OBTENÇÃO DE MATÉRIA PELOS SERES HETEROTRÓFICOS
Unicelularidade e pluricelularidade
Absorção: Dá se antes do processamento e utilização (constituição das estruturas das células, fonte
de energia, participação em reações vitais, permição da formação de novas células e renovação
celular) de nutrientes básicos (água, minerais, vitaminas, proteínas, glícidos, lípidos). É a passagem
de substâncias do meio externo para o meio interno.
Constituição da membrana plasmática: As membrana são complexos lipoproteicos, constituídos
(relativamente ao peso) por 60% a 75% de proteínas e 25% 40% de lípidos, podendo ainda conter
até 10% de glícidos. Esta composição pode variar conforme o tipo de célula.
→ As proteínas presentes na membrana plasmática possuem composição e funções
diversas. Podem ter função estrutural, de transporte de substâncias, de receção de estímulos ou de
enzimas.
→ Os lípidos que constituem a membrana são, maioritariamente, fosfolípidos (presentes
em todas as membranas), colestrol e glicolípidos.
→ Os glícidos membranares situam-se na parte exterior da membrana plasmática. Embora
as suas funções não sejam completamente conhecidas, sabe-se que têm um papel impotante no
reconhecimento de certas substâncias or parte da célula.
Evolução dos modelos da membrana plasmática:
1885 – Nageli e Carmer: existência de membrana celular
1899 – Overton: constituição lipídica
1925 – Gorter e Grendel: teoria da bicamada fosfolipídica
1935 – Davson e Danielli: modelo em que a bicamada fosfolipídica seria revestida interna e
externamente por uma camada de proteínas, que facilmente se ligariam às extremidades polares
hidrofílicas dos fosfolípidos
1954 – Os mesmos propõem a existência de poros proteicos que formariam passagens hidrofílicas
através das quais as substâncias polares poderiam atravessar a membrana. As substãncias não
polares, por sua vez, atravessariam a membrana diretamente através da bicamada fosfolípidica
1972 – Singer e Nicholson: Modelo atual da membrana plasmática – Modelo de mosaico fluido:
→ A membrana é uma estrutura dinâmica, sendo que as moléculas fosfolipídicas podem
mudar de posição na mesma camada (movimento lateral) ou mudar de camada (movimento flipflop)
→ A membrana é constituída por: bicamada fosfolipídica, proteínas extrínsecas e
intrínsecas (algumas que atravessam a membrana de um lado ao outro – transmembranares)
colestrol, glicocálix (conjunto de glicoproteínas e glicolípidos responsáveis pelo reconhecimento de
certas substâncias por parte da célula que estão na superfície externa da membrana plasmática).
Digestão intracelular – importância do sistema endomembranar
Sistema endomembranar: membrana do invólucro nuclear, retículo endoplasmático e complexo e
Golgi – funcionam como uma unidade
Retículo endoplasmático: é um conjunto de citernas achatadas, túbulos e vesículas esféricas,
formando um sistema cvontínuo entre a membrana plasmática e o invólucro nuclear
→ Retículo Endoplasmático Rugoso: maior região de síntese de proteínas (algumas das
quais enzimas) que ocorre nos ribossomas e no transporte das mesmas
→ Retículo Endoplasmático Liso: síntese de lípidos (fosfolípidos) e, logo, elaboração de
novas membranas e transporte de algumas substâncias prejudiciais à célula
Compexo de Golgi/dictiossoma: é composto por conjuntos de sáculos ou cisternas achatadas e
empilhadas de forma regular, na periferia das quais existe uma série de vesículas. Tem uma face
convexa (face de formação), virada para o RE e uma face concâva (face de maturação), onde se
formam vesículas, virada para a membrana plasmática. A face de formação do dictiossoma recebe
materiais (proteínas) do RER através de contactos diretos, por intermédio de canalículos, ou
através da fusão de vesículas. As cisternas, que se vão constituindo na face de formação, vão
substituindo as que se situam na face de maturação, que, por sua vez, dão origem a vesículas de
secreção – armazenando as proteínas produzidas no RER e transportando-as para os locais onde
são necessárias dentro de vesículas. As substâncias provenientes do RE sofrem algumas
transformações durante a sua passagem pelo complexo de Golgi – o que vai permitir que algumas
proteínas se tornem funcionais e algumas enzimas se tornem ativas. O complexo de Golgi está
também envolvido em processos de secreção e de síntese de diversas substâncias, tais como
glicoproteínas e polissacarídeos.
Lisossomas: são pequenas vesículas, mais ou menos esféricas, delimitadas por uma membrana e
que contêm vários tipos de enzimas. Formam-se na fase de maturação do dictiossoma e podem
unir-se a uma vesícula endocítica, formando um corpo de maiores dimensões chamado vacúolo
digestivo. É nesta estrutura que ocorre a digestão das substâncias captadas por endocitose, graças
às enzimas hidrolíticas provenientes do lisossoma – heterofagia. Quando é necessário, os
lisossomas também participam na digestão dos próprios organelos celulares, formando vacúolos
autofágicos – autofagia. Quando o processo de autofagia se descontrola e faz autolise.
Obtenção de matéria pelos seres heterotróficos multicelulares
Na maioria dos seres heterotróficos multicelulares, o processamento dos alimentos até que estes
possam fornecer os seus constituintes para que sejam utilizados pelas células engloba:
→ Ingestão: entrada dos alimentos para o organismoi
→ Digestão: conjunto de processos que permite a transformação de moléculas complexas
dos alimentos em moléculas mais simples
→ Absorção: passagem dos nutrientes resultantes da digestão para o meio interno
Digestão extracelular: ocorre em cavidades que, apesar de se encontrarem dentro do organismo,
fazem parte do meio externo, pois não são mais do que prolongamentos desse meio para o interior
do corpo. Nessas cavidades, são lançados sucos digestivos que contém enzimas, que atuam sobre
os alimentos, transformando-os em substâncias mais simples, capazes de serem absorvidas. Esta
representa uma vantagem evolutiva para os organismos, dado que podem ingerir quantidades
significativas de alimento em cada refeição, que é assim armazenado e vai sendo lentamente
digerido. Desta forma, o organismo não precisa de estar continuamente a captar alimento.
Tubo digestivo incompleto: só possui uma abertura. ex.: hidra e planária
Tubo digestivo completo: duas aberturas: a boca, para a entrada dos alimentos, e o ânus, para a
saída dos resíduos. Esta modificação constitui um importante avanço evolutivo, pois permite uma
digestão e uma absorção sequenciais no tubo digestivo, uma vez que os alimentos se deslocam
apenas num sentido. A digestão pode ocorrer em vários orgãos, por processos mecânicos e ações
enzimáticas distintas. Aqui, após a digestão os nutrientes vão ser absorvidos pelo intestino (que
geralemente tem uma grande superfície de absorção) e vão ser transportados para todas as células
do organismo através do sistema sanguíneo e linfático. Os resíduos dos alimentos (não absorvidos)
continuam o seu trajeto pelo intestino e são expulsos pelo ânus.
