PIBIC – Programa Institucional de Bolsas de Iniciação Científica
Instituto de Botânica
Alterações fisiológicas e metabólicas em plantas de Arabidopsis thaliana
L. com baixa atividade de nitrato redutase em resposta a altas
concentrações de CO2 atmosférico
Bolsista: Bárbara Celeste Messa
Orientadora: Dra. Marília Gaspar
Colaboradores: Dra. Ione Salgado (UNICAMP)
Dr. Marcos P. M. Aidar (IBt)
Projeto Institucional: “INFLUÊNCIA DA ATMOSFERA ENRIQUECIDA EM CO2 EM
COMBINAÇÃO COM OZÔNIO, ÓXIDO NÍTRICO E OUTROS ESTRESSES
AMBIENTAIS
NAS
RESPOSTAS
FISIOLÓGICAS,
BIOQUÍMICAS
MOLECULARES DE PLANTAS CULTIVADAS E NATIVAS TROPICAIS”.
E
PIBIC – Programa Institucional de Bolsas de Iniciação Científica
Instituto de Botânica
Alterações fisiológicas e metabólicas em plantas de Arabidopsis thaliana
L. com baixa atividade de nitrato redutase em resposta a altas
concentrações de CO2 atmosférico
Bolsista: Bárbara Celeste Messa
Aluna do 7o Semestre do Curso de Ciências Biológicas do Centro Universitário São
Camilo
Data de Ingresso PIBIC: agosto/2008 (bolsista renovação)
Orientadora: Marília Gaspar
Colaboradores: Dra. Ione Salgado (UNICAMP)
Dr. Marcos P. M. Aidar (IBt)
Projeto Institucional: “INFLUÊNCIA DA ATMOSFERA ENRIQUECIDA EM CO2 EM
COMBINAÇÃO COM OZÔNIO, ÓXIDO NÍTRICO E OUTROS ESTRESSES
AMBIENTAIS
NAS
RESPOSTAS
FISIOLÓGICAS,
BIOQUÍMICAS
E
MOLECULARES DE PLANTAS CULTIVADAS E NATIVAS TROPICAIS”.
1
1. INTRODUÇÃO E OBJETIVOS
O nitrogênio é o nutriente mineral essencial mais limitante para o crescimento e
desenvolvimento das plantas (Hardy & Havelka, 1975). As plantas adquirem nitrogênio do
solo na forma de nitrato (NO3-), amônia, uréia e aminoácidos. O NO3- é a fonte preferencial de
nitrogênio absorvida do solo pelas plantas superiores (Cheng et al., 1988).
O nitrato absorvido pelas raízes pode ser reduzido ou armazenado nos vacúolos
foliares, ou transportado para a parte aérea, onde será reduzido ou armazenado; esta
propriedade de armazenamento garante uma reserva de nitrogênio que pode ser consumida em
condições de carência nutricional, além de contribuir para a manutenção do volume e da
turgescência celular (Meyer et al., 1993). O nitrato, além de fonte de nitrogênio, age como
molécula sinalizadora em diversos processos celulares (Crawford, 1995).
O primeiro passo da redução de nitrato ocorre no citoplasma e envolve a ação da
enzima nitrato redutase (NR), produzindo nitrito (NO2-), que é a seguir reduzido à amônia
pela ação da enzima nitrito redutase (NiR). A amônia produzida é fixada via complexo de
enzimas GS/GOGAT em aminoácidos, glutamina e glutamato, que servem de substrato para a
produção de todos os aminoácidos precursores das proteínas. A NR também é capaz de
produzir óxido nítrico (NO) e óxido nitroso (NO2) a partir de nitrito (Dean & Harper, 1986;
Rockel et al., 2002). A produção de NO também pode ser mediada por atividade da enzima
óxido nítrico sintase (NOS), tendo como substrato a arginina (Stuehr, 1997).
Devido à complexidade dos metabolismos de carbono e nitrogênio, tem sido difícil
elucidar completamente a relação entre os mesmos. Já se sabe que o fluxo de carbono pode
ser direcionado para a síntese de açúcares ou amido ou para a formação de açúcares
orgânicos, que estão envolvidos na formação de aminoácidos. O fornecimento de altas
concentrações de nitrato a mutantes de tabaco com baixa atividade de NR aumentou o nível
de transcritos das enzimas NR e PEP carboxilase e levou a um acúmulo de ácidos orgânicos
(Scheible et al., 1997). A conexão entre ambos os metabolismos foi investigada utilizando-se
cultivos em dias curtos, plantas com expressão reduzida de NR e adição de açúcares (Scheible
et al., 1997; Matt et al., 1998; Nielsen et al., 1998). Plantas com baixa expressão de NR
acumulam altas concentrações de nitrato e apresentam maior crescimento da parte área
quando comparadas com as plantas selvagens, sugerindo uma mudança na alocação de
carbono devido ao acúmulo de nitrato (Scheible et al., 1997).
As condições ambientais e poluentes interferem na assimilação de nitrato. Um dos
principais poluentes é o NOx, uma mistura de gases contendo principalmente NO2. O NO2
entra na planta através da cutícula ou dos estômatos e resulta em aumento da concentração de
2
nitrato e compostos nitrogenados reduzidos que, após serem transportados para a raiz via
floema, inibem a assimilação de nitrato (Tischner et al., 1988).
Magnusson e colaboradores (1996) demonstraram uma inibição da assimilação de
nitrato em algas do gênero Ulva crescidas em altas concentrações de CO2. O aumento das
concentrações de CO2 atmosférico também afeta o metabolismo de nitrogênio em plantas
vasculares. As taxas mais elevadas de crescimento em alto CO2 resultam em maior demanda
por nutrientes minerais, que podem ser obtidos de duas formas: i) aumentando a eficiência de
uso dos nutrientes, ii) aumentando a taxa de absorção e assimilação dos minerais pelas plantas
(Stitt & Krapp, 1999). Além disso, a aceleração do crescimento e o aumento de biomassa em
alto CO2 podem alterar o status nutricional da planta.
