Universidade Federal de Ouro Preto
Núcleo de Pesquisas em Ciências Biológicas
Laboratório de Imunopatologia
Cinética de migração celular e expressão da enzima óxido nítrico sintase
induzida (iNOS) na pele de cães imunizados com componentes antigênicos de
uma nova vacina contra Leishmaniose Visceral Canina (LVC)
Juliana Vitoriano de Souza
Universidade Federal de Ouro Preto
Núcleo de Pesquisas em Ciências Biológicas
Laboratório de Imunopatologia
Cinética de migração celular e expressão da enzima óxido nítrico sintase
induzida (iNOS) na pele de cães imunizados com componentes antigênicos de
uma nova vacina contra Leishmaniose Visceral Canina (LVC)
Dissertação apresentada ao Programa de Pós-Graduação em Ciências
Biológicas do Núcleo de Pesquisas em Ciências Biológicas da
Universidade Federal de Ouro Preto, como parte integrante dos
requisitos para obtenção do Título de Mestre em Ciências Biológicas,
área de concentração Imunobiologia de Protozoários.
Orientador: Profa Dra. Cláudia Martins Carneiro
Co-Orientador: Prof. Dr. Alexandre Barbosa Reis
Ouro Preto
2008
VITORIANO-SOUZA, J.
Colaboradores
Colaboradores:
Dr. Rodrigo Corrêa Oliveira I
Dr. Rodolfo Cordeiro Giunchetti II
Mestranda Nádia Moreira das Dores II
Dr. Nélder Figueiredo Gontijo III
I - Laboratório de Imunologia Celular e Molecular - IRR - Fiocruz, Belo Horizonte, Minas
Gerais.
II - Laboratório de Imunopatologia - Núcleo de Pesquisas em Ciências Biológicas, Universidade
Federal de Ouro Preto, Ouro Preto, Minas Gerais.
III - Laboratório de Fisiologia de Insetos Hematófagos, Departamento de Parasitologia, Instituto
de Ciências Biológicas, Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais.
i
VITORIANO-SOUZA, J.
Dedicatória
Aos meus queridos pais Terezinha e
Juarez pelo amor e exemplo de vida
ii
“A mente que se abre a uma nova idéia, jamais voltará ao seu tamanho original”
Albert Einstein
iii
VITORIANO-SOUZA, J.
Agradecimentos
A Deus, pela presença constante em todos os dias da minha vida, por iluminar meu caminho e
sempre me dar força para seguir em frente.
Em especial a minha família; aos meus queridos pais (Juarez e Terezinha) e irmãos pelas
orações, pelos conselhos e força dada para realizar este trabalho e principalmente, pelo grande
amor depositado em mim nos momentos bons e ruins de minha vida.
A minha querida família ouro-pretana: A REP. BICHO DO MATO. Agradeço por todos os
momentos inesquecíveis vividos e principalmente, pelas amizades feitas durante todos esses
anos. Em especial, a Tatiana, Aracely, Karla, Lizziani, Handreane, Pauline, Geórgia, Vanessa,
Valdilene, Juliana, Lívia, Vívian, Karina, Isabel, Jussara, Luiza, Fernanda, Fabiana e Amanda.
Vocês contribuíram muito para essa conquista.
Ao professor Alexandre por todos esses anos de convivência e pela orientação. Obrigada pela
oportunidade, confiança depositada, amizade, incentivo, exemplo de determinação e
profissionalismo.
À professora Cláudia pela orientação. Obrigada pela imensa ajuda neste trabalho e
principalmente, pelo exemplo de competência, profissionalismo e equilíbrio.
À Grande amiga Nádia, que esteve presente durante toda esta caminhada, da graduação ao
mestrado. Obrigada pela enorme ajuda na realização deste trabalho, pelas palavras amigas
durante os momentos difíceis e alegrias. Valeu muito!!! Pode ter certeza que ainda vamos rir
muito de tudo isso.
À Paula e Amandinha, por esses dois anos de ótima convivência e amizade. Em pouco tempo,
vocês se tornaram grandes amigas que levarei sempre comigo. Obrigada pelo carinho, força e
incentivo em todos os momentos.
Ao Rodolfo pela imensa colaboração durante todo o trabalho e pelo exemplo de
profissionalismo. Ao Wendel pela força, apoio e por se mostrar sempre disposto a ajudar. À
Raquel e Rafael pela boa convivência no mestrado e laboratório.
À Maria Chaves, técnica do laboratório de Imunopatologia, pela grande contribuição neste
trabalho e, especialmente, pelo cuidado dedicado ao nosso laboratório e aos alunos de iniciação
científica.
A todos os amigos do laboratório de Imunopatologia, pela convivência e colaboração nesse
trabalho. Obrigada a todos por contribuírem para o crescimento do grupo de pesquisas e por
fazerem deste ambiente um excelente local de trabalho, em especial, Bruno e Rodrigo pela ajuda;
Henrique, Samuel, Levi, Jamille, Fernando, Sheler, Ana, Carol, Kátia, Kelly, Taís e Lorena . Ao
professor Wanderson, que apesar do pouco tempo de convívio, sempre se mostrou pronto para
ajudar.
À Lúcia, Érika, Fernanda, Ivo e Thiago pela convivência durante toda a graduação.
A todos os amigos da pós-graduação, professores e à Cida pela atenção e disponibilidade em
resolver todos os nossos pedidos.
iv
VITORIANO-SOUZA, J.
Agradecimentos
A todos os funcionários do Biotério e Canil pelo cuidado e manutenção das colônias de animais,
tão importantes para a realização das pesquisas.
v
VITORIANO-SOUZA, J.
Sumário
LISTA DE GRÁFICOS .............................................................................................................viii
LISTA DE FIGURAS...................................................................................................................ix
LISTA DE ABREVIATURAS......................................................................................................x
RESUMO......................................................................................................................................xii
ABSTRACT ................................................................................................................................xiii
1 - Introdução ..................................................................................................................................1
2 - Revisão da Literatura .................................................................................................................5
2.1 - Leishmaniose Visceral Canina (LVC).................................................................................6
2.2 - Vacinas contra LVC ............................................................................................................7
2.3 - Antígenos e adjuvantes vacinais .........................................................................................9
2.4 - Migração celular...............................................................................................................11
2.5 - Expressão de iNOS na pele ...............................................................................................14
3 - Justificativa ..............................................................................................................................16
4 - Objetivos ..................................................................................................................................18
4.1 - Objetivo Geral: .................................................................................................................19
4.2 - Objetivos específicos:........................................................................................................19
5 - Material e Métodos ..................................................................................................................20
5.1 - Delineamento ....................................................................................................................21
5.2 - Animais..............................................................................................................................22
5.3 - Antígenos e adjuvantes vacinais .......................................................................................22
5.4 - Grupos Experimentais.......................................................................................................23
5.5 - Processamento do material...............................................................................................23
5.6 - Imuno-histoquímica para detecção da expressão da enzima iNOS..................................24
5.7 - Análises Histopatológicas .................................................................................................25
5.8 - Análises quantitativas .......................................................................................................25
5.8.1 - Contagem diferencial (%) ..............................................................................................25
5.8.2 - Análise quantitativa do infiltrado celular na derme e hipoderme .................................25
5.8.3 - Análise quantitativa da área marcada pela enzima iNOs..............................................26
vi
VITORIANO-SOUZA, J.
Sumário
5.9 - Análise estatística..............................................................................................................26
6 - Resultados ................................................................................................................................27
6.1 - Análises Anátomo e Histopatológicas...............................................................................28
6.2 - Cinética de migração celular na derme de cães ...............................................................30
6.3 - Cinética de migração celular seletiva na derme superior de cães ...................................32
6.3.1 - Migração de neutrófilos na derme superior de cães .....................................................32
6.3.2 - Migração de macrófagos na derme superior de cães ....................................................33
6.3.3 - Migração de eosinófilos na derme superior de cães .....................................................34
6.3.4 - Migração de linfócitos na derme superior de cães ........................................................35
6.4 - Cinética de migração celular seletiva na derme profunda de cães ..................................36
6.4.1 - Migração de neutrófilos na derme profunda de cães ....................................................36
6.4.2 - Migração de macrófagos na derme profunda de cães...................................................37
6.4.3 - Migração de eosinófilos na derme profunda de cães ....................................................38
6.5 - Cinética de migração celular na hipoderme de cães........................................................39
6.5 - Cinética de migração celular seletiva na hipoderme de cães...........................................42
6.5.1 - Migração de neutrófilos na hipoderme de cães .............................................................42
6.5.3 - Migração de macrófagos na hipoderme de cães ...........................................................43
6.5.4 - Migração de eosinófilos na hipoderme de cães .............................................................44
6.5.2 - Migração de linfócitos na hipoderme de cães ...............................................................45
6.6 – Análise semiquantitativa da expressão da enzima óxido nítrico sintase induzida (iNOS)
na pele de cães inoculados com diferentes componentes..........................................................46
6.7 - Análise quantitativa da marcação da enzima óxido nítrico sintase induzida (iNOS) na
pele de cães inoculados com diferentes componentes...............................................................47
6.8 - Correlações entre o número de núcleos de núcleos de células e área marcada pela
enzima iNOS na derme e hipoderme .........................................................................................50
6.8.1 - Correlações na derme superior .....................................................................................50
6.8.2 - Correlações na derme profunda ....................................................................................51
7 - Discussão..................................................................................................................................54
8 – Conclusões...............................................................................................................................68
9 - Referências Bibliográficas .......................................................................................................68
vii
VITORIANO-SOUZA, J.
Lista de Gráficos
LISTA DE GRÁFICOS
Gráfico 1: Cinética do infiltrado celular na derme de cães inoculados com diferentes
componentes. .................................................................................................................................30
Gráfico 2: Cinética de migração seletiva de neutrófilos na derme superior de cães inoculados
com diferentes componentes .........................................................................................................33
Gráfico 3: Cinética de migração seletiva de macrófagos na derme superior de cães inoculados
com diferentes componentes. ........................................................................................................34
Gráfico 4: Cinética de migração seletiva de eosinófilos na derme superior de cães inoculados
com diferentes componentes .........................................................................................................35
Gráfico 5: Cinética de migração seletiva de linfócitos na derme superior de cães inoculados com
diferentes componentes. ................................................................................................................36
Gráfico 6: Cinética de migração seletiva de neutrófilos na derme profunda de cães inoculados
com diferentes componentes .........................................................................................................37
Gráfico 7: Cinética de migração seletiva de macrófagos na derme profunda de cães inoculados
com diferentes componentes .........................................................................................................38
Gráfico 8: Cinética de migração seletiva de eosinófilos na derme profunda de cães inoculados
com diferentes componentes .........................................................................................................39
Gráfico 9: Cinética de migração celular na hipoderme de cães inoculados com diferentes
componentes. .................................................................................................................................40
Gráfico 10: Cinética de migração de neutrófilos na hipoderme de cães inoculados com diferentes
componentes. .................................................................................................................................43
Gráfico 11: Cinética de migração de macrófago na hipoderme de cães inoculados com diferentes
componentes antigênicos...............................................................................................................44
Gráfico 12: Cinética de migração de eosinófilos na hipoderme de cães inoculados com diferentes
componentes ..................................................................................................................................45
Gráfico 13: Cinética de migração de linfócitos na hipoderme de cães inoculados com diferentes
componentes ..................................................................................................................................46
Gráfico 14: Análise da produção da enzima iNOS na pele de cães inoculados com diferentes
componentes. .................................................................................................................................48
Gráfico 15: Correlação entre o número de núcleos totais e área marcada pela enzima iNOS na
derme e hipoderme ........................................................................................................................50
Gráfico 16: Correlações na derme superior...................................................................................52
Gráfico 17: Correlações na derme profunda..................................................................................53
viii
VITORIANO-SOUZA, J.
Lista de Figuras
LISTA DE FIGURAS
Figura 1: Delineamento experimental ilustrando as atividades realizadas no presente estudo. ....21
Figura 2: Alterações macroscópicas na pele de cães inoculados intradermicamente com
diferentes componentes antigênicos ..............................................................................................29
Figura 3: Fotomicrografia do infiltrado celular da derme de cães imunizados com saponina,
LBSap e CS. ..................................................................................................................................31
Figura 4: Fotomicrografia do infiltrado celular da hipoderme de cães imunizados com saponina,
LBSap e CS. ..................................................................................................................................41
Figura 5: Fotomicrografia da IMH para detecção da enzima iNOS na derme e hipoderme de cães
imunizados com LBSap.................................................................................................................49
ix
VITORIANO-SOUZA, J.
Lista de Abreviaturas
LISTA DE ABREVIATURAS
%
ºC
µg
µL
µm2
µm
BCG
Ca2+
CD
CD4 +
CD8 +
CMSP
DAB
DTH
eNOS
FML
Percentual
Graus Celsius
Micrograma
Microlitro
Micrômetro quadrado
Micrômetro
Bacillus Calmette Guérin
Cálcio
Cluster differenciation (grupamento de diferenciação)
Marcador de superfície celular da subpopulação de linfócitos
Marcador de superfície celular da subpopulação de linfócitos T
citotóxicos/supressores
Células mononucleares do sangue periférico
Diaminobenzidina
Reação de hipersensibilidade do tipo tardia
Enzima óxido nítrico sintase endotelial
Fucose manose ligante
GRO-α
HE
h
IFN-γ
Ig
IgG
IgG 1
IgG 2
IL-1
IL-2
IL-4
IL-8
IL-10
IL-12
IL-13
IL-18
IMH
iNOS
ISCOMs
kDa
kg
LC
LPG
LPS
LTA
Oncogene-α relacionada ao crescimento
Hematoxilina-eosina
Horas
Interferon-gama
Imunoglobulina
Imunoglobulina da classe G
Imunoglobulina da subclasse G1
Imunoglobulina da subclasse G2
Interleucina 1
Interleucina 2
Interleucina 4
Interleucina 8
Interleucina 10
Interleucina 12
Interleucina 13
Interleucina 18
Imuno-histoquímica
Enzima óxido nítrico sintase induzida
Complexos de Imunoestimulação
Quilodálton (peso molecular)
Quilograma
Leishmaniose cutânea
Lipofosfoglicano
Lipopolissacarídeo
Leishmaniose Tegumentar Americana
x
VITORIANO-SOUZA, J.
LV
LVC
mg
MHC
MIP-1α e β
mL
nNOS
NO
NOS
OCT
OMS
p.i.
PBS
PCR
pH
SMF
SRD
TCRs
TNF- α
UV
WHO
Lista de Abreviaturas
Leishmaniose Visceral
Leishmaniose Visceral Canina
Miligrama
Complexo principal de histocompatibilidade
Proteína inflamatória de macrófagos 1α e β
Mililitro
Enzima óxido nítrico sintase neuronal
Óxido nítrico
Enzima óxido nítrico sintase
Optimal Cutting Temperature Médium
Organização Mundial de Saúde
Pós-imunização
Phosphate buffer saline
Reação da polimerase em cadeia
Potencial hidrogeniônico
Sistema mononucluear fagocitário
Sem raça definida
Receptores de células T
Fator de Necrose Tumoral tipo α
Raios ultra-violeta
World Health Organization
xi
VITORIANO-SOUZA, J.
Resumo
RESUMO
A busca por novos imunobiológicos contra LVC, capazes de ativar uma resposta imune protetora
e duradoura no cão, tem se intensificado na última década. No entanto, ainda são poucos os
trabalhos que explicam os mecanismos iniciais da resposta inata pós-imunização (p.i) in situ. O
presente estudo teve como objetivo central avaliar a cinética de migração celular na pele de cães
frente aos distintos componentes antigênicos da vacina LBSap. Quinze cães sem raça definida
(SRD) de ambos os sexos com idade variando de 8 a 12 meses, foram inoculados
intradermicamente na região dorsal com diferentes componentes antigênicos e divididos em
cinco grupos experimentais: (i) grupo controle salina (CS) recebeu 250 µL de salina estéril 0.9%;
(ii) grupo Saponina (Sap) recebeu 1 mg de saponina; (iii) grupo Saliva (Sal) recebeu 1 ácino de
glândula de Lu. longipalpis por inóculo, (iv) grupo antígeno de L. braziliensis (LB) recebeu 600
µg de proteína de L. braziliensis e (v) grupo vacina de L. braziliensis associado a saponina
(LBSap) recebeu 600 µg de proteína de L. braziliensis e 1 mg de saponina. Após os tempos de 1,
12, 24, 48 e 96 horas, foram realizadas biópsias de pele. Os resultados obtidos indicaram que o
adjuvante saponina isoladamente ou em associação ao antígeno de L. braziliensis induziu forte
reação inflamatória aguda local acompanhada por edema, congestão e hemorragia, no entanto,
essas reações não evoluíram para lesões ulceradas. A morformetria por análise de imagem da
intensidade do infiltrado celular demonstrou aumento do número de núcleos celulares no grupo
Sap na derme. Na hipoderme, esse aumento foi mais intenso que na derme, nos grupos Sap e
LBSap. De modo geral, o perfil celular encontrado nos grupos Sap e LBSap foi composto de
neutrófilos, linfócitos e eosinófilos, sugerindo um perfil misto de resposta imune (tipo1/ tipo2),
fundamental para o controle da infecção. Essas análises ressaltam o papel da saponina como
potente estimulador de resposta celular, importante para o início dos processos de apresentação
antigênica e desenvolvimento da resposta protetora pós-imunização. No grupo Sal, observou-se
aumento de eosinófilos na hipoderme demonstrando o papel da saliva como imunomodulador no
início da infecção por Leishmania. Observou-se também aumento da produção de iNOS nos
grupos Sap e LBSap permitindo-nos inferir que esse aumento poderia ser importante para o
desenvolvimento de um microambiente composto por IFN-γ e de quimiocinas. Desse modo,
podemos concluir que a imunização por LBSap e pelo adjuvante saponina produz um perfil
semelhante ao de resistência a infecções por Leishmania, contribuindo para o melhor
entendimento dos eventos da imunidade inata gerados pela vacina LBSap.
xii
VITORIANO-SOUZA, J.
Abstract
ABSTRACT
The search for new immunobiologicals against Canine Visceral Leishmaniasis (CVL), able to
activate a long-term and protective immune response in dogs, has intensified in the last decade.
However, it still remains to be elucidated that mechanisms of the innate immune response in situ
after immunization (a.i.). The main objective of the present study was to assess the kinetics of
cell migration in the skin dogs with distinct antigenic components of the LBSap-vaccine. In the
current study, 15 mixed breed dogs of both genders aging from 8-12-month old were inoculated
in the dorsal region by intradermic injections and subdivided within five experimental groups: (i)
saline control group (SC) received 250 µL of sterile 0.9% saline; (ii) saponin group (Sap),
received 1 mg of saponin; (iii) Saliva group (Sal), received 1 acini of salivary glands of
Lutzomyia longipalpis; (iv) Leishmania braziliensis antigen group (LB) received 600 µg/protein
of L. braziliensis; and (v) L. braziliensis vaccine plus saponin group (LBSap) received 600
µg/protein of L. braziliensis plus 1 mg of saponin. After the times of 1, 12, 24, 48 and 96 hours,
skin biopsies were performed. The results indicated that saponin adjuvant alone or combined
with L. braziliensis antigen induced strong local acute inflammatory reaction accompanied by
edema, hemorrhage and congestion. However, these reactions not progressed to ulcerated
lesions. The image analysis by morformetry showed the intensity of cell infiltrate and an
increased number of cell-nuclei in dermis of the Sap group. In hypodermis, this increase was
more intense than in the dermis, observed in LBSap and Sap groups. Overall, the cell profile
found in Sap and LBSap was composed of neutrophils, lymphocytes and eosinophils suggesting
a profile of mixed immune response (type1/type2) potentially compatible with effective control
of the etiological agent of CVL. These analyses highlight the role of saponin as a potent
stimulator of cell-immune response and important on antigenic priming events and development
of protective immune response after immunization. In group Sal, it was observed increase of
eosinophils in the hypodermis demonstrating the role of saliva as immunomodulator at the
beginning of Leishmania infection. There was also increased production of iNOS in Sap and
LBSap groups. These data, allowing us to infer that this increase could be important for the
microenvironment composed of cytokines and chemokines with of type-1 profile. Thus, we can
conclude that immunization by LBSap and the saponin adjuvant elicited a potential innateimmune activations status compatible with effective control of the resistance to infection by
Leishmania and contributing to a better understanding of the innate-immunity events induced by
the LBSap vaccine.
xiii
1 - Introdução
VITORIANO-SOUZA, J.
Introdução
As leishmanioses são doenças complexas e cosmopolitas causadas pela invasão das
células do sistema mononuclear fagocitário (SMF) do baço, fígado, medula óssea e pele por
parasitos protozoários hemoflagelados do gênero Leishmania, transmitidos durante o repasto
sangüíneo de fêmeas hematófagas dos gêneros Phlebotomus no Velho Mundo e Lutzomyia no
Novo Mundo (Lainson & Shaw, 1987). Estudos recentes reforçam a hipótese de que os agentes
etiológicos Leishmania infantum e Leishmania chagasi, causadores da leishmaniose visceral
(LV) no Mediterrâneo e América, respectivamente, são variações da mesma espécie (Mauricio et
al., 1999; Maurício et al., 2000; Lukes et al.,2007).
O estabelecimento da infecção por Leishmania depende da fagocitose de formas
promastigotas flageladas por células do SMF e a subseqüente transformação em amastigotas.
Após essa fase, as amastigotas se multiplicam dentro de macrófagos na pele onde foram
inoculadas e posteriormente se disseminam para outros órgãos, causando alterações patológicas
(Pearson, 1993).
Esse grupo de doenças apresenta um amplo espectro de formas clínicas, manifestando-se
através de lesões ulceradas na pele, inflamação destrutiva na mucosa ou infecção visceral
disseminada (kalazar) e são caracterizadas em função da complexidade biológica do parasito,
reservatórios e vetores, bem como das interações parasito-hospedeiro (Saraiva et al., 1989).
A LV ou kalazar é uma doença grave e freqüentemente letal, se não tratada, considerada a
mais devastadora dentre as formas clínicas das leishmanioses (Desjeux, 2004). Geralmente é
restrita geograficamente a regiões que são altamente infestadas pelo inseto vetor, sendo
endêmica em países como Índia, Sudão, Nepal, Bangladesh e Brasil que juntos representam
cerca de 90% dos casos de LV no mundo (WHO, 2005). Até o final da década de 40, a LV era
um problema restrito a poucas regiões do globo e a atenção mundial estava voltada para a
resolução dos problemas ocasionados pelas grandes epidemias bacterianas e virais fatais (Garg &
Dube, 2006).
Atualmente, essa doença encontra-se em expansão em novas áreas (Enserink, 2000) e
centros urbanos (Silva et al., 2000). O número de casos relatados de LV no mundo aumentou
dramaticamente nos últimos anos, devido à migração de populações das áreas rurais para os
centros urbanos. Estes fatores, entre outros, contribuíram para a adaptação do ciclo de
transmissão no ambiente peridomiciliar em grandes cidades do Brasil como Belo Horizonte,
Montes Claros, Rio de Janeiro, Salvador, Fortaleza, Governador Valadares, Araçatuba,
Santarém, Corumbá, Teresina, Natal, São Luís, Camaçari e mais recentemente, nos municípios
de Três Lagoas, Campo Grande e Palmas (Arias et al., 1996; Franca-Silva et al., 2003;
Ministério da Saúde, 2003; Malaquias et al., 2007).
2
VITORIANO-SOUZA, J.
Introdução
Além disso, diversas falhas nos programas de controle de reservatórios e vetores, somado
ao caráter emergente e re-emergente da LV e sua associação às infecções oportunistas tais como
a AIDS (Badaró et al., 1986; Altes et al., 1991; Gradoni et al., 1993) bem como aos recentes
relatos da seleção de cepas resistentes a drogas, levou a Organização Mundial de Saúde (OMS) a
considerar a leishmaniose como uma das doenças parasitárias de maior importância no mundo.
