Foto: Sérgio Fernando Larizzatti
DOMÍNIO ARCHAEA
REINOS CRENARCHAEOTA,
EURYARCHAEOTA E
KORARCHAEOTA
Arqueas
Número de espécies
No mundo: 108
No Brasil: ?
Conhecidas no estado de São Paulo: 4
Estimadas no estado de São Paulo: ?
Methanosphaera sp. *
s microrganismos do Domínio Archaea são considerados únicos, com
propriedades metabólicas extraordinárias e filogenia particular.
Compreendem os microrganismos anaeróbios mais sensíveis ao oxigênio,
os termófilos mais extremos e halófilos cujas enzimas são adaptadas a
elevadas concentrações salinas. Apesar da organização celular procariótica, são organismos
evolutivamente distintos das bactérias (Domínio Bacteria), em função de características de
organização do genoma, expressão gênica, composição celular e filogenia. As Archaea são
divididas em três Reinos: Crenarchaeota, Euryarchaeota e Korarchaeota. O conhecimento da
filogenia e taxonomia do Domínio Archaea é fundamental na compreensão da ecologia
microbiana de ecossistemas extremos. Os estudos taxonômicos ainda são raros, com um
notável direcionamento para as espécies anaeróbias produtoras do gás metano, de ampla
aplicação em tratamentos anaeróbios de resíduos e geração de biogás. Os dados existentes
sobre Archaea no estado de São Paulo restringem-se ao isolamento e identificação de
organismos metanogênicos presentes em sistemas de tratamento anaeróbio e sedimentos.
Não existem registros de isolados do Domínio Archaea caracterizados a nível de espécie de
linhagens em coleções de culturas brasileiras.
* Micrografia eletrônica de varredura de células procarióticas metanogênicas de um biodigestor anaeróbio.
DIVERSIDADE NO DOMÍNIO ARCHAEA
ROSANA FILOMENA VAZOLLER1,2, GILSON PAULO MANFIO1 &
VANDERLEI PEREZ CANHOS3
Coleção de Culturas Tropical (CCT), Fundação André Tosello,
Rua Latino Coelho, 1301, 13087-010 Campinas, SP, e-mail: [email protected]
2
Escola de Engenharia de São Carlos, Universidade de São Paulo
Av. Dr. Carlos Botelho, 1465, 13560-970 São Carlos, SP, e-mail: [email protected]
3
Universidade Estadual de Campinas, Faculdade de Engenharia de Alimentos (FEA)
Rua Professor Zeferino Vaz, s/nº, 13081-970 Campinas, SP, e-mail: [email protected]
1
2
1. Introdução
Os recentes estudos sobre filogenia, baseados na análise de seqüências do ácido ribonucléico ribossomal
(RNAr), revelaram que a vida na Terra é dominada pelos microorganismos. São três as divisões que definem a
organização filogenética atual dos seres vivos, os Domínios Bacteria, Archaea e Eukarya. Os dois primeiros
Domínios compreendem os organismos constituídos por células procarióticas e o último engloba aqueles formados
por células eucarióticas (Sly, 1998).
O Domínio Archaea é caracterizado por microrganismos procarióticos evolutivamente distintos dos
organismos alocados no Domínio Bacteria (Adams, 1995). Uma grande variedade de Archaea possui metabolismo
anaeróbio obrigatório, enquanto que outras espécies são encontradas em ambientes extremos, tais como fontes
geotermais, hábitats com elevada salinidade, solos e sistemas aquáticos altamente ácidos ou alcalinos. Pode-se
dizer que certas espécies de Archaea definem claramente os limites de tolerância biológica nos extremos físicos
e químicos da vida na Terra.
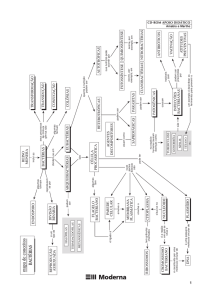
O Domínio Archaea consiste de três divisões, representadas na Figura 1: Crenarchaeota, que contém as
Archaea redutoras de enxofre hipertermófilas; Euryarchaeota, que compreende uma grande diversidade de
organismos, incluindo as espécies metanogênicas, as halófilas extremas e uma espécie hipertermófila; e
Korarchaeota, uma divisão descrita mais recentemente, que engloba organismos hipertermófilos pouco conhecidos,
ainda não cultivados em laboratório. A Figura 1 mostra a árvore filogenética de Archaea, baseada na comparação
de seqüências do RNAr 16S (Madigan et al., 1977; http://www.prenhall.com/~brock).