O fenômeno de aclimatação fotossintética é geralmente mais visível em plantas com
carência de nitrogênio do que em plantas com bom “status” de nitrogênio (Sage, 1994;
Bowler & Press, 1996). Riviere-Rolland e colaboradores (1996) mostraram que altas
concentrações de CO2 diminuem a atividade de Rubisco e o nível de transcritos rbcS em
plantas e ervilha crescidas em condições limitantes de nitrogênio. Estes resultados indicam
que a repressão da expressão da Rubisco pelo acúmulo de açúcares é mais eficiente em
plantas crescidas na ausência de nitrogênio. No entanto, outra explicação possível seria que a
diminuição de Rubisco refletiria uma diminuição no teor de proteínas foliares devido a uma
re-alocação do nitrogênio para as folhas jovens em crescimento (Miller et al., 1997).
Altas concentrações de CO2 induzem um aumento de açúcares solúveis, sendo que o
teor de amido permanece inalterado em plantas de Nicotiana plumbaginifolia crescidas em
meio hidropônico com altas concentrações de nitrato. No entanto, quando as mesmas plantas
foram cultivadas em vaso sob baixas concentrações de nitrato, houve um maior aumento do
conteúdo de amido e apenas um ligeiro aumento no conteúdo de açúcares solúveis (FerrarioMery et al., 1997). Em tabaco, o elevado CO2 levou a um aumento marcante do conteúdo de
amido, mas não do conteúdo de sacarose e açúcares redutores em condições limitantes de
nitrogênio (Geiger et al., 1999).
Com relação à atividade da enzima nitrato redutase, resultados bastante contraditórios
estão disponíveis na literatura. Embora o elevado CO2 tenha induzido um pequeno aumento
na atividade NR em mostarda e Vigna radiata, uma diminuição pôde ser observada em trigo e
milho (Stitt & Krapp, 1999). Li et al. (2006) avaliaram mudanças de expressão gênica e do
perfil metabólico de três diferentes ecótipos de Arabidopsis cultivados em elevado CO2.
Estes autores observaram uma diminuição do nível de transcritos nia2 nos ecótipos Col-0 e
WS, mas não foram detectadas alterações para o ecótipo Cvi-0.
3
O aumento de CO2 atmosférico leva a mudança no padrão de alocação de biomassa e
fenologia que podem incluir aumento da razão raiz: parte aérea, maior ramificação e
florescimento precoce (Bazzaz & Fajer, 1992). Já o acúmulo de nitrogênio provoca o efeito
inverso e leva a um atraso na floração de plantas de Arabidopsis (Stitt & Krapp, 1999). Com
relação à floração, o comportamento de plantas de Arabidopsis thaliana é bastante variável na
presença de altas concentrações de CO2, sendo que um mesmo ecótipo apresenta
comportamentos contrastantes em experimentos distintos (Springer & Ward, 2007).
Arabidopsis como modelo de estudo
Arabidopsis thaliana, é uma dicotiledônea da família Brassicaceae, utilizada como
planta modelo em estudos de fisiologia, bioquímica e biologia molecular. Algumas
características importantes, como seu pequeno porte e desenvolvimento rápido (5-6 semanas
da germinação até o desenvolvimento de novas sementes), associadas ao pequeno genoma
completamente seqüenciado, tornam esta espécie o organismo ideal para estudos de
desenvolvimento vegetal. Além disso, mutações podem ser facilmente induzidas através da
irradiação de sementes ou do tratamento com reagentes químicos, sem a necessidade de
empregar plantas transgênicas.
Foram identificados dois genes codificando a enzima nitrato redutase (NR) em
Arabidopsis thaliana, nia1 e nia2 (Cheng et al., 1988; Wilkinson & Crawford, 1991). O gene
nia2 é responsável por 90% de toda a atividade de NR da planta enquanto nia1 responde pela
atividade residual (10%) (Yu et al.,1998). O duplo mutante nia1nia2, foi utilizado neste
estudo, possui apenas 0,5% de atividade de NR em comparação ao tipo selvagem (Wilkinson
& Crawford, 1993).
Algumas respostas deste duplo mutante cultivado em condições ambientais
normais já são conhecidas, como o acúmulo de nitrato (Wilkinson & Crawford, 1993), baixo
conteúdo de nitrito e arginina, maior susceptibilidade à infecção por Pseudomonas syringae
(Modolo et al., 2006), floração precoce devido à deficiência de NO na fase vegetativa
(Seligman et al., 2008), entre outros.
As respostas de Arabidopsis thaliana em alto CO2 têm sido bastante estudadas. No
entanto, embora algumas plantas transgênicas ou mutantes, deficientes em genes de
fotossíntese, metabolismo de carboidratos e síntese de hormônios, tenham sido analisadas, as
respostas do mutante nia1nia2 não são conhecidas. O estudo da associação entre os
metabolismos de carbono e nitrogênio em alto CO2 tem focado na análise do comportamento
de plantas cultivadas na presença ou ausência de diferentes fontes nitrogenadas. Mutantes
deficientes na assimilação de nitrato ainda não foram analisados neste contexto.
4
Este projeto busca caracterizar o comportamento do mutante nia1nia2 de Arabidopsis
thaliana, cultivado em altas concentrações de CO2, por meio de da análise inicial de alguns
parâmetros fisiológicos e bioquímicos. Pretende-se avaliar de que forma o cultivo destas
plantas na presença de alto CO2 afeta as respostas de fotossíntese e crescimento, os
metabolismos de carbono e nitrogênio e o processo de floração, com relação às plantas
selvagens.
Esta caracterização inicial embasará estudos futuros de análise de expressão de genes
do metabolismo de carboidratos (como INV e SS) e nitrogênio (NR, NiR, GS/GOGAT) e de
genes relacionados com a indução da floração (como FRI e FLC).
2. MATERIAL E MÉTODOS
2.1 Material vegetal
O experimento foi realizado com plantas de Arabidopsis thaliana L. ecótipo
Columbia-0 e plantas da mesma linhagem, mutantes para os genes estruturais nia1nia2 que
codificam a enzima nitrato redutase (NR), que as tornam deficientes na assimilação de
nitrogênio (Wilkinson e Crawford, 1993). As sementes foram gentilmente cedidas pela Dra.
Ione Salgado (UNICAMP) ou adquiridas do banco de germoplasma de Arabidopsis (ABRC).
As plantas foram cultivadas em câmaras de topo aberto conforme descrito em Aidar et al.
