PIBIC-UFU, CNPq & FAPEMIG
Universidade Federal de Uberlândia
Pró-Reitoria de Pesquisa e Pós-Graduação
DIRETORIA DE PESQUISA
UTILIZAÇÃO DE PARÂMETROS MORFOANATÔMICOS NA ANÁLISE
DE PLANTAS DE SOJA (GLYCINE MAX (L.) MERRIL) CULTIVADAS EM
AMBIENTE ENRIQUECIDO COM CO2
Elisa Rodrigues Gontijo1
Universidade Federal de Uberlândia
[email protected]
Neuza Maria de Castro2
Universidade Federal de Uberlândia
[email protected]
Luzimar Campos Silva3
Universidade Federal de Viçosa
[email protected]
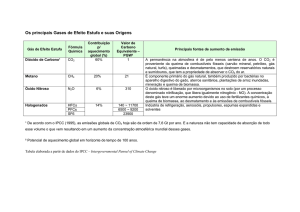
Resumo: A concentração de CO2 atmosférico está aumentando e provocando alterações
fisiológicas e anatômicas nas plantas e estudos sobre como as plantas reagem a estas mudanças
são de vital importância. O presente estudo teve como objetivo avaliar as possíveis alterações
anatômicas na folha de Glycine max (L.) Merril, quando expostas a altas concentrações de CO2,
numa tentativa de contribuir com informações sobre os efeitos causados pelo aumento da
concentração de CO2. Foram analisadas duas linhagens de Glycine max (L.) Merril (soja)
submetidas à concentração atual de CO2 atmosférico encontrado na natureza (≈360 µmol mol-1), e
ao dobro da concentração atual de CO2 (≈720 µmol mol-1). A densidade estomática, a altura da
epiderme, nas duas faces da folha, a espessura do parênquima clorofiliano paliçádico, do
parênquima clorofiliano lacunoso e do parênquima paravenal não apresentaram alterações em
relação à alta concentração de CO2. Foram encontrados estômatos com as células subsidiárias
“inchadas” em folhas desenvolvidas sob alta concentração de CO2. A endogamia das linhagens
analisadas e o controle das variáveis que afetam o desenvolvimento da planta podem estar
relacionados com os resultados encontrados.
Palavras-Chave: anatomia foliar, densidade estomática, estômato anômalo.
1. INTRODUÇÃO
O ciclo do carbono na atmosfera tem sido alterado pela atividade humana nos últimos 150
anos (Pinto et al., 2000). O CO2 emitido pela ação antrópica é da ordem de 8,5 bilhões de toneladas
anuais, e como conseqüência, a concentração de CO2 atmosférico está aumentando gradativamente
(Pinto et al., 2000). Apesar de não existirem estudos conclusivos mostrando a contribuição relativa
das atividades antrópicas ou dos fenômenos naturais na elevação dos níveis de CO2 atmosférico, o
aumento em 25% desse gás na atmosfera, desde a Revolução Industrial suscita a associação entre o
aumento do consumo de combustíveis fósseis e o aumento da concentração de CO2, e dos outros
gases denominados gases do efeito estufa na atmosfera (Lobo, 2003). Segundo Sasaki e
colaboradores (2006), a concentração de CO2 na atmosfera global muito em breve chegará a
aproximadamente 380 µmol mol-1, e poderá alcançar algo próximo a 550-600 µmol mol-1 até final
desse século.
1
Graduada em Ciências Biológicas, Uberlândia-MG.
Professora Associada N1 da Universidade Federal de Uberlândia, Instituto de Biologia.
3
Professora Adjunta da Universidade Federal de Viçosa, Departamento de Biologia Vegetal.
2
Morrisey e Justus (1999) acreditam que um aumento de duas vezes na concentração de CO2
atmosférico levaria a um incremento na temperatura média do globo terrestre, da ordem de 3-4o C,
até o final do século XXI. Assim, uma maior compreensão sobre os efeitos causados pelo CO2 nas
plantas seria de vital importância (Bazzaz, 1990; Bowes, 1993).
Os poluentes, tanto sob a forma líquida como gasosa, aderem à superfície das plantas ou
penetram nas folhas através da cutícula e/ou estômatos, provocando respostas fisiológicas e/ou
estruturais que dependem de uma série de fatores, como, por exemplo: idade da planta e/ou do
tecido exposto, a concentração do poluente, o tempo de exposição, a densidade de tricomas e a
espessura da parede das células (Dickison, 2000).
Para avaliação do impacto causado pela poluição atmosférica na vegetação, o órgão mais
estudado tem sido a folha, abordando-se, principalmente, aspectos como: a abscisão precoce,
manchas, cloroses, necroses e taxas de herbivoria (Domingos et al., 1998). Essas alterações
estruturais, citológicas e/ou ultraestruturais, provocadas pelos poluentes atmosféricos, têm sido
preciosas na avaliação do impacto, causado por esses poluentes, principalmente, porque podem
detectar a degradação de uma área, mesmo quando as injúrias ainda não são visíveis
macroscopicamente (Mazzoni-Viveiros, 2000).