Várias espécies de Archaea são quimiorganotróficas, utilizando diferentes compostos orgânicos como
fonte energética para seu crescimento. É usual a ocorrência em Archaea de vias metabólicas semelhantes
àquelas encontradas em organismos do Domínio Bacteria. Como exemplo disto há o fato de que o catabolismo
da glicose em halófilas extremas e hipertermófilas se processa, basicamente, pela via de Entner-Doudoroff.
Entretanto, alguns organismos apresentam vias bioquímicas raras, como as células metanogênicas, que, através
de sistemas enzimáticos únicos, produzem o gás metano. O metabolismo autotrófico é bastante usual entre as
metanogênicas e em muitos hipertermófilos (Sowers, 1995; Madigan et al., 1997).
Os organismos halófilos extremos habitam locais hipersalinos, geralmente em regiões quentes e secas que
estimulam a evaporação, com conseqüente aumento na concentração de sais na água, sedimentos e solo. Diversos
hábitats hipersalinos têm servido como fonte para estudos de organismos que se desenvolvem em altas concentrações
de íon sódio, da ordem de 105-142 g/L (água do mar = 10.6 g/L), como o Great Salt Lake (Utah, EUA), lagos de
soda (África e Austrália) e o Mar Morto (este último com 40 g/L de íon sódio e 44 g/L de magnésio).
As Archaea metanogênicas, por sua vez, são amplamente distribuídas na natureza em ambientes anóxicos,
cujos potenciais de oxi-redução são da ordem de 300 mV, tais como: sedimentos aquáticos profundos, pântanos,
trato digestivo de ruminantes e animais homeotérmicos, biodigestores anaeróbios de tratamento de resíduos e
aterros sanitários.
18
R.F. Vazoller, G.P. Manfio & V.P. Canhos
A diversidade de Archaea hipertermófilas pode ser observada em ambientes geotérmicos: gêisers, fendas
de vulcões com emissão de vapores (em geral sulforados) e fendas no leito oceânico. Os hábitats mais comumente
estudados localizam-se em países com focos de atividade geotérmica, como os Estados Unidos (notadamente,
no Yellowstone National Park; Combie & Runnion, 1996), Islândia, Itália e Nova Zelândia (Oremland, 1988;
Vazoller, 1989; Madigan et al., 1997).
Em 1995, Carl R. Woese apresentou suas impressões sobre o surgimento do novo Domínio Archaea, em
final de 1977. O autor registrou que o aparecimento repentino das Archaea (inicialmente denominadas
Archaebacteria, Archeobacteria ou Archeote) - negando dogmas da Biologia, como a divisão dos seres vivos em
eucariontes e procariontes, de acordo com a presença de núcleo organizado e maquinaria genética - ocasionou
uma revolução na classificação dos organismos, pois surgia uma terceira forma de vida, cuja estrutura celular
é procariótica, mas, em nível molecular e genético, guarda grandes semelhanças com organismos eucarióticos. A
noção de dicotomia da vida entre eucariontes e procariontes, que ainda domina a Biologia e influencia, em
particular, a percepção sobre o grupo Archaea, está sendo lentamente revista por grupos atuantes em microbiologia.
A diversidade e biologia das Archaea representam uma enorme contribuição à compreensão da Ecologia Microbiana
(Woese, 1995).
Os membros do Reino Crenarchaeota apresentam a ramificação mais profunda na árvore fiologenética
universal, representados por organismos dos gêneros Desulfurococcus, Pyrodictium, Sulfolobus, Thermococcus, Thermofilum
e Thermoproteus. Em seguida, observam-se os grupos de Euryarchaeota, extremamente diversos, incluindo
Archaeoglobus spp., os halófilos extermos Halobacterium spp. e Halococcus spp., os metanogênicos Methanobacterium
spp., Methanococcus spp., Methanosarcina spp., Methanospirillum spp. e Methanothermus, além das metanogênicas halófilas.