(2002), sendo que uma das câmaras foi mantida com atmosfera de 760 ppm de CO2 e outra
com concentração atmosférica normal de CO2 (380 ppm), dentro de uma casa de vegetação. A
concentração interna de CO2 nas câmaras foi monitorada diariamente com o auxílio do
medidor portátil de CO2 435 AG (Testo).
As sementes foram plantadas diretamente no substrato comercial West Garden e
vermiculita expandida média (1:1) e os vasos foram irrigados com solução nutritiva
(Wilkinson e Crawford, 1991) três vezes por semana, sendo que nos demais dias as plantas
foram irrigadas com água destilada. A solução nutritiva das plantas selvagens continha 10mM
de nitrato de potássio e a solução das plantas mutantes continha 10mM de cloreto de potássio,
para compensação osmótica.
2.2. Medidas dos parâmetros fisiológicos e bioquímicos
a) Fluorescência
As medidas da fluorescência da clorofila a foram feitas na 5ª ou 6ª folha da roseta, por
meio de um fluorômetro modulado (PAM 2000, Heinz Walz). As medidas foram realizadas
adotando-se o método do pulso de saturação em folhas pré-adaptadas ao escuro (15 minutos).
Foram dados pulsos (1s) de luz com intensidade luminosa crescente variando de 25 a 1500
5
µmol m-2 s-1. Foram estimados o ETR (taxa aparente de transporte de elétrons) e o yield
(Fv’/Fm’).
b) Determinação do conteúdo endógeno de óxido nítrico (NO)
As análises de emissão de NO foram realizadas em colaboração com a Dra. Ione
Salgado, no Departamento de Bioquímica da UNICAMP. Foram selecionadas três plantas de
cada tratamento. Foram coletados 40mg de folhas intactas de cada amostra, sendo a seguir
colocadas em tubo do tipo eppendorf contendo 30 µl de DAF-2 50 µM em DMSO e 670 µl de
tampão fosfato de potássio 0,1M (pH 7,2). O material foi incubado por 1 hora no escuro, à
temperatura ambiente. O material vegetal foi retirado da solução e a mesma foi centrifugada
para eliminar possíveis resíduos. O sobrenadante foi transferido para novos tubos eppendorf e
congelado à -80°C até o momento da realização da leitura. O branco consistiu em solução
DAF-2 em tampão fosfato, submetida às mesmas condições, sem a incubação de material
vegetal. A leitura das amostras foi realizada em espectrofluorímetro (F 4500, Hitachi, Tokyo,
Japan) com excitação a 495 nm e emissão de 500 a 550 nm.
c) Dosagem da atividade de nitrato redutase in vivo
Para as medidas de atividade de NR, foram selecionadas três plantas de cada
tratamento. Foram coletados 30 mg de material fresco e incubados em 1mL do tampão de
reação descrito por Yu et al. (1998) para Arabidopsis (40 mM KNO3, 0,08 M Na2HPO4,
0,02M NaH2PO4 e 4% de n-propanol, pH 7,5). As amostras foram submetidas a três pulsos de
vácuo de 1 minuto cada, para penetração do tampão no tecido. A seguir, as amostras foram
incubadas no escuro por 1 hora a 30ºC. Após esse período, foram retirados 400 µl de
sobrenadante e ao mesmo foram adicionados 400 µL de ácido sulfanílico a 1% e 400 µL de
naftilelilenodiaminodihidrocloreto (NED) a 0,05%. A reação foi incubada a temperatura
ambiente por 30 minutos. A leitura foi realizada em espectrofotômetro em comprimento de
onda de 540 nm, sendo a atividade de NR determinada segundo Stewart et al. (1986).
d) Floração
Para se avaliar o efeito do cultivo em alto CO2 na indução floral, ou seja, na transição
do estado vegetativo para o estado reprodutivo, foram analisados os seguintes parâmetros,
conforme estabelecido por He et al. (2004): i) número de dias para o aparecimento do
primeiro botão floral desde a semeadura; ii) número de dias para a ocorrência da antese
(abertura do botão floral); iii) intervalo de dias entre o aparecimento do primeiro botão floral e
a antese; iv) número de folhas em roseta e caulinares no dia da antese.
6
e) Massa Fresca, Massa Seca e Teor de água
As plantas foram coletadas ao final do experimento e as folhas foram separadas em
folhas da roseta e caulinares. O peso fresco foi estimado através de pesagem em balança de
precisão. O material foi a seguir acondicionado em sacos de papel Kraft e seco em estufa a 60
°C até peso constante (aproximadamente 48 horas). Após este período, o material foi pesado
novamente para estimativa do peso seco. Para cálculo do teor de água nas folhas, foi usada a
fórmula: H2O%= (MF-MS)/MF, onde MF corresponde a massa fresca e MS a massa seca.
f) Determinação do conteúdo de açúcares totais
Para a quantificação do teor de açúcares solúveis totais (AST), o material vegetal foi
congelado e liofilizado. Cem miligramas de material seco foram adicionados de 1 mL de
etanol 80%, sendo mantidos por 20 min a 80°C. O material foi centrifugado por 5 min a
10000 rpm e o sobrenadante armazenado. O procedimento foi repetido 3 vezes e os
sobrenadantes foram reunidos. O teor de AST do sobrenadante final foi analisado pelo
método fenol-sulfúrico (Dubois et al., 1956), sendo usada glucose (100 µg/mL) como padrão.
As amostras foram lidas em espectrofotômetro no comprimento de onda 490 nm.
h) Determinação de aminoácidos
Duzentos miligramas de material fresco foram extraídos com 2 mL de
metanol:clorofórmio:água (12:5:3 v/v) e mantidos por 24 h a temperatura ambiente. Após
esse período, o material foi centrifugado por 30 min a 2.000 rpm e o sobrenadante foi
coletado. Para cada 4 mL de sobrenadante, foram adicionados 1 mL de clorofórmio e 1,5 mL
de água. O material foi agitado e mantido durante 24 h a temperatura ambiente. A fração
aquosa foi transferida para novo tubo e incubada por 15 h a 38ºC. Posteriormente as amostras
foram centrifugadas a 14.000 rpm por 15 min, o volume do sobrenadante foi medido e
armazenado a -20°C, até o momento da análise. O perfil de aminoácidos foi determinado
HPLC de fase reversa após derivatização com o-phtaldialdeído, segundo Puiatti e Sodek
(1999).