Estudos anatômicos realizados por Alves e colaboradores (2001) com um clone híbrido de
Tradescantia, cultivado na cidade de São Paulo, sob poluição aérea, evidenciaram que na superfície
abaxial da folha, os estômatos das plantas-controle eram maiores que aqueles das plantas expostas
aos poluentes aéreos. Esses mesmos estudos constataram ainda, hipertrofia e hiperplasia do
parênquima clorofiliano lacunoso das plantas expostas à poluição. Essas observações são
importantes indicadores para a avaliação dos efeitos dos poluentes atmosféricos sobre as plantas, na
cidade de São Paulo.
Segundo Azevedo (1995), a epiderme adaxial de folhas de soja submetidas à chuva simulada
com flúor é, na maioria das vezes, a mais afetada apresentando redução acentuada na expansão de
suas células; ocorre também redução nas células do parênquima paliçádico, o que implica na
desorganização do parênquima lacunoso, com aumento no número de camadas e/ou comprimento
das células, alterando o relevo da face abaxial da folha.
No entanto, várias pesquisas têm demonstrado que o desenvolvimento das plantas e sua
produtividade estão diretamente relacionados com o enriquecimento de CO2 (Lin et al., 2001). Altas
concentrações de CO2, frequentemente levam a um aumento da área e volume foliar, bem como, da
razão área/volume foliar (Ceulemans et al., 1997; Norby et al., 1999 apud Lin et al., 2001) e
provocam o aumento da fotossíntese, do crescimento e do rendimento das espécies vegetais (Rogers
et al., 1983; Cure et al., 1986; Kimball, 1986; Bazzaz, 1990 apud Heagle et al., 2002).
No Brasil, tem-se verificado aumento significativo do uso de ambientes protegidos para o
cultivo de hortaliças, principalmente nas regiões Sul e Sudeste, aliado à aplicação de dióxido de
carbono nas plantas, via água de irrigação (Furlan et al., 2002). Segundo estes autores, esta técnica
de enriquecimento da concentração de CO2 tem provocado aumento significativo na produção de
pimentão (cultivar Mayata), possibilitando maior comprimento, diâmetro e número de frutos por
planta, além de aumento da massa e do rendimento (Furlan et al., 2002). Entretanto, efeitos
negativos da “fertilização” com CO2 incluem o carregamento excessivo dos cloroplastos com amido
ou hipertrofia dos tecidos foliares (Larcher, 2000).
Sasaki e colaboradores (2006), também observaram que para o arroz (Oryza sativa L.) há
um aumento da produção com o aumento na concentração de CO2, de acordo com o estudo de.
Canizares e colaboradores (2004) observaram diferenças nas trocas gasosas, transpiração e
condutância estomática, em híbridos de pepino do tipo japonês cultivados em duas épocas do ano,
irrigadas com água comum e enriquecida com CO2.
A aplicação de dióxido de carbono no sistema de irrigação sobre a cultura do tomate
(cultivar Débora-Plus), também proporcionou aumento na produtividade (8,2%) e no peso de frutos
(13%) (Cararo; Duarte, 2002). Além disso, verificaram através de análises da solução do solo, que o
uso de CO2 concentrado parece contribuir para a melhoria das condições nutricionais do tomateiro.
Pinto e colaboradores (2001) observaram aumento de produtividade em cultura de melão (Cucumis
2
melo) submetida à aplicação de dióxido de carbono via água de irrigação, sem alterar a qualidade do
fruto nem os teores de nutrientes das folhas de melão. Pandey e colaboradores (2007) relataram um
aumento significativo na densidade estomática (68,7%), em cultivares de rosa (Rosa hybrida)
mantidos sob alta temperatura e enriquecimento de CO2, quando comparadas com as folhas de
indivíduos cultivados sob temperatura ótima. Utilizando enriquecimento CO2 de 1000 µmol mol-1,
as cultivares de rosa também apresentaram mudanças na abertura estomática.
No entanto, ainda se desconhece como o CO2 afeta o desenvolvimento das folhas (Lin et al.,
2001). Alguns trabalhos têm mostrado que a elevação de CO2 produz algumas alterações na
anatomia foliar (Wullschleger et al., 1997 apud Lin et al., 2001), como um aumento da densidade
estomática ou amento do parênquima paliçádico, mas ainda permanecem dúvidas se as alterações se
devem apenas à elevação de CO2 ou se outros fatores estariam envolvidos.
Atualmente, a cultura da soja (Glycine max (L.) Merril) encontra-se em destaque no cenário
nacional, devido à rentabilidade que proporciona ao produtor rural (Martin et al., 2005), tanto como
fonte de proteína vegetal de alta qualidade, como produto de exportação (Lobo, 2003). A produção
mundial de soja encontra-se na faixa de 236 milhões de toneladas, sendo 58,4 milhões de toneladas
produzidas no Brasil, ocupando mais de 20 milhões de hectares (Embrapa Soja, 2008).
As folhas de soja são tipicamente dorsiventrais, anfiestomáticas, e os estômatos paracíticos
ocorrem em menor número na face adaxial. A epiderme das duas faces da folha apresenta tricomas
tectores e glandulares (Azevedo, 1995; Mussury et al., 2007). O mesofilo é formado por duas
camadas de parênquima paliçádico, duas de parênquima lacunoso e entre estes uma camada de
parênquima paravenal (Azevedo, 1995). Os feixes vasculares são colaterais, envolvidos por uma
bainha de células parenquimáticas e os feixes de maior calibre podem apresentar extensões de
bainha parenquimática conectando-os à epiderme (Mussury et al., 2007).
