Fisiologia de Sistemas II – 2º Módulo
Fisiologia de Sistemas II – 2º Módulo.
Sangue e as suas propriedades visco elásticas.
Viscosidade
A velocidade de cisalhamento, ou seja, a velocidade com que varia a
deformação de um líquido, é proporcional ao débito e inversamente proporcional ao
cubo do raio do vaso sanguíneo, portanto quando o raio do vaso aumenta ou diminui um
pouco, ocorrem grandes variações na velocidade de cisalhamento.
dβ/dt = 4Q/ πr3
dβ/dt – velocidade de cisalhamento
Q – débito
r – raio do vaso
O sangue é um líquido que possui viscosidade anómala, a que se chama
viscosidade aparente, que resulta do facto de o sangue não ser um líquido Newtoniano
(solução homogénea). O sangue não possui uma viscosidade constante, e pode
considerar-se grosseiramente como uma suspensão de glóbulos vermelhos em plasma.
Variação da Viscosidade do sangue
A viscosidade do sangue diminui quando a sua temperatura aumenta, por outro
lado a viscosidade do sangue varia com o hematócrito. O hematócrito é a fracção de
volume do sangue que é devida aos glóbulos vermelhos, dito de outra forma, é o valor
que exprime a relação entre o volume de glóbulos vermelhos no sangue e o volume total
deste.
Portanto se o hematócrito for
zero, só temos plasma. De acordo
com o gráfico, toma-se a viscosidade
do plasma (que corresponde ao
hematócrito = 0), como sendo igual a
1. O gráfico mostra a variação da
viscosidade do sangue, relativamente
ao
plasma,
em
função
do
hematócrito. O hematócrito normal
está à volta dos 40.
Anomalias na viscosidade do sangue
Existem dois tipos de anomalias na viscosidade do sangue: a anomalia de baixa
velocidade de cisalhamento e a anomalia de pequeno raio do vaso.
- Anomalia de baixa velocidade de cisalhamento
Fisiologia de Sistemas II – 2º Módulo
Para baixas velocidades de
cisalhamento (rate of shear), os
glóbulos vermelhos colam-se/aderem
uns aos outros formando rolos, os
chamados “rouleaux” em francês, que
se juntam uns aos outros, ainda,
formando agregados ainda maiores.
Desta maneira para tensões de
cisalhamento
(shearing
stress),
pequenas o sangue comporta-se como
um sólido. O sangue é um pseudo
plástico.
Começa
por ter o
comportamento de um sólido para tensões de cisalhamento pequenas, de seguida chega
à tensão de cedência, e a força que promove a desagregação – a tensão de cisalhamento
– é maior do que a força que promove a agregação dos glóbulos vermelhos, e estes
começam a separar-se uns dos outros. Por outro lado a viscosidade, que corresponde à
inclinação da tangente à curva em cada ponto, é maior para baixas velocidades de
cisalhamento e para baixas tensões de cisalhamento e vai diminuindo com o aumento
destes dois parâmetros. Com o aumento da tensão de cisalhamento os rouleaux vão se
desagregando, e a dada altura os glóbulos vermelhos estão todos separados, a partir de
então a viscosidade passa a ser constante. Chama-se a esta viscosidade, viscosidade
assimptótica. Todos estes acontecimentos sucedem-se por haver fibrinogénio no
sangue, uma vez que é devido ele que os glóbulos vermelho formam agregados.
5:15WMP
Aqui temos a variação da viscosidade com a velocidade de cisalhamento. Em
ambos os eixos estão escalas logarítmicas, o que significa que ocorrem variações muito
grandes. A baixas velocidades de cisalhamento tem-se viscosidade muito alta, com a
formação dos rouleaux, e para altas velocidades de cisalhamento (Shear Rate), temos
os glóbulos isolados e uma viscosidade baixa.
- Arteríolas: as arteríolas têm pequeno raio, portanto a velocidade de cisalhamento é
grande –> (dβ/dt = 4Q/ πr3). Como a velocidade. de cisalhamento é grande nestes vasos
não se formam, geralmente, rouleaux nas arteríolas.
Fisiologia de Sistemas II – 2º Módulo
- Vénulas: as vénulas têm um raio muito maior que as arteríolas, portanto a velocidade
de cisalhamento é muito mais baixa, e é normal a formação de rouleaux nestes vasos.
- Artérias: na aorta por exemplo, uma vez que o fluxo é pulsátil, a velocidade de
cisalhamento varia muito ao longo do ciclo cardíaco, e, consequentemente, a
viscosidade varia também muito durante o ciclo cardíaco nas artérias.
- Anomalia de pequeno raio
Verificou-se que para diâmetros inferiores a 500 µm, à medida que diminui este
diâmetro, a viscosidade diminui. Ora isso sucede porque em todos os vasos há uma
pequena camada (cerca de 5 µm), junto à parede onde só existe plasma. Isto num grande
vaso não é muito significativo, uma vez que se pode considerar tudo uniforme. Mas
num capilar os eritrócitos circulam no eixo do vaso, e tudo o resto é a tal camada de
plasma, de maneira que a viscosidade se torna muito mais próxima da viscosidade do
plasma num capilar do que num grande vaso. A quantidade relativa de plasma é muito
grande num capilar, o que faz diminuir imenso a viscosidade nestes vasos.
