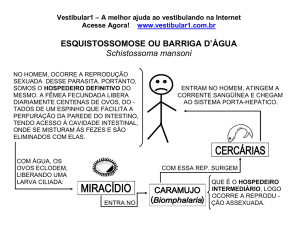
Anais da XI Jornada de Iniciação Científica da UFRRJ
Cinética da produção de antígeno viral em ovos embrionados a partir de
amostras patogênicas e apatogênicas do Vírus da Doença de Newcastle
Paula Amorim Schiavo1, Gabrielle Sales de Oliveira1, Cristiana Portz2, Jorge Granja de Oliveira
Júnior3, Bernardo Oliveira Loureiro4, Carlos Mazur5 & Cláudio de Moraes Andrade6
1. Graduanda em Medicina Veterinária, bolsista IC-PIBIC/UFRRJ. E-mail: [email protected]; 2. Médica Veterinária,
mestranda CPGMV-PV; 3. Médico Veterinário; 4. Graduando em Medicina Veterinária, estagiário LVV/DESP/IV/UFRRJ;
5. Prof. Adjunto DMIV/IV/UFRRJ; 6. Prof. Adjunto DESP/IV/UFRRJ.
Laboratório de Viroses Veterinárias, DESP/IV/UFRRJ, BR 465, Km 7, Seropédica, RJ. 23890-000 .
Palavras-chave: Newcastle, NDV, Viral Cultivation, Paramyxoviridae.
Abstract
rapidamente entre aves susceptíveis, em alguns
casos de maneira aparentemente misteriosa
(MURPHY, et al, 1999). Foi reportada pela primeira
vez em Java, em 1926, como doença
economicamente importante em galinhas domésticas
e perus (KRANVELD, 1926). A primeira descrição
da Doença de Newcastle no Brasil data de 1953,
quando CUNHA & SILVA realizaram o isolamento
da amostra M33 na cidade de Macapá-Amapá,
atribuindo o surto da doença à importação de
carcaças de frangos congelados provenientes dos
Estados Unidos (CUNHA et al, 1955). A partir desta
data, a doença se disseminou por todo o território
nacional, ocasionando graves perdas econômicas
para os avicultores brasileiros (HASTENREITER,
1976). Classificado na família Paramyxoviridae,
gênero Rubulavirus (LAMB et al, 1996), este e muitos
outros paramixovírus aviários se desenvolvem bem
em vários sistemas celulares, mas a técnica mais
sensível para o isolamento primário destes vírus é a
inoculação em ovos embrionados de galinha com 9
a 10 dias de incubação (ALEXANDER, 1989)
(LLENNETE, 1995). Os ovos embrionados de galinha
foram introduzidos na bateria de técnicas virológicas
em 1940 e rapidamente se tornaram um dos fatores
principais no progresso sucedido. Apresentavam
vantagens consideráveis sobre os sistemas animais
usados até então (coelhos, cobaios e camundongos):
é um meio vivo, altamente sensível a vários vírus; é
um sistema fechado, teoricamente estéril,
relativamente barato e pode ser obtido e manipulado
em grandes quantidades. Pode ser inoculado por
várias vias que correspondem às sensibilidades e
aos diferentes graus de adaptação de vários vírus
(MANGUEIRA et al, 1999). A via alantóica é um meio
simples de inoculação de ovos embrionados, sendo
utilizada na produção de grandes quantidades de
We had confronted the viral response of J99, M33
and La Sota strains of Newcastle Disease Virus in
one-hundred-eight 10-days-old SPF chicken embryos
obtained from immunized against Newcastle Disease
hens and non-vaccinated ones, analyzing the
haemagglutinating activity of allantoic fluids collected
with 24, 48, 72 and 96 hours post-inoculation. It was
searched if there was any interference of egg’s origin;
if any strain prevailed over another; if any HA title
prevailed over another when it was considered egg’s
origin. We observed a high HA title in first collects,
followed by a decrease and subsequent increase,
establishing HA highest titles on allantoic fluids
collected with 96 hours post-inoculation, except about
La Sota strain in immunized eggs, which higher title
was obtained in the first collect. J99 was the strain
that provides the greater haemagglutinating titles;
La Sota strain provides lower ones. We concluded
that the best time to collect allantoic fluids is 96 hours
post-inoculation. Chicken embryos obtained from
immunized against Newcastle Disease hens may
difficult the NDV primary isolation, but can be used
for antigen production.