OBTENÇÃO DE MATÉRIA PELOS SERES AUTOTRÓFICOS
Fotossíntese
É um processo realizado pelas cianobactérias, pelas algas e plantas. É utilizada a energia luminosa
para produzir compostos orgânicos a partir de dióxido de carbono e água – convertem energia
luminosa em energia química – 6CO2 + 12H2O → C6H12O6 + 6O2 + 6H2O
As clorofilas a e b (pigmento de cor verde) e os carotenoides (pigmento de cor alaranjada) são
essenciais para a captação da energia luminosa – há uma maior produção de oxigénio nas zonas
iluminadas pelos comprimentos de onda absorvidos por estes pigmentos. A absorção da energia
luminosa pelos diferentes pigmentos está relacionada com a configuração eletrónica dos átomos
que os constituem. Quando os pigmentso fotossintéticos absorvem a luz, os seus eletrões passam
para níveis de energia superiores. Quando estes eletrões excitados são cedidos a outras moléculas
ocorre uma reação de oxidação (perda do eletrão) -redução (ganho do eletrão), gerando-se um
fluzo de eletrões.
Admite-se, atualmente, que os pigmentos fotossintéticos estão dispersos na membrana dos
tilacoides.
O oxigénio libertado na fotossíntese tem origem na água e não no dióxido de carbono.
É necessária luz para dar ínicio ao processo fotossintético, mas a incorporação de dióxido de
carbono não depende diretamente da luz, pois pode-se prolongar na obscuridade.
Fase dependente da luz:
→ Quando as moléculas de clorofila são atingidas pela luz, origina-se uma corrente de
elctrões que se propaga ao longo de uma série de proteínas (acetores) que se encontram dispostas
ao longo da membrana interna do cloroplasto.
→ Este fluxo de eletrões liberta energia que é utilizada para formar ATP (a partir de
moléculas de ADP e de um grupo fosfato). Diz-se, por isso, que há transformação de energia
luminosa em energia química (que fica armazenada na ligação química que se estabeleceu para
formar o ATP). O fluxo de eletrões também permite formar moléculas de NADPH, as quais, tal
como o ATP, são fundamentais para a formação de compostos orgânicos.
→ Os eletrões perdidos pela clorofila são repostos pela molécula de água, que sofre um
fenómeno de fotólise (desdobramento da molécula de água por ação da luz), o que permite a
separação dos átomos de hidrogénio (de onde provêm os eletrões para neutralizar a clorofila e
para a redução do NADP+ a NADPH) e de oxigénio (que é libertado pela planta para o meio
ambiente). - em determinadas situações, as plantas podem só usar o fotossistema I, neste caso não
há redução do NADPH, mas somente produção de ATP, então os eletrões perdidos pela clorofila,
depois de transportados pela cadeia, reressam ao próprio fotosssistema.
Fase não dependente diretamente da luz:
→ Combinação do dióxido de carbono com uma pentose, isto é, um glícido formado por 5
átomos de carbono – ribulose difosfato (ruDP)-, originando um composto intermédio, instável com
seis carbonos.
→ Este composto origina, imediatamente, duas moléculas com 3 átomos de carbono –
ácido fosfoglicérico (PGA)
→ As moléculas de ácido fosfoglicérico são fosforiladas palo ATP e posteriormente
reduzidas pelo NADPH, provenientes da fase luminosa, formando aldeído fosfoglicérico (PGAL).
→ Por cada 12 moléculas de aldeído fosfoglicérico formadas, 10 são utilizadas para
regenerar a ribulose difosfato e duas são utilizadaspara sintetizar compostos orgânicos (glícidos e
outros).
→ Para se formar uma molécula de glicose é preciso que o ciclo se realize 6 vezes,
gastando-se 6 moléculas de dióxido de carbono, 18 moléculas de ATP (3 por cada ciclo) e 12 de
NADPH (2 por cada ciclo).
→ Produtos: glicose (ou frutose, lípidos ou prótidos), água, NADP+ e ADP
Químiossíntese
A quimiossíntese é um processo alternativo à fotossíntese em que se produzem compostos
orgânicos, utilizando como fonte de energia a oxidação de compostos minerais. Fazem
quimiossíntese as bactérias sulfurosas, ferrosas e nitrificantes do solo e algumas archaeobactérias.
Duas fases da quimiossíntese:
→ Fase de produção de ATP e NADPH: Durante esta fase, ocorre a oxidação de compostos
minerais, como o amoníaco, o dióxido de carbono ou o sulfureto de hidrogénio. Esta oxidação
permite a obtenção de protões e eletrões que são transportados ao longo de uma cadeia, no
sentido de produzir ATP e reduzir o NADP+ a NADPH.
→ Ciclo de Calvin/Ciclo do carbono/Ciclo das pentoses: Produção de compostos orgânicos a
partir do dióxido de carbono absorvido, do poder redutor do NADPH e da energia contida no ATP,
gerados na primeira fase.
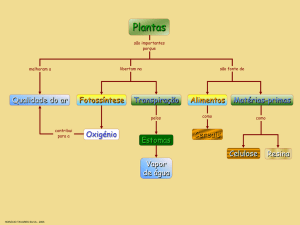
O TRANSPORTE NAS PLANTAS
As plantas, enquanto sres pluricelulares complexos, necessitam de transportar substâncias
minerais até às folhas, para grantir a síntese de compostos orgânicos que aí ocorre.
Posteriormente, esses compostos terão de ser distribuídos a todas as células, de forma a poderem
ser utilizados.
Classificação das plantas:
→ Avasculares
→ Vasculares:
− sem semente (reproduzem-se por esporos): pteridófitas
− com semente: cicas, gingko e coníferas
− com semente e com flor:
• monocotiledóneas (têm um só cotiledone na sementes, folhas estreitas e
compridas, raiz fasciculada, estomas nas páginas superior e inferior da folha,
bandas em U na raiz): arroz, centeio, cevada, etc.
• dicotiledóneas (têm dois cotiledones na semente, folhas arredondadas, raíz
aprumada, estomas só na página inferior, bandas de Caspary na raiz – espessamentos em linha):
feijão, ervilha, fava, etc.
A água e os sais minerais utilizados na síntese de matéria orgânica, entram na planta por absorção,
através da raiz. O dióxido de carbono utilizado durante a fotossíntese entra na planta através de
estomas. Os estomas também controlam a quantidade de água que se evapora pelas folhas, num
processo denominado transpiração – de dia estão abertos e permitem a transpiração, à noite
fecham.
Cerca de 99% da água absorvida pelas raízes é perdida sob a forma de vapor, através das folhas.
