelial" seja mais bem conhecido na literatura médica do que o
termo "sistema monócito macófago", o mesmo deve ser lembrado como um sistema
difuso de fagócitos localizado em todos os tecidos mas especialmente naquelas áreas
onde grandes quantidades de particulas, toxinas e outras substâncias indesejáveis
devem ser destruídas. (GUYTON & HALL, 1996).
12
2.2.8. Eosinófilos
Os eosinófilos normalmente constituem aproximadamente 2% de todos os
linfócitos. Os eosinófilos são fagócitos fracos, apresentam quimiotaxia, mas, em
comparação com os neutrófilos, é duvidoso que os eosinófilos sejam de importância
significativa na proteção contra os tipos comuns de infecção.
Por outro lado, os eosinófilos são frequentementes produzidos nas pessoas
com infecções parasitárias, migrando para os tecidos infectados pelos parasitas.
Embora a maior parte dos parasitas seja muito grande para ser fagocitada pelos
eosinófilos ou por qualquer outro fagócito, os eosinófilos aderem aos parasitas por
intermédio de uma molécula especial da superfície e liberam substâncias que matam
muitos deles. Por exemplo, uma das infecções mais comuns no mundo é a
esquistossomose, uma infecção parasitaria encontrada em um terço da população de
alguns países do terceiro mundo; o parasita literalmente invade qualquer parte do
corpo. Os eosinófilos aderem à forma jovem do parasita e destroem muitos deles.
Atuam de diversas maneiras: (1) liberando as enzimas hidrolíticas dos seus grânulos,
que são lisossomos modificados ; (2)provavelmente, liberando também formas de
oxigênio reativo que são especialmente letais ;(3) liberando dos grânulos um
polipeptídeo altamente tóxico para larva denominado de proteína básica principal. Nos
Estados Unidos, outra doença parasitária que causa eosinofilía é a triquinose, que
resulta da invasão dos músculos pelo parasita Trichinella (verme do porco) após uma
pessoa haver ingerido carne de porco não-cozida.
Os eosinófilos também têm tendência especial de acumular-se nos tecidos
onde ocorreram reações alégicas, tais como os tecidos ao redor dos brônquios nas
pessoas com asma, e na pele depois de reações alérgicas cutâneas. Isto é resultado
no mínimo, em parte, pelo fato que muitos mastócitos e basófilos participam nas
reações alérgicas ; estes mastócitos e basófilos liberam um fator quimiotáxico
eosinófilo que induz os eosinófilos a migrarem na direção do tecido inflamado.
Acredita-se que os eosinófilos detoxificam algumas substâncias indutoras da
inflamação liberadas pelos mastócitos e, provavelmente, também fagocitam e
destroem os complexos alérgeno-anticorpo, evitando desse modo a disseminação do
processo local inflamatório.
13
2.2.9. Basófilos
Os basófilos no sangue circulante são semelhantes aos grandes mastócitos
situados nas proximidades da maioria dos capilares do corpo. Os mastócitos situados
nas proximidades da maioria dos capilares do corpo. Os mastócitos e basófilos
liberam heparina no sangue, uma substância que impede a coagulação sanguínea.
assim como acelera a remoção de partículas de gordura do sangue após uma
refeição gordurosa.
Os mastócitos e basófilos também liberam histamina , assim como pequenas
quantidades de bradicinina e serotonina. Na verdade, são os mastócitos no tecidos
inflamados que, principalmente, liberam estas substâncias durante a inflamação.
Os mastócitos e basófilos desempenham um extraordinário e importante papel
em alguns tipos de reações alérgicas porque o tipo de anticorpo que causa reações
alérgicas, a IgE, tem uma propensão especial de aderir aos mastócitos e basófilos.
Assim, quando um antígeno específico reage com o anticorpo, a consequente ligação
de um com o outro induz a ruptura do mastócito ou basófilo e
liberação de grandes quantidades de histamina, bradicinina, serotonina, heparina,
substância de reação leta da anafilaxia e uma série de enzimas lisossômicas. Estas
por sua vez, produzem reações vasculares e teciduais que originam muitas das
manifestações alérgicas, se não a maioria delas.