Introdução
O Vírus da Doença de Newcastle (VDN), um dos
mais importantes patógenos de aves de várias
espécies, está mundialmente distribuído e produz
quadros clínicos que inclui a morte súbita em 100%
dos casos à doença subclínica, dependendo da cepa
infectante e do hospedeiro (ALEXANDER, 1989). A
Doença de Newcastle é uma das mais contagiosas
de todas as doenças virais, disseminando-se
165
v. 11, n. 2, p. 165-168, 2001
Anais da XI Jornada de Iniciação Científica da UFRRJ
Cinqüenta e quatro (54) ovos embrionados SPF
provenientes de galinhas hiperimunizadas contra a
Doença de Newcastle foram adquiridos junto à
Granja Guedes, Quatis-RJ. Convencionamos
chamar este grupo de ovos “vacinados” (VAC).
Inoculação em ovos embrionados: 100 mL de
diluições 1/10 em PBS das preparações virais foram
inoculadas pela via alantóica em ovos embrionados
de galinha com 10 dias de incubação, de acordo com
as técnicas de isolamento viral (LENNETE, 1955).
A inoculação dos ovos foi feita em baterias de 8 ovos
para cada cepa e dia, sendo 4 destes vacinados e 4
não-vacinados, e destes 2 foram inoculados com a
cepa correspondente, 1 com PBS na quantidade
equivalente e 1 não inoculado, estes 2 últimos como
controles do grupo. Os líquidos alantóicos de 12 ovos,
sendo 6 vacinados e 6 não-vacinados, foram
coletados no 1º dia e testados, como controles do
experimento. Os grupos inoculados foram, portanto,
divididos segundo a tabela 1.
vírus necessárias para o preparo de antígenos para
ensaios sorológicos, vacinas, cultivo de espécimes
clínicos, etc. Vírus inoculados na cavidade alantóica
se propagam nas células endodérmicas da
membrana alantóide com subseqüente liberação de
vírus no fluido alantóico, portanto tanto a membrana
quanto o fluido alantóicos podem ser usados como
fontes de vírus (LENNETE, 1995). ALEXANDER
(1989) preconizou o uso de ovos obtidos de uma
criação soronegativa para VDN, quando o uso de
ovos obtidos de uma criação livre de patógenos
específicos (SPF) não for possível, afirmando que o
uso de ovos de criações com anticorpos para VDN
reduziriam a capacidade do vírus se replicar e o
sucesso do isolamento viral (ALEXANDER, 1989).
No presente estudo, buscamos comparar a resposta
viral das cepas patogênicas M33 (IPIC=1,81)
(BELLUCI et al, 1999) e J99 (IPIC=1,42) (OLIVEIRA
et al, 2000) do VDN e da cepa apatogênica
(ALEXANDER, 1989) La Sota à inoculação em ovos
embrionados de galinhas hiperimunizadas contra a
Doença de Newcastle e em ovos provenientes de
criação SPF, analisando a atividade dos líquidos
alantóicos coletados com 24, 48, 72 e 96 horas pósinoculação. Pesquisamos a interferência da origem
do ovo na replicação viral; se havia predominância
de uma cepa sobre a outra; se algum título
hemaglutinante prevalecia sobre outro, levando em
consideração a origem do ovo.
Tabela 1. Esquema de inoculação e colheita por
cepa, origem do embrião e momento da colheita.
(horas pós-inoculação).
Coleta
(hs)
PBS
C. cego
La Sota
J99
M33
Total
Material e Métodos
Vírus: as suspensões do VDN, cepas La Sota, J99 e
M33, foram previamente cultivadas em ovos
embrionados de galinha. A amostra La Sota foi
adquirida no comércio, sob a forma de vacina
(Newvacin BioVet®). A amostra selvagem J99,
identificada e caracterizada pelo Laboratório de
Referência Animal do Ministério da Agricultura
(LARA-Campinas), foi isolada no Laboratório de
Viroses Veterinárias, DESP/IV/UFRRJ, a partir de
material fecal coletado em Japeri-RJ (OLIVEIRA et
al, 2000). A amostra selvagem M33 do VDN foi obtida
junto ao Instituto de Microbiologia Prof. Paulo de
Góes, da Universidade Federal do Rio de Janeiro
(UFRJ). Ovos embrionados de galinha: cinqüenta e
quatro (54) ovos embrionados SPF (Specific
Pathogens Free) de galinha foram adquiridos junto
à Granja Rezende, Uberaba-MG. Convencionamos
chamar este grupo de ovos “não-vacinados” (NON).