Porém esta água é substituída por outra, transportada, juntamente com sais minerais, num
sistema contínuo de vasos, que se estende desde a raiz, passa pelos caules e chega até às folhas –
xilema. No floema há movimento de água e solutos orgânicos, resultantes da fotossíntese, que se
deslocam, essencialmente, das folhas para os outros órgãos das plantas, pois a fotossíntese não se
verifica em todas as células da planta. O movimento de água e de solutos no interior da planta
através destes tecidos condutores denomina-se translocação.
Xilema/Tecido traqueano/Lenho (na maioria das plantas é constituído por):
→ Elementos condutores: (células mortas onde circulam a água e os sais minerais):
− tracoides: células longas e de extremidades afiladas, as quais contactam entre si,
formando tubos que permitem a passagem de água e de sais minerais
− elementos de vasos: células vasculares com um diâmetro superior ao dos
tracoides, que resultam de células mortas que perderam as paredes transversais e cujas paredes
laterais apresentam espessamentos de lenhina (substância que lhes confere rigidez)
→ Fibras lenhosas: constituídas por células mortas cujas paredes são espessas devido à
deposição de lenhina e desempanham funções de suporte.
→ Parênquima lenhoso: tecido formado por células vivas, pouco diferenciadas, que
desempanha importantes atividades metabólicas nas plantas, tais como a fotossíntese, o
armazenamento ou a secreção de substâncias.
Floema/Tecido crivoso/Líber (na maioria das plantas é constituído por):
→ Células dos tubos crivosos: células vivas muito especializadas, ligadas entre si pelos topos
e cujas paredes de contacto possuem uma série de orifícios, que se assemelham a um crivo; no
inverno, os orifícios destas placas crivosas fcam obstruídos por calose que se dissolve na
primavera; as células dos tubos crivosos são vivas, embora tenham perdido a maior parte dos
organelos.
→ Células de companhia: células vivas que se situam junto das células dos tubos crivosos,
com as quais mantêm numerosas ligações citoplasmáticas, ajudando-as, assim, no seu
funcionamento.
→ Fibras: comprimento variável, desempanham funções de suporte.
→ Parênquima: formado por células vivas pouco diferenciadas que têm funções de reserva.
A maior parte da água e dos iões necessários para as várias atividades da planta é absorvida pelo
sistema radicular.
Normalmente, o meio intracelular das células da raiz é hipertónico relativamente ao extrior, pelo
que a água tende a entrar na planta por osmose, movendo-se desde o solo até aos vasos xilémcos
existentes no interior da raiz.
Os iões minerais, quando presentes no solo em concentrações elevadas, entram nas células da raiz
por difusão simples; no entanto, é usual verificar-se a tal elelevada concentração destes iões no
meio intracelular. Neste caso, os iões entram para as células por transporte ativo.
O transporte ativo de iões através das células da periferia da raiz até ao xilema cria um gradiente
osmótico, que faz com que a água tenda a passar por osmose até ao xilema.
Transporte no xilema
A água e os iões minerais, uma vez chegados ao xilema, constituem a seiva bruta /seiva xilema,
que pode agora ser distribuída pela planta. Uma vez no xilema, as substâncias dissolvidas na água
são transportadas de forma passiva.
Hipóteses para o movimento ascendente de água no xilema:
→ Hipótese da pressão radicular: A ascenção da água do xilema dá-se graças a uma pressão
que se desenvolve ao nível da raiz, graças à ocorrência de forças osmóticas; a contínua acumulação
de iões nas células da raiz tem como consequência a entrada de água para a planta; a
acumulaçãode água nos tecidos provoca uma pressão na raiz que força a água a subir no xilema.
− argumentos a favor:
• exsudação: quando se fazem podas tardias em certas plantas, verifica-se a
saída de água pelas zonas dos cortes efetuados
• gutação: quando a pressão radicular é muito elevada, a água é forçada a
subir até às folhas, onde é libertada sob a forma líquida.
− argumentos contra:
• os valores da pressão radicular nem sempre são suficientes para explicar a
ascensão de água até ao topo das árvores.
• existem mesmo várias espécies que não apresentam pressão radicular
→ Hipótese da Tensão-Coesão-Adesão: A quantidade de vapor de água que sai das folha
por transpiração causa uma tensão na parte superior da planta que provoca a ascensão da água.
Esta tensão ocorre devido às propriedades da água circulante na planta. Então, devido à polaridade
que apresentam, as moléculas de água tendem a ligar-se umas às outras por pontes de hidrogénio,
que se estabelecem entre os átomos de hidrogénio de uma molécula e os átomos de oxigénio de
moléculas próximas. Graças a estas forças de coesão, as moléculas de água mantêm-se unidas
entre si. As moléculas de água têm ainda a capacidade de aderir a outras substâncias,
nomeadamente aos constituintes das paredes do xilema. Por último, estas forças de tensãocoesão-adesão fazem com que se estabeleça uma coluna de água no xilema, desde as raízes até às
folhas. O movimento das moléculas de água, que se perdem por transpiração ao níbel das folhas,
faz mover toda esta coluna no sentido ascendente. Consequentemente, quanto mais rápida for a
transpiração foliar, mais rápida se torhna a absorção radicular – este sistema só funciona quando
existe uma conituidade na coluna de água, isto é, quando não há a interferência de bolhas de água
ou arrefecimento intenso da água.
− argumento a favor: a tensão produzida pela transpiração é suficiente para
provocar a ascensão da água até a uma altura de 150 metros, em tubos de diâmetro de
vasos xilémicos.
Hoje em dia aceita-se a hipótese que é uma união da hipótese da pressão radicular e da tensãocoesão-adesão, embora se considere que o processo dominante é este último.
Transporte no floema:
A seiva floémica/seiva elaborada é constituída por produtos orgânicos (geralmente 10% a 20% de
sacarose, ou outros açucaçres, ácidos aminados e nucleótidos) hormonas e iões inorgânicos – o
que lhe confere uma certa viscosidade.
Hipótese do fluxo de massa:
Os glícidos produzidos nas folhas durante a fotossíntese são convertidos em sacarose antes de
entrarem para o floema, para serem transportados aos locais onde são armazenados ou gastos,
tais como as flores, os frutos, as sementes, os caules ou as raízes.
A passagem da sacarose das células das folhas para as células de companhia do floema ocorre por
transporte ativo. Seguidamente, a sacarose passa destas células para os elementos dos tubos
crivosos, através das ligações citoplasmáticas. O aumento da concentração de sacarose nas células
dos tubos crivosos provoca uma entrada de água, vinda do xilema, nestas células, que ficam
túrgidas. A pressão de turgescência obriga a solução de sacarose a deslocar-se através da placa
crivosa para a célula seguinte do tubo e assim sucessivamente.
O sentido do fluxo é determinado pelas concentrações relativas da sacarose que é produzida e
utilizada, o que gera um gradiente de concentração descrescente, desde o local de produção
(folhas) até ao local de consumo ou armazenamento.