--
14
2.3 IMUNIDADE
2.3.1 Imunidade natural
o corpo humano tem a capacidade de resistir a quase todos os tipos de de
organismo ou toxinas que tendem a danificar os tecidos e órgãos. Esta capacidade é
chamada de imunidade. Grande parte da imunidade é a imunidade adquirida, que se
desenvolve depois do primeiro ataque do organismo por uma doença bacteriana ou
por uma toxina, muitas vezes necessitando de semanas ou meses para se
desenvolver. Uma parte suplementar da imunidade origina-se de processos gerais, e
não de processos direcionados para organismo infeccioso específicos. Esta é
chamada de imunidade natural, que inclui o seguinte:
- Fagocitose de bactérias e outros invasores por glóbulos brancos e células do
sistema de macrófagos teciduais.
- Destruição de organismos presentes no estômago pelas secreções ácidas gástricas
e enzimas digestivas.
- Resistência da pele a invasão de organismos.
- Presença de determinados compostos químicas no sangue que aderem aos
organismos estranhos ou toxinas e os destroem. Alguns desses compostos são (1)
lisozima, um polissacarídeo mucolítico que ataca as bactérias e ocasiona sua
desagregação; (2) polipeptídeos básicos, que reagem e inativam certos tipos de
bactérias Gran - positivos ; (3 ) o complexo do complemento, que será descrito mais
adiante, um sistema de aproximadamente 20 proteínas que pode ser ativados de
diversos modos para destruir bactérias ; (4) linfócitos matadores naturais, que podem
reconhecer e destruir células estranhas, células tumorais e até mesmo algumas
células infectadas.
Esta imunidade natural toma o organismo humano resistente a doenças tais
como algumas paralisias de animais por infecções virais, cólera, suína, peste bovina
e cinomose - uma doença viral que mata grande percentagem de cães afetados. Por
outro lado, muitos animais inferiores são resistentes ou mesmo imune a inúmeras
doenças humanas, tais como poliomilite, caxumba, cólera humana, sarampo e sífilis,
que são destrutivas ou mesmo letais para os seres humanos. (GUYTON & HALL,
1996).
---
--
15
2.3.2 Imunidade Adquirida
Além da sua imunidade natural, o organismo humano tem a habilidade de
desenvolver imunidade extremamente poderosa e específicas contra agentes
invasores tais como bactérias, vírus, toxinas e, até mesmo, tecidos estranhos de
outros animais. Isto é denominado de imunidade adquirida. A imunidade adquirida é
induzida por sistema imune especial formador de anticorpos e linfócitos ativados que
atacam e destroem organismos específicos ou toxinas. É com esse mecanismo imune
e algumas das reações associadas - especialmente as alergias - que o restante deste
capítulo está relacionado. A imunidade adquirida pode muitas vezes
conferir proteção extrema. Por exemplo, certas toxinas,
tais como a toxina
paralisante do botulismo ou a toxina do tétano, podem ser bloqueadas em doses tão
altas quanto 100.000 vezes a quantidade que seria letal sem imunidade. Esta é a
razão por que o processo conhecido como" vacinação "é tão importante na proteção
dos seres humanos contra doenças e toxinas.
2.3.2.1 tipos básicos de imunidade adquirida
Dois tipos básicos mas intimamente relacionados de imunidade adquirida
ocorrem no organismo. Em um deles o organismo desenvolve anticorpos circulantes,
que são globulinas do sangue capazes de atacar agentes invasores. Este tipo de
imunidade é chamada de imunidade humoral ou imunidade célula- B ( porque os
linfócitos-B produzem os anticorpos). O Segundo tipo de imunidade adquirida é
alcançada através da formação de grandes quantidades de linfócitos ativados que são
designados especificamente para destruir o agente estranho. Este tipo de imunidade
é chamado de imunidade celular ou imunidade célula
-
T (porque os linfócitos
ativados são linfócitos - T).
Logo tanto os anticorpos como os linfócitos ativados são formados no tecido
linfóide do corpo.
16
2.3.2.2. os dois tipos de imunidade adquirida que são iniciados pelos antígenos
Desde de que a imunidade adquirida não ocorre até depois da primeira invasão
por organismo ou toxina estranha, está claro que o organismo deve ter algum
mecanismo para reconhecer a invasão inicial. Cada toxina ou tipo de organismo quase
sempre contém um ou mais componentes químicos específicos, na sua configuração,
que são diferentes de todos os outros compostos. Em geral, são proteínas ou grandes
polissacarídeos e são eles que iniciam a imunidade adquirida. Estas substâncias são
chamadas de antígenos.
