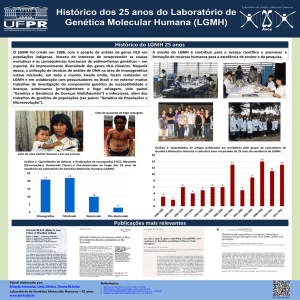
Deriva ∝ 1/2Ne
Por que então é também importante em grandes
populações?
Mutações recentes
Qual a prob de uma mutação que ocorreu pela 1a vez
passar para a próxima geração?
Modelo aleatório, Distribuição de Poisson!
• Mutações recentes
P(i’=0) = e-22i / i!, logo:
• Alelos neutros
Mutações recentes
Qual a prob de uma mutação que ocorreu pela 1a vez
passar para a próxima geração?
Alelos Neutros
São alelos funcionalmente equivalentes aos seus
alelos ancestrais na probabilidade de se replicar e
deixar descendentes na próxima geração
Embora tais alelos tenham sido discutidos por
Wright e Fisher, apenas após a década de 60 sua
importância foi reconhecida
Se k = 4, p = 0.14
Se k = 1, p = 0.61
Conseqüências do “Founder-flush” e de efeitos
fundadores
Genética de Populações
Genética de Populações
Controvérsias acerca da extensão da variação genética
• Estudo da geração e manutenção da variação genética
• Até a década de 1960 apenas conseguiu investigar isto
indiretamente pelo estudo do fenótipo
• Ausência de dados levou a grande controvérsia acerca
da extensão da variação genética
Escola Clássica - muito pouca
variação na natureza
Muller
Escola Balanceada - muita variação
mantida na natureza
Dobzhansky
Escola clássica - mutação é fator mais importante e por
isso mesmo raro.
Morgan e Muller
Mutação surge e é rapidamente selecionada, ou
aumenta de freqüência ou é eliminada da população.
Seleção natural mantém polimorfismo baixo nas
populações - variação era rara.
1
Genética de Populações
Controvérsias acerca da extensão da variação genética
Escola balanceada – Sturtevant e Dobzhanski
sugeriram que existia bastante variação na natureza.
Esta variabilidade seria mantida na população em
forma de polimorfismos balanceados, que fariam com
que ambos alelos permanecessem na população.
Da mesma forma que na escola clássica, seleção tem
papel fundamental na determinação da variação
genética encontrada.
Comparação de seqüências de aminoácidos
Seqüência 1
Seqüência 2
leu
arg
phe
cys
ser
ser
leu
phe
cys
ser
ser
arg
Seqüência 1
Seqüência 2
leu
arg
phe
cys
ser
ser
arg
leu
gap phe
cys
ser
ser
arg
Seqüência
Seqüência
Seqüência
Seqüência
Seqüência
Seqüência
leu
leu
leu
leu
leu
leu
arg
gap
gap
arg
arg
arg
cys
cys
cys
cys
cys
cys
ser ser
ser phe
ser phe
ser ser
ala ser
ile ser
arg
arg
arg
arg
arg
arg
1
2
3
4
5
6
phe
phe
phe
ile
ile
phe
arg
Medição de divergência de seqüência
Década de 60
Seqüenciamento de aminoácidos – permitiu a
comparação de genes homólogos em espécies
diferentes
Eletroforese de proteínas – permitiu a investigação
de polimorfismo em vários loci no genoma.
• Harris e Hopkinson (1966) em humanos
• Hubby e Lewontin (1966) em populações naturais
Medição de divergência de Seqüência
Várias medidas possíveis, mais simples é a comparação do
número proporcional de diferenças:
p = nd / n
nd = número de diferenças
n = número de aminoácidos
Distância é corrigida com Poisson para considerar substituições
múltiplas
Divergência pode ser usada para agrupar
Seqüências de aa de cadeias alfa de hemoglobina
No. de Taxa : 5
Gaps/Dados ausentes : Deleção completa
Método de distância : AA corrigida Poisson
No. de Sítios : 140
d : Estimativa
[1] Homem
[2] Camundongo
[3] Galinha
[5] Salamandra
[6] Carpa
2
16
3
35
39
4
62
63
63
5
68
68
72
74
Tubarão
79
79
83
84
??
2
Relógio Molecular
Década de 60
Seqüenciamento de aminoácidos – permitiu a
comparação de genes homólogos em espécies
diferentes
Eletroforese de proteínas – permitiu a investigação
de polimorfismo em vários loci no genoma.
• Harris e Hopkinson (1966) em humanos
• Hubby e Lewontin (1966) em populações naturais
Mudanças moleculares em genes e seus produtos
acumulam relativamente constantes no tempo
Genética de Populações
Controvérsias acerca da extensão da variação genética
Controvérsias acerca da extensão da variação genética
• Debate começou a ser resolvido na década de 60
quando pesquisadores começaram a investigar variação
natural por metologias moleculares - eletroforese,
seqüenciamento
Eletroforese em gel de amido
Década de 60
Seqüenciamento de aminoácidos - questionou a
quantidade de variação interespecífica
Eletroforese de proteínas - questionou a quantidade
de variação intraespecífica
Década de 60
Maioria das espécies têm grandes quantidades de
variação genética, e ainda era subestimada!