Dado o seu valor econômico e de sua potencialidade de cultivo em diversas condições, a
cultura dessa leguminosa tem sido objeto de constante investimento em novas tecnologias na
agricultura (Bulbovas et al., 2007).
Em 2003, Lobo cultivou duas linhagens de soja, em casa-de-vegetação da Unidade de
Crescimento de Plantas da Universidade Federal de Viçosa, em Viçosa (MG - Brasil), sob elevadas
concentrações de CO2 atmosférico, durante todo seu ciclo de vida. Essas linhagens apresentaram em
média, um incremento de 46% na taxa fotossintética e de 58% na massa seca total. Segundo o autor,
a condutância estomática não foi afetada pelas concentrações de CO2.
A folha é o principal órgão envolvido com a fotossíntese e a transpiração, deste modo, sua
morfologia e anatomia influenciam diretamente o processo fotossintético (Parkhurst, 1986 apud Lin
et al., 2001), e, portanto a produtividade de uma espécie. O presente estudo tem como objetivo
avaliar se ocorrem alterações anatômicas na lâmina foliar de Glycine max (L.) Merril, quando
expostas a alta concentração de CO2, visando contribuir com informações sobre os efeitos causados
pelo aumento da concentração de CO2 sobre esta espécie.
2. MATERIAL E MÉTODOS
2.1 Material Vegetal e Condições de Cultivo
O material vegetal utilizado foi coletado dos indivíduos cultivados por Lobo (2003). As
linhagens de Glycine max (L.) Merril (soja) estudadas foram desenvolvidas pelo Programa de
Melhoramento de Qualidade da Soja do Instituto de Biotecnologia Aplicado à Agricultura
(BIOAGRO, da Universidade Federal de Viçosa - UFV, Viçosa, MG). Foram selecionadas
linhagens de uma população F8, de 118 linhagens endogâmicas recombinantes provenientes de
cruzamentos entre as variedades Barc-8 e Garimpo.
As plantas foram cultivadas em casa de vegetação da Unidade de Crescimento de Plantas da
UFV. As linhagens de soja foram semeadas em vasos de 5 L, com substrato constituído de uma
mistura de solo e areia, na proporção de 3:1, e em seguida, transferidas para 6 câmaras metálicas de
topo aberto (Heagle et al, 1973; Mandle et al, 1973), mantidas em casa de vegetação. Três das
3
câmaras foram submetidas à concentração atual de CO2 atmosférico encontrado na natureza (≈360
µmol mol-1), e as outras três, à concentração de CO2 atmosférico duplicada (≈720 µmol mol-1),
seguindo o método proposto por Olivo (2000).
As plantas foram submetidas aos diferentes tratamentos de concentração de CO2, desde a
semeadura até a colheita. O monitoramento das concentrações de CO2 foi feito com auxílio de um
aparelho coletor modelo WA-161 MK3-A (ADC, UK) e um analisador de gás infravermelho
(IRGA) modelo 225-MK3 (ADC, UK) adotado por Lobo (2003).
Após três meses de cultivo, foi realizada uma coleta aleatória de folhas das plantas dos dois
tratamentos: sendo 3 folhas de cada planta, de cada câmara, perfazendo um total de 36 folhas por
linhagem, que em seguida foram fixadas em FAA (formaldeído 37%, acido acético glacial e etanol
50%, 1:1:18, v/v), conforme Johansen (1940) e conservadas em etanol 70%.
2.2 Análise micromorfométrica
Parte do material fixado foi incluído em historesina (Leica), desidratado em série etílica e
sua inflitração e inclusão seguiram as instruções do fabricante. Foram preparados 3 blocos por
repetição, totalizando 9 blocos por cultivar e 18 blocos por tratamento.
Foram feitos cortes transversais de 6 µm de espessura com o auxílio de um micrótomo
rotativo de avanço automático (modelo RM2155, Leica Microsystems Inc). As lâminas histológicas
com os cortes foram coradas com azul de toluidina pH 4,0 (O’ Brien et al., 1965 apud Kraus e
Arduin, 1997) e montadas em Permount (Kraus e Arduin, 1997).
Desprezando-se a nervura mediana, foram mensuradas: a altura das células epidérmicas da
face adaxial e da face abaxial; do parênquima clorofiliano paliçádico; parênquima clorofiliano
lacunoso e a espessura total do limbo, utilizando-se o software Anati Quanti 2, desenvolvido por
Aguiar et al. (2007) no Laboratório de Anatomia Vegetal da Universidade Federal de Viçosa.
Foram realizadas 135 medidas/repetição/parâmetro, totalizando 405 medidas por tratamento
e 810 medidas por linhagem. As observações anatômicas e a documentação fotográfica foram
realizadas em fotomicroscópio modelo Olympus AX70TRF, Olympus Optical, com sistema UPhoto (Fig.4), no Laboratório de Anatomia Vegetal.