Os organismos termo-acidófilos do gênero Thermoplasma, caracterizados pela ausência de parede celular,
representam um grupo distinto associado aos Euryarchaeota. Methanopyrus sp., um metanogênico hipertermófilo,
capaz de se desenvolver a 110 °C, se encontra bastante próximo ao ponto de radiação de Crenarchaeota e
Euryarchaeota. O Reino Korarchaeota, descrito mais recentemente, engloba organismos hipertermófilos pouco
conhecidos, identificados a partir de seqüências de rDNA 16S isoladas de fontes termais terrestres, porém ainda
não-cultivados em laboratório.
Na prática, as Archaea são estudadas e agrupadas de acordo com o seu metabolismo e fisiologia, sendo
alocadas em diversos grupos, como: as metanogênicas, os halófilos extremos (ou hiperhalófilos) e os termófilos
extremos (ou hipertermófilos). Sem dúvida, as espécies de Archaea mais conhecidas são as metanogênicas, cuja
classificação taxonômica molecular data de 1979, realizada por Balch e colaboradores (Balch et al., 1979).
Provavelmente, pela facilidade de estudos de ambientes anóxicos, ou mesmo pela sua importância em saneamento
ambiental, existem cerca de 66 espécies de metanogênicas descritas, em contraste com aproximadamente 24
espécies de hipetermófilas e 18 de halófilas extremas (Sowers, 1995; Adams, 1995; Madigan et al., 1997). Contudo,
o número de novas espécies descritas nestes dois últimos grupos vem aumentando, em razão do crescente
interesse em estudos da diversidade de Archaea em novos ambientes, que empregam metodologias moleculares
para a identificação dos organismos através de DNA extraído diretamente das amostras.
Os organismos do Domínio Archaea apresentam uma ampla diversidade metabólica e distribuição em
hábitats considerados inóspitos aos demais seres vivos. Pouco se sabe, porém, sobre sua importância funcional
nestes ecossistemas e sobre a atuação dos mesmos nos processos biogeoquímicos em ambientes não extremos.
Além disso, em termos tecnológicos, as descobertas de novos produtos microbianos gerados em ambientes
extremos (e.g., enzimas extremófilas) realça o potencial de exploração deste grupo de organismos como fonte de
novas alternativas para a Biotecnologia.
O emprego dos organismos metanogênicos em saneamento ambiental é amplamente conhecido e busca a
mineralização anaeróbia da matéria orgânica. Porém, as perspectivas se ampliam de forma marcante, por exemplo,
na bioprospecção de enzimas resistentes a condições extremas (e.g., temperaturas da ordem de 80-100°C) presentes
em Pyrococcus spp., Thermococcus spp., Thermofilum spp. e Thermoproteus spp. (Vazoller, 1995).
Em pesquisa básica e Biotecnologia, podem ser relacionadas outras aplicações de Archaea, tais como: a
manipulação genética de moléculas termoestáveis e a produção de compostos químicos e combustíveis (biogás
e álcool). Considerando-se que os bioprocessos de Archaea ocorrem sob temperaturas, salinidade e/ou pH
Archaea
19
marcadamente superiores aos dos organismos comumente empregados em biotecnologia, estes seriam extremamente
atraentes para o desenvolvimento de novas tecnologias. Uma gama diversa de substâncias de interesse industrial
pode ser produzida pelo metabolismo termófilo extremo: ácidos orgânicos, vitaminas, aminoácidos, antibióticos
e produtos bioativos (e.g., estimuladores de crescimento de tecido celular). Algumas Archaea, como Sulfolobus
spp., podem processar reações de biotransformação estereo-específicas em moléculas de hidrocarbonetos
(Vazoller,1995).
No estado de São Paulo, registra-se algum conhecimento sobre Archaea metanogênicas. As pesquisas
originais, voltadas principalmente para aplicações tecnológicas, estão descritas em relatórios internos da Companhia
de Tecnologia de Saneamento Ambiental do Estado de São Paulo (CETESB), em sua maioria coordenados por
Gregori (1979-1980) e Novaes (1982-1988), e em dissertações e teses de Mestrado e Doutorado do Departamento
de Hidráulica e Saneamento da Escola de Engenharia de São Carlos (USP). Publicações técnicas sobre operação
de biodigestores anaeróbios reforçam o interesse da indústria pelo tema (Souza et al., 1991). Os trabalhos sobre
Archaea metanogênicas relacionados no estado de São Paulo são descritos por Badra (1993), Vazoller (1989,
1995, 1997) e Vazoller et al. (1988).