Para uma substância ser antigênica, normalmente deve Ter um alto peso
mOlecular, 8.000 ou mais. Além disso, o processo de antigenicidade normalmente
depende da ocorrência periódica de grupos mOleculares, chamados de epítopos, na
superfície de grandes moléculas, o que explica porque as proteínas e grandes
polissacarídeos são quase sempre antigênicos devido ao fato de ambos possuírem
este tipo de características esterioquímica.
Haptenos. Embora substância com peso molecular menos que 8.000 raramente
atuem como antígenos, mesmo assim a imunidade pode se desenvolver contra
substâncias de baixo peso molecular de uma maneira especial, como se segue: caso
o composto de baixo peso mOlecular, chamado de hapteno, primeiro se combine com
uma substância antigênica, tal como uma proteína, então a combinação resultará
numa resposta imune. Os anticorpos ou linfócitos ativa dos que se desenvolvem bem
contra a associação então reagirão separadamente contra a proteína ou hapteno. Por
esta razão, numa segunda exposição do hapteno alguns anticorpos ou linfócitos
reagirão com ele antes que ele possa se disseminar pelo corpo e causar danos.
Os haptenos que provocam respostas imunes desse tipo são normalmente
drogas de baixo peso molecular, constituintes químicos da poeira, produtos da
decomposição química da descamação de animais, produtos degenerativos da
descamação da pele vários compostos químicos industriais a toxina do veneno da
urtiga, e assim por diante.
--
-- ----
17
2.3.2.3 os linfócitos como base da imundade adquirida
A imunidade adquirida é o produto do sistema linfático do organismo. As
pessoas que possuem carência genética de linfócitos ou aquelas cujos linfócitos foram
destruídos por irradiação ou drogas não podem desenvolver imunidade adquirida . E,
dias após o nascimento, essas pessoas morrem por infecção bacteriana fulminante,
salvo se tratadas com medidas heróicas.
Consequentemente, torna-se claro que os linfócitos são essenciais para a
sobrevivência do ser humano.
Os linfócitos estão localizados amplamente nos linfonodos, mas também são
encontrados em tecidos linfóides especias, tais como o baço, áreas da submucosa do
trato gastrintestinal e na medula óssea. O tecido linfóide esta distribuído
favoravelmente no organismo para interceptar os organismos invasores ou toxinas
antes que possam disseminar-se amplamente.
Em muitos casos, o agente invasor primeiramente penetra nos líquidos
teciduais e, então, é carreado via vasos linfáticos para o linfonodo ou outro tecido
linfóide. Por exemplo, o tecido linfóide do trato gastrintestinal é logo exposto aos
antígenos que invadem o intestino. O tecido linfóide da garganta e faringe (as tonsilas
e adenóides) esta bem localizado para interceptar os antígenos que penetram via trato
respiratório superior. O tecido linfóide nos linfonodos esta exposto aos antígenos que
invadem os tecidos periféricos do corpo. E, finalmente, o tecido linfóide do baço e
medula óssea desempenha o papel específico de interceptar agentes antigênicos que
conseguiram alcançar o sangue circulante.
Dois tipos de linfócitos promovem, respectivamente, a imunidade celular e a
imunidade humoral. Os linfócitos T e B.
Embora a maioria dos linfócitos no tecido linfóide normal pareçam
semelhantes quando estudados ao microscópios estas células são divididas em duas
grandes populações distintas. Uma das populações, os linfócitos T, é responsável
pela formação de linfócitos ativos que produzem a imunidade celular, e a outra
população, os linfócitos B, é responsável pela formação de anticorpos que
proporcionam a imunidade humoral.
Ambos os tipos de linfócitos são derivados originalmente no embrião, a partir
de células-tronco hemopoéticas pluripotenciais que diferenciam e originam os
----
18
linfócitos. Os linfócitos formados finalmente se localizam no tecido linfóide, mas
previamente sofrem diferenciação adicional ou são "pré-processados" da seguinte
maneira.
Os linfócitos destinados a formar linfócitos T ativados primeiro migram e são
"pré-processados" no timo, daí serem chamados de linfócitos 'T'. Eles são
responsáveis pela imunidade celular.