Neste contexto, é importante destacar o papel dos cães como reservatório na LV devido a
alta carga parasitária na pele de animais infectados (Deane & Deane, 1955), alta prevalência da
doença e a demonstração de que eles são capazes de infectar flebotomíneos mesmo após o
tratamento (Molina et al., 1994; Travi et al., 2001; Reis et al., 2006a,b; Giunchetti et al., 2006).
O controle da leishmaniose visceral canina (LVC) preconizado pela OMS baseia-se na
eutanásia dos animais infectados, uma vez que o tratamento com drogas utilizadas com sucesso
em casos humanos como glucanato antimonial de sódio ou stiboglucanato de sódio, mostrou
baixa eficácia em cães (Alencar, 1959; Alvar et al., 1994). Todavia, a eliminação desses cães
nem sempre é aceita por razões éticas e sociais (Desjeux, 2004). Além da eutanásia dos cães
soropositivos, outras estratégias são utilizadas como controle epidemiológico tais como,
tratamento sistemático de todos os casos humanos e utilização de inseticidas residuais como
forma de controle vetorial (Magalhães et al., 1980).
Essas medidas de controle nem sempre são fáceis de serem adotadas, desse modo, alguns
fatores como a gravidade da doença, acompanhada pelas limitações do tratamento e a
possibilidade de surgimento de epidemias levam a OMS a estimular o desenvolvimento de
vacinas contra LVC (O'Hagan & Valiante, 2003). Sendo assim, a imunoprofilaxia pode ser
empregada como uma alternativa promissora para a prevenção da LVC, atuando no sentido de
diminuir a incidência dos casos caninos e, conseqüentemente, da doença humana (Marzochi et
al., 1985; Genaro, 1993; Hommel et al., 1995; Genaro et al., 1996; Gradoni, 2001; Handman,
2001, Desjeux, 2004; Ravindran & Ali, 2004; Requena et al., 2004).
Muitos estudos têm sido realizados com a finalidade de desenvolver uma vacina segura
contra LVC, com capacidade protetora e sem efeitos adversos ao hospedeiro. Entretanto, existem
algumas dificuldades como a grande diversidade antigênica do parasito, bem como o fato desse
protozoário acometer diferentes hospedeiros vertebrados e, desse modo, apresentar vários tipos
de interação parasito-hospedeiro (Handman, 2001).
A elaboração de vacinas deveria levar em consideração fatores como a escolha de
moléculas utilizadas como antígenos, dos adjuvantes, via de administração e tempo requerido
para a imunidade protetora (Cox, 1997). Até o momento, inúmeras vacinas foram testadas a
exemplo das vacinas com parasitos mortos ou parasitos vivo-atenuados, vacinas de antígenos
3
VITORIANO-SOUZA, J.
Introdução
derivados de DNA recombinante, vacinas de peptídeos sintéticos, vacinas de antígenos não
protéicos (lipofosfoglicano – LPG) e vacinas de DNA (Hommel et al., 1995; Gradoni, 2001;
Handman, 2001; Mauel, 2002; Brodskyn et al., 2003; Ravindran & Ali, 2004; Requena et al.,
2004).
A falta de um modelo experimental adequado para testes de vacinas e drogas contra LVC
é um agravante para o desenvolvimento de imunoprofiláticos e novos fármacos. Além disto,
testes de seleção de potenciais antígenos vacinais, sejam no contexto in vitro ou mesmo in vivo,
são limitados. Contudo, o cão pode ser considerado um excelente modelo para o estudo da LV
humana, devido às semelhanças entre a LV canina e a doença humana (Peters & KillickKendrick, 1987) e também devido à sua importância epidemiológica como reservatório da
leishmaniose visceral . O padrão imunopatológico da doença no homem e no cão é similar, com
um longo período de infecção assintomática seguida por anemia, aumento dos linfonodos e febre
(Abranches et al., 1991; Genaro, 1993; Reis, 2001; Giunchetti, 2004; Reis et al., 2006a,b,c;
Giunchetti et al., 2006). Assim como em humanos, a infecção em alguns cães pode permanecer
assintomática (Pinelli et al., 1995), uma das poucas diferenças são as dermatopatias caninas,
raramente detectadas em humanos (Hommel et al., 1995; Reis, 2001; Giunchetti et al., 2006).
Outro agravante ao estudo da LVC é a falta de marcadores imunológicos e reagentes para o cão,
o que retarda o progresso da imunologia canina em detrimento dos estudos em humanos e
camundongos (Moreno & Alvar, 2002; Reis et al., 2005; Reis et al., 2006a; Giunchetti et al.,
2008a).
Considerando a importância da LVC no contexto mundial, como uma importante
protozoonose amplamente distribuída, no presente estudo foi utilizado o cão como modelo
experimental. Neste sentido, se propôs avaliar a cinética de migração celular utilizando antígenos
e adjuvantes vacinais que compõem um imunobiológico (candidato à vacina contra LVC)
recentemente, desenvolvido pela UFOP e FIOCRUZ. Este trabalho pretende contribuir para o
estudo dos aspectos relacionados à cinética do infiltrado inflamatório na pele como estratégia
adicional para o estabelecimento de biomarcadores para avaliação de parâmetros relacionados a
inocuidade de diferentes componentes deste imunoprofilático anti-LVC.
4
2 - Revisão da Literatura
VITORIANO-SOUZA, J.
Revisão da Literatura
2.1 - Leishmaniose Visceral Canina (LVC)
A infecção de cães por parasitos do gênero Leishmania apresenta caráter bastante
variável, podendo desenvolver infecção sintomática resultando em morte, enquanto outros
permanecem assintomáticos, ou ainda apresentam poucos sinais clínicos, sendo assim chamados
de oligossintomáticos (Abranches et al., 1991; Genaro, 1993; Reis, 2001; Reis et al., 2006a). As
principais manifestações clínicas da LVC são linfoadenopatia, anemia, diarréia, alopecia,
dermatite, onicogrifose, perda de peso, caquexia, problemas de locomoção e conjuntivite
(Genaro, 1992; Giunchetti et al., 2006; Reis et al., 2006b).
Muitos trabalhos relacionam a sintomatologia da LVC a mudanças imunológicas
envolvendo células T. Essas mudanças incluem ausência de hipersensibilidade do tipo tardia
(DTH) para antígenos de Leishmania (Cardoso et al., 1998; Solano-Gallego et al., 2000),
diminuição do número de células T no sangue periférico (Pinelli et al., 1994; Bourdoiseau et al.
1997; De Luna et al., 1999; Martinez-Moreno et al., 1995; Reis, 2001; Reis et al., 2006c),
ausência de IFN-γ e da produção de IL-2 por células mononucleares do sangue periférico
(CMSP) in vitro (Pinelli et al., 1994). Todavia, observam-se altos títulos de anticorpos antiLeishmania, presentes em animais sintomáticos, sugerindo que esses não são protetores (Pinelli
et al., 1994; Martinez-Moreno et al., 1995; Reis, 2001; Santos-Gomes et al., 2002; Reis et al.,
2006b).
A resistência a LVC tem sido associada com a ativação da resposta tipo 1 através da
produção de IFN-γ, IL-2 e TNF-α. O principal mecanismo efetor envolvido na resposta protetora
de cães infectados por Leishmania é a ativação de macrófagos por IFN-γ e TNF-α para destruir
amastigotas intracelulares, utilizando a via L-arginina do óxido nítrico (NO) (Pinelli et al., 1994;
Reis et al., 2006a; Reis et al., 2006b; Reis et al., 2006c; Lage et al., 2007). A produção de NO a
partir do aminoácido L-arginina pela óxido nítrico sintase induzida (iNOS) é um mecanismo
chave na defesa por fagócitos nos mamíferos (Nathan et al., 2000).
O papel das citocinas tipo 2 ainda não é bem definido na LVC. Evidências demonstram
que CMSP de cães assintomáticos estimulados com antígenos de Leishmania apresentam
transcritos de mRNA de IL-2, IFN-γ e IL-10, ou seja, um padrão misto de citocinas tipo 1 e tipo
2. Além disso, recentemente nosso grupo de pesquisa demonstrou que 100% dos cães com alta
densidade parasitária no baço expressam IL-10 e que parte destes animais apresentam níveis
relativos de IFN-γ (Lage et al., 2007) demonstrando a participação da IL-10 na modulação da
resposta imune juntamente com a presença de IFN-γ na LVC natural.
6
VITORIANO-SOUZA, J.
Revisão da Literatura
2.2 - Vacinas contra LVC
No século 20, o desenvolvimento e uso de vacinas tornaram-se uma das mais importantes
ferramentas contra doenças infecciosas. No campo das doenças parasitárias, é importante
ressaltar que o entendimento dos padrões similares de resposta protetora na LV nos modelos
murinos, caninos e em humanos possibilitou a descoberta e o estudo de novos imunobiológicos
resultando até o momento, na descrição de três classes de vacinas, que representam diferentes
gerações de antígenos vacinais.
No Brasil, a LEISHVACIN® foi o protótipo de uma vacina utilizada contra LVC, sendo
desenvolvida inicialmente contra Leishmaniose Tegumentar Americana (LTA) pelo grupo
liderado pelo Prof. Wilson Mayrink do departamento de Parasitologia da UFMG, representando
uma vacina de primeira geração. Essa vacina era composta inicialmente por antígenos de
promastigotas sonicadas de cinco cepas de Leishmania, sendo sua composição modificada mais
tarde apresentando apenas uma cepa de L. (L.) amazonensis (PH8) a fim de facilitar a
padronização do antígeno vacinal (Mayrink et al.,1979; Mayrink et al.,1985; Mayrink et al.,
1986; Genaro et al., 1996; Marzochi et al.,1985). Em 1996, Mayrink et al. testaram uma vacina
em cães compostas por L. braziliensis (Cepa MCAN/BR/1972/C348) associada ao adjuvante
BCG (Bacillus Calmette-Guérin) que demonstrou proteção de 90% dos animais vacinados
acompanhados por 470 dias em um estudo fechado de Fase I e II em canil (Mayrink et al., 1996).
Um outro ensaio vacinal conduzido em canil, contou com a avaliação de uma outra
vacina anti-LVC agora composta por L. amazonensis (IFLA/BR/1967/PH8) e L. braziliensis
(MCAN/BR/1972/C348) associada ao adjuvante BCG (Fujiwara, 2003; Giunchetti et al., 2008b).
Os resultados de imunogenicidade evidenciaram indução da atividade linfoproliferativa in vitro
em células T Leishmania-específicas, com expansão de linfócitos T Thy-1+ e T CD8+ circulantes
(Giunchetti et al., 2008a), que vem sendo atribuídos a um perfil imunofenotípico de resistência
contra LVC (Reis et al., 2006c).
Recentemente, o Laboratório Fort Dodge adquiriu a patente de uma vacina de segunda
geração contra LVC, conhecida por LEISHMUNE® e licenciada pelo Ministério da Agricultura.
Essa vacina foi desenvolvida pelo grupo de pesquisas liderado pela Dra. Clarissa Palatinik de
Sousa da UFRJ. A LEISHMUNE® é composta pelo antígeno complexo glicoprotéico ligante de
fucose e manose (FML - presente na superfície do parasito) associado com o adjuvante saponina
(Palatnik et al., 1990). Estudos com essa vacina demonstraram que a combinação desse antígeno
com saponina atingiu uma proteção média de 87,7% e 84% respectivamente em hamsters e
camundongos Balb/C (Palatnik de Sousa et al., 1994a; Palatnik de Sousa et al.,1994b). Em 1996,
foram realizados estudos em área endêmica de São Gonçalo de Amaranto – RN para testar a
7
VITORIANO-SOUZA, J.
Revisão da Literatura
eficácia da vacina FML, onde se observou através de testes sorológicos e de intradermorreação,
uma proteção de 92% no grupo vacinado, correspondendo a 76% de eficácia vacinal (Da Silva et
al., 2001). Outros trabalhos com a LEISHMUNE® também demonstraram sua alta eficácia em
campo (Borja-Cabrera et al., 2002).
Atualmente, nosso grupo de pesquisa vem se dedicando ao estudo de novas composições
vacinais contra LVC. No entanto, esses estudos iniciaram-se com a entrada do Prof. Odair
Genaro (in memorian) no grupo de pesquisas do Prof. Mayrink na década de 80. Desde então,
muitos trabalhos foram realizados utilizando o modelo canino e antígenos brutos de
promastigotas de cepas dermatotrópicas de Leishmania. Diante dos resultados promissores de
uma vacina de antígeno bruto, composta por promastigotas de Leishmania dermatotrópicas
(Mayrink et al., 1996), e com o estabelecimento de novas metodologias para avaliação
imunológica por nosso grupo, que possibilitaram uma avaliação mais acurada do sistema imune
canino e conseqüentemente da história natural da LVC (Reis, 2001; Giunchetti, 2004; Reis et al.,
2005; Giunchetti et al., 2006; Reis et al., 2006a; Reis et al., 2006b; Reis et al., 2006c; Giunchetti
et al., 2008a); Neste sentido, recentemente foi proposto o estudo de dois novos imunobiológicos
compostos por antígenos brutos de promastigotas de L. braziliensis associados ao adjuvante
saponina (LBSap) e/ou saliva de Lutzomyia longipalpis (LBSapSal) (Giunchetti, 2007;
Giunchetti et al., 2007; Giunchetti et al., 2008a).
Os resultados obtidos até o momento demonstraram que esses imunobiológicos (LBSap e
LBSapSal) apresentam forte capacidade indutora de imunogenicidade relacionada aos altos
níveis de isotipos IgG anti-Leishmania, associados aos altos níveis de linfócitos, particularmente
linfócitos T CD8+ circulantes e linfócitos T CD8+ L. chagasi-específicos. Além disto, outros
fatores demonstraram que estes imunobiológicos são indutores de uma resposta imune
compatível com o controle efetivo do agente etiológico da LVC, tais como intensa atividade
linfoproliferativa L. chagasi-específica e aumento da produção de NO durante a estimulação in
vitro por antígenos solúveis de L. chagasi (Giunchetti, 2007; Giunchetti et al., 2007; Giunchetti
et al., 2008a). Estes resultados estimularam nosso grupo de pesquisa a dar continuidade aos
estudos, focalizando em novas avaliações destes imunoprofiláticos.
Neste sentido, apesar da administração destas vacinas em cães terem se mostrado seguras
(Giunchetti, 2007; Giunchetti et al., 2007; Giunchetti et al., 2008a), a reação no local do inóculo
(edema ou formação de nódulos) nos animais que receberam o adjuvante saponina de forma
isolada ou combinada nas diferentes vacinas avaliadas, estimularam a proposição de uma nova
8
VITORIANO-SOUZA, J.
Revisão da Literatura
avaliação buscando o estudo de novos biomarcadores que auxiliassem na compreensão destes
eventos. Além disto, importantes perguntas poderiam ser respondidas neste trabalho como: (i)
“Quais são as populações celulares presentes nos eventos precoces e tardios no local do inóculo
vacinal?”, (ii) “O antígeno, adjuvante e a saliva de Lu. longipalpis selecionam de forma diferente
as populações celulares que migram para o local do inóculo?”. Para responder a estas e outras
perguntas nós estabelecemos um protocolo experimental para estudar estes eventos por meio da
avaliação da cinética da resposta inflamatória e da expressão da enzima iNOS em diferentes
regiões da pele.
2.3 - Antígenos e adjuvantes vacinais
A busca por novos imunobiológicos por nosso grupo resultou em estudos que estimulam
a utilização de antígenos brutos como potenciais candidatos a vacina contra LVC, uma vez que
estes antígenos são capazes de promover resposta imune celular e reduzir a taxa de infecção
(Mayrink et al., 1996) ou mesmo estimular a modulação do sistema imune, desencadeando uma
resposta celular com envolvimento de linfócitos T CD8+ (Araújo, 2006), importantes para
resistência à infecção por L. chagasi (Reis et al., 2006c).
Nas últimas décadas, diversos estudos buscaram avaliar novos adjuvantes, no entanto, seu
mecanismo de ação continua pouco compreendido. Considerando o papel imunogênico, vários
adjuvantes têm sido avaliados em vacinas como o BCG, citocinas que promovem a resposta tipo
1, saponina, emulsões de óleo, polímeros, lipopolissacarídeos, ISCOMs (complexos de
imunoestimulação) e adjuvante completo de Freund (Vogel et al., 1995; Edelman, 1997).
Os adjuvantes são componentes essenciais para os protocolos vacinais, amplamente
definidos pela sua habilidade de aumentar in vivo a capacidade imunogênica de antígenos que
são co-administrados (Afrin & Ali, 1997). O seu uso pode ser justificado por vários motivos: (i)
melhora a imunogenicidade de antígenos; (ii) aumenta a duração de resposta imune; (iii) modula
a avidez, a especificidade e a distribuição de isotipos ou subclasses de anticorpos; (iv) estimula a
imunidade mediada por células; (v) promove a imunidade das mucosas; (vi) aumenta a resposta
imune em indivíduos imaturos ou senescentes; (vii) diminui a dose de antígeno na vacina e
conseqüentemente, reduz os custos de produção (Singh & O’Hagan, 2003).
Dentre os vários adjuvantes podemos citar a saponina que possui um grande potencial
inflamatório. Esse adjuvante pertence a um grupo quimicamente heterogêneo de glicosídeos
esteróis ou glicosídeos triterpenos, comumente encontrado em cascas, caules, raízes e flores de
plantas como, por exemplo, da Quillaja saponaria Molina. Saponinas possuem a habilidade de
9
VITORIANO-SOUZA, J.
Revisão da Literatura
modular a resposta imune mediada por células, bem como aumentar a produção de anticorpos e
possuem a vantagem de necessitarem de uma baixa dose para exercerem seu papel de adjuvante
(Oda et al., 2000). Além da capacidade estimuladora dos componentes da imunidade específica,
saponinas também apresentam algumas reações imunes não específicas como inflamação (de
Oliveira et al., 2001) e estimulam a resposta imune ao antígeno promovendo a síntese de várias
citocinas como IFN-γ, IL-2, IL-4 e IL-10 (Cox & Couter, 1997). Possivelmente, as saponinas são
os adjuvantes ideais para serem utilizados em ensaios vacinais contra Leishmania spp
(Ravindran & Ali, 2004), pois além de apresentarem baixo custo, possuem formulação simples e
geralmente são consideradas seguras (Cox & Coulter, 1997).
Recentemente, muitos trabalhos têm utilizado o extrato de glândula salivar de Lu.
longipalpis, cuja importância na transmissão de Leishmania chagasi na América já é
amplamente reconhecida na literatura, por aumentar a infecciosidade em camundongos desde os
trabalhos pioneiros realizados por Titus & Ribeiro (1988).
Na saliva de flebotomíneos existe uma variedade de moléculas anti-coagulantes, antiplaquetárias e vasodilatadoras que facilitam o repasto sangüíneo e afetam diretamente a
homeostase, bem como as respostas inflamatória e imune do hospedeiro vertebrado. Muitas
dessas moléculas são imunogênicas e levam a produção de anticorpos e/ou de uma resposta
celular pró-inflamatória aumentando a habilidade dos patógenos de se estabelecerem em seus
hospedeiros (Ribeiro et al., 1986).
Kamhawi et al. (2000) demonstraram que a exposição anterior de camundongos a saliva
ou a picadas de flebotomíneos não infectados antes da inoculação por L. major tem efeito
protetor associado a uma forte resposta de hipersensibilidade retardada com a produção de IL-12
e IFN-γ no sítio de inoculação. Este estudo foi fundamental, pois a partir de então, diversos
pesquisadores passaram a considerar os constituintes da glândula salivar como promissores
antígenos vacinais.
Belkaid et al. (1998) demonstraram além da produção de anticorpos específicos antisaliva, a presença de intenso infiltrado inflamatório composto por neutrófilos, eosinófilos e
macrófagos no local da picada por flebotomíneos. Outros autores observaram que a injeção de
soro de camundongos, previamente incubado com saliva induziu rápido infiltrado composto por
neutrófilos e macrófagos, sugerindo a participação de complexos imunes no início do processo
inflamatório (Silva et al., 2005).
Teixeira et al. (2005) observaram em modelo de bolhas de ar para o estudo de inflamação
em camundongos, aumento do recrutamento de macrófagos promovido por extrato de glândula
salivar de Lu. longipalpis. A capacidade da saliva de flebotomíneo induzir imunidade mediada
10
VITORIANO-SOUZA, J.
Revisão da Literatura
por células pode ser caracterizada pelo desenvolvimento de uma reação de hipersensibilidade do
tipo tardia, caracterizada por infiltrado celular no local da picada (Belkaid et al., 2000). A reação
inflamatória induzida pela saliva de flebotomíneo poderia, primariamente, representar uma
estratégia de sobrevivência, uma vez que o local de inflamação possui maior fluxo sanguíneo que
a pele normal e assim facilitaria a alimentação do inseto. Em etapas posteriores, o recrutamento
de células não adequadamente ativadas, provocado pela modulação da saliva, seria um evento
importante para o estabelecimento do parasito, favorecendo seu desenvolvimento nos
macrófagos (Andrade et al., 2007). Considerando a importância da saliva de flebotomíneo no
estabelecimento da infecção por Leishmania, levantamos a hipótese de que uma vacina composta
por saliva de Lu. longipalpis induziria a produção de anticorpos anti-saliva que neutralizariam as
funções imunomoduladoras da saliva, contribuindo para aumentar os níveis de proteção em
posteriores repastos sanguíneos por flebotomíneos infectados. Estes eventos representariam uma
estratégia adicional para aumentar o repertório antigênico vacinal e conseqüentemente, a eficácia
de imunoprofiláticos anti-Leishmania.
Desse modo, devido à importância desses antígenos e adjuvantes na composição da
vacina contra LVC testada pelo nosso grupo, a análise histológica da pele de cães em diferentes
tempos torna-se relevante, podendo contribuir para o entendimento da resposta imune inicial
desencadeada na imunização pelos diferentes componentes antigênicos através da cinética de
migração celular. Estes eventos são fundamentais para uma melhor compreensão das alterações
relatadas no local do inóculo vacinal na presença do adjuvante saponina e da saliva de Lu.
longipalpis (Giunchetti, 2007; Giunchetti et al., 2007; Giunchetti et al., 2008a).
2.4 - Migração celular
A migração celular representa uma propriedade de fundamental importância das células
imunes, uma vez que permitem a mobilização dessas para o local da infecção por patógenos. O
início e a manutenção da inflamação dependem de diversas famílias de moléculas, tais como
prostaglandinas, tromboxanos, leucotrienos, aminas vasoativas, citocinas pró-inflamatórias e
quimiocinas. Estas moléculas agem sobre o endotélio, tornando-o permissivo à migração de
células inflamatórias (Springer, 1995).
O processo de migração leucocitária pode ser dividido em quatro etapas distintas:
marginação, processo onde os leucócitos assumem posição periférica em relação às células
endoteliais e é influenciado pela modificação das condições hemodinâmicas; captura e
rolamento, onde ocorre adesão transitória de leucócitos ao endotélio; adesão, firme união ao
11
VITORIANO-SOUZA, J.
Revisão da Literatura
endotélio e; diapedese, onde ocorre a passagem de leucócitos entre as células endoteliais em
direção ao tecido inflamado/lesionado (Muller, 2003).
Durante o processo inflamatório ocorre influxo de células, acúmulo de mediadores
inflamatórios, como as citocinas, e aumento na deposição da matriz extracelular. Citocinas
regulam tanto a iniciação quanto a manutenção ou resolução da resposta imune inflamatória,
além disso, elas direcionam o tipo de resposta imune e os mecanismos efetores que medeiam a
resistência a patógenos. Entretanto, algumas citocinas quando produzidas em excesso, podem
induzir alterações em decorrência da liberação demasiada de substâncias tóxicas, incluindo
proteases e espécies reativas de oxigênio (Homey et al., 2006).