Figura 1. Árvore filogenética do Domínio Archaea, baseada na comparação de seqüências do RNAr 16S
(baseada em Madigan et al., 1997 http://www.prenhall.com/~brock).
Figura 1. Árvore filogenética do Domínio Archaea, baseada na análise de seqüências do RNAr 16S (baseada em Madigan
et al., 1997; http://www.prenhall.com/~brock).
20
R.F. Vazoller, G.P. Manfio & V.P. Canhos
2. Diversidade
Archaea Halófilas Extremas:
É um grupo de organismos que habita ambientes altamente salinos, como: lagoas com elevada concentração
salina (e.g., Ca, Mg, Na) ou mesmo a superfície de alimentos preservados por salmora. Os hábitats das Archaea
halófilas extremas são denominados hipersalinos e as espécies em cultivo laboratorial podem requerer entre 1,5
e 4 M de NaCl para crescimento. A concentração elevada de sódio é essencial para a manutenção da estabilidade
da parede celular em alguns organismos, como em Halobacterium spp. (Madigan et al., 1997), por exemplo.
Dentre os ambientes naturais hipersalinos mais estudados encontram-se os sistemas lacustres com salinidade
ao redor de 10 vez a da água do mar, com predomínio de sais de sódio e presença de sulfatos e pH ligeiramente
alcalino. As espécies halófilas extremas são, em geral, quimiorganotróficas aeróbias obrigatórias, com raros
representantes halófilos anaeróbios. Podem ainda apresentar a propriedade de síntese de energia mediada pela
luz, na ausência de bacterioclorofilas, através da ação de bacteriorodopsinas (Madigan et al., 1997).
As espécies de Archaea hiperhalófilas são classificadas filogeneticamente em 8 gêneros e 18 espécies,
através da seqüência do RNAr 16S e de características morfo-fisiológicas (composição de parede celular, presença
de flagelos, coloração de Gram, tolerância à salinidade, plasmídeos e conteúdo de G+C do DNA). São descritos
os gêneros: Haloarcula, Halobaculum, Halobacterium, Halococcus, Haloferax, Halorubrum, Natronobacterium e Natronococcus
(Staley et al., 1989; Madigan et al., 1997). Os organismos apresentam, na sua maioria, morfologia celular em forma
de cocos e bacilos. Os ambientes mais estudados localizam-se em zonas hipersalinas de regiões temperadas,
como os lagos salgados em Utah e na Califórnia (EUA), bem como o Mar Morto.
Archaea Metanogênicas:
As Archaea metanogênicas são microorganismos anaeróbios obrigatórios, que requerem condições anóxicas
de crescimento, e altamente redutoras, com potenciais de oxi-redução na ordem de 300 mV (Sowers, 1995).
Provavelmente, a característica mais evidente das metanogênicas está relacionada com sua especificidade
de substratos para crescimento e produção de metano. São conhecidos até o momento dez substratos para a
metanogênese: formiato, monóxido de carbono, metanol, 2-propanol, aminas metiladas, dimetilsulfeto,
metilmercaptanas e acetato, sendo universal o dióxido de carbono, que necessita de hidrogênio como doador de
elétrons.
Os metanogênicos apresentam metabolismo quimiorganotrófico ou autotrófico. Apesar de requererem
condições fastidiosas para crescimento e anaerobiose obrigatória, esses organismos são amplamente distribuídos
na natureza, sendo encontrados em diversos ambientes associados à decomposição de matéria orgânica e/ou
atividades geoquímicas. As arqueas metanogênicas atuam no passo final de consórcios microbianos presentes
em sedimentos aquáticos, pântanos, gêisers, interior de árvores e sistemas de tratamento de resíduos, como
biodigestores anaeróbios e aterros sanitários. Sob o ponto de vista ecológico, o metabolismo metanogênico é
dependente da presença de outros microorganismos, cuja atividade no meio anaeróbio gera os precursores para
a metanogênese (Vazoller, 1995).