Não resolveu a controvérsia, pois a discussão mudou
da quantidade de variação para a significância da
variação genética.
Escola clássica tornou-se a escola da evolução
neutralista
3
Genética de Populações
Genética de Populações
Controvérsias acerca da extensão da variação genética
Controvérsias acerca da extensão da variação genética
A explicação de Motoo Kimura (Kimura 1968)
A explicação de Motoo Kimura (Kimura 1968)
• Variação era muito alta e as mudanças muito rápidas
para serem explicadas pela seleçao natural
Estimativa de Haldane indica 1 substituição a cada 300 gerações
- Estas substituições estavam acontecendo muito mais
rapidamente
1 - 1 subst a cada 28my por 100 aa
2 - Tamanho de genoma de mamífero 4 x 109 bp
3 - 100 aa = 300 pb e 20% de substituição sinônima portanto 1
aa subst ≈ 1.2 subst pb
• - baseado em seq de aa (hb e cyt c) Kimura calculou uma
média de 1 aa p/ 28 my em uma proteína de 100 aa
• - Preço muito alto considerando o conceito de Haldane de
carga genética e custo da seleção (1 a cada 300 gerações)
Genética de Populações
Teoria Neutralista de Evolução Molecular :
Controvérsias acerca da extensão da variação genética
A explicação de Motoo Kimura (Kimura 1968)
Estimativa de Haldane indica 1 substituição a cada 300 gerações
- Estas substituições estavam acontecendo muito mais
rapidamente
4 - Tempo que levaria para estas subst ocorrerem no genoma:
Motoo Kimura (1924-1994)
Publicações de 1968 sobre evolução
neutralista através da deriva
explicam tanto os altos níveis de
variação quanto o relógio molecular
(28 x 106) / (4 x 109/300) / 1.2 = 1.8 anos
- Muito superiores à taxa de subst de 1 a cada 300 gerações
Teoria Neutralista de Evolução Molecular :
Motoo Kimura (1924-1994)
Teoria Neutralista de evolução molecular
Teoria Neutralista não diz respeito apenas à deriva, e
sim à interação entre deriva e mutação.
2N genes em uma população.
Vários aspectos da evolução de
caracteres neutros não dependem do
tamanho populacional.
Taxa de mutação de alelos neutros μ
Ein?
Taxa de produção de novos alelos neutros = 2Nμ
Mas não é α 1/2N?
Taxa de evolução molecular = 1/2N x 2Nμ = μ
Pfixação = 1/(2N)
4
Teoria Neutralista de evolução molecular
Teoria Neutralista de evolução molecular
Taxa de evolução molecular = 1/2N x 2Nμ = μ
Taxa de evolução molecular = 1/2N x 2Nμ = μ
Taxa que mede probabilidade fixação de alelos
novos criados na população.
Explica a existência de um relógio molecular
Nesta equação não existe o efeito do tamanho
populacional na taxa de evolução neutra por
deriva
Deriva é uma força evolutiva importante para alelos
neutros em todas as populações, não apenas as
pequenas!
Modelo Fisher-Wright de deriva genética
Explica a quantidade de polimorfismos na natureza
Taxa de Evolução
Probabilidade de fixação de
uma nova mutação neutra: =
1/2N
2N = 18
requer uma média de 4N
gerações.
Tempo entre fixações
sucessivas = 1/μ gerações.
*N = Tamanho efetivo
populacional.
2N = 100
Qual??
Hartl & Clark (1997)
Principles of Population Genetics
Teoria Neutralista de evolução molecular
Hartl & Clark (1997)
Principles of Population Genetics
Teoria Neutralista de evolução molecular
Infinite Allele Model - Cada mutação produz um novo
alelo. Não existe identidade por estado, apenas ibd.
O impacto da deriva e da mutação pode ser quantificado:
Pnão mutação = (1 - μ)2
À medida que N diminui, F aumenta.
Quando μ aumenta, F diminui.
Deriva aumenta a probabilidade de ibd, enquanto
mutação a diminui
5
Teoria Neutralista de evolução molecular
Equilíbrio entre deriva e mutação na população:
F(t) = F(t-1)
Teoria Neutralista de evolução molecular
1- Feq = Heq = heterozigosidade esperada, logo:
H=
Arredondando (1- μ)2
(4 Nef μ)
=
(1 + 4 Nef μ)
(θ )
(1 + θ )
Chamemos θ = 4 Nefμ, que mede a força da mutação
em relação à deriva
Feq = homozigosidade esperada
Teoria Neutralista de evolução molecular
(θ )
H=
Críticas sobre a teoria neutralista
O que é μ e como medir?
E como medir Nef?
Na ausência de estimativas confiáveis, quaisquer
valores poderiam ser usados…
Grande poder explanatório, mas difícil de se testar!