Na infecção aguda, neutrófilos são recrutados rapidamente para o local de inflamação e
dominam o influxo inicial de leucócitos. Esse papel imediato de neutrófilos na defesa imune os
torna ideais para controlar o recrutamento de outros tipos celulares para o local da infecção, tais
como células T, monócitos, macrófagos e células dendríticas imaturas (van Gisbergen et al.,
2005). Mais tarde, monócitos/macrófagos substituem a população dominante de neutrófilos,
sugerindo um padrão de recrutamento bimodal envolvendo a troca de neutrófilos por
monócitos/macrófagos (Ryan et al., 1997). Os monócitos recrutados para o local lesado
transformam-se em macrófagos fagocíticos, população de células predominante na fase mais
tardia da infecção. Macrófagos são considerados a mais importante célula reguladora na reação
inflamatória. Eles fagocitam, digerem e matam organismos patogênicos, além de destruir
neutrófilos remanescentes (Li et al., 2007).
Interessantemente, trabalhos sugerem a participação de neutrófilos na polarização da
resposta tipo 1. Embora produzam IL-12 e possam migrar em direção a células T no linfonodo,
essa interação é ineficiente. Todavia, a comunicação de neutrófilos com células dendríticas, que
potencialmente interagem com células T poderia direcionar para essa resposta de maneira
indireta, uma vez que algumas evidências indicam que antígenos fagocitados e processados por
neutrófilos são transferidos para células dendríticas para apresentação para células T (Potter et
al., 2001).
Os eosinófilos são também recrutados para os sítios inflamatórios principalmente em
resposta a estímulos alérgicos do trato respiratório, trato gastrointestinal e pele e, geralmente
estão relacionados a infecções helmínticas (Terui et al., 2000). A presença de eosinófilos pode
estar associada com a fase inicial da infecção por Leishmania (Belkaid et al., 2000) e IL-4 tem
sido descrito como forte indutor da migração de eosinófilos (Sanz et al.,1998). Paranhos et al.,
(1993) detectaram eosinofilia em cães inoculados intradermicamente com saliva de Lu.
12
VITORIANO-SOUZA, J.
Revisão da Literatura
longipalpis. Observações similares foram vistas em processo inflamatório em camundongos
imunizados com uma proteína recombinante de 15 KDa de P. papatasi (Valenzuela et al., 2001)
Os macrófagos podem ser descritos como formas evolutivas dos monócitos circulantes
que saíram do sangue e fixaram residência nos tecidos. Seu papel relaciona-se a destruição dos
fatores etiológicos da inflamação ou complementar a função dos neutrófilos na resposta aguda
através da produção de citocinas (Parham, 2001). Entretanto, sua ação principal fica evidente nas
inflamações crônicas atuando como células apresentadoras de antígeno (APCs) aos linfócitos T
na imunidade específica. Uma vez ativados, os macrófagos liberam uma variedade de citocinas
que possuem funções importantes na imunidade inata e ao mesmo tempo na preparação do
cenário para que a resposta imune adaptativa se desenvolva (Egan et al, 1996; Terui et al, 2000).
Os linfócitos T desempenham seu papel na inflamação aumentando a resposta dos
macrófagos pela produção de citocinas pró-inflamatórias. Enquanto os linfócitos B atuam na
produção de anticorpos e estabelecimento de imunidade específica (Egan et al, 1996; Terui et al,
2000; Parham, 2001).
Estudos sobre o papel de neutrófilos no início da infecção por L. major demonstraram
expressão de mRNA para citocinas tipo 1 e quimiocinas em camundongos resistentes, não sendo
observado esse padrão em camundongos susceptíveis. Entretanto, observou-se que a depleção de
neutrófilos resultou em exacerbação das lesões em camundongos BALB/c infectados por L.
major, sugerindo função protetora de neutrófilos (Chen et al., 2005).
Contrário a esses trabalhos existe uma outra linha de pesquisadores que defendem o papel
de neutrófilos através da sua manutenção no local da infecção, como reservatórios para uma
tardia liberação de Leishmania (Aga et al. 2002; Beil et al., 1992). Desse modo, neutrófilos
participariam do início da infecção, permitindo a sobrevivência desses parasitos até a chegada de
macrófagos, onde estas células fagocitariam os neutrófilos sem ativarem os mecanismos
microbicidas, instalando desse modo a infecção (Aga et al. 2002).
É importante citar que na resistência a infecção por Leishmania, além de linfócitos T
CD4+ , também os linfócitos T CD8+ são requeridos durante o controle primário da infecção com
L. major na pele (Belkaid et al., 2002). Embora seja descrito que a principal função protetora
dessas células seria na liberação de IFN-γ no sítio da infecção, elas poderiam também participar
na citólise das células infectadas no local da infecção (Tsagozis et al, 2003).
Visto que a imunidade contra agentes infecciosos requer a ação coordenada de
componentes da imunidade inata e adaptativa, o estudo do recrutamento celular torna-se
relevante, uma vez que o número de células e a composição no estágio inicial depois do estímulo
13
VITORIANO-SOUZA, J.
Revisão da Literatura
influenciarão futuras respostas e o desenvolvimento da resposta imune adquirida (Teixeira et al.,
2005).
2.5 - Expressão de iNOS na pele
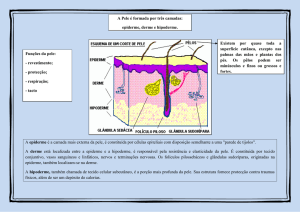
A pele, o maior órgão do corpo, é composta por três principais camadas: a epiderme, a
derme e camada subdermal ou hipoderme. Os principais tipos celulares presentes são
queratinócitos, melanócitos, células endoteliais, adipócitos e células de Langerhans, além de
glândulas sebáceas e sudoríparas. Dentre suas diversas funções, a pele atua como uma eficiente
barreira física, secretando substâncias microbicidas contra agentes agressores (Bos &
Kabsenberg, 1986), ela reage aos estímulos antigênicos disparando o processo inflamatório e
uma potente resposta de células T (Larregina et al., 2001).
Na LVC, a pele exerce uma função de destaque na infecção natural por Leishmania, visto
que se apresenta como local de entrada desse protozoário através da picada de fêmeas
hematófagas de flebotomíneos. Em cães susceptíveis a LVC, observa-se a disseminação do
parasito da pele através dos histiócitos (macrófagos do conjuntivo), células de Langerhans e
células dendríticas para diversos órgãos como baço, fígado, medula óssea, linfonodo, rim,
pulmões e trato digestório (Slappendel et al., 1998; Giunchetti et al., 2006). Além disso,
observa-se que a pele é um importante reservatório de parasitos em cães infectados por L.
chagasi ou L. infantum independente da forma clínica apresentada (Abranches et al., 1991).
Considerando a pele como um órgão extremamente susceptível a infecções de etiologias
diversas, a produção de NO parece ser um elemento chave particularmente no controle in situ de
patógenos intracelulares (Bogdan, 2001). Além da resposta inespecífica da defesa do hospedeiro,
a síntese de NO na pele é responsável por um papel chave na modulação da resposta a estímulos
externos tais como calor, raios ultravioletas, resposta a infecção através de citocinas e
cicatrização, bem como no desenvolvimento de certas condições patológicas (Cals-Grierson &
Ormerod, 2004).
O NO é uma molécula gasosa simples, habitualmente encontrada no ar atmosférico em
pequenas quantidades, altamente tóxica devido à presença de radical livre (elétron extra),
altamente reativa e difusível. Essa molécula é sintetizada pela enzima NOS (óxido nítrico
sintase) em dois passos de oxidação do aminoácido L-arginina produzindo em partes iguais Lcitrulina e NO (Miranda et al., 2000).
Atualmente são identificadas três principais isoformas de NOS, tais como: NOS1,
originalmente isolada do tecido neuronal (nNOS); NOS2 (iNOS), uma isoforma induzida e
14
VITORIANO-SOUZA, J.
Revisão da Literatura
NOS3 (eNOS), predominante em células endoteliais (Michel et al., 1997). Elas existem como
homodímeros com peso molecular entre 130 – 160 KDa e requerem alguns co-fatores para sua
função [flavina dinucleotídeo, flavina mononucleotídeo, tetrahidrobiopterina e nicotinamida
adenina dinucleotídeo reduzida (NADPH)]. Além disso, precisam estar ligadas a calmodulina, no
entanto, somente NOS1 e NOS3 requerem a ligação Ca2+-calmodulina, enquanto NOS2
apresenta uma menor dependência em relação ao cálcio (Nathan et al., 1994b).
As isoformas de NOS constitutivas são expressas em vários tipos celulares com fenótipos
maduros e podem ser moduladas por hormônios circulantes (Molero et al., 2002). A isoforma
NOS2 pode ser induzida em resposta a citocinas inflamatórias (IL-1, TNF-α e IFN-γ) ou em
combinação de citocinas e polissacarídeos de bactérias (LPS) (Stuehr et al., 1991). A síntese de
NO tem sido observada em macrófagos, neutrófilos, linfócitos T helper, fibroblastos, células
epiteliais, osteoclastos, osteoblastos, condrócitos, células endoteliais, células de Langerhans e,
recentemente, em mastócitos (Flora Filho et al., 2000), podendo ter vários efeitos, dependendo
do tipo de célula que o produz, do sítio de liberação e da concentração produzida (Moncada et
al., 1993). As funções do NO até hoje descobertas são complexas e antagônicas, podendo ser
benéfica ou potencialmente tóxica conforme a concentração ou depuração tecidual (Bogdan,
2001).
Dentre as principais funções do NO, pode-se citar a manutenção da homeostase tecidual,
incluindo vasodilatação, neurotransmissão e inibição da agregação e adesão plaquetárias
(Kendall et al., 2001). O NO também atua na resposta de defesa contra fungos, bactérias,
protozoários, bem como células tumorais. Essa molécula pode causar dano ao DNA e morte
celular via apoptose, induzindo fragmentação do DNA (Moncada et al., 2002).
Estudos de Qadoumi et al. (2002) oferecem evidências da expressão diferencial de iNOS
em humanos com leishmaniose cutânea (LC). Análises imuno-histoquímicas da pele revelaram
que a alta expressão da proteína iNOS em lesões clássicas de LC está relacionada ao número
pequeno de parasitos, enquanto que a baixa expressão de iNOs é acompanhada pelo aumento do
número de parasitos.
Considerando a importância do NO como mecanismo de controle de patógenos
intracelulares, aliado ao fato de ser induzido por citocinas importantes que auxiliam na
manutenção do microambiente de uma resposta do tipo 1, a exemplo do INF-γ, a avaliação deste
mediador na forma da iNOS torna-se altamente relevante para se predizer o potencial
imunoprotetor in locu. Neste sentido, o perfil de expressão da iNOS poderia indicar a capacidade
de diferentes estímulos teciduais em induzir uma resposta imune tipo 1, importante para
imunobiológicos candidatos a vacinas contra patógenos intracelulares.
15
3 - Justificativa
VITORIANO-SOUZA, J.
Justificativa
A busca por novos imunoprofiláticos contra LVC deve focalizar na prospecção de
antígenos capazes de intervir no sistema imune do cão, favorecendo o desenvolvimento de
eventos imunológicos específicos relacionados à eliminação do parasito, sendo inócuo ao
hospedeiro, protegendo contra a infecção por Leishmania spp. Dessa forma, a imunização deve
gerar uma resposta imune compatível ao perfil de reposta do tipo 1 que diminuia o risco de
transmissão do parasito. Para alcançar esses objetivos, novos protocolos têm sido desenvolvidos
utilizando o modelo canino e a utilização de novos antígenos e adjuvantes.
Neste contexto, o entendimento dos eventos iniciais da resposta imune ativada por
antígenos vacinais torna-se de grande importância no direcionamento para resposta imune
protetora no cão após a imunização com candidatos vacinais. Além disto, esta abordagem tornase uma importante estratégia de avaliação de candidatos vacinais, pois possibilita o
estabelecimento de biomarcadores para nortear as avaliações de alterações no local do inóculo
vacinal, que compreendem importantes alterações pós-vacinais.
Neste sentido, no intuito de ampliar os estudos sobre a resposta imune desencadeada pelo
imunobiológico contra LVC recentemente patenteado por pesquisadores do Laboratório de
Imunopatologia/NUPEB/UFOP em colaboração com o Laboratório de Imunologia Celular e
Molecular/IRR/FIOCRUZ, torna-se altamente relevante a avaliação no modelo canino da
resposta a este imunoprofilático e seus constituintes. Assim, pretende-se contribuir para a melhor
compreensão dos eventos iniciais pós-vacinais fundamentais para nortear o aprimoramento de
vacinas contra a LVC.
17
4 - Objetivos
VITORIANO-SOUZA, J.
Objetivos
4.1 - Objetivo Geral:
•
Avaliar a cinética de migração celular e a expressão da enzima iNOs na pele de cães
frente a componentes vacinais (L. braziliensis, extrato de glândula salivar de Lu.
longipalpis, saponina e L. braziliensis associado a saponina) de uma nova vacina contra
Leishmaniose Visceral Canina.
4.2 - Objetivos específicos:
Nos intervalos de tempo de 1, 12, 24, 48 e 96 horas após o inóculo:
•
Avaliar as alterações macroscópicas decorrentes dos inóculos intradérmicos dos
diferentes componentes vacinais;
•
Avaliar o estabelecimento do processo inflamatório que se desenvolve na pele de cães
após a imunização intradérmica dos diferentes componentes vacinais;
•
Quantificar as populações celulares mais freqüentes encontradas no local do inóculo
intradérmico dos diferentes componentes vacinais;
•
Avaliar a expressão da enzima óxido nítrico sintase induzida (iNOS) no local do inóculo
intradérmico dos diferentes componentes vacinais.
19
5 - Material e Métodos
VITORIANO-SOUZA, J.
Material e Métodos
5.1 - Delineamento
Nessa primeira figura estão representadas de forma esquemática as atividades realizadas
nesse trabalho.
15 cães
CS
SAP
Sal
LB
LBSAP
Biópsias da pele de cães após
inóculos
Cinética de migração celular
1
12
24
48
96 horas
Processamento, inclusão
em parafina e microtomia (4-5 µm)
Contagem
Diferencial (%)
IMH para detecção
de iNOS
HE
Análises Subjetiva e Morfométrica
Figura 1: Delineamento experimental ilustrando as atividades realizadas no presente estudo.
21
VITORIANO-SOUZA, J.
Material e Métodos
5.2 - Animais
Foram utilizados 15 cães sem raça definida (Canis familiaris) de ambos os sexos, com
idade entre 8 a 12 meses, provenientes da colônia de cães do canil do Biotério Central da
Universidade Federal de Ouro Preto (UFOP). Os cães foram vermifugados a partir do 21° dia de
nascimento e receberam mais duas doses de antihelmínticos com intervalos de 21 dias. A
primeira dose da vacina contra cinomose, adenovírus tipo 2, coronavírus, parainfluenza,
parvovírus e leptospirose canina (Vanguard ® HTLP 5/CV-L, Pfizer Animal Health, New York,
NY, USA) foi aplicada quando os cães atingiram 45 dias de idade, as outras duas doses foram
dadas em intervalos de 21 dias. Antes dos experimentos, todos os animais foram tosados e não
foram observadas reações alérgicas. Foram utilizados três animais em cada grupo.
5.3 - Antígenos e adjuvantes vacinais
O imunobiológico foi preparado a partir de antígeno de L. (Viannia) braziliensis (cepa
MHOM/BR/75/M2903). Esta vacina foi recentemente patenteada pela UFOP em parceria com
pesquisadores do Instituto de Pesquisas René Rachou IRR-FIOCRUZ/ MG (Instituto Nacional
de Propriedade Industrial – patente PI 0601225-6, 17 de fevereiro de 2006). A concentração do
antígeno vacinal (LB) foi de 600 µg/mL diluído em solução salina estéril 0,9%.
O adjuvante saponina (Sigma Chemical Co, St. Louis, MO, U.S.A.) foi diluído em
solução salina estéril 0,9% (1mg/dose) no momento do inóculo para evitar perda na estabilidade.
A formulação de LBSap foi realizada através da associação do antígeno vacinal (600 µg/mL) e
do adjuvante saponina (1mg/dose) diluídos em salina estéril 0,9%.
Utilizou-se o extrato de glândula salivar de Lu. longipalpis (1 ácino/dose) cedida pelo
Laboratório de Entomologia/ICB/UFMG, a partir de colônia vinda de Teresina (Piauí) e
Abaetetuba (Pará) obtidas conforme protocolo estabelecido por Cavalcante et al. (2003).
Os antígenos foram inoculados intradermicamente na região dorsal dos cães em duplicata
para observar possíveis variações. Os inóculos foram realizados através de injeções de 250 µL
(volume total) com os diferentes componentes vacinais. Antes do procedimento, a área foi
demarcada com caneta para delimitar o local de biópsia.
Cada animal, após ser anestesiado (Thiopentax®, São Paulo, Brazil; 7 mg/kg de peso
corporal), recebeu em seu dorso os estímulos de todos os componentes antigênicos vacinais:
saponina (Sap), antígeno vacinal (LB), antígeno vacinal associado a saponina (LBSap) e saliva
de Lu. longipalpis (Sal), sendo que entre esses foram inoculados controles salina (CS) para
detectar possível extravasamento da reação inflamatória. A distância entre cada inóculo foi de
22
VITORIANO-SOUZA, J.
Material e Métodos
aproximadamente 3 cm. Após os tempos de 1, 12, 24, 48 e 96 horas pós-imunização (p.i.) foram
realizadas biópsias de pele para a avaliação da cinética de migração celular provocada pelos
diferentes componentes vacinais. Também foi observada antes de cada biópsia a presença de
alterações no local do inóculo provocadas pelos componentes antigênicos em cada tempo. Todas
as alterações foram fotografadas.
5.4 - Grupos Experimentais
Grupo Adjuvante: Saponina (Sap);
Grupo Saliva: Extrato de glândula salivar de Lu. longipalpis (Sal);
Grupo Antígeno vacinal: Leishmania braziliensis (LB);
Grupo Vacina: Antígeno vacinal de L. braziliensis associado ao adjuvante saponina (LBSap);
Grupo Controle: Salina estéril 0,9% (CS);
Utilizou-se biópsias de pele normal sem estímulo de cada animal, a fim de observarmos
possíveis variações no controle salina.
5.5 - Processamento do material
As biópsias foram realizadas no Biotério da Universidade Federal de Ouro Preto, onde
foram retirados os fragmentos de pele, previamente demarcados, das regiões dos diferentes
inóculos (CS, Sap, Sal, LB, LBSap) além de fragmento de pele normal sem estímulo (CN).
O material coletado foi divido em três amostras, sendo duas amostras congeladas (freezer
-70° C), para posteriores experimentos de PCR e imuno-histoquímica (IMH). Nas amostras
coletadas para IMH foi acrescentado OCT (Optimal Cutting Temperature Médium, Tissue Tek,
Leica Microsystem Nussloch GmbH, Wehrheim, Germany).
A terceira amostra foi fixada em formol tamponado (pH 7,2), processada no histotécnico
(OM CM 69, Metal. OMA LTDA, São Paulo, Brasil), incluída em parafina e cortada em
micrótomo na espessura de cinco µm. Estes procedimentos operacionais são padrões do
laboratório de Imunopatologia do Núcleo de Pesquisas em Ciências Biológicas (NUPEB/UFOP),
seguindo as normas de controle de qualidade do laboratório.
Desse material, retiraram-se dois cortes histológicos de cada bloco que foram depositados
separadamente em lâminas de vidro, sendo uma corada pelo método da Hematoxilina & Eosina
(HE) e a outra submetida à IMH para detecção da expressão da enzima iNOS.
23
VITORIANO-SOUZA, J.
Material e Métodos
5.6 - Imuno-histoquímica para detecção da expressão da enzima iNOS
Para a realização da IMH anti-iNOs, inicialmente os cortes foram desparafinizados em
xilol (dois banhos de 15 minutos cada) e hidratados em concentrações decrescentes de álcoois
(100, 90, 80 e 70%) em banhos de cinco minutos cada. Em seguida, os cortes foram lavados em
água corrente por cinco minutos.
Após essa etapa, realizou-se o bloqueio da peroxidase endógena, onde os cortes foram
incubados por 30 minutos a temperatura ambiente em uma solução de peróxido de hidrogênio
(180 mL de álcool metílico + 20mL de peróxido de oxigênio 30%). Seguiu-se a lavagem em PBS
por três banhos de cinco minutos cada.
A recuperação antigênica dessa reação foi realizada através de solução citrato pH 6,0 por
20 minutos em forno de microondas. Depois de retiradas do forno de microondas, ainda na
solução citrato, as lâminas foram deixadas em temperatura ambiente até seu resfriamento e logo
após, seguiu-se três banhos de PBS de cinco minutos cada.
As bordas das lâminas foram secas e aplicou-se o soro normal de cavalo (R.T.U.
Vectastain Universal Elite ABC; Vector Laboratories, Inc., Burlingame, CA, USA). Essas
lâminas foram incubadas por 30 minutos em câmara úmida a 37º C, com o objetivo de bloquear
sítios antigênicos inespecíficos.
Após o término do bloqueio, secou-se o excesso de soro normal de cavalo e os cortes
foram incubados por 12hs à 18hs em câmara úmida a 4°C com o anticorpo policlonal IgG de
coelho anti-iNOS (Rabbit anti-Mouse iNOS, SC-651, Santa Cruz Biotechnology, INC., CA,
USA) diluído 1:200 em solução diluente de anticorpo com componentes redutores de
background (DakoCytomation, Via Real, Carpinteria, USA).
Após o período de incubação, seguiu-se a lavagem em três banhos de PBS por cinco
minutos cada e novamente, os cortes foram incubados com o anticorpo biotinilado (R.T.U.
Vectastain Universal Elite ABC; Vector Laboratories, Inc., Burlingame, CA, USA) por 30
minutos a 37ºC. Após a incubação com o anticorpo biotinilado, procedeu-se a lavagem das
lâminas com três banhos de PBS de cinco minutos cada.
Os cortes foram então incubados com o complexo ABC (R.T.U. Vectastain Universal
Elite ABC; Vector Laboratories, Inc., Burlingame, CA, USA) por 30 minutos em câmara úmida
a 37°C. Seguiu-se a lavagem em PBS, três banhos de cinco minutos cada.
A revelação da reação foi obtida através da incubação das lâminas em solução de DAB
(50mg de Diaminobenzidina em 250 mL de PBS e 500 µL de Peróxido de Hidrogênio 30%) por
cinco minutos.
24
VITORIANO-SOUZA, J.
Material e Métodos
A reação foi interrompida através da lavagem dos cortes em água corrente por cinco
minutos. Para realizar a contra-coloração, utilizou-se a Hematoxilina de Harris por 10 segundos à
temperatura ambiente seguida pela lavagem dos cortes, por cinco minutos, em água corrente.
O material foi desidratado em álcool absoluto e seco em estufa 56°C. Após a secagem
dos cortes na estufa, as lâminas foram montadas com Bálsamo do Canadá (Pró-Cito) e lamínula
para posteriores análises.
Estes procedimentos operacionais são padrões do laboratório de Imunopatologia do
Núcleo de Pesquisas em Ciências Biológicas (NUPEB/UFOP), seguindo as normas de controle
de qualidade do laboratório.
5.7 - Análises Histopatológicas
As lâminas coradas por HE e submetidas à reação de imuno-histoquímica foram
avaliadas no microscópio óptico, quanto à intensidade do infiltrado inflamatório, bem como às
células inflamatórias predominantes e sua distribuição na derme superior, derme profunda e na
hipoderme. Avaliou-se também a presença de células iNOS positivas, sua localização, sua
distribuição e intensidade de marcação. A avaliação de intensidade foi realizada utilizando-se os
seguintes parâmetros: (-) ausente, (+) discreto, (++) moderado e (+++) intenso.
5.8 - Análises quantitativas
5.8.1 - Contagem diferencial (%)
Foi realizada contagem diferencial (em %) das principais células inflamatórias
(macrófagos, linfócitos, neutrófilos e eosinófilos) na pele de cães imunizados com diversos
antígenos.
A pele dos cães foi subdividida em derme superior, derme profunda e hipoderme a fim de
melhor visualizar a migração dos diversos tipos celulares entre as diferentes camadas da pele.