As Archaea metanogênicas representam um grupo de microrganismos polifilético, compreendendo 3 Ordens,
com 8 Famílias e 21 gêneros. Apresentam morfologia comum às células procarióticas, com forma de bacilos de
diferentes tamanhos, cocos, sarcinas e filamentos. Algumas representantes apresentam propriedade de coloração
Gram-positiva e outros Gram-negativa, sendo a taxonomia baseada essencialmente em métodos moleculares,
através da comparação de seqüências do RNAr 16S. Análises morfo-fisiológicas facilitam a classificação primária
em nível de gênero.
A Ordem Methanobacteriales compreende a Família Methanobacteriaceae, com os gêneros Methanobacterium,
Methanobrevibacter e Methanosphaera, contendo 18 espécies; e a Família Methanothermaceae, com o gênero Methanothermus
e duas espécies. A Ordem Methanococcales congrega as Famílias Methanococcaceae, gênero Methanococcus, com
7 espécies; Methanomicrobiaceae, com os gêneros Methanoculleus, Methanogenium, Methanolacinia, Methanomicrobium
e Methanospirillum, com 13 espécies; Methanocorpusculaceae, gênero Methanocorpusculum, com 5 espécies;
Methanoplanaceae, gênero Methanoplanus, com duas espécies; Methanosarcinaceae, com os gêneros Methanococcoides,
Archaea
21
Methanohalobium, Methanohalophilus, Methanolobus, Methanopyrus, Methanosaeta, Methanosarcina; e Methanothrix, com
19 espécies (Sowers, 1995).
A bioquímica da formação de metano é encontrada apenas nas Archaea metanogênicas. A metanogênese é
uma reação que proporciona um fluxo de prótons nas células e pode suportar um mecanismo quimiosmótico de
produção de energia e ATP. A produção de metano pode, portanto, ser encarada como um marcador taxonômico
para a identificação de Archaea. Alguns dos microrganismos apresentam fluorescência sob iluminação ultravioleta
decorrente da presença do cofator F420 (Oremland, 1988; Vazoller, 1995).
Archaea Hipertermófilas:
A divisão Crenarchaeota compreende microrganismos capazes de crescer em temperaturas extremas (e.g.,
acima do ponto de ebulição da água, com ótimo acima de 80 ºC), conhecidos como hipertermófilos. Estes
organismos foram inicialmente observados em amostras de solos e águas geotermais contendo enxofre elementar
e sulfetos. Posteriormente, os primeiros microrganismos hipertermofílicos foram isolados de comunidades
microbianas em fendas termais vulcânicas no leito oceânico, sob temperaturas de 100 °C. Essas descobertas
culminaram com a descrição do Domínio Archaea (Adams, 1995).
Os hipertermófilos são, em sua maioria, anaeróbios obrigatórios, quimiorganotróficos ou quimiolitotróficos,
que, de alguma forma, utilizam compostos de enxofre no seu metabolismo. O estudo de uma variedade de
hábitats terrestres e aquáticos resultou no isolamento de diversas espécies hipertermófilas, com morfologia
peculiar, em geral esféricas e irregulares, ou mesmo organismos com ausência de parede celular (Thermoplasma
acidophilum). A taxonomia das espécies é baseada em métodos moleculares (análise de seqüência do RNAr 16S),
fisiologia (crescimento sob diferentes temperaturas e pH), morfologia celular, coloração de Gram e conteúdo de
G+C do DNA.
A classificação dos hipertermófilos compreende organismos distribuídos em pelo menos 4 Ordens. A
Ordem Thermoproteales engloba a Família Thermoproteaceae, com os gêneros Pyrobaculum, Thermofilum e
Thermoproteus, isolados de áreas geotérmicas continentais, e a Família Desulfurococcaceae, com os gêneros
Desulforococcus, Pyrodictium, Staphylothermus e Thermodiscus, isolados de regiões marinhas geotérmicas profundas e
não profundas. A Ordem Thermococcales, com os gêneros Pyrococcus e Thermococcus, compreende organismos
isolados de ambientes marinhos profundos e não profundos. A Ordem Sulfolobales, com os gêneros Acidianus,
Desulfurolobus, Metallosphaera, Stygiolobus e Sulfolobus, apresenta organismos isolados de ambiente marinhos não
profundos e águas continentais.