(1 + θ )
E como separar neutralidade de seleção?
Carpa/Lampreia
V ertebrados/
Insetos
Mamíferos/
Répteis
Répteis/Peixes
MamÍferos
Aves/Répteis
Mudanças corrigidas de amino ácido por 100 resíduos
E como separar neutralidade de seleção?
i
j
μ não é constante em genes sob pressão de seleção
diferentes.
10
200
180
160
140
120
abcde
f
g
h
Fibrin
opept
ídeos
1.1 M
Y
220
6
5
na
bi
lo
og
m
Y
e
M
H
8
100
7
8
Substituições possíveis no DNA
purinas
9
G
A
Evolução das
globinas
transições
transversões
5.
80
60
20.0
Pliocene
Miocene
Oligocene
Eocene
Paleocene
100 200 300
400 500
Algonkian
Cambrian
Devonian
Silurian
Ordovician
Cretaceous
Jurassic
Triassic
Permian
0
Carboniferous
20
c
romo
Citoc MY
4
3
2
600
1
Separação dos ancestrais
De plantas e animais
T
C
transições
pirimidinas
Huronian
40
700 800 900 1000 1100 1200 1300 1400
Milhões de anos desde a divergência
6
Mutações sinônimas são mais comumente
fixadas na evolução
Limitações Funcionais
replacement
3
mtDNA Animal
4
Pseudogenes
5
Introns
6
Sítios 4x não-degenerados
Regiões à montante
7
1.000.000.000 anos
Substituições por sítio por
8
Regiões à jusante
9
Sítios não-degenerados
10
Sítios 2x não-degenerados
Diferentes tipos de seqüência evoluem
em taxas diferentes
8
6
4
2
0
3
In
su
lin
m
io
a
gl
ob
in
a
al
bu
in
m
te
i
ap
n
rle
a
ol
u
ip
op cina
ro
1
te
ín
aA
in
te
-1
rfe
ro
n
B1
re
la
xi
na
silent
Mutações sinônimas
Mutações não-sinônimas
10
hi
sto
na
Taxas diferentes entre
mutações silentes e nãosilenciosas (alteração da
seqüência de aa)
Substituições por sítio por
1.000.000.000 anos
12
E como separar neutralidade de seleção?
A evidência de taxa de evolução diferente em
proteínas diferentes apenas favorece a teoria
neutralista se for inconsistente com a idéia da
seleção natural determinando a evolução
molecular
Fisher afirmou que quanto menor o efeito de um
alelo, maior a chance dele ter um efeito benéfico
2
1
0
Teoria da evolução de Fisher
• Mutações aleatórias em um fenótipo qualquer podem
ou não ser favoráveis
E como separar neutralidade de seleção?
Taxa de evolução e relógio molecular entre espécies
diferentes são compatíveis com ambas as
hipóteses.
E dados de variação intraspecífica?
+3
+2
+1
De acordo com o neutralismo, qualquer nível de
variação pode ser explicado por níveis diferentes
de μ e Nef.
7
E como separar neutralidade de seleção?
Se μ é constante para certos genes, a diferença em
heterozigosidade esperada é causada por Nef
diferentes no neutralismo.
E como separar neutralidade de seleção?
Se μ é constante para certos genes, a diferença em
heterozigosidade esperada é causada por Nef
diferentes no neutralismo.
Todos os Heq < 0.25
Valores restritos de θ,
--> logo de Nef
E como separar neutralidade de seleção?
Grande parte da H está entre 0.01 e 0.10, indicando
apenas um aumento de 10 vezes no Nef, enquanto o
N variou em até 14 ordens de magnitude!
Nef ≠ N
mas mesmo assim...
Teoria quase neutra de evolução
E se olharmos para μ?
μ não é a taxa de mutação, e sim a taxa de mutação
NEUTRA - sem efeito na viabilidade
Ohta olhou para certos tipos de mutação:
Se s « 1/N a mutação é seletivamente neutra
Se s » 1/N a mutação é afetada por seleção
Mas e se s ≈ 1/N?
Teoria quase neutra de evolução
Fisher
Efeito da mutação
Prob. substituição adaptativa
Prob. Mutação favorável
Fisher ignorou a probabilidade de fixação
Mutações de efeito fenotípico intermediário são
mais prováveis de terem papel na adaptação
Teoria quase neutra
A taxa neutra de evolução é mais alta em populações
pequenas por incluir algumas mutações que seriam
desfavorecidas em populações maiores.
Quando Nef aumenta, μ diminui
Kimura
Nef μ fica com distribuição mais restrita o que pode
explicar os valores de H encontrados!
Efeito da mutação
8
Teoria quase neutra
As predições do relógio molecular vão pro espaço
μ é função agora de Nef e discrepâncias do relógio
molecular são esperadas
Porque relógio molecular e neutralidade absoluta não
explicam bem os dados observados, teoria neutra é
boa hipótese nula.
9