5.8.2 - Análise quantitativa do infiltrado celular na derme e hipoderme
As células inflamatórias presentes na derme (superior e profunda) e hipoderme foram
quantificadas através da aquisição de 20 imagens aleatórias (área total percorrida igual a 1,5
x 106 µm2) de cada camada. As imagens visualizadas pela objetiva 40x foram digitalizadas
25
6 - Resultados
VITORIANO-SOUZA, J.
Resultados
6.1 - Análises Anátomo e Histopatológicas
Na análise macroscópica não se notou nenhuma reação adversa nos animais imunizados, com
exceção de edema com induração local na pele de cães dos grupos inoculados com o adjuvante
saponina (Sap e LBSap) a partir de 12 horas pós-imunização (p.i.). Essa alteração manteve-se
durante toda a cinética, não resultando em lesões ulceradas. Nos demais grupos não houve
notificação de qualquer alteração.
Durante o recorte do material para biopsia, observou-se que o local da pele inoculado com
Sap e LBSap apresentava-se edemaciado, hiperemiado com focos hemorrágicos no tempo de 12
horas. Nos tempos seguintes, em alguns animais verificou-se necrose na região da pele de cães
desses mesmos grupos. Não se observou qualquer alteração nas demais regiões inoculadas com os
outros antígenos (Figura 2).
As principais alterações histopatológicas observadas na pele de cães imunizados com
diferentes componentes antigênicos foram edema, congestão, hemorragia e degeneração nas fibras
musculares quando associada a intensa inflamação focal. Estas observações foram freqüentes em
toda a cinética, dos tempos iniciais (1 e 12 horas p.i) até o mais tardio (96 horas p.i.) nos grupos
inoculados com Sap e LBSap. No grupo Sal e LB observaram-se apenas discreta hemorragia nos
tempos iniciais (12 e 24 horas p.i.) da cinética.
28
VITORIANO-SOUZA, J.
LBSap
Resultados
CS
Sal
CS
Sap
CN
LB
CS
CS
LB
A
CN
Sap
CS
Sal
CS LBSap
B
C
D
E
LBSap
Sap
F
Sap
LBSap
G
H
1,86 cm
CS
Figura 2: Alterações macroscópicas na pele de cães inoculados intradermicamente
com diferentes componentes antigênicos. Em A, está representada a distribuição dos
componentes vacinais no dorso do animal; em B e C, inóculo intradérmico; em D,
são mostradas as alterações macroscópicas nos grupos Sap e LBSap; em E, está
representada a fotomicrografia da pele invertida, onde observam-se as reações
provocadas pelos grupos Sap e LBSap; em F, G e H, são representados os grupos Cs,
Sap e LBSap, demonstrando edema e focos hemorrágicos nos grupos Sap e LBSap,
além de uma região com presença de exsudato purulento em LBSap (H).
29
VITORIANO-SOUZA, J.
Resultados
6.2 - Cinética de migração celular na derme de cães
A cinética de migração celular na derme de cães está representada no Gráfico 1 e Figura 3.
Nessa cinética não foram observadas diferenças significativas entre os grupos analisados em relação
aos diferentes tempos.
Foi observado na derme aumento (P<0.05) do infiltrado celular após imunização com Sap
nos tempos de 1 (130,8±30,1),12 (187,2±38,9), 24 (154±32,81) e 96 horas p.i. (254,8±57,34) em
relação ao grupo CS (57±12,15; 58,62±13,24; 44,07±2,917 e 40,35±5,798), respectivamente; e em
12 (187,2±38,9) e 24 (154±32,81)
e 96 horas p.i. (254,8±57,34) em relação ao grupo Sal
(82,7±21,48; 68,35±16,13 e 35±21,21). Adicionalmente, em 96 horas p.i. observou-se aumento
(P<0.05) do infiltrado em Sap (254,8±57,34) em relação a LB (35±5,162). No tempo de 24 horas
p.i., observou-se ainda aumento (P<0.05) do infiltrado no grupo LB (119,3±43,53) em relação a CS
(44,07±2,917).
400
Número de núcleos
200
0
1h
400
1h
12h
24h
48h
24h
12 h
96h
48h
96h
200
0
CS
Sap
Sal
LB
LBSap
Tempo
Gráfico 1: Cinética do infiltrado celular na derme de cães inoculados com diferentes componentes: CS
(controle salina), Sap (saponina), Sal (extrato de glândula salivar de Lu. longipalpis), LB (antígeno de
L. braziliensis) e LBSap (antígeno de L. braziliensis associado a saponina). As análises foram realizadas
1, 12, 24, 48 e 96 horas p.i. Os resultados estão expressos como média ± desvio padrão. As diferenças
significativas (P<0.05) da análise comparativa entre os grupos estão demonstradas pelas linhas.
30
VITORIANO-SOUZA, J.
Resultados
24 h
96 h
Controle
1h
B
C
D
E
F
G
H
I
LBSap
Saponina
A
50 µm
Figura 3: Fotomicrografia do infiltrado celular da derme de cães imunizados com saponina,
LBSap e CS. Nessa figura está representado o infiltrado celular discreto observado nestes
grupos nos tempos de 1, 24 e 96 horas p.i (aumento de 40x, barra: 50 µm).
31
VITORIANO-SOUZA, J.
Resultados
6.3 - Cinética de migração celular seletiva na derme superior de cães
6.3.1 - Migração de neutrófilos na derme superior de cães
Buscando-se ampliar o estudo do infiltrado inflamatório para se determinar os tipos celulares
que seriam recrutados para o local do inóculo pelos componentes antigênicos da vacina LBSap foi
realizada a contagem diferencial de células na derme profunda e hipoderme, sendo os resultados
expressos em percentual.
No Gráfico 2, estão demonstrados os resultados da cinética de neutrófilos na derme superior,
bem como as comparações entre os grupos estudados. Em relação a cinética, o grupo Sap induziu
aumento (P<0.05) de neutrófilos no tempo de 12 horas p.i. (42±8,485) em relação ao tempo inicial
de 1 hora p.i. (6±0). Por outro lado, nos tempos de 48 (9,5±2,121) e 96 horas p.i. (9±12,73), o grupo
Sap apresentou redução (P<0.05) neutrófilos quando comparado ao tempo de 12 horas p.i.
(42±8,485).
A avaliação entre os diferentes inóculos considerando a população de neutrófilos na derme
superior, revelou aumento (P<0.05) no grupo Sap (42±8,485) quando comparado aos grupos CS
(4,5±6,364), Sal (6,5±0,7071), LB (13,67±10,21) e LBSap (6,667±3,215) no tempo de 12 horas p.i.
32
VITORIANO-SOUZA, J.
Resultados
100
Percentual de neutrófilos (%)
50
a
b
b
0
1h
100
1h
12h
24h
48h
24 h
12 h
96h
48 h
96 h
50
0
CS
Sap
Sal
LB
LBSap
Tempo
Gráfico 2: Cinética de migração seletiva de neutrófilos na derme superior de cães inoculados com
diferentes componentes: CS (controle salina), Sap (saponina), Sal (extrato de glândula salivar de Lu.
longipalpis), LB (antígeno de L. braziliensis) e LBSap (antígeno de L. braziliensis associado a
saponina). As análises foram realizadas 1, 12, 24, 48 e 96 horas p.i. No Gráfico da cinética, as
diferenças estatísticas estão representadas pelas letras “a” e “b”, referentes aos tempos de 1 e 12 horas
p.i, respectivamente. Os resultados estão expressos como média ± desvio padrão. As diferenças
significativas (P<0.05) da análise comparativa entre os grupos estão demonstradas pelas linhas.
6.3.2 - Migração de macrófagos na derme superior de cães
No Gráfico 3, estão representados os resultados de cinética de macrófagos na derme superior,
bem como as comparações entre os grupos estudados.
A avaliação da cinética de migração de macrófagos indicou redução no tempo de 12 horas
p.i. (47,67±2,082) no grupo imunizado com Sap em relação ao tempo de 1 hora p.i. (82,33±4,041).
Na avaliação entre os diferentes inóculos foi observado queda no número de macrófagos do
grupo Sap (47,67±2,082) quando comparado ao CS (85±7,071) no tempo de 12 horas p.i. De forma
similar, foi observada redução de macrófagos no grupo LBSap (43,67±4,726) em 48 horas p.i. em
relação aos grupos CS (71,33±11,55), Sal (68,67±10,97) e LB (83,33±4,619) (P<0.05).
33
VITORIANO-SOUZA, J.
Resultados
Percentual de macrófagos (%)
100
50
a
0
1h
100
1h
12h
24h
12 h
48h
24 h
96h
48 h
96 h
50
0
CS
Sap
Sal
LB
LBSap
Tempo
Gráfico 3: Cinética de migração seletiva de macrófagos na derme superior de cães inoculados com
diferentes componentes: CS (controle salina), Sap (saponina), Sal (extrato de glândula salivar de Lu.
longipalpis), LB (antígeno de L. braziliensis) e LBSap (antígeno de L. braziliensis associado a
saponina). As análises foram realizadas 1, 12, 24, 48 e 96 horas p.i. Os resultados estão expressos
como média ± desvio padrão. No Gráfico de cinética, a diferença estatística está representada pela
letra “a” referente ao tempo de 1 hora. As diferenças significativas (P<0.05) da análise comparativa
entre os grupos estão demonstradas pelas linhas.
6.3.3 - Migração de eosinófilos na derme superior de cães
No Gráfico 4, estão demonstrados os resultados de cinética de eosinófilos na derme superior,
bem como as comparações entre os grupos estudados.
Na avaliação da cinética de migração de eosinófilos observou-se aumento significativo
(P<0.05) desse tipo celular no tempo de 96 horas p.i. (5±0) em relação aos tempos de 12 (0±0), 24
(0±0) e 48 horas p.i. (0,3333±0,5774) no grupo LBSap. A análise estatística entre os grupos não
revelou diferença significativa.
34
VITORIANO-SOUZA, J.
Resultados
Percentual de eosinófilos (%)
10
b,c,d
5
0
1h
10
1h
12h
24h
48h
24 h
12 h
96h
48 h
96 h
5
0
CS
Sap
Sal
LB
LBSap
Tempo
Gráfico 4: Cinética de migração seletiva de eosinófilos na derme superior de cães inoculados com
diferentes componentes: CS (controle salina), Sap (saponina), Sal (extrato de glândula salivar de Lu.
longipalpis), LB (antígeno de L. braziliensis) e LBSap (antígeno de L. braziliensis associado a
saponina). As análises foram realizadas 1, 12, 24, 48 e 96 horas p.i. Os resultados estão expressos
como média ± desvio padrão. No Gráfico de cinética, as diferenças estatísticas estão representadas
pelas letras “b”, “c” e “d”, referentes aos temos de 12, 24 e 48 horas p.i. respectivamente.
6.3.4 - Migração de linfócitos na derme superior de cães
Na avaliação da cinética de migração de linfócitos na derme superior observou-se aumento
(P<0.05) de linfócitos no grupo LBSap no tempo de 48 horas p.i. (40,67±6,11) em relação ao tempo
de 1 hora (9,333±0,5774). Nas cinéticas dos demais grupos, não foram observados diferenças como
demonstrado no Gráfico 5.
Adicionalmente, a avaliação considerando a comparação entre os grupos revelou aumento
(P<0.05) de linfócitos nos grupos Sap (24,67±3,786) e LBSap (24,67±4,041) no tempo de 24 horas
em relação ao CS (13,67±3,512), Sal (7,667±1,528) e LB (10,67±4,726). No tempo de 48 horas,
observou-se um aumento (P<0.05) do número de linfócitos no grupo LBSap (40,67±6,11) quando
comparado aos grupos CS (16,67±4,509) e LB (12,33±1,528).
35
VITORIANO-SOUZA, J.
Resultados
Percentual de linfócitos (%)
100
50
a
0
1h
100
1h
12h
24h
48h
24 h
12 h
96h
48 h
96 h
50
0
CS
Sap
Sal
LB
LBSap
Tempo
Gráfico 5: Cinética de migração seletiva de linfócitos na derme superior de cães inoculados com
diferentes componentes: CS (controle salina), Sap (saponina), Sal (extrato de glândula salivar de Lu.
longipalpis), LB (antígeno de L. braziliensis) e LBSap (antígeno de L. braziliensis associado a
saponina). As análises foram realizadas 1, 12, 24, 48 e 96 horas p.i. Os resultados estão expressos
como média ± desvio padrão. No Gráfico de cinética, a diferença estatística está representada pela letra
“a” referente ao tempo de 1 hora. As diferenças significativas (P<0.05) da análise comparativa entre os
grupos estão demonstradas pelas linhas.
6.4 - Cinética de migração celular seletiva na derme profunda de cães
6.4.1 - Migração de neutrófilos na derme profunda de cães
No Gráfico 6, estão demonstrados os resultados de cinética de neutrófilos na derme profunda,
bem como as comparações entre os grupos estudados.
Ao longo da cinética não houve diferença significativa em nenhum grupo estudado. Na
análise comparativa entre os grupos, notou-se aumento (P<0.05) de neutrófilos no grupo Sap
(38,33±11,59) em 48 horas p.i. em relação ao grupo LB (3±1,732).
36
VITORIANO-SOUZA, J.
Resultados
100
Percentual de neutrófilos (%)
50
0
1h
100
1h
12h
24h
12 h
48h
24 h
96h
48 h
96 h
50
0
CS
Sap
Sal
LB
LBSap
Tempo
Gráfico 6: Cinética de migração seletiva de neutrófilos na derme profunda de cães inoculados com
diferentes componentes: CS (controle salina), Sap (saponina), Sal (extrato de glândula salivar de Lu.
longipalpis), LB (antígeno de L. braziliensis) e LBSap (antígeno de L. braziliensis associado a
saponina). As análises foram realizadas 1, 12, 24, 48 e 96 horas p.i. Os resultados estão expressos
como média ± desvio padrão As diferenças significativas (P<0.05) da análise comparativa entre os
grupos estão demonstradas pelas linhas.
6.4.2 - Migração de macrófagos na derme profunda de cães
A cinética de migração de macrófagos na derme profunda, bem como as comparações entre
os grupos está representada no Gráfico 7.
Em relação a cinética, nenhuma diferença estatística entre os grupos ao longo do tempo foi
observada. A análise comparativa entre grupos mostrou redução (P<0.05) do percentual de
macrófagos no grupo LBSap (17,67±10,02 e 38,67±7,767) quando comparado ao CS (77,67±8,145 e
77,33±4,509) nos tempos de 24 e 48 horas p.i., respectivamente. Adicionalmente, houve redução
significativa em 48 horas p.i. do percentual de macrófagos no grupo LBSap (38,67±7,767) quando
comparado ao grupo LB (80,67±9,452). Também, foi verificado no grupo Sap (48,33±11,68)
redução (P<0.05) de macrófagos quando comparados aos grupos CS (77,33±4,509) em 48 horas p.i.
37
VITORIANO-SOUZA, J.
Resultados
100
Percentual de macrófagos (%)
50
0
1h
100
1h
12h
24h
12 h
48h
96h
48 h
24 h
96 h
50
0
CS
Sap
Sal
LB
LBSap
Tempo
Gráfico 7: Cinética de migração seletiva de macrófagos na derme profunda de cães inoculados com
diferentes componentes: CS (controle salina), Sap (saponina), Sal (extrato de glândula salivar de Lu.
longipalpis), LB (antígeno de L. braziliensis) e LBSap (antígeno de L. braziliensis associado a
saponina). As análises foram realizadas 1, 12, 24, 48 e 96 horas p.i. Os resultados estão expressos
como média ± desvio padrão. As diferenças significativas (P<0.05) da análise comparativa entre os
grupos estão demonstradas pelas linhas.
6.4.3 - Migração de eosinófilos na derme profunda de cães
No Gráfico 8, está representada a avaliação da cinética de migração de eosinófilos onde se
observa aumento (P<0.05) desse tipo celular no tempo de 96 horas p.i. (21±5,657) no grupo Sap em
relação a todos os tempos anteriores: 1 (0,3333±0,5774), 12 (0±0), 24 (0,3333±0,5774) e 48 horas
(2,667±1,155). De forma similar, neste mesmo tempo, também foi observado aumento (P<0.05) de
eosinófilos no grupo LBSap no tempo de 96 horas p.i. (23,5±21,92) em relação ao tempo de 12
(0±0) e 24 horas p.i. (0,3333±0,5774). Na comparação entre os grupos não foram observadas
diferenças significativas no número de eosinófilos ao longo dos tempos.
Na derme profunda, não foram observadas diferenças ao longo dos tempos e nem entre os
grupos avaliados, quanto à migração de linfócitos.
38
VITORIANO-SOUZA, J.
Resultados
40
b,c
a,b,c,d
Percentual de eosinófilos (%)
20
0
1h
1h
40
12h
24h
12 h
48h
96h
48 h
24 h
96 h
20
0
CS
Sap
Sal
LB
LBSap
Tempo
Gráfico 8: Cinética de migração seletiva de eosinófilos na derme profunda de cães inoculados com
diferentes componentes: CS (controle salina), Sap (saponina), Sal (extrato de glândula salivar de Lu.
longipalpis), LB (antígeno de L. braziliensis) e LBSap (antígeno de L. braziliensis associado a
saponina). As análises foram realizadas 1, 12, 24, 48 e 96 horas p.i. Os resultados estão expressos
como média ± desvio padrão. No Gráfico de cinética, as diferenças estatísticas estão representadas
pelas letras “a”, “b”, “c” e “d”, referentes aos tempos de 1, 12, 24 e 48 horas p.i, respectivamente.
6.5 - Cinética de migração celular na hipoderme de cães
Os principais resultados do infiltrado celular total na hipoderme de cães imunizados com
diferentes componentes antigênicos estão representados no Gráfico 9 e Figura 4. Na análise da
cinética foi observado aumento (P<0.05) do infiltrado celular nos grupos Sap (791,4±80,63;
994,6±118,5 e 1098±268,4) e LBSap (529±120,3; 711,7±124 e 598,4±217,1) a partir de 12 horas até
48 horas p.i. quando comparado ao tempo inicial de 1 hora (160,1±14,21- Sap e 77,07±22,45LBSap).
Na análise comparativa do infiltrado celular entre os grupos foi observado aumento (P<0.05)
do infiltrado celular nos grupos Sap (791,4±80,63; 994,6±118,5 e 1098±268,4) e LBSap (529±120,3;
711,7±124 e 598,4±217,1)
nos tempos de 12, 24 e 48 horas quando comparados com CS
(65,53±21,59; 60,43±25,41 e 49,03±14,09), Sal (79,75±45,27; 93,17±63,99 e 64,83±10,88) e LB
(248,4±81,62; 210,4±123,6 e 94,85±32,6). Além disto, o grupo Sap (791,4±80,63; 994,6±118,5 e
1098±268,4) apresentou aumento significativo do infiltrado celular nos mesmos tempos de 12, 24 e
39
VITORIANO-SOUZA, J.
Resultados
48 horas quando comparado ao grupo LBSap (529±120,3; 711,7±124 e 598,4±217,1).
Adicionalmente, no tempo de 1, o grupo Sap (160,1±14,21) apresentou aumento do infiltrado celular
em relação aos grupos CS (39,82±8,476), e em 96 horas, aumento de Sap (596,2±271,5) em relação
a CS (41,13±5,197) e Sal(36,6±11,38), respectivamente.
1500
a
a
1000
a
a
a
a
Número de núcleos
500
0
1h
1500
1h
12h
24h
48h
48h
24h
12h
96h
96h
1000
500
0
CS
Sap
Sal
LB
LBSap
Tempo
Gráfico 9: Cinética de migração celular na hipoderme de cães inoculados com diferentes
componentes: CS (controle salina), Sap (saponina), Sal (extrato de glândula salivar de Lu.
longipalpis), LB (antígeno de L. braziliensis) e LBSap (antígeno de L. braziliensis associado a
saponina). As análises foram realizadas 1, 12, 24, 48 e 96 horas p.i. Os resultados estão expressos
como média ± desvio padrão. No Gráfico de cinética, a diferença estatística está representada pela
letra “a” referente ao tempo de 1 hora. As diferenças significativas (P<0.05) da análise comparativa
entre os grupos estão demonstradas pelas linhas.
40
VITORIANO-SOUZA, J.
Resultados
24 h
96 h
Controle
1h
B
C
D
E
F
G
H
I
LBSap
Saponina
A
50 µm
Figura 4: Fotomicrografia do infiltrado celular da hipoderme de cães imunizados com
saponina, LBSap e CS. Nessa figura está representado o infiltrado celular intenso observado
nestes grupos quando comparado ao controle nos tempos de 1, 24 e 96 horas p.i (aumento de
40x, barra: 50 µm).
41
VITORIANO-SOUZA, J.
Resultados
6.5 - Cinética de migração celular seletiva na hipoderme de cães
6.5.1 - Migração de neutrófilos na hipoderme de cães
No Gráfico 10, estão representados os resultados referentes a cinética de neutrófilos na
hipoderme.
A hipoderme foi o terceiro compartimento cutâneo avaliado. Neste sentido, a avaliação
cinética de neutrófilos revelou redução (P<0.05) do percentual de neutrófilos no grupo LBSap, no
tempo de 96 horas p.i. (35±11,31) quando comparado aos tempos de 1 (67,33±4,619) e 24 horas p.i.
(66,67±5,508). Adicionalmente, foi observado no grupo LB redução (P<0.05) de neutrófilos nos
tempos de 48 (16,67±9,074) e 96 horas p.i. (19,5±9,192) em relação aos tempos de 1 (51,33±13,65),
12 (64,67±1,155) e 24 horas p.i. (66,67±6,658).
Na análise comparativa entre grupos foi observado aumento (P<0.05) de neutrófilos nos
grupos Sap (56,67±29,67) e LBSap (67,33±4,619) em relação ao CS (11,33±4,041), no tempo inicial
de 1 hora p.i., similarmente, foi observado no tempo de 24 horas aumento (P<0.05) de neutrófilos
nos grupos Sap (76,67±6,658), LB (66,67±6,658) e LBSap (66,67±5,508) quando comparado aos
grupos CS (29±11,31) e Sal (37,33±16,5). No tempo de 48 horas foi observado aumento (P<0.05) de
neutrófilos nos grupos Sap (65,67±4,041) e LBSap (48,67±3,512) quando comparado aos grupos CS
(22,33±2,517), Sal (22±11,53) e LB (16,67±9,074).
42
VITORIANO-SOUZA, J.
Resultados
100
a
Percentual de neutrófilos (%)
50
a,c
b
a,b,c
b
a,b,c
48h
96h
0
1h
1h
100
12h
24h
12 h
24 h
48 h
96 h
50
0
CS
Sap
Sal
LB
LBSap
Tempo
Gráfico 10: Cinética de migração de neutrófilos na hipoderme de cães inoculados com diferentes
componentes: CS (controle salina), Sap (saponina), Sal (extrato de glândula salivar de Lu. longipalpis),
LB (antígeno de L. braziliensis) e LBSap (antígeno de L. braziliensis associado a saponina). As
análises foram realizadas 1, 12, 24, 48 e 96 horas p.i. Os resultados estão expressos como média ±
desvio padrão No Gráfico de cinética, as diferenças estatísticas estão representadas pelas letras “a”, “b”
e “c”, referentes aos temos de 1, 12 e 24 horas p.i., respectivamente. As diferenças significativas
(P<0.05) da análise comparativa entre os grupos estão demonstradas pelas linhas.
6.5.3 - Migração de macrófagos na hipoderme de cães
No Gráfico 11, estão representados os resultados referentes a cinética de macrófagos na
hipoderme.
Na cinética de migração de macrófagos, notou-se redução significativa (P<0.05) desta
população celular no grupo CS (39±22,72) em 12 horas p.i. em relação ao tempo de 1 hora p.i.
(74,33±2,309) No grupo LB, no entanto verificou-se aumento significativo (P<0,05) de macrófagos
no tempo de 48 horas p.i. (66±11) em relação aos tempos de 12 (25,67±5,859) e 24 horas p.i.
(27±5,196).