O experimento foi executado em um delineamento inteiramente casualizado, em esquema de
parcelas divididas, ficando na parcela principal a concentração atmosférica de CO2 e na subparcela
a linhagem, sendo utilizadas três repetições por tratamento combinado. Os dados foram submetidos
à Análise de Variância e as médias, foram comparadas pelo Teste de Tukey, a 5% de probabilidade
(p≤0.05).
2.3 Densidade Estomática
Para a realização dos cálculos de densidade estomática, pequenos fragmentos da lâmina foliar
foram diafanizados em solução de hidróxido de sódio seguindo o protocolo de Arnott (1959),
modificado por Lersten (1967 apud Kraus e Arduin, 1997) por 24 horas. A coloração do material
diafanizado foi feita com uma solução de safranina 0,5% em etanol 30% (Kraus e Arduin, 1997).
Os fragmentos corados foram montadas em lâminas histológicas com Permount (Kraus e Arduin,
1997) e analisados em microscópio (modelo Olympus CBR, Olympus Japan) acoplado a câmara
clara (modelo LB 1,25x, Olympus) para o estudo da densidade estomática. O material foi
documentado em fotomicroscópio modelo Olympus AX70TRF, Olympus Optical, com sistema UPhoto.
2.4 Análise Micromorfólogica - Microscopia Eletrônica de Varredura
As amostras foram desidratadas em série etílica e submetidas ao ponto crítico com gás
carbônico no “Critical Pointer Dryer” (modelo CPD 030, Bal-Tec, Balzers, Liechtenstein). O
4
material seco foi afixado em suportes metálicos (“stubs”), com fita adesiva dupla face e metalizado
com pó de ouro, no metalizador modelo FDU010, Bal-Tec, Balzers.
A documentação fotográfica foi feita no Microscópio Eletrônico de Varredura (marca LEO,
modelo VP1430) acoplado a câmera digital, do Núcleo de Microscopia e Microanálise da UFV.
3. RESULTADOS
A epiderme da folha de soja completamente expandida, em vista frontal, apresenta as células
fundamentais com paredes anticlinais sinuosas (Fig. 1). Em secção transversal, a epiderme é
uniestratificada (Fig. 5) revestida por uma camada de cera epicuticular em formato de grânulos (Fig.
1).
1
2
Figura 1 e 2: Figura 1 - Glycine max (L.) Merril - Epiderme da folha vista em Microscopia
Eletrônica de Varredura. Figura 2 - Glycine max (L.) Merril - Detalhe da epiderme da face
abaxial da folha após Diafanização.
A folha é anfiestomática (Fig. 5), os estômatos são do tipo paracítico (Fig. 2) e ocorrem em
maior número na epiderme abaxial.
Os tricomas tectores (Fig. 3) e glandulares (Fig. 4 e 5) ocorrem nas duas faces da folha,
preferencialmente, sobre as nervuras. O tricoma tector, levemente elevado pelas células epidérmicas
3
fundamentais (Fig. 3), é formado por uma célula basal globosa, uma célula mediana e uma célula
apical alongada, todas revestidas por uma cutícula ornamentada (Fig. 1)
Figura 3 e 4 : Figura 2 - Glycine max (L.) Merril - Tricoma tector da epiderme abaxial de folha
vista em Microscopia Eletrônica de Varredura. Figura 3 - Glycine max (L.) Merril - Tricoma
glandular da face abaxial da folha vista em Microscopia Eletrônica de Varredura, detalhando a
célula apical (CA), o pedúnculo (Pe) e a célula basal (CB).
O mesofilo é dorsiventral, com duas camadas de parênquima clorofiliano paliçádico (PCP),
voltado para a face adaxial, uma camada de parênquima paravenal (PP) aparentemente descontínuo
e duas camadas de parênquima clorofiliano lacunoso (PCL), voltadas para a face abaxial da folha
5
(Fig. 5). Os feixes vasculares (FV) são colaterais, com o xilema voltado para a face adaxial e o
floema voltado para a face abaxial.
Os feixes vasculares são envolvidos por uma bainha parenquimática, a endoderme (Ed), e os
feixes maiores apresentam extensões de bainha (EB) parenquimática que os une a epiderme (Fig. 5
e 6). Algumas células das extensões de bainha são idioblastos contendo cristais prismáticos de
oxalato de cálcio (Fig. 6 - Cr). Na nervura mediana ocorre uma pequena quantidade de colênquima
e de fibras sobre o floema.
As plantas de Glycine max analisadas, não apresentaram grandes alterações anatômicas,
quando submetidas à elevadas concentrações de CO2.
5
6
Figuras 5 e 6. Glycine max (L.) Merril – Fig 5. Aspecto geral do limbo em secção transversal.
Glycine max (L.) Merril; Fig. 6. Detalhe de um feixe vascular evidenciando a endoderme (Ed) e
a célula da extensão de bainha (EB) contendo um cristal prismático de oxalato de cálcio (Cr).
CB - célula basal; CM - célula mediana; Et – estômato; PCL - parênquima clorofiliano
lacunoso, PCP - parênquima clorofiliano paliçádico; PP - parênquima paravenal, TG - tricoma
glandular.