Organismos do gênero Thermoplasma, Ordem Thermoplasmatales, isolados de áreas geotérmicas continentais,
apesar de apresentarem temperatura máxima de crescimento ao redor de 67 ºC, são considerados, para estudo,
junto ao grupo dos hipertermófilos (Adams, 1995). Existem ainda alguns organismos não classificados, isolados
de ambiente marinho não profundo e profundo, incluindo-se neste grupo os organismos do gênero Hyperthermus
(Adams, 1995).
As Archaea hipertermófilas redutoras do íon sulfato são alocadas no gênero Archaeoglobus e as Archaea
hipertermófilas metanogênicas, nos gêneros Methanococcus, Methanopyrus, Methanothermus (Adams, 1995). Assim
como os demais gêneros de hipertermófilos, estes organismos também são isolados de ambientes marinhos
geotermais.
Os organismos dos gêneros Thermotaga e Aquifex, filogeneticamente alocados no Domínio Bacteria, são, em
geral, estudados em conjunto com as Archaea devido ao seu hábitat marinho hipertermófilo.
3. Estado do conhecimento
A existência de um terceiro Domínio de seres vivos, Archaea, suscitou indagações da comunidade científica
sobre a sua evolução, biologia e papéis ecológicos. Muito embora este grupo de microrganismos apresente uma
grande diversidade metabólica, um ponto comum é indiscutível, a sua ocorrência em hábitats com extremos de
temperatura e pH, salinidade elevada e anaerobiose estrita. As condições iniciais de formação da Terra certamente
possibilitaram o surgimento das Archaea, representadas, por exemplo, pelos hipertermófilos e metanogênicos
22
R.F. Vazoller, G.P. Manfio & V.P. Canhos
utilizadores de hidrogênio. Contudo, é certo que a história da evolução dos seres vivos no planeta está relacionada
ao Domínio Archaea, fato evidenciado pelo parentesco filogenético entre Archaea e Eukarya (Adam, 1995;
Sowers, 1995; Woese, 1995).
Relatos sobre Archaea na literatura são concentrados em estudos conduzidos em regiões temperadas do
globo, sendo ainda incipientes os estudos em regiões tropicais. Na América Latina, países como Brasil, Colômbia,
México e Uruguai destacam-se no conhecimento sobre as Archaea metanogênicas em biorreatores anaeróbios
(diversos trabalhos citados no Anais dos Simpósios Internacionais de Digestão Anaeróbia, 1988 e 1991).
Em especial, no Brasil, foram desenvolvidos dois programas de formação de recursos humanos para
estudo da biologia de microorganismos metanogênicos. Em 1982, um projeto conduzido pela Sociedade Brasileira
de Microbiologia na Universidade de São Paulo promoveu o treinamento de cerca 50 profissionais sobre
microorganismos presentes em processos biológicos anaeróbios, destacando-se procedimentos práticos para o
isolamento de organismos anaeróbios estritos, como Archaea metanogênicas. Em 1987, o projeto Formação de
Recursos Humanos em Microbiologia da Digestão Anaeróbia, financiado pela FINEP e CNPq [Projeto CNPq
701651/87.3-SBIO/FV (CG), 19871991] e conduzido na CETESB, envolveu essencialmente treinamento de
pessoal em técnicas laboratoriais para isolamento de microorganismos anaeróbios estritos.
O grupo de pesquisadores da CETESB (Companhia de Tecnologia de Saneamento Ambiental do Estado
de São Paulo), no período de 1978 a 1990, trabalhou efetivamente na pesquisa de metanogênicos, sendo uma das
instituições pioneiras no isolamento de culturas, algumas delas preservadas por métodos convencionais de
subcultivo e estoque à temperatura ambiente ou sob nitrogênio líquido (Vazoller & Rech, 1992). Na coleção de
culturas da CETESB, ainda no ano de 1990, encontravam-se estocadas a temperatura ambiente ou sob nitrogênio
líquido culturas de Arcahea metanogênicas originárias de amostras de biodigestores de lodo de esgoto, isolados
pioneiramente pela Bióloga Dra. Rosana Elda Gregori (relatórios internos da CETESB 1979-1980). As culturas
estocadas foram identificadas como pertencentes ao gênero Methanobacterium (Vazoller, 1989) e listadas no Catálogo
Nacional de Culturas (Canhos et al., 1989). Estudos posteriores (Vazoller, 1988) revelaram o isolamento de
metanogênicas do gêneros Methanosaeta (sin. Methanothrix) e Methanosarcina em sistemas de tratamento anaeróbio
de esgotos sanitários.
