Fisiologia I
Data: 17 de Dezembro de 2007 (18ª aula)
Docente: Prof. A. Escalda
Desgravado por: David N. Moreira, Joana S. Jorge e M.ª Carolina Ferreira
Tema: Músculo Cardíaco
Já todos ouviram falar do músculo cardíaco: “o coração é uma bomba” que
aspira o sangue, apresentando um músculo estriado com características especiais…
- É chamado estriado pois apresenta estrias, tem também um
sarcómero (com actina e miosina), e é diferente do músculo-esquelético (ou
estriado): as aurículas formam um um sincício (syncytium), independente do
sincício dos ventrículos (a contracção das aurículas dá-se independentemente
da contracção dos ventrículos).
- Na função do coração, onde os ventrículos se contraem depois das
aurículas, tem que haver uma coordenação muscular. Esta coordenação é feita
por um tecido especial, que comanda o batimento cardíaco, primeiramente a
contracção das aurículas e posteriormente a dos ventrículos, o tecido nodal. O
nódulo sino-auricular (que está ao nível da aurícula direita) despolariza e leva o
impulso a toda a massa da aurícula (poeticamente comparado com o atirar uma
pedra a um lago, originando ondas concêntricas, que se vão espalhando e
despolarizando toda a aurícula). O estímulo chega então ao outro nódulo
aurículo-ventricular, situado no pavimento da aurícula direita, na transição para
o ventrículo. É neste nódulo que se dá um atraso, chamado atraso fisiológico,
sendo este muito importante para a função do coração (a contracção das
aurículas tem que ser diferenciada em tempo da contracção dos ventrículos,
para que o sangue saia do coração e não passe da aurícula para o ventrículo,
seguido do ventrículo para a aurícula).
1/11
- O atraso fisiológico na propagação do impulso é então feito pelo
nódulo aurículo-ventrícular, havendo a distribuição do impulso pela rede de
Purkinje, e consequente contracção dos ventrículos.
A despolarização percorreu então todo o coração, que vai voltar a
repolarizar (voltar a um estado calmo), para que se dê uma nova
despolarização.
Uma característica muito importante do coração é a sua capacidade de
responder a estímulos nervosos, capacidade essa que nos vai condicionar o diaa-dia. Temos um ritmo quando estamos sossegados, cerca de 60
batimentos/min, havendo um determinado volume sistólico (volume de sangue
em cada sístole que é lançado na aorta no lado esquerdo ou na artéria
pulmonar no lado direito), que vai aumentar quando fazemos exercício, quando
nos excitamos por qualquer motivo ou nos zangamos com alguém, através das
catecolaminas produzidas).
O sistema nervoso autónomo (simpático e parasimpático) influência o
funcionamento do coração.
Propriedades do coração:
Automatismo – capacidade de gerar estímulos para poder gerar
contracção (presente no músculo liso e coração)
Dromotropismo (ou condutibilidade) – o estímulo vai ser conduzido por
um tecido especial de condução ou até inclusivamente conduzido pela própria
massa muscular.
Batmotropismo (ou excitabilidade) – tecido pode ser excitável
Inotropismo (ou contractilidade) – permite uma maior ou menor força
por parte do coração, consoante a quantidade de sangue presente no coração.
Tonicidade – (presente em todos os tipos de músculo)
2/11
Apresenta também
bastantes canais proteícos – Gap
junctions – que levam a que a
despolarização passe mais
rapidamente de célula para
célula (condutibilidade mais
rápida), permitindo uma mais
rápida contracção cardíaca.
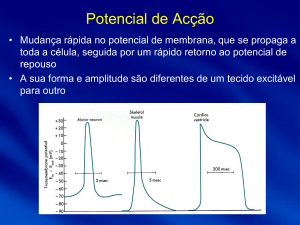
A despolarização tem
diferentes potenciais de acção: o potencial de acção ao nível do nódulo sinusal
é diferente do potencial de acção ao nível do nódulo auriculoventricular, que é
por sua vez diferente do das fibras de purkinje.
São bases iónicas que estão na base dos
diferentes potenciais de acção (há vários canais
que abrem e fecham, tanto para o sódio, como
para o potássio, cloro, cálcio, deixando passar os
diferentes iões, levando às diferentes
configurações nos potenciais de acção).
No gráfico:
Massa ventricular (no ventrículo): todos
eles partem dos cerca de 90 mV. No ventrículo, o
plateau (ou planalto) é maior do que ao nível da
aurícula (que pode até nem existir nesta). O potencial de repouso é negativo,
até à entrada de Na+, que faz com que o potencial suba, passando o valor zero
→ Há uma subida de potencial, uma despolarização. Posteriormente, a descida
do potencial de acção é devida à entrada de Cl- para a célula. O plateau que se
3/11
segue (planalto) é característico da entrada de mais Na+ e, principalmente, de
Ca2+. O K+ sai, voltando o potencial de repouso (como ocorre na repolarização
de qualquer célula).