A densidade estomática não variou, nas duas faces da folha, nas plantas cultivadas com o
dobro da concentração de CO2 atmosférica (≈ 720 µmol mol-1), quando comparadas com as plantas
cultivadas na concentração de CO2 atmosférica encontrada no ambiente (≈ 360 µmol mol-1).
Não foi observada diferença na espessura total do limbo das plantas submetidas à alta
concentração de CO2. No entanto, verificou-se que a espessura total do limbo difere entre as duas
linhagens analisadas (Tabela 1).
Tabela 1: Prova de médias da espessura do limbo.
Linhagem
1
2
Nota: médias seguidas de mesma letra maiúscula nas linhas não
pelo teste de Tukey a 5% de probabilidade.
Limbo
150,9 A
139,7 B
diferem estatisticamente entre si
A análise individual dos tecidos foliares mostrou que da epiderme da face adaxial e da face
abaxial não variou em relação ao tratamento com CO2. Entretanto, uma linhagem apresentou
diferença na espessura do limbo em relação à outra linhagem (Tabela 2).
Tabela 2: Prova de médias da espessura das faces adaxial e abaxial.
Linhagem
Epiderme adaxial
Epiderme abaxial
1
13,5 A
12,5 B
2
12,1 A
11,6 B
Nota: médias seguidas de mesma letra maiúscula na linha não diferem estatisticamente entre si pelo
teste de Tukey a 5% de probabilidade.
6
O parênquima clorofiliano paliçádico, parênquima clorofiliano lacunoso e o parênquima
paravenal não apresentaram alterações quanto à espessura, em relação aos diferentes tratamentos
com CO2 nem entre duas as linhagens avaliadas.
Não foram observadas alterações nos tricomas e nem nas a espessura células epidérmicas
fundamentais das folhas dos indivíduos submetidos aos dois tratamentos. No entanto, alguns
estômatos das plantas cultivadas em ambiente com o dobro da concentração de CO2 apresentavam
as células subsidiárias visivelmente maiores (Fig. 7).
Figura 4: Glycine max (L.) Merril - Epiderme da folha de plantas desenvolvidas em ambiente
7 vista em Microscopia Eletrônica
com o dobro da concentração de CO2 encontrado no ambiente,
de Varredura.
Apesar de não avaliado estatisticamente, foi observada uma maior quantidade de cera
epicuticular nas folhas das plantas submetidas ao dobro da concentração de CO2.
4. DISCUSSÃO
A presença de folhas anfiestomáticas, estômatos paracíticos e mesofilo dorsiventral
observadas nas linhagens estudadas estão de acordo com o descrito na literatura (Azevedo, 1995;
Mussury et al., 2007).
Os estudos sobre as alterações da anatomia foliar de plantas cultivadas expostas à elevada
concentração de CO2 podem ajudar a esclarecer os efeitos potenciais do aumento do CO2 global
sobre a produtividade das plantas. As plantas respondem ao aumento de CO2 atmosférico adaptando
sua absorção de CO2 e perda de água. Essas adaptações são conseguidas com mudanças na abertura
estomática e/ou densidade estomática (Miyazawa et al., 2006).
Apesar das plantas analisadas, neste trabalho, cultivadas em ambiente com o dobro da
concentração de CO2 atmosférico não terem apresentado alteração na densidade estomática,
Woodward e Kelly (1995) registraram um significativo aumento na densidade estomática de
indivíduos de Glycine max crescendo em ambiente enriquecido com um CO2 a 910 ppm, o que pode
ser a causa de um resultado diferente do presente experimento.
É amplamente conhecido que as altas concentrações de CO2 atmosférico têm efeitos sobre a
densidade estomática. Vários trabalhos têm relatado o comportamento de diferentes espécies, como:
a não alteração da densidade estomática em diferentes variedades laranjeiras (Estiarte et al., 1994);
ou aumento da densidade estomática em algumas espécies herbáceas (Ferris; Taylor, 1994; Larcher,
2000); ou ainda, redução da densidade estomática (Woodward, 1987; Beerling; Woodward, 1995).
Segundo Fernández e colaboradores (1998), diferenças nas respostas de densidade
estomática em plantas submetidas a concentrações elevadas de CO2 podem ser devidas não apenas a
variação nas concentrações de CO2, mas também a outros fatores, como por exemplo: velocidade do
vento (Retuerto; Woodward, 1993); densidade do fluxo luminoso (Tichá, 1982) e temperatura
(Beerling; Chaloner, 1993). Trabalhando com folhas de Populus trichocarpa x P. deltóides,
Miyazawa e colaboradores (2006) demonstraram que os resultados conflitantes encontrados na
7
literatura para esta espécie, com um aumento ou redução da densidade estomática, em resposta a
diferentes concentrações de CO2, poderiam estar relacionados a diferenças da intensidade luminosa
e/ou umidade durante o crescimento das espécies usadas nestes experimentos. Demonstraram ainda,
que o desenvolvimento dos estômatos e das demais células epidérmicas é afetado por todas estas
variáveis.