Em 1992, o Departamento de Hidráulica e Saneamento da Escola de Engenharia de São Carlos da
Universidade de São Paulo obteve seus primeiros resultados no estudo de metanogênicas em aterros sanitários
(Badra, 1993), destacando-se neste sistema organismos dos gêneros Methanobacterium e Methanococcus. Em 1995,
estudos adicionais sobre a metanogênese termofílica da vinhaça resultaram no isolamento de duas culturas de
metanogênicos termófilos, identificados como Methanobacterium sp. e Methanosarcina sp. (Vazoller, 1995), através
do emprego de métodos clássicos de caracterização de Archaea, tais como: morfologia, fluorescência sob luz
UV, produção de metano e avaliação do consumo e utilização de substratos orgânicos e inorgânicos. Nenhum
método de preservação de culturas metanogênicas foi indicado nos trabalhos realizados por Badra (1993) e
Vazoller (1989; 1995).
Recentemente, técnicas moleculares estão sendo aplicadas no estudo de organismos metanogênicos no
Laboratório de Processos Anaeróbios, Departamento de Hidráulica e Saneamento da Escola de Engenharia de
São Carlos da Universidade de São Paulo, em conjunto com a Coleção de Culturas Tropical, da Fundação André
Tosello, Campinas. Esse trabalho constitui-se em uma iniciativa pioneira para a introdução de metodologias
moleculares para o estudo de Archaea em reatores anaeróbios, empregando técnicas de amplificação de rDNA
16S a partir de DNA isolado diretamente de amostras do reator, identificação dos organismos através de
seqüenciamento e análise filogenética do rDNA 16S, estudos de diversidade (DGGE de região espaçadora
rDNA 16S-23S) e detecção de organismos específico por hibridização in-situ com sondas fluorescentes.
Não há registros no estado de São Paulo de resultados de pesquisa sobre ocorrência e diversidade de
Archaea em ambiente tropical/sub-tropical. No entanto, existem vários hábitats extremamente atraentes para o
desenvolvimento de tais estudos, destacando-se regiões estuarinas de mangue (Baixada Santista e Cananéia),
regiões costeiras do estado, sedimentos aquáticos de lagos e rios do cerrado paulista, fontes termais (Araçatuba)
e sistemas anaeróbios de tratamento de rejeitos diversos (esgotos sanitários, vinhaça de processo de produção de
etanol e efluentes de indústria de papel e celulose).
Archaea
23
4. Prioridades para o desenvolvimento dos estudos
Percebe-se claramente que o Domínio Archaea é pouco conhecido e explorado em todos os ramos da
Microbiologia. No que se refere à sistemática e taxonomia, não há nenhum esforço acadêmico concentrado,
mesmo que individual, na direção de aprofundar o conhecimento sistemático deste grupo de organismos no
estado de São Paulo. Algumas iniciativas atuais ainda são incipientes e necessitam de apoio de agências de
fomento e desenvolvimento científico para a sua consolidação. É necessário um esforço direcionado à formação
de profissionais em microbiologia anaeróbia, isolamento (solos, sedimentos, água, sistemas de biorreatores etc.) e
taxonomia polifásica de Archaea. Os cursos de Pós-Graduação em Microbiologia devem introduzir e estimular o
conhecimento sobre a microbiologia de Archaea, reconhecida internacionalmente como prioritária para
desenvolvimento, particularmente em ecologia microbiana e bioprospecção para biotecnologia.
5. Bibliografia
Anais do V International Anaerobic Digestion Symposium. 1988. Bologna, Itália.
Anais do VI International Anaerobic Digestion Symposium. 1991. São Paulo, SP, Brazil.