A análise comparativa entre grupos mostrou redução (P<0.05) do percentual de macrófagos
no tempo de 1 hora considerando os grupos Sap (18,33±5,033) e LBSap (19,33±7,024) quando
comparados aos CS (74,33±2,309) e Sal (52,33±14,74), e de forma similar no grupo LB (39±8,544)
em relação ao grupo CS (74,33±2,309). No tempo de 24 horas p.i., observou-se redução de
43
VITORIANO-SOUZA, J.
Resultados
macrófagos nos grupos Sap (14,67±6,658) e LBSap (15,33±6,028) em relação ao grupo CS
(57,33±9,292). Subseqüentemente, no tempo de 48 horas foi observada redução (P<0.05) de
macrófagos nos grupos LBSap (26±2) e Sap (8,333±4,163) em relação aos grupos CS
(66,67±3,215), Sal (60,33±23,44) e LB (66±11).
100
b,c
Percentual de macrófagos (%)
50
a
0
1h
100
1h
12h
24h
12 h
48h
24 h
96h
48 h
96 h
50
0
CS
Sap
Sal
LB
LBSap
Tempo
Gráfico 11: Cinética de migração de macrófago na hipoderme de cães inoculados com diferentes
componentes antigênicos: CS (controle salina), Sap (saponina), Sal (extrato de glândula salivar de Lu.
longipalpis), LB (antígeno de L. braziliensis) e LBSap (antígeno de L. braziliensis associado a
saponina). As análises foram realizadas 1, 12, 24, 48 e 96 horas p.i. Os resultados estão expressos
como média ± desvio padrão. No Gráfico de cinética, as diferenças estatísticas estão representadas
pelas letras “a”, “b” e “c” referentes aos temos de 1, 12 e 24 horas p.i., respectivamente. As diferenças
significativas (P<0.05) da análise comparativa entre os grupos estão demonstradas pelas linhas.
6.5.4 - Migração de eosinófilos na hipoderme de cães
No Gráfico 12 estão representados os resultados referentes a cinética de eosinófilos na
hipoderme. Observou-se aumento de eosinófilos (P<0.05) no grupo Sap nos tempos finais de 48
(7±2,646) e 96 horas p.i. (13±2,828) em relação aos tempos de 1 (0,3333±0,5774), 12
(0,6667±0,5774) e 24 horas p.i. (1,667±1,528). Também, nós observamos aumento significativo no
tempo de 96 horas p.i. (13±2,828) em relação ao tempo de 48 horas p.i. (7±2,646).
44
VITORIANO-SOUZA, J.
Resultados
Na análise comparativa entre grupos, verificou-se um aumento (P<0.05) de eosinófilos no
tempo de 1 hora p.i. no grupo Sal (5±3,606) quando comparado ao grupo LB (0±0). Este aumento de
eosinófilos no grupo Sal (5,333±2,309) se manteve no tempo de 12 horas p.i. quando comparados
aos grupos CS (0,3333±0,5774), Sap (0,6667±0,5774), LB (0±0) e LBSap (0,6667±1,155).
40
Percentual de eosinófilos (%)
20
a,b,c,d
a,b,c
0
1h
40
1h
12h
24h
12 h
48h
96h
48 h
24 h
96 h
20
0
CS
Sap
Sal
LB
LBSap
Tempo
Gráfico 12: Cinética de migração de eosinófilos na hipoderme de cães inoculados com diferentes
componentes: CS (controle salina), Sap (saponina), Sal (extrato de glândula salivar de Lu.
longipalpis), LB (antígeno de L. braziliensis) e LBSap (antígeno de L. braziliensis associado a
saponina). As análises foram realizadas 1, 12, 24, 48 e 96 horas p.i. Os resultados estão expressos
como média ± desvio padrão. No Gráfico de cinética, as diferenças estatísticas estão representadas
pelas letras “a”, “b”, “c” e “d” referentes aos tempos de 1, 12, 24 e 48 horas p.i., respectivamente. As
diferenças significativas (P<0.05) da análise comparativa entre os grupos estão demonstradas pelas
linhas.
6.5.2 - Migração de linfócitos na hipoderme de cães
A cinética de migração de linfócitos na hipoderme está representada no Gráfico 13. Na
avaliação de migração de linfócitos foi observado aumento (P<0.05) no grupo Sap no tempo de 48
horas p.i. (19±4) em relação ao tempo de 24 horas p.i. (7±5,292). No entanto, nenhuma diferença
significativa foi observada na análise entre os grupos.
45
VITORIANO-SOUZA, J.
Resultados
100
Percentual de linfócitos (%)
50
c
a
0
1h
100
1h
12h
24h
12 h
48h
24 h
96h
48 h
96 h
50
0
CS
Sap
Sal
LB
LBSap
Tempo
Gráfico 13: Cinética de migração de linfócitos na hipoderme de cães inoculados com diferentes
componentes: CS (controle salina), Sap (saponina), Sal (extrato de glândula salivar de Lu.
longipalpis), LB (antígeno de L. braziliensis) e LBSap (antígeno de L. braziliensis associado a
saponina). As análises foram realizadas 1, 12, 24, 48 e 96 horas p.i. Os resultados estão expressos
como média ± desvio padrão. No Gráfico de cinética, as diferenças estatísticas estão representadas
pela letra “c” referente ao tempo de 1 hora p.i., respectivamente.
6.6 – Análise semiquantitativa da expressão da enzima óxido nítrico sintase induzida (iNOS)
na pele de cães inoculados com diferentes componentes
Após a realização da imuno-histoquímica para detecção da enzima iNOS, as lâminas foram
avaliadas de maneira semiquantitativa para análise da imunorreatividade de iNOS na derme e
hipoderme, bem como em queratinócitos, glândulas sebáceas e sudoríparas. Desse modo, os padrões
utilizados foram denominados como ausente (-), discreto (+), moderado (++) e intenso (+++).
No grupo Sap observou-se imunorreatividade em todas as camadas da pele (derme e
hipoderme), bem como anexos. Essa marcação variou de moderada a intensa nos queratinócitos,
discreta a intensa na derme, ausente a intensa na hipoderme.
Em relação ao grupo Sal, a marcação nos queratinócitos oscilou de moderada a intensa,
durante toda a cinética. Na derme, a intensidade de marcação variou de discreta a intensa e na
hipoderme houve variação de ausente a intensa.
46
VITORIANO-SOUZA, J.
Resultados
Já no grupo LB, observou-se o mesmo padrão de marcação de queratinócitos observado nos
grupos acima descritos, variando de moderada a intensa durante toda a cinética. Na derme, a
intensidade de marcação variou de discreta a intensa, sendo observada na hipoderme oscilação da
intensidade de ausente a intensa.
Resultados encontrados no grupo LBSap em relação aos queratinócitos e à derme,
demonstraram intensa marcação em todos os tempos da cinética. Na hipoderme, a intensidade variou
de moderada a intensa (Figura 5).
Interessantemente, observamos no grupo CS imunorreatividade em relação ao queratinócitos,
variando de discreta a intensa. Essas marcações também foram observadas na derme e hipoderme
com intensidade variando de ausente a intensa.
Houve também marcação nas glândulas presentes, fibroblastos e células endoteliais em todos
os grupos analisados. Como observado nos resultados supracitados, a intensidade de marcação
variou dentro do mesmo grupo demonstrando perfil heterogêneo que poderia refletir a diferença na
maturidade de células recém-migradas para o local do inóculo após estímulos vacinais.
6.7 - Análise quantitativa da marcação da enzima óxido nítrico sintase induzida (iNOS) na pele
de cães inoculados com diferentes componentes
Buscando ampliar o estudo da cinética do infiltrado inflamatório utilizando a vacina LBSap e
seus constituintes bem como do extrato de glândula salivar de Lu. Longipalpis foi empregada a
análise de imagem para um estudo quantitativo dos níveis de iNOS na pele dos cães.
A avaliação longitudinal da produção da iNOS revelou um aumento (P<0.05) nos grupos
Sap (1258±264) e LBSap (1304±289,8) no tempo de 12 horas p.i. em relação ao tempo inicial de 1
hora p.i. (353,3±52,75-Sap e 306,8±6,445-LBSap). Na análise comparativa entre grupos observou-se
que o grupo LB (496,7±163,6) apresentou aumento na área marcada para iNOS no tempo de 1 hora
(P<0.05) quando comparado ao grupo CS (117,4±86,99). Em 12 horas p.i., tanto o grupo Sap
(1258±264) como LBSap (1304±289,8) apresentaram áreas marcadas para iNOS maiores (P<0.05)
em relação aos grupos CS (269,6±170,5), Sal (613,5±189,3) e LB (376,6±68,23). No tempo de 24
horas, o grupo LBSap (1166±366,4) manteve o aumento (P<0.05) de área marcada para iNOS
quando comparado aos grupos CS (146,9±136,5) e LB (454,4±238,3). De forma similar, no tempo
de 48 e 96 horas p.i. foi observado aumento (P<0.05) da área marcada para iNOS no grupo LBSap
(1028±142,9 e 1022±416,9) quando comparado aos grupos CS (130,2±12,38 e 68,35±46,16), Sal
(468,2±161,4 e 205,2±93,07) e LB (438,3±89,87 e 346±280,8). Além disto, de maneira semelhante
ao grupo LBSap, o grupo Sap (1157±357) apresentou aumento (P<0.05) da área marcada para iNOS
47
VITORIANO-SOUZA, J.
Resultados
no tempo final de 96 horas em relação aos grupos CS (68,35±46,16), Sal (205,2±93,07) e LB
(346±280,8). Os resultados referentes a análise quantitativa da enzima iNOS estão representados no
Gráfico 14.
2000
a
a
Área marcada (mm2)
1000
0
1h
2000
1h
12h
24h
12 h
48h
96h
24 h
48 h
96 h
1000
0
CS
Sap
Sal
LB
LBSap
Tempo
Gráfico 14: Análise da área marcada pela enzima iNOS na pele de cães inoculados com diferentes
componentes: CS (controle salina), Sap (saponina), Sal (extrato de glândula salivar de Lu. longipalpis),
LB (antígeno de L. braziliensis) e LBSap (antígeno de L. braziliensis associado a saponina). As
análises foram realizadas nos tempos de 1, 12, 24, 48 e 96 horas p.i. Os resultados estão expressos
como média ± desvio padrão. No Gráfico de cinética, a diferença estatística está representada pela letra
“a” referente ao tempo de 1 hora p.i. As diferenças significativas (P<0.05) da análise comparativa entre
os grupos estão demonstradas pelas linhas.
48
VITORIANO-SOUZA, J.
Resultados
1h
12 h
96 h
50 µm
50 µm
Derme
50 µm
A
50 µm
C
50 µm
Hipoderme
50 µm
B
D
25 µm
E
50 µm
F
50 µm
I
25 µm
G
H
J
Figura 5: Fotomicrografia da IMH para detecção da enzima iNOS na derme e hipoderme de
cães imunizados com LBSap. Nessa figura está representada a intensa área marcada pela
enzima iNOS neste grupo nos tempos de 1, 12 e 96 horas p.i na derme e hipoderme (A – F,
aumento de 40x, barra: 50 µm); em G, neutrófilos iNOS-positivo (aumento 100x, barra:
25µm) do grupo Sap; em H-J, controle negativo da reação.
49
VITORIANO-SOUZA, J.
Resultados
6.8 - Correlações entre o número de núcleos de núcleos de células e área marcada pela enzima
iNOS na derme e hipoderme
Foi realizada análise de correlação entre a expressão da enzima iNOS e número de núcleos de
células nos diferentes grupos experimentais, onde observou-se na derme correlação positiva entre
número de núcleos de células totais presentes derme e área marcada pela enzima iNOS no grupo Sap
(P = 0.0265 e r = 0.6924). Já na hipoderme, verificou-se correlação positiva entre número total de
núcleos de células e área marcada da pela enzima iNOS no grupo LBSap (P=0.0427 e r=0.5683).
Nos demais grupos não houve correlações entre esses parâmetros avaliados (Gráfico 15).
Derme
Número de núcleos/1,5x106 µm2
400
Sap
Hipoderme
1000
LBSap
500
200
P=0.0427
P=0.0265
r=0.6924
0
0
1000
r=0.5683
0
2000
Área marcada pela
0
iNOS/ 1,5x106
1000
2000
µm2
Gráfico 15: Correlação entre o número de núcleos totais e área marcada pela enzima iNOS na derme
e hipoderme. Neste Gráfico está demonstrada a associação positiva nos grupos Sap ( ) e LBSap ( )
na derme e hipoderme, respectivamente, bem como os valores de P e r (correlação de Pearson).
6.8.1 - Correlações na derme superior
Utilizando-se o teste de Pearson, foi possível verificar algumas correlações na derme
superior. Neste sentido, o grupo Sal apresentou correlação negativa entre o percuntual de neutrófilos
e a área marcada pela enzima iNOS (P=0.0476 e r=-0.6072) e entre neutrófilos e número de núcleos
de células (P=0.0289 e r=-0.6277). Observou-se nesse grupo correlação positiva entre macrófagos e
área marcada pela enzima iNOS (P=0.0024 e r=0.8113).
50
VITORIANO-SOUZA, J.
Resultados
Nesta camada, pôde-se notar ainda, associação positiva no grupo LB entre o percentual de
macrófagos e área marcada pela enzima iNOS (P=0.0445 e r=0.5876). Os resultados referentes as
correlações na derme superior estão representados no Gráfico 16.
6.8.2 - Correlações na derme profunda
As correlações observadas na derme profunda estão representadas no Gráfico 17. No grupo
Sal, como demonstrado acima na derme superior, verificou-se correlação negativa entre o percentual
de neutrófilos e o número de núcleos de células (P=0.0446 e r=-0.5434). No grupo Sap notou-se
correlação positiva entre eosinófilos e número de núcleos celulares (P=0.0402 e r=0.5742). No
grupo LBSap observou-se correlação entre o percentual de macrófagos e a área marcada pela enzima
iNOS (P=0.0346 e r=-0.5880). Na hipoderme não foi observada nenhuma correlação significativa.
51
VITORIANO-SOUZA, J.
100
SAL
Resultados
100
P=0.0476
LB
r=-0.6072
50
50
P=0.0445
0
0
500
1000
iNOS
100
SAL
P=0.0289
macrófagos (%)
Neutrófilos (%)
r=0.5876
0
0
500
1000
iNOS
100
SAL
r=-0.6277
50
50
P=0.0024
r=0.8113
0
0
100
0
200
0
500
1000
iNOS
Número de núcleos
Gráfico 16: Correlações na derme superior. Neste Gráfico estão representadas as correlações no
grupo Sal ( ): neutrófilos (%) x área marcada pela iNOS; neutrófilos (%) x número de núcleos;
macrófagos (%) x área marcada pela enzima iNOS. No grupo LB ( ), associação entre macrófagos
(%) x área marcada pela enzima iNOS. Os valores de P e r estão demonstrados no gráfico
(correlação de Pearson).
52
VITORIANO-SOUZA, J.
20
P=0.0446
SAL
r=-0.5434
Eos inófilos (%)
Ne utrófilos (%)
100
Resultados
50
0
0
100
P=0.0402
r=0.5742
10
Sap
0
200
200
0
400
Número de núcle os
Macrófagos (%)
100
P=0.0346
r=-0.5880
LBSap
50
0
1000
0
2000
iNOS
Gráfico 17: Correlações na derme profunda. Neste Gráfico estão representadas as correlações
existentes no grupo Sal ( ): neutrófilos (%) x número de núcleos; no grupo LBSap ( ): macrófagos
(%) x área marcada pela enzima iNOS; no grupo Sap ( ) eosinófilos (%) x número de núcleos. Os
valores de P e r estão demonstrados no gráfico (correlação de Pearson).
53
7 - Discussão
VITORIANO-SOUZA, J.
Discussão
A Leishmaniose Visceral (LV) é uma das mais relevantes e importantes doenças
parasitárias (WHO, 2005) em expansão em diversas regiões do mundo (Enserink, 2000). O
caráter emergente e re-emergente dessa protozoonose deve-se a diversas falhas no controle de
reservatórios e vetores, a associação do parasito às infecções oportunistas tais como a AIDS
(Badaró et al., 1986; Altes et al., 1991; Gradoni et al., 1993), bem como aos recentes relatos da
seleção de cepas resistentes a drogas.
Um dos pontos fundamentais do controle da LV preconizado pela OMS e pelo Ministério
da Saúde do Brasil (2003) baseia-se na eutanásia dos cães infectados, uma vez que o tratamento
com drogas utilizadas com sucesso em casos humanos como glucanato antimonial de sódio ou
stiboglucanato de sódio mostra baixa eficácia em cães (Alencar, 1959; Alvar et al., 1994; Baneth
& Shaw, 2002; Noli & Auxilia, 2005). Todavia, a eliminação desses cães nem sempre é aceita
por razões éticas e sociais (Desjeux, 2004). Diante dessas dificuldades, a imunoprofilaxia pode
ser empregada como uma alternativa promissora para a prevenção da LV, atuando no sentido de
diminuir a incidência dos casos caninos e conseqüentemente, da doença humana (Marzochi et
al., 1985; Genaro, 1993; Hommel et al., 1995; Genaro et al., 1996a; Gradoni, 2001; Handman,
2001; Desjeux, 2004; Ravindran & Ali, 2004; Requena et al., 2004).
Até o momento diversas vacinas anti-LVC utilizando diferentes antígenos e adjuvantes
foram testadas no Brasil, sendo algumas delas com resultados muito promissores (Genaro et al.,
1996; Mayrink et al.,1996; Palatnik et al., 1990; Palatnik de Sousa et al., 1994a; Palatnik de
Sousa et al.,1994b; Da Silva et al., 2001; Borja-Cabrera et al., 2002; Giunchetti, 2007;
Giunchetti et al., 2007; Giunchetti et al., 2008a; Giunchetti et al., 2008b).
Buscando o desenvolvimento de uma vacina eficaz e segura, nosso grupo de pesquisa
vem estudando dois novos imunobiológicos contra LVC. Os resultados obtidos demonstraram
que estas vacinas (LBSap e LBSapSal) apresentam forte capacidade indutora de
imunogenicidade estimulando altos níveis de isotipos IgG anti-Leishmania (IgG1 e IgG2) e de
linfócitos, particularmente linfócitos T CD8+ circulantes e linfócitos T CD8+
L. chagasi-
específicos in vitro. Além disto, outros fatores demonstraram que estes imunobiológicos
induzem uma resposta imune compatível com o controle efetivo do agente etiológico da LVC,
tais como intensa atividade linfoproliferativa L. chagasi-específica e aumento da produção de
NO durante a estimulação in vitro por antígenos solúveis de L. chagasi (Giunchetti, 2007;
Giunchetti et al., 2007; Giunchetti et al., 2008a). É importante ressaltar que nestes estudos foram
observadas alterações no local do inóculo vacinal, quando a saponina foi utilizada como
adjuvante ou quando associada aos antígenos vacinais. No entanto, estas alterações
56
VITORIANO-SOUZA, J.
Discussão
apresentaram-se brandas, e as vacinas LBSap e LBSapSal foram consideradas seguras para
utilização em cães.
A avaliação das alterações no local do inóculo vacinal através dos aspectos
anatomopatológicos (macroscópicos) como formação de edema e úlceras, ou pelos aspectos
histopatológicos e/ou imunopatológicos como produção de NO, dosagem de citocinas e
quimiocinas representam biomarcadores importantes para análise da inocuidade vacinal e da
imunogenicidade no contexto da resposta imune inata que ocorre in situ, ou seja, no local da
vacinação. A compreensão destes eventos nos permitirá em breve propor um novo protocolo de
pré-testes para selecionar futuros potenciais candidatos vacinais e a conduzir os ensaios de fase I
e II, no sentido de prosseguir para os ensaios clínicos de campo. Desta forma, Sesardic (2006)
propõe a avaliação de estudos pré-clínicos para a identificação da melhor composição vacinal,
que se apresente inócuo e segura, como controle de qualidade adicional para o desenvolvimento
de candidatos vacinais.
Neste sentido, o presente estudo buscou avaliar o processo inflamatório induzido por
antígenos e adjuvantes vacinais no local do inóculo durante uma cinética incluindo os tempos de
1, 12, 24, 48 e 96 horas. Considerando, que a imunidade contra agentes infecciosos requer a ação
orquestrada de componentes da imunidade inata e adaptativa, o estudo do recrutamento celular
torna-se muito relevante, uma vez que o número de células e a composição do infiltrado celular
no estágio inicial depois do estímulo influenciarão futuras respostas e o desenvolvimento de uma
resposta imune adquirida (Teixeira et al., 2005).
Embora muitos pesquisadores concentram esforços na análise do perfil de citocinas da
reação inflamatória (Cohen, 1976; Dunn et al.,1989; Berg et al., 1995), o estudo do perfil celular
apresenta grande importância, visto que é indicador do processo inflamatório local e implica
diretamente no perfil de citocinas presentes nesse microambiente (Stwart et al., 1984; De Souza,
2000).
Neste sentido, importantes trabalhos realizados por Tafuri (1992) e Tafuri et al., (1993)
buscaram avaliar a cinética celular por meio do perfil histopatológico da intradermorreação de
Montenegro (IRM) na pele de cães procedentes de áreas endêmicas para Leishmaniose
Tegumentar. Estes autores, avaliaram a cinética de aspectos histopatológicos em diferentes
tempos (6, 12, 24, 48 e 72 horas, 7 e 14 dias) após a inoculação de dois antígenos Leishvacin® e
P10.000G. Neste estudo, foi observado que em 24 horas, ocorre um infiltrado inflamatório
discreto composto por neutrófilos, e subseqüentemente, após 48 e 72 horas, composto
principalmente por intenso infiltrado de células mononucleares, com declínio da intensidade da
reação após 7 e 14 dias do inóculo. É importante ressaltar que os trabalhos realizados por Tafuri
57
VITORIANO-SOUZA, J.
Discussão
(1992) e Tafuri et al., (1993) assemelham-se a nosso trabalho considerando o modelo
experimental utilizado, mas difere em relação à região onde foram realizados os inóculos. No
presente estudo decidiu-se pela realização dos inóculos na região dorsal em primeiro lugar
porque a drenagem linfática da região abdominal é extremamente heterogênea e em segundo
lugar para evitar o contato direto com a baia, o que poderia, eventualmente, interferir no
experimento.
Adicionalmente, Moreira (2008) ao avaliar a cinética de migração celular na derme de
hamsters inoculados com componentes antigênicos das vacinas LBSap e da Leishmune®,
observou intenso infiltrado celular promovido pelo adjuvante saponina e pela vacina Leishmune®
em relação ao grupo controle (salina). De forma interessante, esse aumento não foi observado no
grupo LBSap sugerindo que nesse modelo, o antígeno de Leishmania atuaria na modulação da
resposta da saponina. Esses antígenos promoveram recrutamento seletivo de populações
celulares no grupo saponina onde houve migração precoce (1, 12 e 24 horas) e persistente (88, 96
e 168 horas) de macrófagos e diminuição de neutrófilos no tempo tardio (168 horas) associado
ao aumento de linfócitos (96 e 168 horas), reforçando o papel do adjuvante saponina na indução
do tráfego celular conforme sugerido por Kensil (1996).
Sendo assim, buscando ampliar o entendimento de como antígenos e adjuvantes vacinais
poderiam induzir resposta imune, torna-se importante avaliar no modelo cão potenciais vacinas
contra LVC para minimizar possíveis variações em decorrência das particularidades das
respostas observadas entre diferentes modelos experimentais (Gradoni, 2001). Desse modo, nós
decidimos analisar os eventos da resposta imune celular “recentes” como a migração celular e
expressão da enzima iNOS, induzidos pelos componentes antigênicos da vacina LBSap, usando a
região dorsal de cães como modelo experimental de vacinação intradérmica.