A presença de células subsidiárias hipertrofiadas, em alguns dos estômatos das folhas das
plantas cultivadas sob alta concentração de CO2, é marcante (Fig. 21-23), mas nada foi encontrado
na literatura que pudesse esclarecer seu aparecimento. No entanto, estes estômatos aparentemente
anômalos podem ser resultados da endogamia que resultaram nas linhagens estudadas. Azevedo
(1995) relata a ocorrência de estômatos com células subsidiárias colapsadas, próximos a áreas
necrosadas, em folhas de soja submetidas à chuva ácida, e Alves (2001), relata que a superfície
abaxial de folhas expostas a poluentes aéreos, em São Paulo, apresentou estômatos menores que as
folhas de plantas cultivadas afastadas dos poluentes.
Nas demais células da epiderme não
foram observadas alterações visíveis, a não ser um aparente aumento na quantidade de cera
epicuticular, nas folhas das plantas cultivadas sob alta concentração de CO2. O mesmo foi
observado por Thomas e Harvey (1983) em indivíduos de Glycine max (L.) cultivados com 910
ppm.
A espessura total do limbo das plantas analisadas, submetidas ao dobro da concentração de
CO2 atmosférico (≈ 720 µmol mol-1) não apresentou diferença significativa. Folhas de soja
submetidas ao enriquecimento de CO2, com concentrações de 910 ppm, por Thomas e Harvey
(1983), apresentaram um aumento na espessura total do limbo, em decorrência do aparecimento de
uma terceira camada de parênquima clorofiliano paliçádico, enquanto o parênquima lacunoso não
apresentou aumento significativo. Segundo estes autores a potencial contribuição desta terceira
camada de parênquima clorofiliano paliçádico na capacidade fotossintética requer maiores estudos.
No entanto, vale à pena ressaltar que, apesar de não ter havido acréscimo do parênquima
clorofiliano paliçádico nas folhas estudadas, Lobo (2003) verificou incremento de 46 % na taxa
fotossintética nas plantas cultivadas sob ≈ 720 µmol mol-1 de CO2.
A diferença entre os resultados encontrados neste trabalho e a literatura poderia ser
explicada pela a atuação simultânea de diferentes fatores no desenvolvimento da folha, de modo
semelhante ao que ocorreu com as diferenças de densidade estomática observadas em Populus
trichocarpa x P. deltoides, mostrado por Miyazawa e colaboradores (2006). No entanto, para
esclarecer que fatores estariam ou não atuando no desenvolvimento foliar seriam necessários novos
experimentos que controlassem todas as variáveis que afetam o desenvolvimento da folha.
As conseqüências que o aumento da concentração de CO2 provoca em cada espécie vegetal
são muito diferentes (Hunt et al., 1991 apud Murray, 1995). Os estudos sobre os efeitos nas
comunidades vegetais devem ser aprofundados, e associados ao desenvolvimento de novas
metodologias (Murray, 1995).
5. AGRADECIMENTOS
Agradecimentos sinceros à Universidade Federal de Uberlândia, em especial, à Professora
Doutora Neuza Maria de Castro. À Universidade Federal de Viçosa pelo acesso ao Laboratório de
Anatomia Vegetal e à Professora Doutora Luzimar Campos Silva pelos ensinamentos. E ao
Professor Doutor Francisco de Almeida Lobo pelo material e todo apoio dado à pesquisa.
6. REFERÊNCIAS
Aguiar, T.V.; Sant’ Anna-santos, B.F.; Azevedo, A.A.; Ferreira, R.S. Anati Quanti, 2007. Software
de análises quantitativas para estudos em anatomia vegetal. Planta Daninha, Viçosa-MG, v.25, n.4,
p. 649-659.
8
Alves, E. S.; Giusti, P.M.; Domingos, M.; Saldiva, P.H.N.; Guimaraes, E.T.; Lobo, D.J. A., 2001.
Estudo anatômico foliar do clone híbrido 4430 de Tradescantia: alterações decorrentes da poluição
aérea urbana. Revista Brasileira de Botânica, São Paulo, v. 24, n.4, p. 567-576.
Azevedo, A. A. Ação do Flúor, em Chuva Simulada, Sobre a Estrutura Foliar de Glycine max (L.)
Merril. 1995. Dissertação (Doutorado em Ciências), Universidade de São Paulo, São Paulo.
Bazzaz, F.A., 1990. The responses of natural ecosystems to the rising global CO2 levels. Annual
Rev Ecol Syst, v. 21, p. 167-196.
Beerling, D.J.; Chaloner, W.G., 1993. The impact of atmospheric CO2 and temperature change on
stomatal density: observations from Quercus robur lammas leaves. Annals of Botany, n.71, p. 231235.
Beerling, D.J.; Woodward, F.I., 1995. Stomatal responses of variegated leaves to CO2 enrichment.
Annals of Botany, n. 75, p. 507-511.
Bowes, G., 1993. Facing the inevitable: plants and increasing atmospheric CO2Ann. Rev Plant
Physiol Plant Molecular Biology, v. 44, p. 309-332.
Bulbovas, P.; Souza, R. S.; Moraes, R. M.; Luizão, F.; Artaxo, P., 2007. Plântulas de soja “Tracajá”
expostas ao ozônio sob condições controladas. Pesquisa Agropecuária Brasileira, Brasília, v.42, n.5,
p. 641-646.