Adams, M.W.W. 1995. Thermophilic Archaea: an overview. In: Archaea a laboratory manual: Thermophiles.
Robb, F.T. & Place, A.R. (eds.). New York: Cold Spring Harbor Laboratory Press. p. 3-7.
Badra, R.J. 1993. Isolamento e caracterização de bactérias metanogênicas de um aterro experimental. Dissertação
de Mestrado. São Carlos: Escola de Engenharia de São Carlos/USP.
Balch, W.E.; Fox, G.E.; Magrum, J.L.; Woese, C.R. & Wolfe, R.S. 1979. Methanogens: reevaluation of a
unique biological group. Microbiological Reviews, 43:260-296.
Canhos, V.P.; Souza, S. & Canhos, D.A.L. (eds.). 1989. Catálogo nacional de linhagens, vol. I - Bactérias.
Campinas: Fundação Tropical de Pesquisas e Tecnologia André Tosello.
CETESB Relatórios internos 1979 a 1980.
Combie, J. & Runnion, K. 1996. Looking for diversity of Yellowstone extremophiles. Journal of Industrial
Microbiology, 17:214-218.
a
Madigan, M.T.; Martinko, J.M. & Parker, J. 1997. Brock biology of microorganisms. 8 Ed. Caps. 15, 16 e
17. New Jersey: Prentice-Hall. p. 606-768.
Sly, L.I. 1997. Australian Microbial Diversity Is its preservation in good hands? In: Proceedings of the Annual
Scientific Meeting of the Australian Society for Microbiology. Adelaide: Australian Collection of
Microorganisms.
Souza, M.E.; Fuzaro, G. & Polegato, A.R. 1991. Thermophilic anaerobic digestion of vinasse in pilot plant
UASB reactor. In: Proceedings of Sixth International Symposium on Anaerobic Digestion, São Paulo,
Brazil. p. 191-200.
Sowers, K.R. 1995. Methanogenic Archaea: an overview. In: Archaea a laboratory manual. Methanogens.
Sowers, K. R. & Schreier, H. J. (eds). New York: Cold Spring Harbor Laboratory Press. p. 3-13.
Staley, J.T.; Bryant, M.; Pfennig, N. & Holt, J.G. (eds.). 1989. Bergeys manual of systematic bacteriology.
Archaeobacteria. Vol. 3. Baltimore: Williams & Wilkinson. p. 2171-2253.
Vazoller, R.F. 1989. Estudos sobre isolamento, caracterização e crescimento de culturas puras de bactérias
metanogênicas provenientes de biodigestores de lodo de esgoto. Dissertação de Mestrado. São Paulo:
Instituto de Ciências Biomédicas/USP.
Vazoller, R.F. 1995. Avaliação do ecossistema microbiano de um biodigestor anaeróbio de fluxo ascendente e
manta de lodo, operado com vinhaça sob condições termofílicas. Tese de Doutorado. São Carlos: Escola
de Engenharia de São Carlos/USP.
24
R.F. Vazoller, G.P. Manfio & V.P. Canhos
Vazoller, R.F. 1997. Microbial aspects of thermophilic anaerobic biodigestion of vinasse. In: Proceedings of
ISME 7 Progress in Microbial Ecology. Santos. p. 527-532.
Vazoller, R.F. & Rech, C. 1992. A simple method for storage of methanogenic bacteria in liquid nitrogen.
Revista Brasileira de Microbiologia, 23:54-56.
Vazoller, R.F.; Rech, C.M.; Figueiredo, M.G. & Giaj-Levra, L.A. 1988. Bacterial identification of granular
sludge from domestic sewage UASB-reactor. In: Proceedings of the 5th International Symposium on
Anaerobic Digestion, Bologna, Itália. p. 61-64.
Woese, C.R. 1995. When is a prokayote not a prokaryote? In: Archaea A laboratory manual. Thermophiles.
Robb, F.T. & Place, A.R. (eds.). New York: Cold Spring Harbor Laboratory Press. p. xv-xvi.
Oremland, R.S. 1988. Biogeochemistry of methanogenic bacteria. In: Biology of anaerobic microorganisms.
Zehnder, A.J.B. (eds). Cap. 13. New York: John Wiley & Sons. p. 641-705.