A outra população de linfócitos - os linfócitos 8, destinados a formar anticorpos
- são pré-processados no fígado, durante a metade da vida fetal, e na medula óssea,
durante o final da vida fetal e após o nascimento. Esta população celular foi
inicialmente descoberta nas aves, nas quais o pré-processamento ocorre na bolsa de
Fabrícius, uma estrutura ausente nos mamíferos. Por esta razão, estes linfócitos são
chamados de linfócitos "8", e são os responsáveis pela imunidade humoral.
2.3.2.4. pré-processamento dos linfócitos T e 8
Embora todos os linfócitos do corpo se originem das células-tronco linfocíticas
comprometidas estas células-tronco por si sós são incapazes de formar os linfócitos T
ativados ou os anticorpos. Previamente elas devem sofrer diferenciação adicional em
áreas apropriadas de processamento no timo ou em áreas de processamento de
célula 8.
O timo pré-processa os linfócitos T. Os linfócitos T, após sua produção na
medula óssea, inicialmente migram para o timo, onde se dividem rapidamente e, ao
mesmo tempo, desenvolvem extrema diversidade para reagir contra diferentes
antígenos específicos. Isto é, um dos linfócitos tímicos desenvolve reatividade
específica contra um antígeno. Depois, o linfócito seguinte desenvolve especificidade
contra outro antígeno. Isto continua até que haja diferentes linfócitos tímicos com
reatividades específicas contra, literalmente, milhões de antígenos diferentes. Estes
diferentes tipos de linfócitos T processados agora deixam o timo e disseminam-se
pelo organismo, para se fixarem no tecido linfóide.
O timo também determina que qualquer linfócito T que o deixa não reagirá
contra proteínas ou outros antígenos presentes nos tecidos do próprio corpo ; caso
--
19
contrário, os linfócitos T seriam letais para o corpo em somente poucos dias. O timo
seleciona quais os linfócitos T que serão liberados, expondo-os, virtualmente, a todos
os "antígenos próprios" específicos dos tecidos do corpo. Se um linfócito T reage, ele
é destruído e fagocitado, em vez de ser libertado, sendo o que acontece com 90% dos
linfócitos T. Deste modo, somente as células que finalmente são liberadas são
aquelas não reativas contra os antígenos do próprio corpo
-
ao contrário, somente
contra os antígenos de origem externa tais como uma bactéria , uma toxina ou,
mesmo, um tecido transplantado de outra pessoa.
A maior parte do pré-processamento dos linfócitos T no timo ocorre
rapidamente antes do nascimento de uma criança e durante alguns meses após o
nascimento. Após este período, a remoção do timo diminui (mas não elimina) o
sistema imune linfótico T. Entretanto, a remoção do timo vários meses antes do
nascimento evita o desenvolvimento de toda a imunidade celular. Como este tipo de
imunidade celular é o principal responsável pela rejeição de orgãos transplantados,
tais como coração e rim, podem-se transplantar orgãos com pouca probabilidade de
rejeição se timo for removido do animal num período razoável antes do nascimento.
O fígado e a medula óssea pré-processam os linfócitos B. Sabe-se muito
pouco sobre os detalhes do pré-processamento dos linfócitos B do que aqueles do
pré-processamento dos linfócitos T. Nos seres humanos, considera-se que os
linfócitos B são processados, no fígado, durante o segundo trimeste da vida fetal e, na
medula óssea, durante o final da vida fetal e após o nascimento.
Os linfócitos B são diferentes dos linfócitos T por dois motivos: primeiro, em vez
de a célula inteira desenvolver retividade contra o antígeno, como ocorre com os
linfócitos T, o linfócito B secreta ativamente anticorpos que são agentes reativos.
Estes agentes são grandes moléculas protéicas capazes de se combinar com o
agente antigênico e destruí-Io. Os linfócitos B têm maior diversidade do que os
linfócitos T, produzindo muitos e muitos milhões
-
talvez bilhões
-
de tipos de
anticorpos com diferentes reatividades específicas.
Após o pré-processamento, os linfócitos B, como os linfócitos T, migram para o
tecido linfóide através do organismo, onde se fixam nas proximidades das áreas de
linfócitos T. (GUYTON & HALL, 1996).