3. RESULTADOS E DISCUSSÃO
Ao analisar a taxa aparente de transporte de elétrons (ETR) em diferentes intensidades
luminosas (PAR) (Fig. 1), constatou-se que não houve diferenças entre as plantas selvagens
cultivada em diferentes concentrações de CO2. No entanto, as taxas de transporte de elétrons
das plantas selvagens foram em média superiores às das plantas mutantes nia, independente
do tratamento. O efeito do cultivo em alto CO2 pôde ser observado para as plantas nia que
apresentaram um PAR de saturação menor (em torno de 400 µmol m-2 s-1) quando cultivadas
7
em 380 ppm de CO2, sendo que em 760 ppm o par de saturação das plantas nia esteve em
torno de 600 µmol m-2 s-1. Além disso, uma queda mais acentuada na taxa de transporte de
elétrons foi observada para as plantas mutantes cultivadas em 380 ppm de CO2.
200
180
ETR ( mol m-2s-1)
160
140
WT 380
120
Nia 380
100
WT 760
80
Nia 760
60
40
20
0
0
200
400
600
800
1000
1200
1400
1600
PAR
Figura 1. Curva de indução da fotossíntese em plantas selvagens (WT) e mutantes (nia) de
Arabidopsis thaliana cultivadas em 380 e 760ppm de CO2. ETR: taxa aparente de transporte de
elétrons e PAR: radiação fotossinteticamente ativa. Plantas com 34 dias. Barras indicam o desviopadrão da média.
Nas plantas nia, a baixa atividade da enzima nitrato redutase (NR) resulta não somente
em menor teor endógeno de óxido nítrico (NO), mas em outras alterações metabólicas
importantes. As folhas destas plantas são mais amareladas, o que sugere menor conteúdo de
clorofila ou alterações na razão clorofila a e b. Embora o teor de clorofilas não tenha sido
quantificado em mutantes Arabidopsis do tipo nia, Yamamoto-Katou e colaboradores (2006)
observaram, em plantas mutantes de tabaco, que a diminuição da atividade de NR resulta em
folhas amareladas, fenótipo similar ao observado em Arabidopsis, e diminuição da
subunidade maior da proteína Rubisco. Estes dois fatores poderiam explicar a menor taxa de
transporte de elétrons e menor eficiência fotossintética dos mutantes nia de Arabidopsis. O
cultivo em alto CO2 parece compensar parcialmente esta deficiência das plantas mutantes.
O efeito da disponibilidade de nitrogênio também afeta a resposta fotossintética em
alto CO2, como discutido por Stitt & Krapp (1999). A concentração de CO2 atmosférico e a
disponibilidade de diferentes formas nitrogenadas (NO3- ou NH4+) influencia a partição da
energia derivada da fotossíntese. Por exemplo, em concentração ambiente de CO2, plantas de
tomate que receberam NO3- como única fonte de nitrogênio tiveram maiores taxas de
transporte de elétrons (ETR) do que plantas que receberam NH4+ (Searles & Bloom, 2003).
Devido à deficiência em assimilar nitrato, as plantas nia de Arabidopsis receberam, no
8
presente estudo, solução nutritiva com predominância de NH4+ como fonte de nitrogênio,
enquanto a solução nutritiva das plantas selvagens possuía as duas fontes nitrogenadas, com
predominância de NO3-. De maneira similar ao observado em tomate, as plantas que
assimilaram NO3- como fonte preferencial de nitrogênio (selvagens) tiveram maiores taxas de
transporte de elétrons.
Na Figura 2, pode-se observar a emissão de óxido nítrico em folhas intactas de plantas
de Arabidopsis submetidas aos diferentes tratamentos. Plantas deficientes nos dois genes
estruturais de nitrato redutase (nia) apresentam níveis reduzidos de óxido nítrico quando
comparadas com as plantas selvagens (WT). Quando cultivadas em CO2 ambiente, os níveis
de NO do mutante nia são próximos de zero (similares aos obtidos para a amostra controle
sem material biológico – branco). No entanto, quando cultivadas em concentrações elevadas
de CO2, os níveis endógenos de NO nas folhas aumentam ligeiramente. O efeito do CO2 na
produção endógena de NO é mais visível nas plantas selvagens, sendo que um aumento
significativo pôde ser observado nestas plantas.
Alguns autores demonstraram que altas concentrações de CO2 podem afetar os
processos de nitrificação e denitrificação por bactérias do solo, alterando a produção de gases
como o óxido nitroso e o óxido nítrico (Hinkson, 1996) e sua liberação posterior na
atmosfera. Gould e colaboradores (2003) observaram um aumento dos níveis de NO em
células de tabaco em suspensão submetidas a diversos estresses abióticos, como altas
temperaturas, estresse osmótico e salinidade.
Concentração de NO (nmol.g-1.min-1)
0,70
0,60
0,50
0,40
0,30
a
0,20
0,10
c
d
0,00
WT 380
WT 760
Nia 380
Nia 760
Tratamentos
Figura 2. Quantificação do teor de NO em folhas de A. thaliana selvagem (WT) e mutante (nia) em
duas diferentes concentrações atmosféricas (380 e 760ppm). Os dados correspondem ao valor máximo
de emissão de NO obtido no comprimento de onda 515 nm. As barras correspondem ao desvio padrão
da média (n = 3).
9
No entanto, não existem relatos do efeito direto do CO2 atmosférico na produção de
óxido nítrico em plantas. Aparentemente, o maior teor de NO nas plantas selvagens de
Arabidospsis cultivadas em alto CO2 pode estar relacionado com um aumento da atividade da
enzima nitrato redutase, observado nestas mesmas plantas (Fig. 3). Uma diferença marcante
pôde ser observada entre as plantas selvagens, sendo que as cultivadas em 760 ppm de CO2
apresentaram taxas de atividade NR 2,5 vezes superiores às das plantas controle. Entretanto,
um aumento mais discreto da atividade de NR nas plantas nia foi observado em elevado CO2.
As plantas mutantes nia em CO2 ambiente foram as que apresentaram as menores atividades
da enzima NR (Fig. 3).