Os resultados obtidos através das observações macroscópicas demonstraram apenas
inchaço local após o inóculo com Sap e LBSap, sugerindo o envolvimento de inflamação aguda
local em resposta ao adjuvante saponina, importante para desencadear resposta imune específica
(Jancar, 2001). No entanto, o fato dessas alterações não evoluírem para lesões ulceradas, nos
permite inferir que a saponina sozinha ou associada ao antígeno não apresenta alterações
significativas no local do inóculo, mostrando-se segura para ser empregada como adjuvante
vacinal. Giunchetti et al. (2007) também observaram resultados similares em cães vacinados com
LBSap demonstrando que a vacina é bem tolerada, bem como o adjuvante saponina. Outros
estudos com adjuvantes, como BCG usados em vacinas humana e canina anti-Leishmania
(Mayrink et al., 1996; Giunchetti et al., 2008b), também relatam fortes reações inflamatórias,
muitas vezes associadas com lesões cutâneas ulceradas, responsáveis pelo recrutamento inicial
58
VITORIANO-SOUZA, J.
Discussão
de células inflamatórias. Parra et al. (2007), ao analisarem a toxicidade e inocuidade da
Leishmune® observaram que as principais reações notificadas como dor, anorexia e inchaço local
foram moderadas, indicando que a vacina foi bem tolerada e todos os efeitos foram
espontaneamente revertidos, antes da imunização subseqüente.
Algumas observações histopatológicas como edema, hemorragia e congestão foram
verificadas em maior intensidade nos grupos Sap e LBSap. Essas reações locais são observadas
freqüentemente em imunizações vacinais com este adjuvante e podem variar de acordo com a
pureza da saponina utilizada (Kensil, 1996). Em um trabalho utilizando inóculo de 1 mg de
saponina pela via subcutânea em ovelhas, observou-se reações inflamatórias variando de
moderadas a intensas com edema e duração do evento por volta de 2 a 3 dias (Houdayer et
al.,1978).
No presente estudo, alterações discretas foram observadas em biópsias de pele de cães
inoculados apenas com Sal e LB nos tempos de 12 e 24 horas p.i. Estas alterações podem ser
resultantes da própria imunização que induziu um discreto processo inflamatório associado ao
aumento da permeabilidade vascular no local. Provavelmente estas alterações foram menos
intensas em função da ausência do adjuvante saponina nestes grupos.
De forma interessante, a quantificação do infiltrado celular na derme através da
morfometria por análise de imagem revelou que o adjuvante saponina apresenta papel importante
para o recrutamento celular nos tempos de 1, 12, 24 e 96 horas p.i. na derme de cães,
representado pelo aumento do número de células no infiltrado inflamatório no grupo Sap. Esses
resultados estão de acordo com outros estudos, que descrevem o papel de saponina na modulação
da resposta imune celular, no aumento da produção de anticorpos, em reações imunes não
específicas como a inflamação e também no tráfego celular (de Oliveira et al., 2001; Kensil,
1996). Nos demais grupos, não foram observadas diferenças quando se avaliou a derme.
Na hipoderme, o aumento do número de células observado nos grupos Sap e LBSap
revelou ser maior em relação a derme, possivelmente, devido à maior vascularização desta
camada. Em nosso estudo, observamos aumento do infiltrado celular na hipoderme de cães dos
grupos Sap e LBSap nos tempos de 12, 24 e 48 horas quando comparados com CS, Sal e LB.
Além disto, o grupo Sap apresentou aumento do infiltrado celular nos mesmos tempos (12, 24 e
48 horas) quando comparado ao grupo LBSap. Adicionalmente, no tempo de 1 hora, o grupo Sap
apresentou aumento do infiltrado celular em relação aos grupos CS, e em 96 horas, aumento em
relação aos grupos CS e Sal. Durante toda a cinética, o grupo Sap induziu maior recrutamento
celular em relação ao grupo LBSap. Novamente, esses resultados corroboram para o papel do
adjuvante saponina no tráfego celular em direção ao local do inóculo. Além disto, é possível
59
VITORIANO-SOUZA, J.
Discussão
especular que há uma modulação no recrutamento celular em alguns tempos da cinética, quando
associado o antígeno de L. braziliensis ao adjuvante saponina.
Uma vez que se observou o aumento do infiltrado celular, acionado pelos componentes
antigênicos vacinais, durante diversos momentos da cinética, decidiu-se avaliar também qual
seria o perfil de células recrutadas nas diferentes camadas da pele (derme superior, derme
profunda e hipoderme). Na derme superior, foi observado maior recrutamento de neutrófilos no
grupo Sap, enquanto nos grupos Sap e LBSap houve aumento de linfócitos associado à redução
de macrófagos. Já na derme profunda, maior recrutamento de neutrófilos no grupo Sap e redução
de macrófagos nos grupos LBSap; e na hipoderme, maior recrutamento de neutrófilos e redução
de macrófagos nos grupos Sap e LBSap e aumento de migração de eosinófilos no grupo Sal.
De modo geral, encontramos um perfil bem semelhante nos grupos Sap e LBSap com
aumento de neutrófilos e linfócitos em alguns tempos da cinética e queda de macrófagos quando
comparado aos demais grupos, no entanto, verificamos também na análise longitudinal de
eosinófilos, um importante aumento nos últimos tempos no grupo Sap na derme profunda e
hipoderme e no grupo LBSap na derme superior e profunda.
Esses resultados nos permitiram verificar e destacar a participação de neutrófilos na
resposta imune inata induzida por vacina e adjuvantes. Esta hipótese ganha força, considerando
os diversos estudos em que tem sido discutido o papel de neutrófilos como um tipo celular com
funções mais complexas que simples células fagocitárias, demonstrando seu papel na
determinação da resposta imune e cooperação com outros tipos celulares para melhorar a
resposta efetora do sistema imune (Appelberg, 2006).
Similar a esses resultados, em trabalhos realizados por Moreira (2008) em nosso
laboratório, utilizando-se o modelo hamster, verificou-se aumento de neutrófilos circulantes no
grupo inoculado com saponina, sugerindo o papel de neutrófilos durante o processo de
imunização. Além disto, trabalhos recentes têm mostrado o papel de neutrófilos na imunidade
adquirida como célula apresentadora de antígenos para linfócitos T virgens (Beauvillain et al.,
2007), uma vez que neutrófilos seriam as primeiras células a migrarem, em grande quantidade
para o local da infecção. Assim, eles poderiam se acumular precocemente no local da infecção,
antes das células dendríticas e macrófagos, e poderiam interagir com células T CD8+ antes das
células dendríticas (Beauvillain et al., 2007). De fato, tem sido descrito, o importante papel da
utilização de saponina como adjuvante na indução de células T CD8+ (Kensil, 1996; BorjaCabrera, 2004; Giunchetti et al., 2007; Giunchetti et al., 2008a) e, neste contexto, os neutrófilos
poderiam ter uma importante contribuição. É importante ressaltar ainda, que outros autores
sugerem que neutrófilos poderiam atuar de forma indireta na apresentação de antígenos bem
60
VITORIANO-SOUZA, J.
Discussão
como no recrutamento de outros tipos celulares tais quais as células T, monócitos, macrófagos e
células dendríticas imaturas, por meio de produção de quimiocinas (Scapini et al., 2000). A
comunicação entre neutrófilos e células dendríticas imaturas poderia resultar na maturação das
últimas, e assim no aumento da capacidade de apresentação de antígenos e proliferação das
células T (Appelberg, 2006).
Considerando o exposto e os resultados obtidos neste trabalho, é possível hipotetizar que
os neutrófilos estariam exercendo um papel altamente relevante na linha de frente da resposta
celular imediata decorrente do processo de imunização e, orquestrando as demais células a
migrarem após algumas horas para o local do inoculo, participando efetivamente do início da
resposta adaptativa. Desta maneira, os neutrófilos poderiam apresentar um papel importante no
direcionamento da resposta imune através da produção de quimiocinas. Após ativação, estas
células produziriam diversas quimiocinas (IL-8, GRO-α, MIP-1α e β) que atuariam em
receptores de células tipo 1. Desta forma, tem sido relatado que os neutrófilos, através da
secreção de quimiocinas, induziriam uma polarização para a resposta de células tipo 1, pela
atração preferencial desse tipo celular para o local do inóculo (van Gisbergen et al., 2005). Sendo
assim, novas abordagens para o estudo de vacinas baseadas na indução de linfócitos T
citotóxicos, através do uso preferencial de adjuvantes que favoreçam o recrutamento seletivo de
neutrófílos seriam muito relevantes.
A avaliação cinética longitudinal realizada neste trabalho revelou aumento de eosinófilos
nos grupos Sap e LBSap nos tempos finais (48 e 96 horas). Este aumento sugere que a
imunização com estes antígenos possa ter gerado um perfil de reposta mista tipo1/tipo2 e nos
permite hipotetizar que essas células, juntamente com os neutrófilos participariam efetivamente
na resposta inata e no direcionamento da resposta específica para o local da imunização. De fato,
tem sido proposto que a imunização com as vacinas LBSap ou LBSapSal em cães, induziriam
um perfil de resposta imune humoral compatível com uma resposta mista do tipo 1 e tipo 2
(Giunchetti, 2007; Giunchetti et al., 2007; Giunchetti et al., 2008a). De forma interessante,
Moreira (2008) não relatou nenhuma alteração na contagem desse tipo celular, sugerindo que a
ausência de diferença nesse trabalho deve-se aos distintos modelos avaliados, sendo um em cão e
outro em hamster, que em muito diferem quanto ao tipo de resposta.
Um interessante estudo realizado por Pearson et al. (1987) demonstrou que eosinófilos
apresentam capacidade microbicida frente a Leishmania. Esses resultados sugerem que a
ativação de eosinófilos em microambientes contendo IFN-γ levaria a estimulação da produção de
vários mediadores inflamatórios. Assim, o recrutamento de eosinófilos observado poderia
representar um mecanismo leishmanicida adicional que compensaria a ausência do envolvimento
61
VITORIANO-SOUZA, J.
Discussão
precoce de macrófagos (Beil et al., 1992; Oliveira et al., 1997 e 1998; Watanabe et al., 2004).
Em camundongos BALB/c, os neutrófilos são as células predominantes nos primeiros três dias
após infecção por Leishmania, acompanhados de um discreto infiltrado de eosinófilos (Andrade
et al., 1984; Beil et al., 1992). Após análise histopatológica, observou-se a presença de parasitos
íntegros e destruídos no interior de neutrófilos e também de eosinófilos, sugerindo a participação
destas células no controle inicial da infecção por Leishmania (Lima et al., 1998).
Em relação a queda percentual de macrófagos, poderíamos associá-la ao intenso
recrutamento inicial de neutrófilos, células mais prevalentes da inflamação aguda (Terui et al.,
2000) e não à inibição da migração dessas células pelos componentes antigênicos, visto que os
macrófagos são células importantes na apresentação antigênica em vacinas que utilizam saponina
(Kensil, 1996). Sendo assim, a presença desses dois tipos celulares no mesmo infiltrado nos
permite sugerir que esteja ocorrendo cooperação entre macrófagos e neutrófilos. Neutrófilos
apoptóticos seriam fagocitados por macrófagos e assim transfeririam seus componentes
microbicidas, contribuindo para o aumento da apresentação antigênica (Appelberg, 2006).
Soehnlein et al. (2007) demonstraram que os produtos secretados por neutrófilos disparam uma
resposta ativa em monócitos e macrófagos que aumentam funcionalmente, a capacidade
fagocitária e a formação de radicais reativos de oxigênio. Estes fenômenos sugerem que esse
mecanismo contribuiria para estimular as funções dos macrófagos na imunidade inata.
Estudos recentes, realizados em nosso laboratório no modelo hamster, observaram que a
saponina promove um recrutamento precoce e persistente de macrófagos e que os demais grupos
vacinais demonstram uma manutenção dos níveis normais desta população celular (Moreira,
2008). Esses resultados diferem dos nossos apresentados acima, onde em alguns tempos,
observamos redução dessa população no período inicial da cinética avaliada. Talvez essas
diferenças possam ser explicadas pelo fato de que, no cão, houve um recrutamento superior de
neutrófilos em detrimento dos outros tipos celulares.
Além disto, o aumento de linfócitos nos tempos de 24 horas dos grupos Sap e LBSap e
em 48 horas no grupo LBSap na derme superior, demonstra mais uma vez, o papel do adjuvante
saponina agindo de forma rápida e efetiva no recrutamento celular para o local do inóculo
(Kensil, 1996). A co-participação dessas células da imunidade adaptativa em conjunto com a
resposta inata foi recentemente descrita por Berg & Forman (2006). Este seria um mecanismo
adicional, uma vez que os linfócitos T CD8+ produziriam IFN-γ através da interação com
citocinas IL-12 e IL-18, independente do mecanismo de interação TCR-MHC, e desse modo,
atuariam na destruição de células infectadas, participando do controle de vários tipos de infecção
e inicialização da resposta específica (Berg & Forman, 2006). A presença de linfócitos T CD4+
62
VITORIANO-SOUZA, J.
Discussão
também é crucial, uma vez que produzem IFN-γ ativando macrófagos, aumentando a capacidade
microbicida e produção de citocinas que atuarão na indução da resposta adaptativa e no controle
da infecção por Leishmania (Bittar et al.,2007).
Esses resultados corroboram com os de Moreira (2008) onde também houve
envolvimento desse tipo celular no grupo LBSap, embora no cão esse recrutamento tenha sido
mais precoce (24 e 48 horas) que no modelo hamster (96 e 168 horas).
Rezende (2003), utilizando a análise por citometria de fluxo, mostrou que adjuvantes
associados a uma vacina anti-rábica inoculada através de uma bolha de ar no dorso de
camundongos Swiss foram capazes de promover recrutamento intenso de neutrófilos, de
macrófagos e de linfócitos nos tempos de 24 e 72 horas, importante para o estabelecimento da
resposta específica.
Diante do exposto, podemos inferir que o recrutamento seletivo de linfócitos
principalmente linfócitos T CD8+ produtores de IFN-γ iriam contribuir para uma resposta imune
protetora contra LVC.
É importante notar que o grupo inoculado apenas com extrato de glândula salivar de Lu.
longipalpis apresentou recrutamento similar ao observado com inóculo de salina, tanto na derme
quanto na hipoderme. Em estudos com camundongos BALB/c, verificou-se um infiltrado celular
inflamatório compreendido por neutrófilos, macrófagos e eosinófilos depois de repetida
exposição a picadas de Lu. longipalpis. Adicionalmente, observaram que inóculos de extrato de
glândula salivar em animais não expostos induzia um recrutamento similar aos animais
inoculados com salina (Silva et al., 2005).
O maior recrutamento de eosinófilos observado no inóculo do grupo Sal sugere o
desenvolvimento de uma resposta do tipo 2 importante para criar um microambiente que permita
o estabelecimento de parasitos do gênero Leishmania na infecção natural. Entretanto, de forma
interessante, alguns autores relatam que indivíduos expostos a sucessivas picadas de insetos
apresentam um perfil de resposta mista (tipo 1/tipo 2), onde a resposta imune do hospedeiro seria
iniciada com uma resposta DTH positiva e posteriormente, predominaria uma resposta de
hipersensibilidade imediata até seu desaparecimento (Silva et al., 2005). Belkaid et al. (1998),
demonstram o surgimento de proteção resultante da pré-exposição ao extrato de glândula salivar
de Phlebotomus papatasi, já observada em habitantes de áreas endêmicas para leishmanioses.
O grupo LB também não promoveu intenso recrutamento celular, demonstrando a
importância da utilização concomitante de um adjuvante para melhorar a imunogenicidade desse
antígeno no local do inóculo vacinal.
63
VITORIANO-SOUZA, J.
Discussão
Outra questão importante levantada neste trabalho seria se o recrutamento dessas células
inflamatórias poderia contribuir para o desenvolvimento de um microambiente favorável ao
desenvolvimento de uma resposta eficaz contra futuras infecções por Leishmania. Nos últimos
anos, trabalhos mostraram que a expressão de iNOS após inflamação ou infecção (Nathan & Xie,
1994 a, b) é bem caracterizada e aceita como componente chave da resposta adaptativa a
estímulos estranhos ou patógenos virulentos e este aumento de NO relaciona-se ao controle da
infecção por parasitos do gênero Leishmania (Hall & Titus, 1995).
Desse modo, avaliamos a expressão da enzima iNOS na pele de cães imunizados com
diferentes componentes antigênicos. A análise semiquantitativa demonstrou que ocorre
expressão da enzima em todos os grupos, incluindo o CS, nas diferentes camadas da pele. Esses
resultados nos permitem inferir que todos os estímulos foram capazes de aumentar a expressão
da enzima iNOS, contribuindo para a produção de NO, participando desde processos de resposta
não-específica até na modulação da resposta imune (Bogdan, 2001).
A expressão de iNOS no grupo controle salina sugere a participação do NO na fisiologia
da pele normal. Esses resultados corroboram com os trabalhos de Shimizu et al. (1997) que
observaram marcação pela enzima iNOS em queratinócitos de pele normal de humanos, que
poderia ser importante no controle da proliferação dos queratinócitos. Outra possível justificativa
para a presença de iNOS na pele normal poderia ser devido a exposição de animais aos raios
ultra-violeta (UV) (Kuhn et al., 1998), uma vez que os animais são de uma região tropical.
Uma outra abordagem adotada neste estudo teve como objetivo realizar a quantificação
da área marcada pela enzima iNOS. Neste sentido, foi observado no tempo inicial de 1 hora,
maior área marcada pela iNOS no grupo LB. Este resultado pode ser justificado pela presença de
macrófagos recrutados por este estímulo, considerados grandes produtores de NO, na derme
superior. Os grupos Sap e LBSap apresentaram maior área marcada em relação aos demais
grupos, sendo que esse aumento observado foi significativo a partir de 12 horas até o tempo final
em LBSap e no grupo Sap nos tempos de 12 e 96 horas. As maiores produções de iNOS nesses
grupos provavelmente estão relacionadas ao grande número de células recrutadas para o local,
que uma vez ativadas, produziriam grandes quantidades de NO para a erradicação do parasito.
Além disto, esses resultados também são demonstrados nas correlações entre número de núcleos
e expressão de iNOS nos grupos Sap e LBSap na derme e hipoderme.
É importante ressaltar que a enzima iNOS é expressa em uma variedade de tipos
celulares, incluindo macrófagos, neutrófilos e fibroblastos em resposta a uma variedade de
estímulos tais como citocinas e LPS (Liew, 1997). Vários trabalhos têm demonstrado que
citocinas do tipo 1 como IFN-γ, TNF-α e IL-18 induzem a expressão de iNOS (Liew &
64
VITORIANO-SOUZA, J.
Discussão
O’Donnell, 1993; Bogdan et al.,1993), enquanto, citocinas do tipo 2 (IL-4, IL-13 e IL-10)
diminuem a atividade leishmanicida em modelo murino e em humanos por regularem
negativamente a expressão de iNOS (Vouldoukis et al., 1997). Considerando essas afirmações,
algumas ponderações são importantes para tentar explicar as principais correlações entre a iNOS
e os tipos celulares (%).
Neste sentido, na derme superior, os grupos Sal e LB apresentaram correlação positiva
entre o percentual de macrófagos e a área marcada pela iNOS, sugerindo que estes estímulos
ativariam os macrófagos para produção de NO.
No entanto, é importante ressaltar que alguns trabalhos demonstraram claramente que
componentes da saliva de Lu. longipalpis, como o “maxadilan”, inibem a produção de NO (Hall
& Titus, 1995), peróxido de hidrogênio (H2O2) (Titus & Ribeiro, 1990) e apresentação de
antigênica por macrófagos (Theodos & Titus, 1993). Entretanto, devemos considerar que a saliva
dos fleblótomos possui um amplo repertório de moléculas farmacologicamente ativas que
interferem com a homeostase, inflamação e resposta imune (Titus & Ribeiro, 1990). Desse
modo, essas moléculas poderiam atuar na ativação de macrófagos e assim, disparariam uma
resposta imune específica contra esses antígenos. Valenzuela et al. (2002) hipotetizaram que
proteínas salivares de P. papatasi (como por exemplo, PpSP15) poderiam induzir uma resposta
DTH e macrófagos ativados liberariam radicais de oxigênio, NO e IL-12, atuando como
microbicidas em parasitos do gênero Leishmania.
Adicionalmente, as correlações negativas no grupo Sal entre iNOS e a população de
neutrófilos, bem como entre neutrófilos e o número de núcleos presentes no local do inóculo,
sugerem que o extrato de glândula salivar de Lu. longipalpis não foi capaz de induzir uma
resposta inflamatória aguda com presença de neutrófilos. Como mencionado anteriormente, a
importância da atuação destas células nos mecanismos adicionais de resistência, como por
exemplo na participação da elaboração de uma resposta imune do tipo 1, a baixa capacidade
indutora de neutrófilos na presença de extrato salivar de Lu. longipalpis, poderia refletir em uma
maior susceptibilidade a infecção por Leishmania (Charlab et al.,1999; Cavalcante et al., 2003;
Titus et al., 1988).
Na derme profunda, verificou-se que além da correlação negativa no grupo Sal entre
neutrófilos e o número de células (inflamação), ocorreu também uma correlação positiva no
grupo Sap, entre número de células e eosinófilos, sugerindo a contribuição destas células na
resposta inata provocada pelo forte poder adjuvante da saponina.
Alguns trabalhos descreveram que a atividade inicial de iNOS ocorre primariamente
durante a resposta do tipo 1 e que em um tempo mais tardio atuaria no controle da resposta que
65
VITORIANO-SOUZA, J.
Discussão
poderia causar danos locais (Bogdan, 2001). Esses resultados nos permitem inferir que o
adjuvante saponina (sozinho ou em associação com antígeno de L. braziliensis) induziria um
microambiente no local do inóculo capaz de estimular a polarização do sistema imune para um
perfil de resistência determinante do sucesso de vacinas contra LVC. Considerando que alguns
patógenos infecciosos, como Leishmania, são capazes de inibir a expressão de iNOS,
contribuindo para a limitada expressão inicial de iNOS durante a infecção (Nandan et al., 1999),
a associação do antígeno de L. braziliensis com saponina parece ser muito promissora.
O papel do NO em reações inflamatórias na pele é complexo, pois, em baixas doses
possui propriedades pró-inflamatórias por induzir vasodilatação e recrutamento de neutrófilos,
enquanto que em altas concentrações, regula negativamente a expressão de moléculas de adesão,
suprime ativação e induz apoptose de células inflamatórias (Ross et al., 2001). Desse modo, o
NO participaria do balanço de resposta tipo 1/tipo 2. Esses altos níveis de NO observados nos
grupos imunizados com saponina poderiam estar relacionados aos altos níveis de citocinas tipo 1
(IFN-γ e TNF-α) (Kaim et al., 2006), fundamentais para a ativação de mecanismos efetores,
como produção de NO para o controle de parasitos do gênero Leishmania.
De maneira geral, os resultados obtidos neste trabalho demonstram que a vacina LBSap,
bem como o adjuvante saponina, isoladamente foram capazes de induzir um intenso infiltrado
celular na pele de cães, necessário para os eventos iniciais de apresentação antigênica. Além
disto, este trabalho mostrou que estes componentes são seguros, uma vez, que não evoluíram
para lesões ulceradas locais em todos os tempos da cinética. O recrutamento seletivo de
neutrófilos e a intensa área marcada pela enzima iNOS provocados por Sap e LBSap produziu
um perfil que se assemelha ao observado em resistência a infecções por Leishmania.
66
8 – Conclusões
VITORIANO-SOUZA, J.
Conclusões
Os resultados obtidos neste trabalho indicam que o adjuvante saponina e a vacina LBSap
mobilizam uma forte reação inflamatória no local do inóculo ativando os mecanismos da
imunidade inata e adaptativa, importantes nos processos de apresentação de antígenos
contribuindo para a eficiência da imunização. Além disto, os grupos Sap e LBSap induziram
apenas um pequeno edema local, demonstrando-se seguras e inócuas.
O microambiente formado pelo recrutamento diferencial das células (neutrófilos,
linfócitos e eosinófilos) e aumento da expressão da enzima iNOS induzidos por LBSap e Sap
permite o estabelecimento de uma resposta inata capaz de direcionar uma resposta adaptativa
eficaz contra futuras infecções por L. chagasi.