Canizares, K. A. L.; Rodrigues, J.D.; Goto, R., 2004. Crescimento e índices de troca gasosa em
plantas de pepino irrigadas com água enriquecida com CO2. Horticultura Brasileira, Brasília, v.22,
n.4, p. 706-711.
Cararo, D.C.; Duarte, S.N.,2002. Injeção de CO2 e lâminas de irrigação em tomateiro sob estufa.
Horticultura Brasileira, Brasília, v. 20, n. 3, p. 432-437.
Dickison, W. C., 2000. Ecological Anatomy. In: Integrative Plant Anatomy. San Diego: Academic
Press, cap.8, p. 295-331.
Domingos, M.; Klumpp, A.; Klump, P.G., 1998. Air pollutants impact on the Atlantic forest in the
Cubatão region, São Paulo, Brazil Ciência e Cultura, São Paulo, v.50, p. 230-236.
Embrapa Soja. Soja em números (Safra 2006/2007). Londrina, 2008. Disponível em: <
http://www.cnpso.embrapa.br/index.php?op_page=294&cod_pai=516>. Acesso 2008.em: 03 jun.
Estiarte, M.; Pennuelas, J.;Kimball, B.A.; Idso, S. B.; LaMorte, R.L.; Pinter, P.J.; Wall, G.W.; Jr;
Garcia, R.L., 1994. Elevated CO2 effects on stomatal density of wheat and sour orange trees.
Journal of Experimental Botany, v.45, p. 1665-1668.
Fernandez, M.D.; Pieters, A.; Donoso, C.; TezarA, W.; Azkue, M.; Herrera, C.; Rengifo, E.;
Herrera, A., 1998. Effects of a natural source of very high CO2 concentration on the leaf gas
exchange, xylem water potential and stomatal characteristics of plants of Spatiphylum cannifolium
and Bauhinia multinervia. New Phytologit, v. 138, p. 689-697.
Ferris, R.; Taylor, G., 1994. Stomatal characteristics of four native herbs following exposure to
elevated CO2. Annals of Botany, v. 73, p. 477-453.
Furlan, R.A.; Rezende, F.C.; Alves S, D.R.B.; Folegatti, M.V., 2002. Lâmina de irrigação e
aplicação de CO2 na produção de pimentão cv. Mayata, em ambiente protegido. Horticultura
Brasileira, Brasília, v.20, n.4, p. 547- 550.
Heagle, A. S.; Body, D. E.; Heck, W. W., 1973. An open-top field chambers to assess the impact of
air pollution on plants. Journal of Environmental Quality. V. 42. p. 141-153.
Heagle, A.S.; Burns, J.C.; Fisiier, D.S.; Miller, J.E., 2002. Effects of carbon dioxide enrichment on
leaf chemistry and tmosférica by Twospotted Spider Mites (Acari: Tetranychidae) on White
Clover, Entomological Society of America, v.31, n.4, p. 594-601.
Johansen, D. A. Plant, 1940. Microtechnique. New York, McGraw-Hill Book Co. Inc., p.523
Kraus, J. E.; Arduin, M. Manual básico de métodos em morfologia vegetal. Seropédica: Edur, 1997.
198 p.
Larcher, W. A., 2000. Planta sob Estresse. In: Ecofisiologia Vegetal. São Carlos: Rima, cap. 6, p.
470-475
Lin, J.; Jach, M.E.; 2001. Ceulemans, R. Stomatal density and needle anatomy of Scots pine (Pinus
sylvestris) are affected by elevated CO2. New Phytologit, v.150, p. 665-674.
9
Lobo, F. A., 2003. Alterações na assimilação e metabolismo do carbono em plantas de soja sob
condições de atmosfera enriquecida com CO2. 2003. Tese (Doutorado) – Fisiologia Vegetal,
Universidade Federal de Viçosa,Viçosa.
Mandle, R. H.; Weinstein, L. H.; McCune, D. C.; Keveny, M., 1973. A cylindrical open-top
chamber for the exposure of plants to air pollutants in the field. Journal of Environmental Quality.
V. 2. p. 371-376.
Martin, T.N.; Dutra, L.M.C.; Jauer, A.; Storck, L.; Zabot, L.; Uhry, D.; Santi, A.L.; Stefanelo, C.;
Filho, O.A.L., 2005. Tamanho ótimo de parcela e numero de repetições em soja (Glycine max (L.)
Merril). Ciência Rural, Santa Maria, v.35, n. 2, p. 271-276.
Mazzoni-Viveiros, S. C., 2000. Diversificação do uso da anatomia: poluição atmosférica na Mata
Atlântica. In: Tópicos atuais em Botânica. 1 ed. Brasília: Embrapa Recursos Genéticos e
Biotecnologia, v. 1, p. 101-106.
Miller, A.; Tsai, C.; HemphilL, D.; Endres, M.; Rodermel, S.; Spalding, M., 1997. Elevated CO2
effects during leaf ontogeny. Plant Physiology, v. 115, p. 1195-1200.