Os resultados do efeito de altas concentrações de CO2 sobre a atividade de NR são
bastante controversos. Embora alguns autores tenham observado uma diminuição da atividade
de NR em trigo, milho e tabaco (Purvis et al., 1974; Hocking & Meyer, 1991; Ferrario-Mery
et al., 1997), um pequeno aumento de atividade pôde ser observado em mostarda (Maevskaya
et al., 1990) e Vigna radiata (Sharma & Sen Gupta, 1990). Fonseca et al. (1997)
demonstraram que o cultivo de Plantago major em elevado CO2 resulta em aumento do nível
de transcritos nia nas folhas e, principalmente, nas raízes e este aumento é acompanhado por
um aumento de 50% da atividade NR nas folhas e 100% de aumento nas raízes. Geiger et al.
(1999) observaram um estímulo acentuado da atividade de NR em plantas jovens de tabaco
cultivadas em alto CO2, mas um aumento similar não foi observado em plantas mais velhas.
350
b
pmoles NO2 s-1 g-1 MF
300
250
200
150
a
100
a
50
a
0
WT 380
WT 760
Nia 380
Nia 760
Tratamentos
Figura 3. Dosagem de atividade da enzima nitrato redutase (NR) em folhas de A. thaliana selvagem
(WT) e mutante (nia) em duas diferentes concentrações atmosféricas (380 e 760ppm). As barras
correspondem ao desvio padrão da média (n = 3).
10
Segundo estes autores, a modulação da atividade de NR pelo CO2 não está relacionada
com o status nutricional da planta, e sim com suas taxas de crescimento relativo.
É importante ressaltar que, embora o mutante nia tenha apresentado taxas de atividade
NR mais baixas que o selvagem, as mesmas são bastante superiores ao 0,5% de atividade
descrita por Wilkinson & Crawford (1993). Uma das explicações pode ser o fato de
Wilkinson & Crawford (1993) terem medido a atividade de NR in vitro, enquanto que neste
trabalho foi medida a atividade in situ. Além disso, Matt et al. (2001) demonstraram que
plantas nia são capazes de compensar a baixa atividade NR por meio de alterações no ciclo
diurno de atividade da enzima. No caso dos mutantes, a NR permanece ativa durante todo o
período luminoso e não é observada retroinibição pelo produto. As taxas de atividade NR nas
plantas Nia são 2 a 3 vezes menores do que nas selvagens na primeira parte do período
luminoso, mas esta diferença tende a diminuir no restante do período. No presente estudo, as
plantas foram coletadas para dosagem de atividade NR no período da tarde, o que pode
explicar diferenças menos acentuadas entre as plantas selvagens e mutantes.
Os dados de massa seca das folhas (Fig. 4) mostram que, embora as plantas cultivadas
em alto CO2 tenham apresentado melhor status fotossintético e maior atividade de NR, o seu
desenvolvimento final foi similar ao das plantas controle, cultivadas em CO2 ambiente. No
entanto, fica claro o menor desenvolvimento foliar das plantas nia em relação às plantas
selvagens.
30
a
a
Massa seca (g)
25
20
b
15
b
10
5
0
WT380
WT 760
nia 380
nia 760
Tratam entos
Figura 4. Massa seca foliar total (folhas da roseta e folhas caulinares) de plantas de A. thaliana
selvagem (WT) e mutante (nia) em duas diferentes concentrações atmosféricas (380 e 760ppm). As
barras correspondem ao desvio padrão da média (WT, n = 14; nia, n = 9).
Ao analisar o teor de água nas folhas (Fig. 5), foi possível observar uma maior
porcentagem de água nas folhas das plantas selvagens cultivadas em elevado CO2. Estes
11
dados estão de acordo com a literatura, sendo que diversos autores já demonstraram
diminuição da condutância estomática e aumento da eficiência do uso da água (WUE) em
plantas de diversas espécies quando cultivadas em alto CO2 (Long et al., 2004; Souza et al.,
2008).
95
c
c
Teor de água (%)
94
93
b
92
a
91
90
WT 380
WT 760
nia 380
nia 760
Tratam entos
Figura 5. Teor de água foliar total (folhas da roseta e folhas caulinares) de plantas de A. thaliana
selvagem (WT) e mutante (nia) em duas diferentes concentrações atmosféricas (380 e 760ppm). As
barras correspondem ao desvio padrão da média (WT, n = 14; nia, n = 9).
Embora um efeito direto do CO2 não seja observado neste parâmetro para as plantas
mutantes nia, há uma diferença significativa no teor de água das folhas destas plantas em
relação às selvagens. Estes dados estão de acordo com observações visuais feitas durante o
experimento de que as plantas nia suportavam melhor a falta de água e recuperavam melhor o
status hídrico após um período sem rega.
O menor desenvolvimento das plantas nia também pôde ser observado por um atraso
na floração nestas plantas (Tabela 1). Na planta selvagem em condições controle, o
aparecimento do botão floral se deu, em média, 46 dias após a semeadura. Na presença de
elevado CO2, as plantas selvagens apresentaram floração precoce, com diferença de
aproximadamente 10 dias entre os dois tratamentos.
Nas plantas mutantes, o CO2 não exerceu efeito sobre o aparecimento do botão floral,
mas houve diminuição significativa no intervalo de dias entre o aparecimento do botão e a
abertura da flor. No entanto, o número de indivíduos nia que tiveram sua floração avaliada foi
baixo com relação às plantas mutantes, pois foram poucos os que apresentaram flor ao
término do experimento.
12
Tabela 1: Parâmetros de indução floral de plantas de A. thaliana selvagem (WT) e mutante (nia) em
duas diferentes concentrações atmosféricas (380 e 760ppm). Os dados correspondem à média ± desvio
padrão; as letras indicam as diferenças estatísticas.
Dias para aparecimento
do primeiro botão floral
Dias para antese
Intervalo de dias entre
aparecimento do botão e
antese
Número de folhas da
roseta e caulinares no dia
da antese
n
WT 380
46,4 (±4,9) a
WT 760
37,8 (±6,5) b
nia 380
50 (±4,2) a
nia 760
51,1 (±2,3) a
53,1 (±4,6) a
6,7 (±1,5) b
44,6 (±7,4) a
6,8 (±1,8) b
61 (±1,4) b
11 (±2,8) a
59,3 (±2,3) b
7,8 (±2,2) b
26,9 (±1,9) a
27,8 (±5,5) a
30,5 (±3,5) a
29,2 (±9,4) a
19
19
2
6
Elevadas concentrações de CO2 podem acarretar mudanças na alocação de biomassa e
na fenologia, incluindo aumento da razão raiz: parte aérea, maior ramificação, maior
perfilhamento em gramíneas e florescimento precoce (Bazzaz & Fajer 1992; Tissue et al.