O recrutamento de eosinófilos pelo grupo Sal reforça o papel imunomodulador de
proteínas da glândula salivar de Lu. longipalpis, fundamentais no estabelecimento da infecção
por Leishmania. Estes resultados reforçam a importância de uma vacina que tenha proteínas de
glândula salivar de Lu. longipalpis em sua composição de forma que haja bloqueio destas
funções.
De modo geral, este trabalho ressalta a importância do estudo de migração celular abrindo
novas perspectivas na prospecção e testes de antígenos e adjuvantes vacinais no modelo cão.
68
9 - Referências Bibliográficas
VITORIANO-SOUZA, J.
Referências Bibliográficas
ABRANCHES, P.; SANTOS-GOMES, G.; RACHAMIN, N.; CAMPINO, L.; SCHUNUR, L.R.;
FAFFE, C.L. An experimental model for canine visceral leishmaniasis. Parasite
Immunology. 13: 537-550, 1991.
AFRIN, F. & ALI, N. Adjuvanticity and the protect immunity elicited by Leishmania
donovani antigens encapsulated in positively charged liposomes. Infection and Immunity.
66 (6): 2371-2377, 1997.
AGA, E.; KATSCHINSKI, D.M.; ZANDBERGEN, G.V.; LAUFS, H.; HANSEN, B.;
MULLER, K.; SOLBACH, W.; LASKAY, T. Inhibition of the spontaneous apoptosis of
neutrophil granulocytes by the intracellular parasite Leishmania major. J Immunol. 169:
898-905, 2002.
APPELBERG, R. Neutrophils and intracellular pathogens: beyond phagocytosis and killing.
Trends in Microbilogy. 15 (2): 2007.
ALENCAR, J.E. Calazar canino: Contribuição para o estudo da epidemiologia do calazar
no Brasil. Fortaleza, Imprensa Oficial. (Tese da Universidade Federal do Ceará), 1959.
ALTES, J.; SALAS, A.; RIERA, M.; UDINA, M.; GALMES, A.; BALANZAT, J.
BALLESTEROS, A.; BUIADES, J.; SALVA, F.; VAILALONGA, C. Visceral
Leishmaniasis: another H.I.V. associated opportunistic infection? Report of eight
cases and review of literature. AIDS(5): 201-20, 1991.
ALVAR, J.; MOLINA, R.; SAN ANDRÉS, M.; TESOURO, M.; NIETO, J.; VITUTIA, M.;
GONZALES, F.; SAN ANDRÉS, M.D.; BOGGIO, J.; RODRIGUEZ, F.; SANIZ, A.;
ESCACENA, C. Canine leishmaniasis: clinical, parasitological and entomological follow
up after chemotherapy. Annals of Tropical Medicine and Parasitology. 88(4): 371-378,
1994.
ANDRADE, Z.A.; REED, S.G., ROTERS, S.B.; SADIGURSKY, M. Immunopathology of
experimental cutaneous leishmaniasis. Am J Pathol. 114:137-148,1984.
ANDRADE, B.B.; DE OLIVEIRA, C.I.; BRODSKYN, C.I.; BARRAL. A.; BARRAL-NETO,
M. Role of Sand Fly Saliva in Human and Experimental Leishmaniasis: Current
Insights. Scad. J. Immunol. 66: 122-127, 2007.
ARAÚJO, M. S. S. Alterações imunológicas no sangue periférico de cães submetidos à
imunoprofilaxia para leishmaniose visceral canina (Tese de Doutorado em Ciências).
Belo Horizonte: Instituto de Ciências Biológicas da Universidade Federal de Minas Gerais,
180p. 2006.
ARIAS, J.R.; MONTEIRO, P.S.; ZICKER, F. The reemergence of visceral leishmaniasis in
Brazil. Emerg. Infec. Dis. 2: 145-146, 1996.
BADARÓ, R.; ROCHA, H.; CARVALHO, E.M.; QUEIROZ, A.C.; JONES, T.C. Leishmania
donovani: an opportunistic infection associated with progressive disease in three
immunocompromised patients. Lancet ii: 647-649, 1986.
70
VITORIANO-SOUZA, J.
Referências Bibliográficas
BANETH, G.; SHAW, S.E. Chemotherapy of canine leishmaniosis. Vet Parasitol.. 106 (4):
315-24, 2002.
BEAUVILLAIN, C.; DELNESTE, Y.; SCOTET, M.; PERES, A.; GASCAN, H.;
GUERMONPREZ, P.; BARNABA, V.; JEANNIN, P. Neutrophils efficiently cross-prime
naive T cells in vivo. Blood. 110 (8): 2965-2973, 2007.
BEIL, WJ, MEINARDUS-HAGER G, NEUGEBAUER DC, SORG C. Differences in the onset
of the inflammatory response to cutaneous leishmaniasis in resistant and susceptible
mice. J Leukoc Biol; 52: 135-142, 1992.
BELKAID, Y.; KAMHAWI, S.; MODI, G.; VALENZUELA, J.; NOBEN-TRAUTH, N.;
ROWTON, E.; RIBEIRO, J.; SACKS, D.L. Developmente of a natural model of
cutaneous Leishmaniasis: powerful effects of vector saliva and saliva and saliva
preexposure on the long-term outcome of Leishmania major infection in the mouse ear
dermis. J. Exp. Med. 188: 1941-1953, 1998.
BELKAID, Y.; VALENZUELA, J.G.; KAMHAWI, S.; ROWTON, E.; SACKS, D.L.;
RIBEIRO, J.M. Delayed-type hypersensitivity to Phlebotomus papatasi sand fly bite: An
adaptative response induced by the fly? Proc.Natl. Acad. Sci. USA. 97: 6704-9, 2000.
BELKAID, Y.; VON STEBUT, E.; MENDEZ, S.; LIRA, R.; CALER, E.; BERTHOLET, S.;
UDEY, M.C.; SACKS, D. CD8+ T cells are required for primary immunity in C57BL/c
mice following low-dose, intradermal challenge with Leishmania major. J Immunol 168:
3992-4000. 2002.
BERG, D.J.; LEACH, M.W.; KÜHN, R.; RAJEWSKY, K.; MÜLLER, W.; DAVIDSON, N.J.;
RENNICK, D. - Interleukin 10 but not interleukin 4 is a natural suppressant of
cutaneous inflammatory responses. J Exp Med 182(1): 99-108, 1995.
BERG, R.E.; FORMAN, J. The role of CD8 T cells in innate immune and in antigen nonspecific protection. Current Opinion in Immunology. 18: 338-343, 2006.
BITTAR, R.C.; NOGUEIRA, R.S.; VIEIRA-GONÇALVES, R.; PINHO-RIBEIRO, V.;
MATTOS, M.S.; OLIVEIRA-NETO, M.P.; COUTINHO, S.G.; DA-CRUZ, A.M. T-cell
responses associated with resistance to Leishmania infection in individuals from
endemic areas for Leishmania (Viannia) braziliensis. Mem. Inst. Oswaldo Cruz. 102(5):
625-30, 2007.
BOGDAN, C.; VODOVOTZ, Y.; PAIK, J.; XIE, Q.W.; NATHAN, C. Traces of bacterial
lipopolysaccharide suppress IFN-gamma-induced nitric oxide synthase gene expression
in primary mouse macrophages. J Immunol 151: 301–309, 1993.
BOGDAN, C. Nitric Oxide and the immune response. Nature Immunology. 2(10):907-916,
2001.
BORJA-CABRERA, G.P.; CORREIA PONTES, N.N.; DA SILVA, V.O.; PARAGUAI DE
SOUZA, E.; SANTOS, W.R.; GOMES, E.M.; LUZ, K.G.; PALATNIK, M.; PALTNIK DE
SOUZA, C.B. Long lasting protection against canine Kala-azar using the FML-QuilA
71
VITORIANO-SOUZA, J.
Referências Bibliográficas
saponin vaccine in ana rea endemic of Brazil (São Gonzalo do Amarante, RN). Vaccine,
20: 3277-3284, 2002.
BORJA-CABRERA, G.P.; CRUZ MENDES, A.; PARAGUAI DE SOUZA, E.; HASHIMOTO
OKADA, L.Y.; DE A TRIVELLATO, F.A.; KAWASAKI, J.K.; COSTA, A.C.; REIS, A.B.;
GENARO, O.; BATISTA, L.M.; PALATNIK, M.; PALATNIK-DE-SOUSA, C.B. Effective
immunotherapy against canine visceral leishmaniasis with the FML-vaccine. Vaccine.
22 (17-18): 2234-43, 2004.
BOS, D.J & KASENBERG, M.L. The skin immune system. Immunology Today. 7: 235-240,
1986.
BOURDOISEAU, G.; BONNEFONT, C.; MAGNOL, J.P.; SAINT-ANDRE, I.; &
CHABANNE, L. Lymphocyte subset abnormalities in canine leishmaniasis. Vet.
Immunol. Immunopathol. 56, 345-351, 1997.
BRODSKYN, C.; DE OLIVEIRA, C.I.; BARRAL, A.; & BARRAL-NETTO, M. Vaccines in
leishmaniasis: advances in the last five years. Expert. Rev. Vaccines. 2, 705-717, 2003.
CALS-GRIERSON, M.M.; ORMEROD, A.D. Nitric oxide function in the skin. Nitric Oxide.
10: 179-193, 2004.
CARDOSO, L.; NETO, F.; SOUSA, J.C.; RODRIGUES, M.; CABRAL, M. Use of a
leishmanin test in the detection of canine Leismania-specific cellular immunity. Vet.
Parasitol. 79: 213-220, 1998.
CAVALCANTE, R.R.; PEREIRA, M.H.; & GONTIJO, N.F. Anti-complement activity in the
saliva of phlebotomine sand flies and other haematophagous insects. Parasitology 127, 8793. 2003.
CHARLAB, R., VALENZUELA, J. G., ROWTON, E. D., & RIBEIRO, J. M. Toward an
understanding of the biochemical and pharmacological complexity of the saliva of a
hematophagous sand fly Lutzomyia longipalpis. Proc. Natl. Acad. Sci. U.S.A. 96, 1515515160, 1999.
CHEN, L.; ZHANG, Z.; WATANABE, T.; YAMASHITA, T.; KOBAYAKAWA, T.;
KANEKO, A.; FUJIWARA, H.; SENDO, F. The involvement of neutrophils in the
resistance to Leishmania major infection in susceptible but not in resistance mice.
Parasitology International. 54: 109-118, 2005.
COHEN, S. Cell mediated immunity and the inflammatory system. Hum Pathol 7(3): 24964, 1976.
COX, F.E.G. Designer vaccines for parasitic diseases. International Journal of Parasitology.
27: 1147-1157, 1997.
COX, J.C & COUTER, A.L. Classification and review of their modes of action. Vaccine. 15:
248-256, 1997.
72
VITORIANO-SOUZA, J.
Referências Bibliográficas
DA SILVA, V.O.; BORJA-CABRERA, G.P.; CORREIA PONTES, N.N.; PARAGUAI DE
SOUZA, E.; LUZ, K.G.; PALATNIK, M.; PALATINIK DE SOUZA, C.B. A phase III trial
of efficacy of the FML vaccine agaisnt canine kala-azr in an endemic area of Brazil (São
Gonçalo ddo Amarante, RN). Vaccine, 19: 1082-1092, 2001.
DEANE, L.M.; DEANE, M.P. Leishmaniose visceral urbana (no cão e no homem) em
Sobral, Ceará. Hospital. 47: 75-87 ,1955.
DE LUNA, R.; VUOTTO, M.L.; IELPO, M.T.; AMBROSIO, R.; PIANTEDOSI, D.;
MOSCATIELLO, V.; CIARAMELLA, P.; SCALONE, A.; GRADONI, L.; & MANCINO,
D. Early suppression of lymphoproliferative response in dogs with natural infection by
Leishmania infantum. Vet,. Immunol. Immunopathol. 70:95-103, 1999.
DESJEUX, P. Leishmaniasis: current situation and new perspectives. Comp Immunol.
Microbiol. Infect. Dis. 27, 305-318, 2004.
DE SOUZA, R.C.A. - Estudo da cinética celular da resposta inflamatória causada pela
crioterapia experimental em pele normal de camundongos utilizando o método de
bolsas de ar no subcutâneo. Florianópolis. Tese Mestrado – Universidade Federal de
Santa Catarina, 2001.
DE OLIVEIRA, C.A.C.; PEREZ, A.C.; MERINO, G.; PRIETO, J.G.; ALVAREZ, A.I.
Protective effects of Panax ginseng on muscle injury and inflammation after eccentric
exercise. Comp. Biochem. 130 (3): 369-377, 2001.
DUNN, C.J.; HARDEE, M.M.; STAITE, N.D. - Acute and chronic inflammatory responses to
local administration of recombinant IL-1 alpha, IL-beta, TNF alpha, IL-2 and IFN
gamma in mice. Agents actions 27(3-4): 290-3, 1989.
EDELMAN, R. Adjuvants for the future. In: Levine, M.M.; Woodrow, G.C.; Kaper, J.B.;
Cobon, G.S. (eds.), New Generation Vaccines. Marcel Deker Inc., New York. 173-192,
1997.
EGAN, P.J.; KIMPTON, W.; SEOW, H.F.; BOWLES, V.M.; BRANDON, M.R.; NASH, A.D.
Inflammation-induced changes in the phenotype and cytokine profile of cells migrating
through skin e afferent lymph. Immunology 89(4): 539-46, 1996.
ENSERINK, M. Infectious diseases. Has leishmaniasis become endemic in the U.S.? Science.
290: 1881-1883, 2000.
FLORA FILHO, R.; ZILZERSTEIN, B. Óxido nítrico: o simples mensageiro percorrendo a
complexidade. Metabolismo, síntese e funções. Ver. Ass. Med. Brasil. 46 (3):265-71, 2000.
FRANCA-SILVA, J.C.; DA COSTA, R.T.; SIQUEIRA, A.M.; HADO-COELHO, G.L.; DA
COSTA, C.A.; MAYRINK, W.; VIEIRA, E.P.; COSTA, J.S.; GENARO, O., &
NASCIMENTO, E. Epidemiology of canine visceral leishmaniosis in the endemic area of
Montes Claros Municipality, Minas Gerais State, Brazil. Vet. Parasitol. 111, 161-173,
2003.
73
VITORIANO-SOUZA, J.
Referências Bibliográficas
FUJIWARA, R.T. Aspectos da imunidade celular e humoral da vacinação experimental na
leishmaniose visceral canina (Tese de Doutorado em Parasitologia). Belo Horizonte:
Instituto de Ciências Biológicas da Universidade Federal de Minas Gerais, 201p, 2003.
GARG, R.; DUBE, A. Animal models for vaccine studies for visceral leishmaniasis. Indian J.
Med. Res. 123: 439 – 454, 2006.
GENARO, O.; DA COSTA, C.A.; BREYNER, E.T.; REIS, A.B.; SILVA, A.R.; TROPIA,
M.J.M., TAFURI, W.L., DIAS, M., & MAYRINK, W. The course of experimental visceral
leishmaniasis in dogs. Mem. Inst. Oswaldo Cruz 87, 105, 1992.
GENARO, O. Leishmaniose visceral canina experimental (Tese de Doutorado em
Parasitologia). Belo horizonte: Instituto de Ciências Biológicas da Universidade Federal de
Minas Gerais. 202 p. Doutorado em Parasitologia, 1993.
GENARO, O.; DE, T.V; DA COSTA, C.A.; HERMETO, M.V.; AFONSO, L.C.; &
MAYRINK, W. Vaccine for prophylaxis and immunotherapy, Brazil. Clin. Dermatol. 14,
503-512, 1996.
GIUNCHETTI, R.C. Influência do parasitismo sobre os aspectos imunopatológicos na
leishmaniose visceral canina (Dissertação de Mestrado em Imunoparasitologia). Belo
Horizonte: Instituto de Ciências Biológicas da Universidade Federal de Minas Gerais, 101p.,
2004.
GIUNCHETTI, R.C.; MAYRINK, W.; GENARO, O.; CARNEIRO, C.M.; CORREIAOLIVEIRA, R.; MARTINS-FILHO, O.A.; MARQUES, M.J.; TAFURI, W.L.; REIS, A.B.
Relationship between canine visceral leishmaniosis and the Leishmania (Leishmania)
chagasi burden in dermal inflammatory foci. J. Comp. Pathol., 135:100-107, 2006.
GIUNCHETTI, R.C. Avaliação da imunogenicidade de dois novos imunobiológicos
candidatos a vacina contra Leishmaniose Visceral Canina (Dissertação de Doutorado em
Ciências Biológicas). Ouro Preto: Núcleo de Pesquisas em Ciências Biológicas, Universidade
Federal de Ouro Preto, 233p. 2007.
GIUNCHETTI, R.C.; CORRÊA-OLIVEIRA, R.; MARTINS-FILHO, O.A.; TEIXEIRACARVALHO, A.; ROATT, B.M.; AGUIAR-SOARES, R.D.O.; VITORIANO-SOUZA, J.;
MOREIRA, N.D.; MALAQUIAS, L.C.C.; GONTIJO, N.F. CASTRO, L.B.M.; LANA, M.;
REIS, A.B. Immunogenicity of a killed Leishmania vaccine with saponin adjuvant in
dogs. Vaccine. 25(44):7674-86, 2007.
GIUNCHETTI, R.C.; CORRÊA-OLIVEIRA, R.; MARTINS-FILHO, O.A.; TEIXEIRACARVALHO, A.; ROATT, B.M.; AGUIAR-SOARES, R.D.O.; COURA-VITAL, W.;
ABREU, R.T.; MALAQUIAS, L.C.C.; BRODSKYN, C.; OLIVEIRA, C.I.; COSTA, D.J.;
LANA, M.; REIS, A.B. A killed Leishmania vaccine with sand fly saliva extract and
saponin adjuvant displays immunogenicity in dogs. Vaccine. 26 (5): 623-638, 2008a.
GIUNCHETTI, R.C.; REIS, A.B.; DA SILVEIRA-LEMOS, D.; MARTINS-FILHO, O.A.;
CORRÊA-OLIVEIRA, R.; BETHONY, J.; VALE, A.M.; DA SILVA QUETZ, J.; BUENO,
L.L. FRANÇA-SILVA, J.C.; NASCIMENTO, E.; MAYRINK, W.; FUJIWARA, R.T.
74
VITORIANO-SOUZA, J.
Referências Bibliográficas
Antigenicity of a whole parasite vaccine as promising candidate against canine
leishmaniasis. Res. Vet. Sci. doi:10.1016/j.rvsc.2007.09.008. 2008b.
GRADONI, L.; SCALONE, A.; GRAMICIA, M. HIV-Leishmania co-infections in Italy:
serological data as a indication of the sequence of aquisition of two infections.
Transactions of Royal Society of Tropical Medicine and Hygiene 87:94-96, 1993.
GRADONI, L. An update on antileishmanial vaccine candidates and prospects for a canine
Leishmania vaccine. Vet. Parasitol. 100, 87-103, 2001.
HANDMAN, E. Leishmaniasis: Current Status of Vaccine Development. Clinical
Microbiology Reviews. 14(2): 229-243, 2001.
HALL L.R. & TITUS R.G. Sandfly vector saliva selectively modulates macrophage
functions that inhibit killing of L. major and nitric oxide production. J. Immunol. 155:
3501-3506, 1995.
HOMMEL, M.; JAFFE, C.L; TRAVI, B.; MILON, G. Experimental models for leishmaniasis
and for testing anti-leishmanial vaccines. Ann. Trop. Med. Parasitol. 89 (Suppl. 1): 55-73,
1995.
HOMEY, B.; STEINHOFF, M.; RUZICKA, T.; LEUNG, D.Y.M. Cytokines and chemokines
orchestrate atopic skin inflammation. J. Allergy Clin. Immunol. 118 (1): 178-189, 2006.
HOUDAYER, M.; ROUZE, P.; DALSGAARD, K. AND METZGER, J.J. Adjuvant Effect of
Quil A in porcine humoral immune response. Ann. Immunol. Cell. Biol. (Institute
Pauster), 129C, 197, 1978.
JANCAR, S. Imunidade natural e inflamação. In: Imunologia. CALISH,V. & VAZ, C. (Eds).
Editora Revinter, Rio de Janeiro, p.11-29, 2001.
KAIM, U.; MORITZ, A.; FAILING, K.; BAUMGÄRTNER, W. The regression of a canine
Langerhans cell tumour is associated with increased expression of IL-2, TNF-alpha,
IFN-gamma and iNOS mRNA. Immunology. 118(4):472-82, 2006.
KAMHAWI, S. The biology and immunomodulatory properties of sand fly and its role in
the establishment of Leishmania infections. Microbes Infect. 2: 1765-1773, 2000.
KENDALL, H.K.; MARSHALL, R.I.; BARTOLD, P.M. Nitric oxide and tissue destruction.
Oral Dis. 7 (1): 2-10, 2001.
KENSIL, C.R. Saponin as Vaccine Adjuvants. Critical Reviews in Therapeutic Drug Carrier
Systems. 13 (1&2): 1-55, 1996.
KUHN, A.; FEHSEL, K.; LEHMANN, P.; KRUTMANN, J.; RUZICKA, T.; KOLBBACHOVEN, V. Aberrant timing in epidermal expression of inducible nitric oxide
synthase after UV irradiation in cutaneous lupus erythematosus. J.Invest. Dermatol. 149153, 1998.
75
VITORIANO-SOUZA, J.
Referências Bibliográficas
LAGE RS, OLIVEIRA GC, BUSEK SU, GUERRA LL, GIUNCHETTI RC, CORREAOLIVEIRA R, REIS AB. Analysis of the cytokine profile in spleen cells from dogs
naturally infected by Leishmania chagasi. Vet. Immunol. Immunopathol. 115(1-2):135-45,
2007.
LAINSON, R.; SHAW J.J. Evolution, classification and geographical distribution. The
Leishmaniasis. London: Academic Press, 1–120, 1987.
LARREGINA, A.T. and L.D. FALO, Jr. Dendritcs cells in the context of skin immunity. In
Dendritcs Cells, 2nd Ed. M. T. Lotze and A.T. Thomson, eds. Academic Press. London. 301314, 2001.
LI, J.; CHEN, J. KIRSNER, R. Pathophysiology of Acute Wound Healing. Clinics in
Dermatology. 25: 9-18, 2007.
LIMA, G.M.A.C.; VALLOCHI, A.L.; SILVA, U.R.; BEVILACQUA, E.M.A.F.; KIFFER,
M.M.F.; ABRAHAMSOHN, I.A.. The role of polymorphonuclear leukocytes in the
resistence to cutaneous Leishmaniasis. Immunol Lett. 64: 145-151, 1998.
LIEW, F.Y.; O’DONNELL, C.A. Immunology of Leishmaniasis. Adv Parasitol 32: 161–259,
1993.
LIEW, F.Y.; WEI, X.Q.; PROUDFOOT, L. Cytokines and nitric oxide as effector molecules
against parasitic infections. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 352 (1359): 1311-5,
1997.
LUKES, J.; MAURICIO, I.L.; SCHÖNIAN, G.; DUJARDIN, J.; SOTERIADOU, K.; DEDET,
J.; KUHLS, K.; TINTAYA, K. W. Q.; JIRKU, M.; CHOCHOLOVA, E.; HARALAMBOUS,
C.; PRATLONG, F.; OBORNI´K, M.; HORA´K, A.; AYALA, F.J. AND MILES, A.M.
Evolutionary and geographical history of the leishmania donovani complex with a
revision of current taxonomy. PNAS. 104 (22): 9375-9380, 2007.
MAGALHAES, P.A.; MAYRINK, W.; DA COSTA, C.A.; MELO, M.N.; DIAS, M.; BATISTA,
S.M.; MICHALICK, M.S.; WILLIAMS, P. Kala-azar in the Rio Doce, Minas Gerais.
Results of prophylactic measures. Rev. Inst. Med. Trop. Sao Paulo 22, 197-202, 1980.