TRANSPORTE NOS ANIMAIS
Os animais necessitam de receber nutrientes e oxigénio para as suas células e têm de eliminar
dióxido de carbono e outros produtos resultantes do metabolismo.
A forma mais eficaz de estas substâncias atravessarem a membrana celular é sob a forma
dissolvida, o que implica que as células sejam banhadas por um meio líquido.
Nos animais mais simples, não existe um sistema de transporte especializado, pois têm poucas
camadas de células e o facto de estarem em contacto direto com o meio permite que o oxigénio se
difunda, de forma direta, da água para as células. Os nutrientes difundem-se da cavidade para
gatrovascular para as células e os produtos de excreção, resultantes do metabolismo, são lançados
diretamente no meio.
Nos animais mais complexos existem orgãos especializados na absorção de nutrientes e oxigénio e
na excreção de substâncias tóxicas. A condução destas substâncias, entre os órgãos onde são
absorvidos e as células do resto do organismo, é feita através de um sistema de transporte
especializado – o sistema circulatório.
Sistemas circulatórios abertos/lacunares: A hemolinfa abandona os vasos sanguíneos e contacta
diretamente com as células. O líquido circulante é designado hemolinfa por não existir qualquer
diferença entre o sangue e a linfa. A hemolinfa é bombeada por um coração tubular, ao longo de
vasos, até aos tecidos. Ao chegar aos tecidos, a hemolinfa sai dos vasos e passa para um sistema de
cavidade/lacunas que no seu conjunto formam o hemocélio. No hemocélio, a hemolinfa entra em
contacto com as células, fornecendo-lhes nutrientes e recebendo produtos de excreção. Após a
irrigação dos tecidos, a hemolinfa entra novamente no coração tubular, através dos ostíolos
(orifícios do coração). A entrada da hemolinfa no coração ocorre devido à força de sucção que é
gerada quando este órgão relaxa. Seguidamente, os ostíolos fecham, o coração contrai e a
hemolinfa é novamente impulsionada ao longo dos vasos. Nestes sistemas, a hemolinfa tem pouca
velocidade, embora estes animais possuam uma elevada taxa metabólica – isto porque os gases
respiratórios não são transportados pelos fluídos circulantes, existindo um sistema respiratório que
conduz os gases diretamente até aos tecidos.
Sistemas circulatórios fechados: O líquido circulante designa-se sangue e, em codições normais,
nunca abandona os vasos. Devido à contração do coração, o sangue é distribuído por todo o
organismo no interior de vasos, cujo calibre vai diminuindo: artérias – arteríolas – capilares –
vénulas – veias. Os capilares têm apenas uma camada de células e formam uma rede em cada um
dos órgãos, de forma a atingirem praticamente todas as células. As trocas realizam-se entre o
sangue dos capilares e a linfa que envolve as células – linfa intersticial. O sangue fornece oxigénio e
nutrientes e recebe produtos resultantes do metabolismo celular.
Aqui, a velocidade dos fluídos circulantes é maior, pois sendo estes animais mais complexos, há
necessidade de se fazerem as trocas mais rapidamente.
Sistema circulatório fechado simples:
São os peixes que têm este tipo de circulação. É circulação simples, porque apenas o sangue
venoso atravessa o coração.
O sangue venoso, proveniente dos diferentes orgãos, entra na aurícula através de uma pequena
dilatação – o seio venoso. A contração da autícula impele o sangue para a segunda câmara, mais
musculosa – o ventrículo. A contração do ventrículo impulsiona o sangue para fora do coração,
passando por uma dilatação – o cone arterial – até atingir as brânquias. Nas brânquias efetuam-se
trocas gasosas – hematose branquial – sendo o sangue recolhido numa artéria de grande calibre –
a aorta – que se ramifica (artérias – arteríolas – capilares) conduzindo o sangue a todo o corpo.
Nos tecido dos diferentes órgãos, o sangue circula numa rede de capilares, que permitem a
libertação de oxigénio e de nutrientes e a recolha de dióxido de carbono e de outros produtos
metabólicos. A rede de capilares une-se ( capilares – vénulas – veias) e o sangue regressa ao
coração.
O sangue atinge a aorta com baixa pressão, diminuindo a eficácia de oxigenação dos restantes
tecidos.
Sistema circulatório duplo incompleto:
Os anfíbios têm este tipo de circulação. É uma circulação dupla, pois o sangue efetua dois trajetos
diferentes, passando duas vezes pelo coração. A circulação está dividida em circulação
pulmonar/pequena circulação e em circulação sistemática/grande circulação. É incompleta devido
ao facto de poder ocorrer mistura parcial de sangue venoso com sangue arterial, ao nível do
ventrículo.
Os anfíios apresentam o coração dividido em três cavidades: duas aurículas e um ventrículo.
O sangue, proveniente dos diferentes órgãos, dá entrada na aurícula direita, enquanto que na
aurícula esquerda entra sangeu proveniente dos pulmões. A contração das aurículas conduz o
sangue para o ventrículo, que, por sua vez, impulsiona uma parte do sangue para os pulmões e
outra para os restantes órgãos.
Como as duas aurículas encaminham o sangue para um único ventrículo, o sangue venoso e o
arterial poderiam misturar-se, o que íriaempobrecer a oxigenação dos tecidos. No entanto, devido
à estrura anatómica do ventrículo, o sangue venoso, proveniente da aurícula direita, tende a ser
enviado para o circuito pulmonar, equanto que o sangue arterial, proveniente da aurícula
esquerda, é conduzido à aorta para realizar o circuito sistémico.
A pequena circulação corresponde ao trajeto que o sangue efetua desde o ventrículo, através da
artéria pulmonar, até aos pulmões, onde é oxigenado, transformando-se em sangue arterial,
voltando à aurícula esquerda, pelas veias pulmonares.
A grande circulação corresponde ao trajeto do sangue desde o ventrículo até aos diferentes
órgãos. Nos tecidos desses órgãos, o sangue passa de arterial a venoso, regressando ao coração
por veias que terminam no seio venoso, o qual dá entrada na aurícula direita.
A grande vantagem deste duplo circuito é permitir que o sangue circule com uma pressão mais
elevada, logo, há um fluxo mais vigoroso do sangue.
Sistema circulatório duplo e completo:
É um sistema duplo por ter pequena e grande circulação e completo por existirem dois ventrículos
e o sangue venoso e arterial não se misturarem.
O sangue venoso entra a aurícula direita e o arterial na esquerda. A entrada do sangue dá-e na
diástole auricular (auriculas relaxadas). A sua contração (sístole auricular) conduz o sangue aos
respetivos ventrículos. A contração do ventrículo direito impulsiona o sangue para os pulmões
através da artéria pulmonar que se ramifica até aos capilares alveolares, onde se realiza a
hematose pulmonar. Ao mesmo tempo, dá-se a sístole ventricular do ventrículo esquerdo, que faz
o sangue atravessar a aorta ( curvatura para a direita nas aves e esquerda nos mamíferos), que se
ramifica ( artérias – arteríolas – capilares) e leva o sangue aos principais órgãos. Aqui o sangue
torna-se venoso e volta para o coração pelas veias cavas.