1997). No caso específico de Arabidopsis thaliana, o tempo de floração é bastante variável
em função das condições ambientais, por ser esta uma planta bastante sensível às condições
de luz e temperatura. Com relação à concentração atmosférica de CO2, um levantamento
recente da literatura feito por Springer e Ward (2007), mostra uma ampla variação do
comportamento de florescimento nas mais diferentes espécies, incluindo Arabidopsis
thaliana, para a qual diferentes autores demonstraram atraso, precocidade ou nenhuma
alteração na floração destas plantas quando cultivadas em alto CO2 (VanderKooij &
DeKok,1996; Ward & Strain, 1997).
Com relação às condições ambientais de cultivo de Arabidopsis do experimento
realizado, as câmaras se encontram em casa de vegetação, sem controle de temperatura,
luminosidade e umidade. Embora estes parâmetros variem, a variação é a mesma em todas as
câmaras. No entanto, estas condições dificultam comparações de florescimento com os
demais trabalhos da literatura. He et al. (2004) demonstraram que o óxido nítrico reprime a
floração em plantas de Arabidopsis ao estudar os mutantes nox1 e cue1-5 que produzem
maiores quantidades de NO do que as plantas selvagens. Seligman e colaboradores (2008)
observaram floração precoce em mutantes nia de Arabidopsis deficientes na produção de NO,
corroborando os dados de He et al. (2004). Neste estudo, a floração precoce foi observada nas
plantas que produziram mais NO. No entanto, nos dois trabalhos as plantas foram cultivadas
em condições controladas de luz e temperatura, sendo a temperatura média de 22oC e a
luminosidade máxima de aproximadamente 100 µmol m2 s-1. Além disso, no caso do
experimento de Seligman et al. (2008), as plantas nia receberam solução nutritiva com
13
aproximadamente 5 vezes mais nitrogênio do que a das plantas selvagens, diferentemente do
presente estudo, no qual as soluções foram ajustadas para uma mesma molaridade de
nitrogênio.
Nas condições estudadas, o fenótipo de florescimento precoce não foi observado nas
mutantes nia, e o CO2 não influenciou a floração nestas plantas. No entanto, com relação às
plantas selvagens em CO2 ambiente, o tempo de floração é compatível com o observado para
plantas WT por He et al. (2004), sendo que a precocidade na floração estimulada pelo alto
CO2 corrobora o que foi observado previamente para outras espécies (Tissue et al. 1997;
Springer & Ward, 2007).
Na figura 6, observa-se que as plantas selvagens apresentaram um aumento
significativo no de teor de açúcares solúveis nas folhas em CO2 elevado. Um pequeno
aumento na concentração de AST foi observado nas plantas mutantes em alto CO2, que
b
Concetração AST (ug/mg MF)
2,0
1,8
1,6
a
ac
1,4
1,2
c
1,0
0,8
0,6
0,4
0,2
0,0
WT 380
WT 760
Nia 380
Nia 760
Tratamentos
Figura 6. Quantidade de açúcar solúvel em folhas de A. thaliana selvagem (WT) e mutante (nia) em
duas diferentes concentrações atmosféricas (380 e 760ppm). As barras correspondem ao desvio padrão
da média (n = três plantas por triplicata em cada tratamento).
apresentaram níveis similares aos da planta selvagem em condições ambientais normais. Este
aumento no teor de AST em alto CO2 é um fenômeno bastante conhecido e discutido na
literatura e está relacionado ao aumento das taxas fotossintéticas. Esse mesmo efeito foi
observado por Moore et al. (1998) em plantas de A. thaliana ecótipo Columbia cultivadas em
400 e 1000 ppm de CO2.
Ao analisar o conteúdo dos diferentes aminoácidos nas folhas (Fig. 7), observa-se que
os aminoácidos predominantes nos diferentes tratamentos são Asp (asparagina), Glu
(glutamato) e Glu (glutamina). Um efeito do aumento de CO2 atmosférico não foi visível no
14
teor dos diferentes aminoácidos nas plantas selvagens. Já as plantas nia apresentaram um
aumento na quantidade de Asp, Glu, Gln e Arg (arginina) quando cultivadas em
concentrações elevadas de CO2.
Geiger et al. (1999) cultivaram plantas de tabaco em CO2 ambiente e elevado e
verificaram que a quantidade de arginina nas folhas é 8,5 vezes maior em alto CO2. Estes
autores também observaram um aumento nos teores de Glu e Gln em alto CO2. Esse resultado
é similar ao observado no presente estudo.
Figura 7. Conteúdo de aminoácidos em plantas de A. thaliana selvagem (WT) e mutante (nia) em
duas diferentes concentrações atmosféricas (380 e 760ppm). As barras correspondem ao desvio padrão
da média (n=3).
Como pode ser observado na figura 8, glutamina e glutamato são precursores da
síntese de arginina. A síntese de NO pela enzima óxido nítrico sintase (NOS) é dependente de
arginina, como demonstrado por Corpas et al. (2006) em tecidos de plântulas de ervilha em
diferentes estágios do desenvolvimento.
15
Figura 8. Vista parcial da rota metabólica de síntese de Arginina, com seus precursores e a
participação do CO2 nos diferentes pontos da via. Fonte: Cambuí (2009).
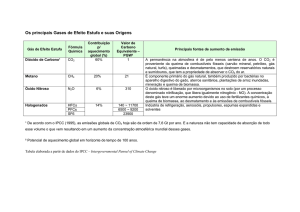
Levando-se em conta o conjunto dos resultados obtidos, pode-se sugerir que o
aumento de CO2 atmosférico leva a um aumento da síntese de NO nas plantas selvagens via
aumento da atividade da enzima nitrato redutase (NR). Já nas plantas nia, a atividade de NR
deficitária não pouco afetada em alto CO2. Portanto, o aumento de NO observado nestas
plantas mutantes poderia estar relacionado a um aumento de glutamina e glutamato e,
consequentemente de arginina, levando a um aumento de NO via enzima óxido nítrico sintase
(NOS).