MALAQUIAS, L.C.; DO CARMO, R.R., DO AJ, J.R.; GIUNCHETTI, R.C.; CORREAOLIVEIRA, R.; REIS, A.B. Serological screening confirms the re-emergence of canine
leishmaniosis in urban and rural areas in Governador Valadares, Vale do Rio Doce,
Minas Gerais, Brazil. Parasitol. Res. 100, 233-239, 2007.
MARTINEZ-MORENO, A.; MORENO, T.; MARTINEZ-MORENO, F.J.; ACOSTA, I.;
HERNÁNDEZ, S. Humoral and cell-mediated immunity in natural and experimental
canine leishmaniasis. Vet. Immunol. Immunopathol. 48: 209-220, 1995.
MARZOCHI, M.C.A.; COUTINHO, S.G.; SOUZA, W.J.S.; TOLEDO, L. M.; GRIMALD, J.R.;
MOMEN, H.; PACHECO, R.S.; SABROZA, P.C.; SOUZA, M.A.; RANGEL JR, F.B.;
TRAMONTANO, N. Canine visceral leishmaniasis in Rio de Janeiro, Brazil. Clinical,
parasitological, therapeutical and epidemiological findings. (1977-1983). Mem. Inst.
Oswaldo Cruz. 80: 349-357, 1985.
76
VITORIANO-SOUZA, J.
Referências Bibliográficas
MAUEL, J. Vaccination against leishmania infections. Curr. Drug targets. Immune. Endocr.
Metabol. Disord. 2, 201-226, 2002.
MAURICIO, I.L.; HOWARD, M.K.; STOTHARD, J.R.; MILES, M.A. Genomic diversity in
the leishmania donovani complex. Parasitology 119 ( PT 3), 237-246, 1999.
MAURÍCIO, I.L.; STOTHARD J.R.; MILES M.A.. The strang case of Leishmania chagasi.
Parasitol. Today. 16:188-189, 2000.
MAYRINK, W.; DA COSTA, C.A.; MAGALHAES, P.A.; MELO, M.N.; DIAS, M.; LIMA,
A.O.; MICHALICK, M.S., & WILLIAMS, P. A field trial of a vaccine against American
dermal leishmaniasis. Trans. R. Soc. Trop. Med. Hyg. 73, 385-387, 1979.
MAYRINK, W.; WILLIAMS, P.; DA COSTA, C.A.; MAGALHAES, P.A.; MELO, M.N.;
DIAS, M.; OLIVEIRA, L.A.; MICHALICK, M.S.; FERREIRA, C.E.; BARROS, G.C. An
experimental vaccine against American dermal leishmaniasis: experience in the State of
Espirito Santo, Brazil. Ann. Trop. Med. Parasitol. 79, 259-269, 1985.
MAYRINK, W.; ANTUNES, C.M.; DA COSTA, C.A.; MELO, M.N.; DIAS, M.,
MICHALICK, M.S.; MAGALHAES, P.A.; DE OLIVEIRA, L.A.; & WILLIAMS, P.
Further trials of a vaccine against American cutaneous leishmaniasis. Trans. R. Soc.
Trop. Med. Hyg. 80, 1001, 1986.
MAYRINK, W.; GENARO, O.; FRANÇA-SILVA, J.C.; da COSTA, R.T.; TAFURI, W.L.;
ROTONDO, A.SILVA.; REIS, A. B.; WILLIAMS, P. & COSTA, C. A. Phase I and II
Open Clinical Trials of a Vaccine Against Leishmania chagasi Infections in Dogs. Mem
Inst Oswaldo Cruz, 80(6): 695-697, 1996.
MICHEL, T.; FERON, O. Nitric Oxide Syntases: which, where, how, and why? J. Clin.
Invest. 100: 2146-2152, 1997.
MINISTÉRIO DA SAÚDE. Manual de vigilância e controle da leishmaniose visceral,
Secretaria de Vigilância em Saúde, Brasília, Brasil, 122p. 2003.
MIRANDA, K.M.; ESPEY, M.G.; JOURD’HEUIL, D.; GRISHAM, M.B.; FUKUTO, J.M.;
FEELISCH, M.; WINK, D.A. The chemical biology of nitric oxide, in: L.J. Ignarro (Ed.),
Nitric Oxide: Biology and Pathobiology. Academic Press, San Diego, 41-55 p, 2000.
MOLERO, L.; GARCIA-DURAN, M.; DIAZ-RECASENS, J.; RICO, L.; CASADO, S.;
LOPEZ-FARRE, A. Expression of estrogen receptor subtypes and neuronal nitric oxide
syntase in neutrophils from women and men:regulation by estrogen. Cardiovasc. Res.
56: 43-51, 2002.
MOLINA, R.; AMELA, C.; NIETO, J.; SAN-ANDRES, M.; GONZALEZ, F.; CASTILLO,
J.A.; LUCIENTES, J.; ALVAR, J. Infectivity of dogs naturally infected with Leishmania
infantum to colonized Phlebotomus perniciosus. Trans. Roy. Soc. Trop. Med. Hyg. 88,
491-493, 1994.
77
VITORIANO-SOUZA, J.
Referências Bibliográficas
MONCADA, S.; HIGGS, A. The L-arginine-Nitric Oxide Pathway. New Eng. J. Med. 329
(27):2002-12, 1993.
MONCADA, S.; ERUSALIMSKY, J.D. Does nitric oxide modulate mitochondrial energy
generation andapoptosis? Nat Rev Mol Cell Biol. 3 (3): 214-20, 2002.
MORENO, J; ALVAR, J. Canine Leishmaniasis: epidemiological risk and the experimental
model. Trends in Parasitology. 18(9): 399-405, 2002.
MOREIRA, N.D. Cinética da migração celular intradérmica em hamsters inoculados com
diferentes constituintes antigênicos de duas vacinas contra Leishmaniose Visceral
Canina. (Tese de mestrado). Núcleo de Pesquisas em Ciências Biológicas, Universidade
Federal de Ouro Preto, Ouro Preto, 2008.
MULLER, W.A. Leucocyte – endothelial – cell interations in leucocyte transmigration and
the inflammatory response. Trends in Immunology. 24(6): 326-333, 2003.
NANDAN, D.; LO, R.; REINER, N.E. Activation of phosphotyrosine phosphatase activity
attenuates mitogen-activated protein kinase signaling and inhibits c-FOS and nitric
oxide synthase expression in macrophages infected with Leishmania donovani. Infect.
Immun. 67 (8): 4055-63. 1999.
NATHAN, C.; XIE, Q. Nitric oxide synthases: Roles, tolls, and controls. Cell. 78: 915-918,
1994a.
NATHAN, C.; XIE, Q. Regulation of biosynthesis of nitric oxide. J. Biol. Chem. 269: 1372513728, 1994b.
NATHAN, C.; SHILOH, M.U. Reactive oxygen and nitrogen intermediates in the
relationship between mammalian hosts and microbial pathogens. Proc. Natl. Acad. Sci.
97: 8841-8848, 2000.
NOLI, C.; AUXILIA, S.T. Treatment of canine Old World visceral leishmaniasis: a
systematic review. Vet Dermatol. 16 (4): 213-32, 2005.
ODA, K.; MATSUDA, H.; MURAKAMI, T.; KATAYAMA, S.; OHGITANI, T.;
YOSHIKAWA, M. Adjuvant and haemolytic activities of 47 saponins derived from
medicinal and food plants. Biol. Chem. 381 (1): 67-74, 2000.
O'HAGAN, D.T. & VALIANTE, N.M. Recent advances in the discovery and delivery of
vaccine adjuvants. Nat. Rev. Drug discov. 2: 727-735, 2003.
OLIVEIRA, S.H.P.; FONSECA, S.G.; ROMÃO. P.R.T.; FERREIRA, S.H.; CUNHA, F.Q.
Nitric oxide mediates the microbicidal activity of eosinophils. Mem. Inst. Osw. Cruz. 92:
233-235, 1997.
OLIVEIRA, S.H.P.; FONSECA, S.G.; ROMÃO, P.R.T.; FIGUEIREDO, F.; FERREIRA, S.H.;
CUNHA, F.Q. Microbiology activity of eosinophils is associated with acitivation of the
arginine-no pathway. Parasite Immunol 20: 405-412, 1998.
78
VITORIANO-SOUZA, J.
Referências Bibliográficas
PARHAM, P. ed. - O sistema immune. Artmed, Porto Alegre. 202-240 pp., 2001
PALATNIK, C.B.; PREVIATO, J.O.; MENDONÇA-PREVIATO, L.; BOROJEVIC, R. A new
approach to phylogeny of Leishmania: species-specificity of glycoconjugate ligands for
promastigote internalization into murine macrophages. Parasitol. Research. 76: 289-293,
1990.
PALATNIK DE SOUSA, C.B.; MORENO, M.B; PARAGUAI DE SOUZA, E.; BOROJEVIC,
R. The FML vaccine (fucose-mannose ligand) protects hamsters from experimental
kala-azar. Cienc. Cult. (J. Braz. Assoc. Adv. Sci.). 46:290-296, 1994a.
PALATNIK DE SOUSA, C.B.; PARAGUAI DE SOUZA, E.; GOMES, E.M.; BOROJEVIC, R.
Experimental murine Leishmania donovani infection immunoprotection by fucose
mannose ligand (FML). Braz. J. Med. Biol. Res. 27: 547-551, 1994b.
PARANHOS, M.W.; DOS SANTOS, I..; SHERLOCK, G.; OLIVEIRA, AND DE
CARVALHO, L. Development of eosinophilia in dogs intradermically inoculated with
sand fly saliva and Leishmania (Leishmania) chagasi staionary-phase promastigotes.
Mem. Inst. Oswaldo Cruz 88: 249–251, 1993.
PARRA, L.E.; BORJA-CABRERA, G.P.; SANTOS, F.N.; SOUZA, L.O.P.;
PALATNIK-DE-SOUSA, C.B.; MENZ, I. Safety trial using the Leishmune®
vaccine against canine visceral leishmaniasis in Brazil. Vaccine. 25 2180–2186,
2007.
PEARSON RD, UYDESS IL, CHAPMAN SW, STEIGBIGEL RT. Interaction of human
eosinophis with Leishmania donovani. Ann. Trop. Med. Parasitol. 81: 735-739, 1987.
PEARSON, R.D. Pathology of Leishmaniasis. In Warren KSS (ed). Immunology and
molecular, 1993.
PETERS, W.; KILLICK-KENDRICK, R.. The Leishmaniasis in Biology and Medicine.
Academic Press, New York: 941, 1987.
PINELLI, E.; GONZALO, R.M; BOOG, C.J.P.; RUTTEN, V.P.M.G., DEL REAL, G.;
RUITENBERG, J. Leishmania infantum-specific T cell lines derived from asymptomatic
dogs that lyse infected macrophages in a major histocompatibility complex- restricted
manner. Eur. J. Immunology. 25: 1594-1600, 1994.
PINELLI, E.; KILLICK-KENDRICK, R.; WAGENAAR, J.; BERNADINA, W., DEL REAL,
G.; RUITENBERG, E.J. Cellular and humoral responses in dogs experimentally and
naturally infected with Leishmania infantum. Infect. Immun. 62: 229-235, 1995.
POTTER, N.S.; HARDING, C.V. Neutrophils process exogenous bacteria via an alternative
class I MHC processing pathway for presentation of peptide to T lymphocytes. J.
Immunol. 167: 2538-2546, 2001.
QADOUMI, M.; BECKER, I.; DONHAUSER, N.; RÖLLINGHOFF, M.; BOGDAN, C.
Expression of inducible nitric oxide synthase in skin lesions of patients with American
Cutaneous Leishmaniasis. Infection and Immunity. 70 (8): 4638-4642, 2002.
79
VITORIANO-SOUZA, J.
Referências Bibliográficas
RAVINDRAN, R. & ALI, N. Progress in vaccine research and possible effector mechanisms
in visceral leishmaniasis. Curr. Mol. Med. 4, 697-709, 2004.
REIS, A.B. Avaliação de parâmetros laboratoriais e imunológicos de cães naturalmente
infectados com Leishmania (Leishmania) chagasi, portadores de diferentes formas
clínicas da infecção (Tese de Doutorado em Ciências). Belo Horizonte: Instituto de Ciências
Biológicas da Universidade Federal de Minas Gerais, 180p. 2001.
REIS, A.B.; CARNEIRO, C.M.; CARVALHO, M.G.; TEIXEIRA-CARVALHO, A.;
GIUNCHETTI, R.C.; MAYRINK, W.; GENARO, O.; CORREA-OLIVEIRA, R., &
MARTINS-FILHO, O.A. Establishment of a microplate assay for flow cytometric
assessment and it is use for the evaluation of age-related phenotypic changes in canine
whole blood leukocytes. Vet. Immunol. Immunopathol. 103, 173-185, 2005.
REIS, A.B.; MARTINS-FILHO, O.A.; TEIXEIRA-CARVALHO, A.; CARVALHO, M.G.;
MAYRINK, W.; FRANÇA-SILVA, J.C.; GIUNCHETTI, R.C.; GENARO, O.; CORRÊAOLIVEIRA, R. Parasite density and impaired biochemical/hematological status are
associated with severe clinical aspects of canine visceral leishmniasis. Research in
Veterinary Science.81: 68-75, 2006a.
REIS, A.B.; TEIXEIRA-CARVALHO, A.; VALE A.M.; MARQUES, M.J. GIUNCHETTI, R.
C.; MAYRINK, W.; GUERRA, L.L.; ANDRADE, R.A.; CORREIA-OLIVEIRA, R.;
MARTINS-FILHO, O.A. Isotype patterns of immunoglobulins: hallmarks for clinical
status and tissue parasite density in Brazilian dogs naturally infected by Leishmania
(Leishmania) chagasi. Vet. Immunol. Immunopathology. 112, 102-116, 2006b.
REIS A.B.; TEIXEIRA-CARVALHO A.; GIUNCHETTI R.C.; GUERRA L.L.; CARVALHO
M.G.; MAYRINK W.; GENARO, O.; CORREA-OLIVEIRA, R.; MARTINS-FILHO, O.A.
Phenotypic features of circulating leucocytes as immunological markers for clinical
status and bone marrow parasite density in dogs naturally infected by Leishmania
chagasi. Clin Exp Immunol. 146(2):303-11, 2006c.
REQUENA, J. M., IBORRA, S., CARRION, J., ALONSO, C., & SOTO, M. Recent advances in
vaccines for leishmaniasis. Expert. Opin. Biol. Ther. 4, 1505-1517, 2004.
REZENDE, F.C.B. Estudo da cinética do infiltrado celular inflamatório induzido por
adjuvantes de vacinas em bolhas de ar formadas no subcutâneo de camundogos.
Universidade Federal de Santa Catarina. Programa de Pós-Graduação em Biotecnologia
(mestrado), Florianópolis, 2003.
RIBEIRO, J.M.C.; ROSSIGNOL, P.A.; SPIELMAN, A. Blood-finding strategy of capillaryfeeeding sandfly, Lutzomyia longipalpis. Comp. Biochem. Physiol. A. 83: 683-686, 1986.
ROSS, R.; RESKE-KUNZ, A.B. The role of NO in contact hypersensitivity. International
Immunopharmacology. 1469–1478, 2001.
RYAN, G.B.; MAJNO, G. Acute inflammation. Am. J. Pathol. 86: 185-274, 1997.
80
VITORIANO-SOUZA, J.
Referências Bibliográficas
SANTOS-GOMES, G.M.; ROSA, R.; LEANDRO, C.; CORTES, S.; ROMÃO, P.; SILVEIRA,
H. Cytokine expression during the outcome of canine experimental infection by
Leishmania infantum. Vet. Immunol. Immunopathol. 88: 21-30, 2002.
SANZ, M.J.; MARINOVA-MUTAFCHIEVA, L.; GREEN, P.; LOBB, R.R.; FELDEMANN,
M.; NOURSHARGH, S. IL-4 induced eosinophil accumulation in rat skin is dependent
on endogeous TNF-alpha and alpha 4 integrin/VCAM-1 adhesion pathways. J.
Immunol. 160: 5637-5645, 1998.
SARAIVA, N.G.; VALDERRAMA, L.; LABRADA, M.; HOLGUIN A.F.; NAVAS, C.;
PALMA, G.; WEIGLE, K.A. The relationship of Leishmania braziliensis subspecies and
immune response to disease expression in New World leishmaniasis. J. Infec. Dis., v.
159, p. 725-735, 1989.
SCAPINI, P.; LAPINET-VERA, J.A.; GASPERINI, S.; CALZETTI, F., BAZZONI, F.;
CASSATELLA, M.A. The neutrophil as a cellular source of chemokines. Immunological
Reviews. 177, 195-203, 2000.
SHIMIZU, Y.; SAKAI, M.; UMEMURA, Y.; UEDA, H. Immunohistochemical Localization
of Nitric Oxide Synthase in Normal Human Skin: Expression of Endothelial-type and
Inducible-type Nitric Oxide Synthase in Keratinocytes. The Journal of Dermatology., v.
24: 80-87, 1997.
SESARDIC, D. Regulatory considerations on new adjuvants and delivery systems. Vaccine
24S2. S2/86–S2/87, 2006.
SILVA, E.S.; GONTIJO, C.M.; PACHECO, R.S.; FIÚZA, V.O.; BRAZIL, R.P. Visceral
Leishmaniasis in the Metropolitan Region of Belo Horizonte, State of Minas Gerais,
Brazil. Mem. Inst. Oswaldo Cruz. 96: 285-291, 2000.
SILVA, F.; GOMES, R.; PRATES, D.; MIRANDA, J.C.; ANDRADE, B.; BARRAL-NETO,
M.; BARRAL, A. Inflammatory cell infiltration and high antibody production in
BALB/c mice caused by natural exposure to Lutzomyia longipalpis bites. Am. J. Trop.
Med. Hyg.. 72: 94-8, 2005.
SINGH, M.; O’HAGAN, D.T. Invited review recent advances in veterinary vaccine
adjuvants. Int. J. Parasitol. 33 (5-6): 469-478, 2003.
SLAPPENDEL, R.J.; GREENE, C.E.; APUD CARDOSO, L.; CABRAL, M. Leishmaniasis
and canine leishmaniasis. Revista Portuguesa de Ciências Veterinária. XCIII (527). 122141, 1998.
SOLANO-GALLEGO, L.; LULL, J.; RAMOS, G., et al. The Ibizian hound presents a
predominantly cellular immune response against natural Leishmania infection. Vet.
Parasitol. 90: 37-45, 2000.
SOEHNLEIN, O., KENNE, E.; ROTZIUS, P.; ERIKSSON, E.E.; LINDBOM, L. Neutrophil
secretion products regulate anti-bacterial activity in monocytes and macrophages. Clin
Exp Immunol. 151 (1): 139-45, 2008.
81
VITORIANO-SOUZA, J.
Referências Bibliográficas
SPRINGER, T.A. Traffic signals on endonthelium for lymphocyte recirculation and
leukocyte emigration. Annu. Rev. Physiol. 57: 827-872, 1995.
STUEHR, D.J.; CHO, H.J.; KWON, N.S.; WEISE, M.F.; NATHAN, C.F. Purification and
characterization of the cytokine-induced macrophage nitric oxide synthase: an FADand FMN- cointaining flavoprotein. Proc. Natl. Acad. Sci. USA, 88: 7773-7777, 1991.
STWART, R.J.; HOLLOWAY, L.J.; ISBISTER, W.H. Peritoneal neutrophilia: a potential
indicator of the surgical acute abdomen. Aust N Z J Surg 54(6):565-8, 1984.
TAFURI, W.L. Histopatologia da Intradermorreção de Montenegro no cão em várias
condições experimentais: Cinética da Reação e Estudo Comparativo com os Antígenos
Leishavacin e P10.000G. (Tese de mestrado). Faculdade de Medicina, Universidade Federal
de Minas Gerais, Belo Horizonte, 1992.
TAFURI, W.L.; RASO, P.; HERMETO, M.V.; VIEIRA-DIAS, D.; MAYRINK, W.
Comparative histopathologic study of the skin test in dogs from an endemic area of
tegumentary leishmaniasis, using 2 antigens: Leishvacin and P10.000G. Rev. Soc. Bras.
Méd. Trop. 26(1): 11-14; 1993.
TEIXEIRA, C.R.; TEIXEIRA, M.J.; GOMES, R.B.B.; SANTOS, C.S.; ANDRADE, B.B.;
RAFFAELE-NETO, I.; SILVA, J.S.; GUGLIELMOTTI, A.; MIRANDA, J.C.; BARRAL,
A.; BRODSKYN, C.; BARRAL-NETO, M. Saliva from Lutzomyia longipalpis Induces
CC Chemokine Ligand 2/Monocyte Chemoattractant Protein-1 Expression and
Macrophage Recruitment. The Journal of Immunology. 175: 8346-8353, 2005.
TERUI, T.; OZAWA, M.; TAGAMI, H. Role of neutrophils in induction of acute
inflammation in T-cell-mediated immune dermatosis, psoriasis: a neutrophil-associated
inflammation-boosting loop. Exp Dermatol 9(1): 1-10, 2000.
THEODOS, C.M. AND R.G. TITUS. Salivary gland material from the sand fly Lutzomyia
longipalpis has an inhibitory effect on macrophage function in vitro. Parasite Immunol.
15: 481–487, 1993.
TITUS, R.G. AND RIBEIRO, J.. The role of vector saliva in transmission of arthropodborne diseases. Parasitol. Today 6: 157–160, 1990.
TITUS, R.G.; RIBEIRO, J.M. Salivary gland lysates from the sand fly Lutzomyia longipalpis
enhace Leishmania infectivity. Science. 239: 1306-1308, 1988.
TRAVI, B.L.; TABARES, C.J.; CADENA, H.; FERRO, C.; OSORIO, Y. Canine visceral
leishmaniasis in Colômbia: relationship between clinical and parasitologic status and
infectivity for sand flies. Am. J. Trop. Med. Hyg. 64: 119-124, 2001.
TSAGOZIS P.; KARAGOUNI, E.; DOTSIKA, E. CD8(+) T cells with parasite-specific
cytotoxic activity and a Tc1 profile of cytokine and chemokine secretion develop in
experimental visceral leishmaniasis. Parasite Immunol. 25: 569-579, 2003.
VALENZUELA, J.G. High-throughput approaches to study salivary proteins and genes
from vectors of disease. Insect Biochemistry and Molecular Biology. 32:1199–1209, 2002.
82
VITORIANO-SOUZA, J.
Referências Bibliográficas
VAN GISBERGEN, K.P.J.M.; GEIJTENBEEK, T.B.H.; VAN KOOYK, Y. Close encounters
of neutrophils and DCs. Trends in Immunology. 26 (12), 2005.
VOGEL, F.R.; POWELL, M.F. A compendium of vaccine adyuvants and excipients. In:
Powell, M.F., Neuman, M.J. (Eds.), Vaccine Design, the subunit and adyuvants approach.
Plenum Press, New York. 141-228, 1995.
VOULDOUKIS, I.; BECHEREL, P.A.; RIVEROS-MORENO, V.; AROCK, M.; DA SILVA,
O.; DEBRE, P.; MAZIER, D.; MOSSALAYI, M.D. Interleukin- 10 and interleukin-4
inhibit intracellular killing of Leishmania infantum and Leishmania major by human
macrophages by decreasing nitric oxide generation. Eur J Immunol 27: 860–865, 1997.
WATANABE, Y.; HAMAGUCHI-TSURU, E.; MORIMOTO, N.; NISHIO, Y.; YAGYU, K.;
KONISHI, Y.; TOMINAGA, M.; MIYAZAKI, J.; FURUYA, M.; TOMINAGA, A. IL-5Induced Eosinophils Suppress the Growth of Leishmania amazonensis In Vivo and Kill
Promastigo6tes In Vitro in Response to Either IL-4 or IFN-gamma. DNA Cell Biol. 23:
412-418, 2004.
WORLD HEATH ORGANIZATION. Magnitude of the problem. Avaible:
http://www.who.int/leishmaniasis/burden/magnitude/burden_magnitude/en/index.ttml, 2005.
83