Nódulo sino-auricular: Aqui, o potencial de repouso não está
´´sossegado`` (há uma ligeira subida dos potenciais de repouso, característica
dos potenciais dos pace-makers, nos nódulos sino-auricular, sinusal e auriculoventricular). Há no início uma descida ligeira, devido à saída de K+.
Posteriormente ocorre uma subida do potencial de acção (entrada lenta de Na +
e de Ca2+), seguida de uma entrada rápida de Ca2+ e saída de K+ por último.
São então
necessárias bases
iónicas: Na+, K+, Cl-,
mas acima de tudo
de Ca2+, muito
importante para a
despolarização da
fibra do miocárdio.
Este, como no músculo-esquelético, encontra-se ao nível do retículo
sarcoplasmático (que se encontra bastante desenvolvido).
A sua entrada é feita livremente através das trocas feitas com o Na+ na
bomba Na+/Ca2+/K+.
A sua excreção dá-se para a parte externa da membrana plasmática,
através de uma bomba ATPase dependente.
O Ca2+ existente, abundantemente, no retículo sarcoplasmático, vai-se
unir à troponina C, fazendo deslizar a tropomiosina, levando à interacção da
actina e da miosina, dando-se a contracção muscular (tal como no músculo
esquelético).
4/11
Ao nível do retículo sarcoplasmático, bem como do líquido extracelular,
também existe a calcecrestina (ou calcecristina?) (proteína que capta o Ca2+,
retendo-o no retículo sarcoplasmático). No entanto, quando ele sai do retículo
sarcoplasmático e vai ter com a troponina C, o Ca2+ fica disperso no líquido
extracelular. O fosfolambão, por sua vez, capta o Ca2+, trazendo-o de volta para
o retículo sarcoplasmático (processo que dispende energia).
Como o coração contrai e relaxa muitas vezes por minuto, é necessário
que haja uma eficaz libertação e também uma captação do Ca2+ (esta pelo
fosfolambão) para dentro do retículo, para que não ocorram falências
cardíacas.
Resumo:
Ca2+ + troponina C → contracção muscular
No relaxamento muscular → Ca2+ trazido de volta ao
retículo sarcoplasmático pelo fosfolambão (proteína)
Nota: há outros factores que fazem variar estes fenómenos.
Neste gráfico
podem ser comparadas as
tensões dos músculos
esquelético e cardíaco
(praticamente iguais). No
entanto, como a distensão do sarcómero no músculo-esquelético é bastante
maior, a distensibilidade e contracção deste músculo é maior (à volta do
pericárdio, músculo cardíaco, existe também uma membrana que impede que
o coração se distenda muito).
5/11
Ciclo cardíaco (ou Revolução cardíaca)
Consiste numa diástole, numa sístole e novamente uma diástole, sendo que os
fenómenos que se passam do lado direito do coração são precisamente os
mesmos que se passam do lado esquerdo.
Ao enchimento ventricular segue-se a contracção isovolumétrica e um
esvaziamento e relaxamento ventricular e posteriormente volta a acontecer o
enchimento ventricular, existindo válvulas que comandam estes fenómenos: do
lado esquerdo a válvula
auriculo-ventricular ou
mitral e a aórtica
(válvula semilunar
correspondente e do
lado direito a válvula
tricúspida, que é a
válvula auriculoventricular e a válvula
pulmonar ou direita.
6/11
Este gráfico permite perceber como se dá o funcionamento do ciclo cardíaco:
No ponto A a válvula mitral abre pelo que o ventrículo ainda não está cheio de
sangue mas vai aumentando de volume devido ao enchimento. Quando o
ventrículo está cheio a válvula mitral fecha (ponto C) e para que o sangue saia
do ventrículo é necessária uma determinada pressão, havendo uma fase de
aumento de tensão, uma contracção dentro do ventrículo (neste caso, o
esquerdo) e abre-se a válvula aórtica (ponto D), o que permite a saída do
sangue. Ao sair há uma injecção rápida e posteriormente mais lenta do sangue
que leva a uma diminuição gradual da pressão dentro do ventrículo e fecho da
válvula aórtica (ponto F). Quando esta fecha há um relaxamento isovolumétrico
que leva a que o sangue flua para dentro do ventrículo novamente, repetindose o ciclo.
O Sistema Nervoso
Simpático e
Parassimpático vão
alterar os potenciais
de repouso da célula.
Neste gráfico é
possível observar um
potencial de repouso
padrão.
Mas, se houver uma
estimulação simpática
ocorre um aumento da
contractilidade e da frequência cardíaca devido a um maior aumento do
potencial de repouso (entrada mais rápida de cálcio) ocorrendo a
despolarização.
Se houver uma estimulação parassimpática, o potencial de repouso é mais
baixo e demora mais tempo até atingir o potencial de despolarização, o sódio e
o cálcio entram mais lentamente e depois despolariza.