4. REFERÊNCIAS BIBLIOGRÁFICAS
Aidar, M.P.M.; Martinez, C.A.; Costa, A.C.; Costa, P.M.F.; Dietrich, S.M.C. &
Buckeridge, M.S. 2002. Effect of atmosferic CO2 enrichment on the establishment of
seedlings of jatobá, Hymenaea courbaril L. (Leguminosae, Caesalpinioideae). Biota
Neotrópica 2: 1-10.
Bazzaz, F.A. & Fajer, E.D. 1992. Plant life in a CO2-rich world. Scientific American 266:
68-74.
Bowler, J.M. & Press, M.C. 1996. Effects of elevated carbon dioxide, nitrogen form and
concentration on growth and photosynthesis of fast- and slow-growing grass. New
Physiologist 132: 391-401.
Cambuí, C.A. 2009. Absorção e assimilação de uréia em folhas de bromélia: clonagem e
expressão de aquaporinas, atividade e citolocalização de urease e análise de aminoácidos.
Tese de doutorado, Universidade de São Paulo, São Paulo.
16
Cheng, C., Dewdney. J., Nam. H., den Boer, B.G.W. & Goodman, H.M. 1988. A new
locus nia 1 in Arabidopsis thaliana encoding nitrate reductase. The EMBO Journal 7:
3309 – 3314.
Corpas, F.J., Barroso, J.B., Carreras, A., Valderrama, R., Palma, J.M., Leon, A.M.
León, Sandalio, L.M. & del Rio, L.A. 2006. Constitutive arginine-dependent nitric oxide
synthase activity in different organs of pea seedlings during plant development. Planta
224: 246-254.
Crawford, N.M. 1995. Nitrate: nutrient and signal for plant growth. The Plant Cell
7: 859-868.
Dean, J.V. & Harper, J.E. 1986. Nitric oxide and nitrous oxide production by soybean and
winged bean during the in vivo nitrate reductase assay. Plant Physiology 82: 718-723.
Dubois, M., Gilles, A., Hamilton, J.K., Rebers, P.A. & Smith, F. 1956. Colorimetric
method of determination of sugars and related substances. Analytical Chemistry 28: 350355.
Ferrario-Mery, S., Thibaud, M.-C., Betsche, T., Valadier, M.-H. & Foyer, C.H. 1997.
Modulation of carbon and nitrogen metabolism and of nitrate reductase in transformed
Nicotiana plumbaginifolia during CO2 enrichment of plants grown in pots and hydroponic
culture. Planta 202: 510-521.
Fonseca, F., Bowsher, C. & Stulen, I. 1997. Impact of elevated atmospheric carbon dioxide
on nitrate reductase transcription and activity in leaves and roots of Plantago major.
Physiologia Plantarum 100: 940-480.
Geiger, M., Haake, V., Ludewig, F., Sonewald, U. & Stitt, M. 1999. Influence of nitrate
and ammonium nitrate supply on the response of photosynthesis, carbon and nitrogen
metabolism and growth to elevated carbon dioxide in tobacco. Plant, Cell and
Environment 22: 1177-1199.
Gould, K. S., Lamotte, O., Klinguer, Pugin ,A. & Wendehenne, D. 2003. Nitric oxide
production in tobacco leaf cells: a generalized stress response? Plant, Cell and
Environment 26:1851–1862.
Hardy, R.W.F & Havelka, V.P. 1975. Nitrogen fixation research: a key to world food.
Science 188: 633-643.
He, Y., Tang, R.H., Hao, Y., Stevens, R.D., Cook, C.W., Ahn, S.M., Jing, L., Yang, Z.,
Chen, L., Guo, F., Fiorani, F., Jackson, R.B., Crawford, N.M. & Pei, Z.M. 2004.
Nitric oxide represses the Arabidopsis floral transition. Science 35: 1968-1971.
Hinkson, C.L. 1996. Gas exchange, growth response and nitrogen dynamics of Quercus
agrifolia atmosferic CO2 and water stress. Master Thesis, San Diego State University.
17
Hocking, P.J. & Meyer, C.P. 1991. Effects of CO2 Enrichment and Nitrogen Stress on
Growth, and Partitioning of Dry Matter and Nitrogen in Wheat and Maize. Australian
Journal of Plant Physiology 18, 339–356.
Li, P., Sioson, A., Mane, S.P., Ulanov, A., Grothaus, G., Heath, L.S., Murali, T.M., Hans
J. Bohnert, H.J. & Grene, R. 2006. Response diversity of Arabidopsis thaliana ecotypes
in elevated CO2 in the field. Plant Molecular Biology 62: 593-609.
Long ,S.P., Ainsworth, E.A., Rogers, A. & Ort, D.R. 2004. Rising atmospheric carbon
dioxide: plants FACE the future. Annual Reviews of Plant Biology 55: 591–628.
Maevskaya, S.N., Andreeva, T.F., Voedudskaya, S.Y.U., Cherkanova, S. 1990. Effect of
high carbon dioxide concentration on photosynthesis and nitrogen metabolism in leaf
mustard plants. Fiziologia Rastenii 37, 921–927.
Magnusson, G., Larsson, C. & Axelsson, L. 1996. Effects of high CO2 treatment on nitrate
and ammonium uptake by Ulva lactuga grown in different nutrient regimes. Scientia
Marina 60: 179-189.
Matt, P., Geiger, M., Walch-Liu, P., Engels, C., Krapp, A. & Stitt, M. 2001. Elevated
carbon dioxide increases nitrate uptake and nitrate reductase activity when tobacco is
growing on nitrate, but increases ammonium uptake and inhibits nitrate reductase activity
when tobacco is growing on ammonium nitrate. Plant, Cell & Environment 24: 11191137.
Matt, P., Schurr, U., Krapp, A. & Stitt, M. 1998. Growth of tobacco in short day conditions
leads to high starch, low sugars, altered diurnal changes of the nia transcript and low
nitrate reductase activity and an inhibition of amino acid synthesis. Planta 207: 27-41.
Meyer, C., Hirel, B., Morot-Gaudry, J.F. & Caboche, M. 1993. L’utilisation de l’azote par
les plantes. La Recheche 257: 956-962.