O facto de não haver mistura de sangue permite uma elevada eficáia de oxigenação dos tecidos e,
por isso, uma maior capacidade energética. Parte da energia é utilizada por estes animais para a
manutenção de uma temperatura constante (homeotermia) – animais mais evoluidos – mais
adaptados.
Fluídos circulantes
Os fluidos circulantes existem como veículos de transporte e servem para responder às
necessidades das células e para protegerem o organismo.
Isto porque as células, para sobreviverem, precisam constantemente de nutrientes, oxigénio,
hormonas, de removerem os produtos de desassimilação, e proteção; e o nosso corpo precisa de
ter uma temperatura homogénea. Assim, nos vertebrados, existem dois tipos de fluidos
circulantes: o sangue e a linfa.
O sangue é formado por uma parte líquida – o plasma – e por elementos figurados – as hemácias
(ou glóbulos vermelhos), os leucócitos (ou glóbulos brancos) e as plaquetas. É um líquido
vermelho, dada a existência de uma grande quantidade de glóbulos vermelhos.
O plasma transporta os elementos figurados do sangue, nutrientes, dióxido de carbono (resultante
da respiração celular), produtos de excreção, hormonas (que controlam as atividades celulares),
anticorpos e células do sistema imunitário.
As hemácias são células sem núcleo, que transportam oxigénio e algum dióxido de carbono,
também, mas em menor quantidade que o plasma.
Os leucócitos servem apenas para a defesa do organismo, já que ao fazerem fagocitose, destroem
os corpos estranhos e, logo, tentam acabar com as possíveis consequências dos mesmos, como
doenças, infeções, etc., e também ajudam na coagulação do sangue, a fim de parar hemorragias.
As plaquetas são também células sem núcleo, cuja principal função é a coagulação do sangue.
Para existir a circulação do sangue tem de haver um órgão propulsor – o coração – e vasos
condutores: as artérias, as veias e os capilares. Quanto maior for a capilarização, maior é a área de
contacto, logo, maior será a irrigação e a distribuição.
A linfa é constituída por leucócitos e plasma (que transporta o mesmo que o sangue, exceto
hemácias e plaquetas). É um líquido amarelado e, visto que transporta essencialmente leucócitos,
é muito importante na defesa do organismo.
No sistema linfático não existe órgão musculoso que faça movimentar a linfa. A linfa movimentase devido à existência de válvulas, semelhantes às das grandes veias sanguíneas, que asseguram o
seu fluxo numa única direção. O movimentos lento da linfa é devido à pressão osmótica do sangue
e à pressão mecânica e ocasional provocada pelos músculos esqueléticos.
A linfa circula nos capilares linfáticos, vasos linfáticos e gânglios linfáticos. Os vasos linfáticos de
maior calibre reúnem-se no canal torácico e canal linfático ou grande veia linfática.
A linfa divide-se em dois tipos: a intersticial e a circulante. A intersticial é formada pelo plasma e
leucócitos que, ao atravessarem as paredes dos capilares, vão ocupar os espaços entre as células
e, assim, esta vai ter um contacto maior com as células. Quando a linfa intersticial se encontra em
excesso, é recolhida por um conjunto de capilares estreitamente ligados aos capilares sanguíneos
– capilares linfáticos e, torna-se, então, linfa circulante.
Os materiais estranhos ao organismo, como bactérias e partículas virais, transportados pelos vasos
linfáticos até aos gânglios, vão ser, aqui, eliminados pelos leucócitos. Assim se explica que, quando
há uma infeção, há um inchaço nessa zona, visto que os gânglios, nessa altura, sofrem um
aumento.
OBTENÇÃO DE ENERGIA
Todas as células necessitam de energia para a realização das suas atividades. Assim, os compostos
orgânicos (considerados depósitos energéticos) são degradados de forma a libertarem energia,
formando ATP. Uma parte desses compostos orgânicos é utilizada pelas células dos seres
autotróficos que os produzem, sendo outra parte usada pelos heterotróficos (fluxo energético que
tem início no sol e continua através dos seres vivos).
Trabalho
mecânico, biossínteses,
calor, transporte ativo,
movimentos celulares
Metabolismo celular: conjunto de reações que as células realizam que são essenciais à vida.
→ Catabolismo: reações em que os compostos orgânicos são degradados em moléculas
mais simples, ocorrendo libertação de energia.
→ Anabolismo: reações em que ocorre formação de moléculas mais complexas a partir
de moléculas mias simples, ocorrendo consumo de energia.
A energia libertada nos processos catabólicos realiza-se por etapas, pois se fosse libertada de uma
vez, provocaria um elevado aumento de temperatura, o que podera colocar em risco a vida.
Assim, as células obtêm energia através de uma série de reações em cadeia, nas quais as
substâncias orgânicas vão sendo lentamente degradadas, libertando-se a energia em etapas. Esta
energia não pode ser utilizada diretamente nas atividades celulares, sendo, por isso, acumulada
em compostos intermédios como o ATP (transportador universal de energia, a nível celular).
Existem diversas vias catabólicas capazes de transferir a energia contida nos compostos orgânicos
para moléculas de ATP. Nestas vias, intervêm compostos (como o NAD) que transportam os
protões e eletrões do hidrogénio, desde o substrato até a um acetor final:
Respiração: Via catabólica que visa a obtenção de energia em que o acetor final de eletrões é uma
molécula inorgânica.
→ Respiração aeróbia: quando o acetor é o oxigénio (maioria dos animais e plantas).
→ Respiração anaeróbia: quando o acetor final é, por exemplo, o nitrato ou o sulfato
(algumas bactérias).
Fermentação: Processo catabólico que visa a obtenção de energia em que não existe um acetor de
eletrões externo, ou seja, no final do processo degrativo, os eletrões são captados por uma
molécula orgânica que deriva do subtracto inicial (bactérias e outros seres vivos na ausência de
oxigénio – anaeróbios facultativos).
Fermentação
É um processo simples e, em termos evolutivos, primitivo; ocorre no hialoplasma das células e tem
duas etapas:
→ Glicólise
→ Redução do piruvato
Respiração aeróbia:
À medida que as células evoluíram, as suas necessidaes energéticas aumentaram. Nas células eu
carióticas, surgiram organelos especializados (mitocôndrias) capazes de realizar a oxidação
completa do ácido pirívico obtido na glicólise, originando compostos muito simples (água e
dióxido de carbono).