0
V
6
6
24
N
6
6
V
3
3
2
2
2
12
48
N
3
3
2
2
2
12
V
N
3
3
3
3
2
2
2
2
2
2
12 12
108
72
V
3
3
2
2
2
12
96
N
3
3
2
2
2
12
V
3
3
2
2
2
12
N
3
3
2
2
2
12
Após a inoculação os embriões foram incubados a
37ºC. O líquido alantóico foi coletado com 24, 48,
72, 96 horas pós-inoculação, após serem resfriados
a 4ºC por 30 minutos. O sobrenadante foi estocado
a -20ºC, após centrifugação por 10 minutos a 600 x
g. Prova de Hemaglutinação (HA): foi realizada em
placas de 96 cavidades e fundo em “U”. Diluições
geométricas do líquido alantóico foram feitas em
PBS, adicionadas do sistema revelador e incubadas
por 30 minutos em temperatura ambiente. O título
foi definido a partir da maior diluição de líquido
alantóico capaz de exercer atividade hemaglutinante
completa (BRASIL, 1994). Sistema revelador:
Suspensão de hemácias de galinha a 1% em PBS,
coletadas de aves doadoras derivadas de ovos SPF,
166
Anais da XI Jornada de Iniciação Científica da UFRRJ
mantidas no Laboratório de Viroses Veterinárias da
UFRRJ especificamente para este fim.
Os ovos provenientes de galinhas hiperimunizadas
contra a Doença de Newcastle ofereceram os
maiores títulos hemaglutinantes, quando analisadas
individualmente cada cepa por intervalo. O único
título hemaglutinante médio proporcionado por ovos
não-vacinados que prevaleceu sobre o de ovos
vacinados, quando pesquisados os maiores títulos
por intervalo, foi dado pela cepa M33 às 24 horas.
As cepas patogênicas ofereceram os maiores títulos
hemaglutinantes, exceto na primeira coleta. A cepa
apatogênica apresentou seu maior título
hemaglutinante médio, analisando-se todos os
resultados em conjunto, na primeira coleta, em ovos
vacinados. NAGAI et al. (1979) afirmaram que os
altos títulos proporcionados por cepas patogênicas
do VDN, em relação a cepas apatogênicas, quando
ambas eram inoculadas na cavidade alantóica de
ovos embrionados com 11 dias de incubação deviase ao fato de que as primeiras se replicavam em
várias
membranas
embrionárias
e
extraembrionárias, enquanto cepas apatogênicas se
replicavam apenas no endoderma, de acordo com a
susceptibilidade das glicoproteínas virais à clivagem
proteolítica (NAGAI, et al, 1979). Em todas as
amostras se observou um decréscimo dos títulos nas
primeiras coletas, seguido de aumento subseqüente,
com estabelecimento de alto título nas últimas
coletas, refletindo a recuperação dos títulos
encontrados ainda no inóculo, nos primeiros dias, e
o aumento segundo a replicação, como esperado,
nos dias seguintes.
Resultados e Discussão
Conforme esperado, nenhum dos líquidos alantóicos
controle deste experimento apresentou atividade
hemaglutinante, não interferindo, portanto, no
resultado final. Para a análise comparativa dos
demais líquidos alantóicos, calculamos o título
hemaglutinante médio de cada grupo, somando-se
os títulos hemaglutinantes de cada indivíduo do grupo
e dividindo pelo número de indivíduos. Estes estão
relacionados nas figuras 1 e 2.
O v o s v a c in a d o s
2 50 0
2 04 8
Títulos
2 00 0
1 50 0
1 28 0
1 02 4
7 68
5 12
1 00 0
5 00
6 40
3 84
2 56
35
26
0
2
1 02 4
0
24 hs
48 hs
72 hs
96 hs
C o l h e it a
L a S o ta
M 33
J 99
Figura 1. Cinética da produção de antígeno viral em
ovos provenientes de plantel imunizado.
Conclusões
O vos n ão-vac in ados
· Títulos decrescentes para a cepa La Sota em
ovos provenientes de galinhas hiperimunizadas com
esta cepa demonstram que o uso de tais ovos no
isolamento de amostras de campo pode interferir no
resultado.