Miller, A., Tsai, C.-H., Hemphill, D., Endres, M., Rodermel, S. & Spalding, M. 1997.
Elevated carbon dioxide effects during leaf ontogeny. Plant Physiology 115: 1195-1200.
Modolo, L.V., Augusto, O., Almeida, I.M.G., Pinto-Maglio, C.A.F., Oliveira, H.C.,
Seligman, K. & Salgado, I. 2006. Decreased arginine and nitrite levels in nitrate
reductase-deficient Arabidopsis thaliana plants impair nitric oxide synthesis and the
hypersensitive response to Pseudomonas syringae. Plant Science 17: 34-40.
Moore, B.D., Cheng, S.H., Rice, J. & Seemann, J. R. 1998. Sucrose cycling, Rubisco
expression, and prediction of photosynthetic acclimation to elevated atmospheric CO2.
Plant, Cell and Environment 21: 905–915.
Nielsen, T.H., Krapp, A., Röper-Schwarz, U. & Stitt, M. 1998. The sugar-mediated
regulation of genes encoding the small subunit of Rubisco and regulatory subunit of ADP
18
glucose pyrophosphorylase is modified by phosphate and nitrogen. Plant, Cell and
Environment 21:443-454.
Puiatti, M. & Sodek, L. 1999. Waterlogging affects nitrogen transport in the xylem of
soybean, Plant Physiol. Biochem. 37: 767–773.
Purvis, A.C., Perters, D.B. & Hageman, R.H. 1974. Effect of carbon dioxide on nitrate
accumulation and nitrate reductase induction in corn seedlings. Plant Physiology 53: 934941.
Riviere-Rolland, H., Contard, P. & Betsche T. 1996. Adaptation of pea to elevated
atmospheric CO2: Rubisco, PEP carboxylase and chloroplast phosphate translocator at
different levels of nitrogen and phosphate nutrition. Plant, Cell and Environment 19: 109117.
Rockel, P. F., Strube, F., Rockel, A., Wildt, J. & Kaiser, W.M. 2002. Regulation of nitric
oxide (NO) production by plant nitrate reductase in vivo and in vitro. Journal of
Experimental Botanical 53: 1-8.
Sage, R.F. 1994. Acclimatation of photosynthesis to increasing atmospheric carbon dioxide,
the gas exchange perspective. Photosynthesis Research 39: 351-368.
Scheible, W.R., Lauere, M., Schulze, E.D., Caboche, M. & Stitt, M. 1997. Accumulation
of nitrate in the shoot acts as signal to regulate shoot-root allocation in tobacco. Plant
Journal 11: 671-691.
Searles, P. S. & Bloom, A. J. Nitrate photo-assimilation in tomato leaves under short-term
exposure to elevated carbon dioxide and low oxygen
Seligman, K., Saviani, E.E., Oliveira, H.C., Pinto-Maglio, C. A.F., & Salgado, I. 2008.
Floral transition and nitric oxide emission during flower development in Arabidopsis
thaliana is affected in nitrate reductase-deficient plants. Plant Cell Physiology. 49: 11121121.
Sharma, A. & Sen Gupta, U.K. 1990. Carbon dioxide effects on photosynthesis and related
enzymes in Vigna radiate L. Wilczec. Indian Journal of Plant Physiology 33: 340-346.
Souza, A.P., Gaspar, M., Silva, E.A., Ulian, E.C., Waclawovsky, A.J., Nishiyama-Jr.,
M.Y., Santos, R.V., Teixeira, M.M., Souza, G.M. & Buckeridge, M.S. 2008. Elevated
CO2 increases photosynthesis, biomass and productivity, and modifies gene expression in
sugarcane. Plant Cell & Environment 31: 1116-1127.
Springer, C.J. & Ward, J.K. 2007. Flowering time and elevated atmospheric CO2. New
Physiologist 176: 243–255.
19
Stewart, G.R., Popp, M., Holzapfel, I., Stewart, J.I. & Dickie-Eskew, A. 1986.
Localization of nitrate reduction interns and its relationship to environmental and
physiological characteristics. New Phytologist 104: 373-384.
Stitt, M. & Krapp, A. 1999. The interaction between elevated carbon dioxide and nitrogen
nutrition: the physiological and molecular background. Plant, Cell and Environment 22:
583-621.
Stuehr, D.J. 1997. Structure-function aspects in the nitric oxide synthases. Annual Review
Pharmacology Toxicology 37: 339-359.
Tischner, R., Peuke, A., Godbold, D.L., Feig, R., Merg, G. & Hüttermman A. 1988. The
effect of NO2-fumigation on aseptically grown spruce seedlings. Journal of Plant
Physiology 133: 243-246.
Tissue, D.T., Thomas, R.B. & Srain B.R. 1997. Atmospheric CO2 enrichment increases
growth and photosynthesis of Pinus taeda, a 4-year experiment in the field. Plant, Cell &
Environment 20: 1123-1134.
VanderKooij, T.A.W. & DeKok, L.J. 1996. Impact of elevated CO2 on growth and
development of Arabidopsis thaliana L. Phyton – Annales Rei Botanicae 36: 173–184.
Ward, J.K.& Strain, B.R. 1997. Effects of low and elevated CO2 partial pressure on growth
and reproduction of Arabidopsis thaliana from different elevations. Plant, Cell &
Environment 20: 254–260.
Wilkinson, J.Q. & Crawford, N.M. 1991. Identification of the Arabidopsis chl3 gene as the
nitrate-reductase structural gene nia 2. Plant Cell 3: 461-471.
Wilkinson, J.Q. & Crawford, N.M. 1993. Identification and characrization of a chlorateresistent mutant of Arabidopsis thaliana with mutations in both nitrate reductase structural
genes nia 1 and nia 2. Molecular General Genetics 239: 289-297.
Yamamoto-Katou, A., Katou, S., Yoshioka, H.,
Doke, N. & Kawakita, K.
Nitrate
Reductase is Responsible for Elicitin-induced Nitric Oxide Production in Nicotiana
benthamiana. Plant & Cell Physiology 47: 726-735.
Yu, X., Sukumaran, S. & Márton, L. 1998. Differential expression of the Arabidopsis nia 1
and nia 2 genes. Plant Physiology 116: 1091-1096.
20