4 etapas da respiração aeróbia:
→ Glicólise
→ Formação de acetil-coenzima A: na presença de oxigénio, o ácido pirúvico entra na
mitocôndria, onde é descarboxilado (perde uma molécula de dióxido de carbono) e oxidado
(perde um hidrogénio que é usado para reduzir o NAD+ , formando NADH + H+).
→ Ciclo de Krebs/Ciclo do ácido cítrico: conjunto de reações metabólicas que conduz à
oxidação completa da glicose, é um processo catalizado por várias enzimas (descarboxilases e
desidrogenases). São precisos dois ciclos para a degradação de cada molécula de glicose.
Cadeia transportadora de eletrões/Cadeia respiratória e fosforilação oxidativa: a cadeia
transportadora de eletrões encontra-se ordenada na membrana interna das mitocôndrias, de
acordo com a sua afinidade para eletrões. Os eletrões transportados pelo NADH e pelo FADH 2 são
cedidos aos acetores, iniciando um fluxo, ao longo do qual estas moléculas vão sendo
sucessivamente reduzidas e oxidadas. Este fluxo é condicionado pela disposição das moléculas
transportadoras de eletrões. Cada transportador tem mais afinidade para os eletrões do que o
transportador anterior, garantindo-se desta forma um fluxo unidirecional até ao oxigénio. O
oxigénio, depois de receber os eletrões, capta os protões presentes na matriz da mitocôndria,
formando água. À medida que os eletrões passam de transportador em transportador, liberta-se
energia. Essa energia é utilizada para fosforilar o ADP, formando ATP – fosforilação oxidativa.
Balanço nergético de ATP's formados:
Total de ATP necessário para
desencadear o processo
Fermentação
Respiração aeróbia
2
2
TROCAS GASOSAS EM SERES MULTICELULARES
Todos os seres vivos necessitam necessitam de oxigénio e compostos de carbono nas suas células,
de removerem o dióxido de carbono e de energia.
Trocas gasosas nas plantas
Durante a respiração, a planta capta oxigénio e liberta dióxido de carbono; durante a fotossíntese
capta dióxido de carbono e liberta oxigenio; durante a transpiração, a planta liberta vapor de água
que tinha captado pela raíz assim como os sais minerais.
Os estomas controlam as trocas gasosas entre a planta e o meio externo, graças à capacidade de
que têm de abrir e fechar.
As células das folhas e os respetivos espaços intercelulares possuem um elevado conteúdo em
água, devido ao contínuo fluxo de água desde a raiz até à folha, o que leva à saída de vapor de
água da folha para o exterior, por transpiração.
O movimento de abertura e fecho dos estomas é condicionado por alterações na turgescência das
células estomáticas/guarda, que têm uma estrutura diferente das células vizinhas, já que possuem
muitos cloroplastos e a parte da parede celular que delimita o ostíolo (pore) é mais espessa que a
parte encostada às células vizinhas. A existência de microfibrilas de celulose provoca o
afastamento das células estomáticas (quando túrgidas), aumentando a abertura dos ostíolos.
Quando uma célula vegetal está túrgida, aumenta de volume e, consequentemente, exerce
pressão sobre a parede – pressão de turgescência. Nas células de guarda, a região mais fina da
parede deforma-se mais facilmente que a região espessa, levando à abertura do estoma. Quando
as células de quarda perdem água, a pressão de turgescência diminui, permitindo às células
aproximarem-se, fechando o estoma. Esta variação na turgescência pode ser provocada pela
concentração de certos iões nas células, concentração de dióxido de carbono, a luz, a
temperatura, o vento ou o conteúdo de água no solo:
→ O mecanismo mais aceite para explicar a variação de turgescência é o transporte
ativo do ião potássio: a elevada concentração de potássio no interior das células estomáticas faz
com que entre água por osmose para as mesmas; as células guardas ficam túrgidas e abre-se o
estoma; vai haver transporte ativo do ião potássio para dentro destas células – equanto isto
ocorrer o estoma permanece aberto; o ião potássio para de entrar nas células guarda e começa a
sair por difusão, tal como a água – o estoma fecha.
→ A diminuição de concentração de dióxido de carbono nos espaços intracelulares das
células das folhas causa a abertura dos estomas. Isto porque durante a fotossíntese consome-se
dióxido de carbono e formam-se solutos no interior das células estomáticas, o que provoca a
entrada de água nas mesmas, logo a abertura dos estomas.
→ Como a luz determima a ocorrência de fotossíntese, também vai determinar a
abertura dos estomas.
Trocas gasosas nos animais
As trocas gasosas nos animais exigem a existência de superfícies respiratórias. Aqui, as trocas
gasosas podem ocorrer por difusão direta (entra as células e o meio exterior) ou difusão indireta
(quando os gases são transportados por um fluido circulante) – hematose.
Características comuns das superfícies respiratórias:
→ Pouca espessura, geralmente apenas uma camada de células entre o meio externo e interno;
→ Apresentam-se sempre húmidas, o que facilita a difusão dos gases;
→ São muito vascularizadas, para facilitar o contacto com o fluido circulante;
→ A sua morfologia permite uma grande superfície de contacto entre o meio externo e interno.
Nos animais menos complexos (hidra) a camada de células exterior realiza trocas com o meio
aquático envolvente, equanto qua a camada de células interior realiza essas trocas com a água
que circula na cavidade gastrovascular.
Outros animais, um pouco mais evoluídos, têm tegumento que permite que, mesmo vivendo num
ambiente terrestre, possam realizar trocas diretamente do meio envolvente (hematose cutânea),
por terem uma pele húmida e um sistema circulatório muito próximo da pele.
Mesmo no caso da aves e dos répteis ovíparos, os embriões desenvolvem-se graças à trocas
gasosas efetuadas através dos milhares de poros existentes na superfície dos ovos.
A maioria dos animais aquáticos faz respiração através de brânquias/guelras. Os peixes ósseos têm
as brâquias protegidas na câmara branquial, protegidas por uma estrutura óssea móvel –
opérculo. Nestes peixes, as brânquias são banhadas por uma corrente contínua de água, que entra
pela boca e sai pelas fendas operculares. O movimento de abertura e fecho da boca e dos
opérculos ajuda a esta circulação. As brânquias são constituídas por séries de filamentos duplos,
inseridos obliquamente em estruturas ósseas denominadas arcos branquiais. Em cada filamento
branquial existe um vaso sanguíneo por onde o sangue entra na brânquia e um por onde o sangue
sai. Entre estes dois vasos sanguíneos existe uma densa rede de capilares, que estão contidos em
dilatações do filamento branquial – lamelas. A água que entra pela boca do peixe passa por entre
as lamelas, cruzando-se em sentido contrário com o sangue, que circula nos capilares sanguíneos –
mecanismo contracorrente.