7/11
Comparando as curvas controle e a da estimulação simpática, no segundo a
inclinação é mais abrupta, diminuindo o tempo para a despolarização mas,
tanto a estimulação simpática como a parassimpática e a normal não
aumentam a amplitude do potencial de acção. A grande alteração dá-se no
potencial de repouso, isto é, o potencial do simpático é mais rápido enquanto o
parassimpático tem um potencial mais negativo e uma inclinação mais
alongada no tempo
até atingir o potencial
de repouso. Percebese assim que o
simpático aumente a
frequência e o
parassimpático a
baixe.
O volume sistólico
também vai ter
alterações. Ao fazer
uma estimulação simpática há um aumento do volume sistólico.
Pode dizer-se que o Sistema Nervoso Simpático aumenta a frequência cardíaca
e o volume sistólico e o Sistema Nervoso Parassimpático diminui a frequência e
o volume sistólico.
A força desenvolvida durante
a contracção do músculo
(inotropismo) é também
aumentada pela estimulação
simpática, tendo uma força
de contracção inotrópica
positiva, isto é, aumenta-a.
8/11
Por sua vez, o Parassimpático diminui a força de contracção, o que se traduz
num efeito inotrópico negativo.
Mas o aumento da força de contracção não se pode dar constantemente, tem
limites, não podendo aumentar constantemente porque o coração para
aguentar essa força vai ter que distender durante o relaxamento. Além disso, a
membrana do pericárdio limita a distensibilidade do coração.
Este facto é
explicado pela Lei
de Frank-Starling,
em que a
interacção da
actina e da
miosina leva a que
estas não fiquem
em contacto:
Em a) a actina e a miosina estão bastante integradas uma na outra e pode-se
desenvolver uma pequena força de contracção.
Se o músculo for distendido vai haver uma maior interacção entre a actina e a
miosina até determinado ponto porque quando se distende muito, estas
deixam de interagir e não existe contracção muscular.
Se se aumentar a força ao nível do relaxamento, a
distensão cardíaca vai aumentar também a força de
contracção até um determinado nível.
Fran-Starling idealizou que se se aumentar o aporte de
sangue ao coração, atribui-se-lhe uma pré-carga (além da
carga habitual que varia entre os 70 ou 80ml por cada batimento, podendo ir
até aos 120-130mL por batimento. Não há uma expulsão completa do sangue:
ficam sempre 40-50 mL no ventrículo) levando a que tenha que existir uma
9/11
distensibilidade maior. À medida que se vai distendendo o coração, este vai-se
contrair ainda mais. Todavia, quando se ultrapassa o nível de interacção entre a
actina e a miosina deixa de haver contracção e o coração entra em falência.
Esquematicamente, pode dizer-se que o coração tem uma determinada carga
que faz distender o elemento elástico e, consequentemente, há um aumento
de tensão ao nível do elemento contráctil, levando a que este se possa contrair.
Assim, há um movimento da carga: existe contracção.
Se se aumentar muito a carga até determinado nível há contracção e a partir
desse nível deixa de se contrair.
A pré-carga é o enchimento ventricular e a pós-carga é a dificuldade na saída de
sangue dos ventrículos. Estes dois factores condicionam o débito cardíaco, a
frequência cardíaca e, por conseguinte, o inotropismo; No fundo, condicionam
a função cardíaca em si.
O débito cardíaco (quantidade
de volume de sangue expulso
por unidade de tempo,
normalmente em minutos)
depende do volume da
frequência cardíaca e do
volume ou débito sistólico:
Débito cardíaco = Frequência
cardíaca x Volume sistólico
Este débito influencia a pressão arterial que, por sua vez, aumenta com a
resistência periférica:
Débito cardíaco = pressão arterial x resistência
Por sua vez, a Pré-carga condiciona o encurtamento ou da fibra miocárdia. Este
aumento ou encurtamento da fibra do miocárdio vai determinar também o
aumento do débito sistólico.
10/11
A pressão arterial muito elevada aumenta o tamanho do ventrículo esquerdo e
se o ventrículo está maior há mais sangue lá dentro e aumenta o débito
sistólico. Trata-se de um ciclo quase vicioso.
Se se aumentar a contractilidade a força inotrópica positiva do coração
também vai aumentar o encurtamento da fibra do miocárdio que conduz ao
débito cardíaco e ao aumento da pressão arterial.
Se ocorrer um aumento da pressão arterial, há pós-carga que condiciona o
encurtamento da fibra do miocárdio, ou seja, a pré-carga aumenta o
encurtamento da fibra do miocárdio e a pós-carga diminui o encurtamento
dessa mesma fibra.
Esta tabela é um resumo de quase tudo o que foi falado de músculos e é
importante. O sítio de regulação do cálcio no músculo liso é a calmodulina que
por sua vez se une ao caldesmon.
11/11