12:27WMP
Músculo Cardíaco
As células do músculo cardíaco subdividem-se e vão se ligar a outras células. As
células estão ligadas por discos intercalares, que são junções gap de grande condutância,
que permitem a passagem de iões, e a fácil propagação de potenciais de acção de uma
fibra muscular para outra. As junções gap também impedem que as fibras musculares se
separem umas das outras . O músculo tem aspecto estreado. As células musculares
cardíacas formam um sincício (syncytium) (o que as distingue das células musculares
esqueléticas), que contrai todo ao mesmo tempo.
No coração existe um sincício para as aurículas e um sincício para os
ventrículos, separados pelo esqueleto fibroso das válvulas. As válvulas
auriculoventriculares são a bicúspide ou mitral e a tricúspide, e as válvulas semilunares
são a aórtica e a pulmonar. Existe apenas um ponto em que há comunicação entre o
músculo auricular e o músculo ventricular, pelas fibras condutoras do potencial de
acção, que é o feixe de His.
- Estrutura
As fibras musculares têm muitas mitocôndrias, em consequência da grande
necessidade de ATP para a contracção muscular. As fibras musculares são muito
vascularizadas, existem muitos capilares, quase um capilar por fibra, de modo a que
todo o músculo receba oxigénio. Existem umas invaginações do sarcolema, os tubulos
T, na zona da banda Z, e desta maneira o líquido intersticial penetra bem no interior das
fibras musculares. O retículo sarcoplasmático é muito desenvolvido e rodeia as fibras.
Fisiologia de Sistemas II – 2º Módulo
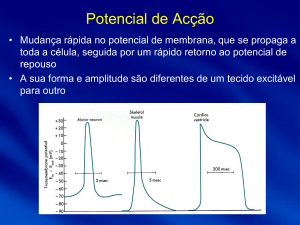
- Potencial de acção do músculo cardíaco
1- resposta rápida: (ocorre nas fibras miocárdicas atriais e ventriculares, no feixe de His
e nas fibras de Purkinje)
O potencial de acção das fibras cardíacas apresenta um patamar, e é diferente do
potencial de acção do músculo esquelético. Este patamar resulta do seguinte: começa
por ocorrer a entrada de iões sódio na célula, por canais de abertura rápida permeáveis
ao Na+, e que dão origem ao spike, e de seguida, mais lentamente, começam a abrir os
canais de cálcio, e a entrada de Ca++ na célula origina, portanto, o tal plateau que dura
0.2/0.3 segundos, e o músculo está contraído durante todo este tempo. De seguida
ocorre a repolarização pelo aumento da condutância ao potássio que vai sair da célula e
obrigar o potencial de membrana a baixar para o potencial de repouso.
2- resposta lenta: (ocorre no nodo sinoatrial – SA – e no nodo atrioventricular – AV)
A despolarização nestes locais do coração resulta do influxo de Ca++ e de Na+
para o interior da célula por canais lentos originando uma resposta mais lenta.
Existe um período refractário absoluto (PRA) durante o qual não pode ser
disparado outro potencial de acção, e existe um período refractário relativo (PRR) onde
já pode ocorrer um potencial de acção sobreposto sobre o anterior.
A amplitude do potencial de acção depende da concentração exterior de Na+, da
concentração exterior de K+ e da concentração exterior de Ca++ mas, por sua vez, o
potencial de repouso não depende da concentração exterior de Na+. Para aumentos da
concentração exterior de K+ o potencial de repouso da membrana varia muito,
aproximando-se do valor dado pela equação de Nernst, mas para baixas concentrações
de K+ o potencial de repouso já não varia tanto.
Fisiologia da Contracção
O cálcio é muito importante na contracção do coração. O que sucede é o
seguinte: quando um potencial de acção se espalha ao longo do sarcolema e dos tubulos
T durante o período do plateau, aumenta a condutância ao Ca++ que vai penetrar no
sarcoplasma. Este cálcio que entra devido ao potencial de acção vai provocar a abertura
Fisiologia de Sistemas II – 2º Módulo
dos canais de Ca2+ existentes no retículo sarcoplasmático (RS) e a consequente saída de
Ca2+ do retículo para o sarcoplasma, ou seja, vai aumentar ainda mais a concentração de
cálcio no interior da célula. Todo este cálcio vai-se ligar à proteína troponina C,
formando o complexo Ca++-troponina, que vai modificar a ligação da troponina à
tropomiosina, libertando assim o locais de ligação entre a miosina e a actina, dando
assim o início ao ciclo das pontes cruzadas, e portanto à contracção das fibras
musculares (sístole). Seguidamente começa a saída de cálcio do citoplasma para o
retículo sarcoplasmático e para o meio extracelular. Os canais existentes no retículo
sarcoplasmático fecham-se e o RS passa a captar Ca2+ do citoplasma activamente, com
gasto de ATP. A diminuição da concentração citoplasmática de Ca2+ reverte formação
do complexo Ca2+-troponina, e portanto, a tropomiosina bloqueia de novo os locais de
ligação entre a miosina e a actina ocorrendo o relaxamento (diástole).