Mecanismo contracorrente: O sangue flui no sentido contrário ao da água, o que permite
aumentar significativamente a eficiência da hematose branquial. À medida que o sangue flui
através dos capilares vai ficando cada vez mais rico em oxigénio e, uma vez que circula no sentido
contrário da água, vai contactando com a água que é sucessivamente mais rica em oxigénio.
Assim, o coeficiente de difusão é elevado, que permite que cerca de 80% do oxigénio de difunda
para o sangue e o dióxido de carbono para a água.
Trocas gasosas através de traqueias
As superfícies respiratórias invaginadas dentro do corpo reduzem significativamente as perdas de
água por evaporação. Os inscectos e outros antrópedes terrestres possuem um sistema de
respiração constituído por uma rede de traqueias, que se encontra no interior do corpo. Estas
traqueias vão-se ramificando em tubos cada vez mais finos, que terminam nas traquíolas, que
contactam diretamente com as células. As traqueias dos insetos contactam com o exterior através
de pequenos orifícios localizados na superfície do corpo – espiráculos.
Nos insetos mais primitivos, os espiráculos encontram-se permanentemente abertos, não
havendo controlo do ar que circula através deles. Já nas espécies mais evoluídas, os espiráculos
possuem filtros, bem como estruturas semelhantes a válvulas (ostíolos) que controlam o fluxo de
ar. Nos menores insetos não ocorre ventilação ativa, mas, nos maiores, tal ocorre por movimentos
musculares que contraem as traqueias.
No sistema traqueal, as traqueias servem de condutas de ar e mantêm-se abertas devido à
existência, na sua parede, de uma estrutura dequitinizada enrolada em forma de hélice. As trocas
gasosas ocorrem por difusão direta do epitélio das traquíolas para as células. O sistema
circulatório não é, por isso, utilizado para o transporte dos gases respiratórios.
Trocas gasosas através de pulmões
Todos os vertebrados terrestres possuem pulmões.
Os anfíbios possuem pulmões mais simples e efetuam igualmente trocas gasosas através da pele.
Os répteis, mais adaptados à vida terrestre que os anfíbios, possuem pulmões um pouco mais
desenvolvidos. As aves e os mamíferos possuem aparelhos respiratórios muito complexos.
As aves são animais com um metabolismo muito elevado, pelo que necessitam de elevadas
quantidades de oxigénio. Para tal, apresentam uma grande superfície respiratória e uma eficiente
ventilação pulmonar. Para além dos pulmões, as aves possuem sacos aéreos, localizados por todo
o corpo, que constituem reservas de ar, melhorando assim a eficacia da ventilação. Para além de
ajudarem as aves a tornarem-se menos densas, os sacos aéreos, contribuem para a dissipação e
calor resultante do metabolismo, sobretudo durante o voo. No sistema respiratório das aves, o ar
circula apenas num sentido, num circuito que passa pelos sacos aéreos posteriores, pulmões e
sacos aéreos anteriores. A hematose ocorre apenas nos pulmões, nos parabrônquios, que são
finos canais, abertos nas duas extremidades. Os sacos aéreos é que permitem que o ar circule
apenas num sentido. Para que o ar precorra todo o sistema respiratório de uma ave, são
necessários dois ciclos ventilatórios. Na primeira inspiração, o ar circula pela traqueia até aos
sacos posteriores. Durante a expiração, esse ar passa para os pulmões. Durante a segunda
inspiração, passa ar novo para os sacos posteriores e o ar dos pulmões passa aos sacos anteriores.
Quando o ar contido nos sacos posteriores passa para os pulmões, o ar dos sacos aéreos
anteriores é expulso para o exterior. Aqui, o ar também circula nos parabrônquios no sentido
oposto ao da circulação sanguínea.
No caso dos mamíferos, a superfície respiratória +e constituída por milhões de alvéolos
pulmonares, dispostos em caho à volta dos bronquíolos. Neste tipo de pulmões, o ar circula em
dois sentidos opostos – inspiração e expiração.
No caso do Homem, mesmo depois de uma respiração profunda, permanece sempre algum ar nos
pulmões – ar residual.
REGULAÇÃO NERVOSA E HORMONAL EM ANIMAIS
Todos os sistemas biológicos são abertos e todos os organismos interagem como o seu meio
ambiente, assim, todos recebem estímulos. Ao receberem estímulos, podem alterar o seu
metabolismo e fisiologia. A evolução para a multicelularidade, e particularmente os elevados
níveis de organização dos animais, exigiu o desenvolvimento de complexos mecanismos de
comunicação entre as células e os órgãos.
Homeostasia:mecanismo que os seres vivos possuem que equilibram as alterações induzidas pelo
meio externo, para que exista uma constância no meio interno. Quando a homeostasia é rompida,
o ser vivo fica doente, só quando este é reposta, pelos mecanismos homeostáticos, é que o ser
vivo volta ao seu estado normal.
No sentido de evitar a perda de homeostasia, a atividade dos órgãos é controlada e regulada,
respondendo ás alterações, quer do meio interno como externo, através de mecanismos de
retroalimentação ou feedback. O sistema de feedback positivo alimenta a resposta, o feedback
negativo reverte o sistema para um determinado ponto – é utilizado por sistemas reguladores que
pretedem manter a estabilidade.
Nos animais os mecanismos de feedback são assegurados pelo sistema nervoso (para respostas
rápidas que envolvem a propagação de alterações eletroquímicas ao longo das membranas
celulares) e pelo sistema hormonal (respostas mais lentas que envolvem a libertação de
mensageiros químicos para os fluídos circulantes, até atingirem os órgãos-alvos – orgãoes
recetores capazes de detetar a presença destes mensageiros químicos).
O sistema hormonal/endócrino não é independente do sistema nervoso, os dois interagem no
sentido de regularem os mecanismos homeostáticos. De facto, grande parte da atividade
endócrina é controlada, direta ou indiretamente, pelo hipotálamo. Esta região do cerébro
estabelece a região entre o sistema nervoso e o hormona, quer do ponto de vista anatómico, quer
do fisiológico. O hipotálamo recebe diversos estímulos, quer de outras partes do cérebro, quer do
hormonas que o atingem pela corrente sanguínea. Em resposta a estes estímulos, os neurónios
hipotalámicos produzem neuro-hormonas que atingem a hipófise. A hipófise é uma glândula
proxima do hipotálamo que tem uma natureza endócrina (lóbulo anterior) e nervosa (lóbulo
posterior). Algumas hormonas produzidas pelo hipotálamo são encaminhadas até ao lóbulo
posterior da hipófise, sendo aí lançadas para a corrente sanguínea, atingindo as células-alvo.