Miyazawa, S.; Livingston, N. J.; Turpin, D. H., 2006. Stomatal development in new leaves is
related to the stomatal conductance of mature leaves in poplar (Populus trichocarpa x P. tmosfér).
Journal of Experimental Botany, v.57, n.2, p. 373-380.
Morrisey, W. e Justus, J.H., 1999. Global climate change. CRS Issue Brief for Congress. 5p.
Mott, K.A., 1990. Sensing of atmospheric CO2 by plants. Plant, Cell & Environment, v.13, n.7, p.
731-737.
Murray, D.R., 1995.Plant responses to carbon dioxide. American Journal of Botany, v. 82, n. 5, p.
690-697.
Mussury, R.M.; BetonI, R.; Silva, A.M.; Decian, V.; Scalon, S.P.Q.; Barbosa,E.L. Morfoanatomia
de Glycine max (L.) Merril infectadas por Phakopsora pachyrhizi H. Sydow & Sydow. Revista
Brasileira de Biociências, Porto Alegre, v. 5, p. s459-s461, 2007. Suplemento 1.
Olivo, M. N. 2000, 2000. Crescimento e respostas fisiológicas de duas espécies de batata (Solanum
sp) sob ambientes enriquecidos com CO2. Dissertação (Mestrado em Fisiologia Vegetal),
Universidade Federal de Viçosa,Viçosa, MG.
Pandey, R.; Chacko, P.M.; Choudhary, M.L.; Prasad, K.V.; Pal, M., 2007. Higher than optimum
temperature under CO2 enrichment influences stomata anatomical characters in rose (Rosa hybrida).
Scientia Horticulturae, n.113, p. 74-81.
Pinto, J. M.; Botrel, T. A.; Machado, E. C. Uso de dióxido de carbono na agricultura. Santa Maria.
Ciência Rural, v.30, n.4, p.919-925, 2000.
Pinto, J.M.; Botrel, T.A.; Machado, E.C.; Feitosa Filho, J.C., 2001. Aplicação de CO2 via água de
irrigação em relação à produtividade do meloeiro. Scientia Agricola, v.58, n.1, p.33-38.
Retuerto, R.; Woodward, F.I., 1993. The influences of increased CO2 and water supply on growth,
biomass allocation and water use efficiency of Sinapis alba L. grown under different wind speeds.
Oecologia, n.94, p. 415-427.
Sanz, M. J.; Penas, G. S.; Calatayud, V.; Gallego, M. T.; Cervero, J., 2002. La contaminacion
tmosférica de los bosques: guia para la identification de daños visibles causados por ozono.
Ministério Del médio Ambiente. Madrid.
Sasaki, H.; Hara, T.; Ito, S.; Uehara, N.; Kim, H.; Lieffering, M.; Okada, M.; Kobayashi, K., 2006.
Effect of free-air CO2 enrichment on the storage of carbohydrate fixed at different stages in rice
(Oryza sativa L.). Field Crops Research.
Tichá, I., 1982. Photosynthetic characteristics during ontogenesis of leaves. 7. Stomatal density and
sizes. Photosynthetica, n. 16, p. 375-471.
Thomas, J.F.; Harvey, C.N., 1983. Leaf anatomy of four species grown under continuous CO2
enrichment. Botanical Gazette, n.144, v.3, p. 303-309.
Woodward, F. I., 198.Stomatal numbers are sensitive to increases in CO2 from pre-industrial levels.
Nature, n. 327, p. 617-618.
Woodward, F. I.; Kelly, C.K., 1995.The influence of CO2 concentration on stomatal density. New
Phytologist, v. 131, p. 311-327.
10
USE OF MORPHOLOGICAL PARAMETERS IN THE ANALYSES OF
SOYBEN PLANTS (GLYCINE MAX (L.) MERRIL) IN HIGH CO2
CONCENTRATION
Elisa Rodrigues Gontijo
Universidade Federal de Uberlândia
[email protected]
Neuza Maria de Castro
Universidade Federal de Uberlândia
[email protected]
Luzimar Campos Silva
Universidade Federal de Viçosa
[email protected]
Abstract: The concentration of atmospheric CO2 is increasing and causing changes in plant
physiology and plant anatomy. Studies of how plants respond to these changes are of vital
importance. The objective of this study was to evaluate possible anatomical alterations in leaves of
Glycine max (L.) Merrill (soybean) when exposed to high concentrations of CO2 in an attempt to
contribute information on the effects caused by increased concentration of CO2. Two lines of G.
max grown under the current atmospheric CO2 concentration (≈ 360 μmol mol-1) and under
double the current CO2 concentration (≈ 720 μmol mol-1) were analyzed. Samples obtained from
fully expanded leaves were subjected to routine anatomical procedures. Epidermal peels and
scanning electron microscopy were used to observe possible changes in leaf surfaces. The stomata
density, the thicknesses of epidermal in both leaf surfaces, the thicknesses of palisade and spongy
mesophyll layers and paravenal parenchyma showed no changes compared to the high
concentration of CO2. However, were found stomata with subsidiary cells "swollen" in leaves
developed under high concentration of CO2. The inbreeding of strains analyzed and control
variables that affect the development of the plant may be related to the results.
Keywords: leaf anatomy, stomata density, anomalous stomata.
11