2500
2048
Títulos
2000
1500
1040
1000
· O alto título encontrado na primeira coleta para
a amostra La Sota se deve à adaptação da cepa ao
embrião.
768
500
0
570
512
272
144
24 hs
256
96
48 hs
192
72 hs
· Todas as amostras apresentaram queda dos
títulos hemaglutinantes às 48 e 72 horas. Na
produção de antígeno viral, o melhor momento para
a coleta de líquidos alantóicos é às 96 horas pósinoculação de ovos embrionados com 10 dias de
incubação.
96 hs
C olh e ita
L a Sota
M 33
J 99
Figura 2. Cinética da produção de antígeno viral em
ovos SPF.
167
v. 11, n. 2, p. 165-168, 2001
Anais da XI Jornada de Iniciação Científica da UFRRJ
· Os títulos encontrados na última colheita são
compatíveis com as características de cada amostra
(vacinal ou selvagem), reforçando resultados
encontrados em outros experimentos (NAGAI et al,
1979).
KRANVELD, F.C. A Poultry disease in the Dutch East
Indies. Ned. Nd. Bl. Diergeneesk, 28: 448.450. 1926.
LAMB, R. & KOLAKOFSKY, D. Paramyxoviridae The
Viruses and their Replication. In: Fundamental
Virology. 3rd ed. Fields, B.N., Knipe, D.M., Howley,
P.M. et al., Lippincott-Raven Publishers, Philadelphia,
p. 577-604. 1996.
Agradecimentos e Auxílio Financeiro
Agradecemos a FAPERJ e ao PIBIC pelo suporte
financeiro.
LENNETE, E.H.; LENNETE, D.A.; LENNETE, E.T.:
Diagnostic Procedures for Viral, Rickettsial, and
Chlamydial Infections. 7th ed. American Public Health
Association, p. 22. 1995.
Referências Bibliográficas
MANUGUERRA, J.C.; HANNOUN, C. Grippe et
autres viroses respiratoires. Institut Pasteur. p.187.
1999.
ALEXANDER, D.J. Newcastle disease. In: A
Laboratory Manual for the Isolation and Identification
of Avian Pathogens. 3rd ed. The American Association
of Avian Pathologists. p. 114-120. 1989.
MURPHY, F.A.; GIBBS, E.P.J.; HORZINEK, M.C.;
STUDDERT, M.J. Veterinary Virology. 3 rd ed.
Academic Press. p. 419. 1999.
BELLUCI, M.S.P.; OLIVEIRA Jr., J.G.; PORTZ, C.;
ORSI, M.A.; DORETTO Jr., L.; MAZUR, C.;
ANDRADE, C.M. Identificação e caracterização da
amostra M33 do Vírus da Doença de Newcastle.
Resumos da IX Jornada de Iniciação Científica da
UFRRJ, p. 294-295. 1999.
NAGAI, Y.; SHIMOKATA, K.; YOSHIDA, T.;
HAMAGUCHI, M.; IINUMA, M.; MAENO, K.;
MATSUMOTO, T.; KLENK, H.D.; ROTT, R. The
spread of a pathogenic and an apathogenic strain of
Newcastle disease virus in the chick embryo as
depending on the protease sensitivity of the virus
glycoproteins. J. Gen. Virology, 45: 263-272. 1979.
BRASIL – MAARA. Programa Nacional de Sanidade
Avícola. Atos Legais, p. 83. 1994.
CUNHA, R.G. & SILVA, R.A. Isolamento e
identificação do vírus da Doença de Newcastle no
Brasil. Soc. Bras. Med. Vet. 23: 17-33, 1955.
OLIVEIRA JR, J.G.; PORTZ, C.; SCHIAVO, P.A.;
DORETTO JR, L.; ORSI, M.A.; MAZUR, C.;
ANDRADE, C.M. Isolation and biologic
characterization of the Newcastle disease virus in
Rio de Janeiro State. Virus Reviews and Research, 5
(2), suppl. 1: 56. 2000.
HASTENREITER, H. La Maladie de Newcastle au
Brésil. Bull. Off. Int. Epiz. 85(7-8): 813-817. 1976.
168







![Ppt0000079 [Somente leitura]](http://s1.studylibpt.com/store/data/003752968_1-1257c89e89e4e7c354469b531a0c1e3a-300x300.png)