Porém, outras hormonas hipotalámicas atuam sobre o lóbulo anterior da hipófise, estimulando ou
inibindo a produção e libertação de hormonas hipofisárias que vão controlar o funcionamento de
diversos tecidos do organismo, incluindo outras glândulas endócrinas. O sistema nervoso e o
hormonal relacionam-se estruturalmente.- muitas glândulas endócrinas são células nervosas;
encontram-se quimicamente relacionados - as hormonas podem ser usadas como sinais entre o
sistema nervoso e o hormonal; relacionam-se funcionalmente, uma vez que a coordenação do
organismo envolve os dois sistemas e cada um é afetado pelo outro.É pela interação e
coordenação dos sistemas nervoso e hormonal que o indivíduo vai assegurando as respostas
adequadas às diversas solicitações que interna e externamente desafiam os padrões
homeostáticos da vida.
O sistema nervoso regula a temperatura corporal através de mecanismos de retroalimentação
negativa.
A unidade do sistema nervoso é a célula nervosa – o neurónio, apesar de seream as células de glia
a sustentarem-nos.
O conjunto do axónio e da bainha de mielina (que apenas se encontra nos vertebrados e
invertebrados mais complexos) formam a fibra nervosa. As fibras nervosas reunidas em feixes
envolvidos por uma capa de tecido conjuntivo constituem os nervos.
.
Os neurónios são células altamente estimuláveis, capazes de detetar pequenas alterações do
meio. Em resposta a estas variações, verifica-se uma alteração elétrica, que percorre a sua
membrana – impulso nervoso. A passagem do impulso nervoso de um neurónio para outro faz-se
através das sinapses.
As sinapses podem ser químicas e elétricas. Nas sinapses químicas, quando o impulso nervoso
atinge as extremidades do axónio, libertam-se, para a fenda sináptica, substâncias químicas
designadas neurotransmissores.
O sistema nervoso regula a temperatura corporal através de mecanismos de retroalimentação
negativa.
→ Animais homeotérmicos: animais que têm a capacidadede regular a sua temperatura para um
nível constante.
− Animais endotérmicos: animais que regulam a sua temperatura corporal produzindo calor por pr
→ Animais poiquilotérmicos: animais que cuja temperatura varia com as alterações da
temperatura do meio.
− Animais ectotérmicos: animais cuja taxa metabólica não contraria as alterações de
temperatura e, portanto, dependem largamente das fontes de calor externas.
Um fator limitante é qualquer fator ambiental que condiciona a vida dos seres vivos. Isto é, a vida
só é possível dentro de determinados valores desse facto, pois em casos extremos os seres vivos
não são capazes de regular a sua atividade de forma a compensarem a ação desse fator).
Na termorregulação estão envolvidos fenómenos de vasodilatação, vasoconstrição e trocas de
calor.
Osmorregulação: processo que permite a manutenção do equilíbrio de água e sais no organismo.
→ Animais osmoconformantes: animais que não regulam a concentração de sais dos seus fluidos
corporais, e, portanto,.esta concentração varia de acordo com a concentração do meio.
→ Animais osmorreguladores: animais que regulam a concentração de sais dos seus fluidos
corporais.
A hormona antidiurética (ADH) está envolvida no processo de osmorregulação de alguns animais.
A hormona antidiurética, ou vasopressina, promove a acumulação de líquidos por parte dos rins e
contribui para reter a quantidade adequada de água. Quando um doente está desidratado, existem
recetores especiais no coração, pulmões, cérebro e aorta que indicam à hipófise a necessidade de
produzir mais quantidade de hormona antidiurética. As concentrações no sangue dos eletrólitos,
tais como o sódio, o cloro e o potássio, devem ser mantidas dentro da uma margem estreita para
que as células possam funcionar normalmente. As concentrações elevadas destes elementos, os
quais são percebidos pelo cérebro, estimulam a libertação desta hormona. A dor, o nervosismo, o
exercício físico, as baixas concentrações de açúcar no sangue, a angiotensina, as prostaglandinas e
certos medicamentos, como a clorpropamida, os medicamentos colinérgicos e outros que se
empregam para tratar a asma e o enfisema, também estimulam a libertação da hormona
antidiurética.
Hormonas vegetais
Tropismos: movimentos das plantas que envolvem crescimento na direção de um estímulo
ambiental, ou na direcçção oposta a esse estímulo. Podem ser negativos ou positivos conforme o
movimento se realiza em direção ao estímulo ou se afasta dele.
Quanto à natureza do estímulo que os origina, os tropismos podem ser: fototropismo,
gravitropismo, tigmotropismo, termotropismo, quimiotropismo e hidrotropismo.
Nastias: movimentos que não envolvem crescimento direcionado relativamente ao estímulo.
Podem ser fotonastias, tigmonastias, termonastias, quimionastias ou hidronastias.
O principal fator interno de regulação das reações de desenvolvimento e crescimento nas plantas
são hormonas vegetais/fitohormonas (auxinas, giberelinas, citoquininas, ácido abscístico e
etileno).
A regulação íntrinseca efetuada pelas hormonas vegetais está dependente de fatores extrínsecos,
como a luz ou a temperatura.
O controlo de fatores extrínsecos como a luz (manipulação do fotoperíodo) ou a temperatura (uso
de estufas) também é muito utilizado em hortofloricultura, permitindo obter flores, frutos e
legumes fora da época habitual do seu desenvolvimento.
Meristemas: células que têm uma grande capacidade de divisão e diferenciação.
As hormonas vegetais raramente agem sozinhas. É a produção coordenada das diversas hormonas
que permite à planta sobreviver às mudanças das estações do ano, bem como às condições
climatéricas extremas, e continuar o seu desenvolvimento.
Respostas das plantas em relação a estímulos ambientais
Ácido
abscísico
Estimula a abscisão. Inibe a germinação de sementes e o desenvolvimento de
gomos. Promove o fecho dos estomas em plantas com stresse hídrico.
Reprodução sexuada nas plantas
Órgãos masculinos: Estames: antera (têm sacos polínicos), filete.
Órgãos femininos: Carpelo: estigma, estilete, ovário (têm os óvulos).
As flores podem ser hermafroditas ou unissexuadas.
Para que haja reprodução é necessário que ocorra polinização.
→ Polinização direta: polinização na mesma flor.
→ Polinização cruzada: entre flores da mesma população. - permite maior variabilidade.
Ciclos de vida
Ciclo de vida da espirogia:
Após a fecundação, os filamentos desagregam-se, ficando cada um dos ovos rodeado por uma
parede espess, em estado de vida latente, geralmente durante o inverno, até que as condições
ambientais voltem a ser favoráveis. Então, o núcleo de cada ovo divide-se por meiose, formando
quatro núcleos haploides – três desses degenream.
Ciclo de vida do polipódio:
Tipo de reprodução
Vantagens
Desvantagens
Sexuada
Proporciona uma grande
variabilidade de características
na descendência.
A diversidade de características
permite às espécies não só
maior capacidade de
sobrevivência, caso haja
mudanças ambientair, mas
também proporciona evolução
para novas formas.
Processo lento.
Grande dispêndio de energia,
quer na formação dos gâmetas,
quer nos processos que
desencadeiam a fecundação.