Modelo físico do músculo cardíaco
Montando um elemento contráctil, um elemento elástico (em série) e uma carga,
obtém-se o modelo de um músculo. A tensão estabelecida pela carga no elemento
elástico chama-se tensão inicial, e a carga chama-se pré carga. Ora ao contrair o
elemento contráctil, o elemento elástico sofre extensão e o músculo fica com o mesmo
tamanho. Neste caso temos uma contracção isométrica. Continuando a contrair o
músculo consegue vencer a carga e diminui de tamanho. A carga passa a ter o nome de
pós carga.
Existe uma lei (ND: a professora não disse o nome), que foi estabelecida 1º
empiricamente, que diz que quanto maior for o comprimento inicial do elemento
contráctil (músculo), maior será a força de contracção, portanto quanto mais distendido
estiver o coração mais fortemente contrairá depois. Isto sucede apenas até um certo
ponto, pois se o coração estiver distendido demais, se as fibras estiverem muito
compridas, a força de contracção passa a diminuir com o aumento do comprimento.
Vamos agora ver a nível do sarcómero o que isto quer dizer. Para 2,2 µm de
comprimento do sarcómero a tensão é máxima e continua máxima até 1,95 µm e de
seguida começa a decrescer. Para 3,65 µm a tensão é nula. Ora este comprimento de
sarcómero corresponde à situação em que os filamentos actina não estão em contacto
com os filamentos de miosina, sendo normal não haver força nenhuma. Para um
comprimento de sarcómero igual a 2,2 µm todas as cabeças de miosina estão em
contacto com a actina, e portanto temos a tensão máxima. Se formos diminuindo o
comprimento do sarcómero até 1,95µm continuamos a ter tensão máxima, uma vez que
no meio dos filamentos de miosina não há cabeças para se ligarem à actina.
Continuando a diminuir o comprimento do sarcómero os filamentos de actina começam
a sobrepor-se e nesta situação tem-se perda de força, e quando se chega a 1,6µm os
filamentos de miosina começam a enrugar. No entanto verificou-se que a diminuição de
tensão, de força, não era justificada apenas por esta razão, é um facto que
comprimentos mais pequenos do sarcómero conduzem a uma perda de força, mas não
justificam na totalidade essa perda. O que justifica também esta perda de força, é uma
variação na velocidade com que o cálcio sai do RS, e uma variação da sensibilidade dos
filamentos de actina e miosina à concentração de cálcio. Portanto para maiores
comprimentos de sarcómero a curva de tensão do músculo é de determinada forma, e
para comprimentos menores, vamos ter a mesma tensão se tivermos uma grande
concentração de cálcio, uma vez que a afinidade ao cálcio é menor.
30:00WMP
Fisiologia de Sistemas II – 2º Módulo
Propriedades da fibra cardíaca
1 – Automatismo ou cronotropismo
2 – Condutibilidade ou dromotropismo
3 – Excitabilidade ou batmotropismo
4 – Contractilidade ou inotropismo
5 – Tónus
Crono significa tempo, dromo vem de correr, batmo é de linear e inos vem de fibra.
Automatismo ou cronotropismo
32:49WMP
Como se sabe o coração tem automatismo, no nodo sinosal (SA) formam-se
espontaneamente potenciais de acção o mesmo se passa no nodo AV. No caso de
doença estes potenciais podem mesmo formar-se em qualquer parte do músculo
cardíaco, são os focos ectópicos(?). Mas no coração saudável a frequência é marcada
pelo nodo sinosal. Cada potencial gera normalmente um batimento cardíaco, desta
forma, o ritmo cardíaco, ou a frequência cardíaca é marcada pelo ritmo de geração de
impulsos pelo nodo SA.
Alterações na frequência cardíaca por modificações do pacemaker potencial:
a) o potencial de acção no nodo sinosal é
precedido por um pré potencial (PP) que pode
ocorrer a diferentes velocidades (a inclinação do PP
varia ), assim quanto mais lento for este pré
potencial menor a frequência cardíaca
b) se for aumentado o limiar de excitação da
célula, a frequência também vai baixar, uma vez
que o PP vai ser mais longo, ou seja, menos
impulsos por unidade de tempo ou seja menor
frequência.
c) se a célula for hiperpolarizada, então a
diferença entre o limiar de excitação e o potencial
diastólico máximo aumenta, e portanto o PP fica
mais longo e consequentemente a frequência
diminui.
O sistema nervoso simpático vai actuar sobre as células do nodo sinosal de modo
a aumentar a frequência cardíaca. O que o SNS faz é aumentar a inclinação do PP de
modo a que este fique mais curto, e os potenciais sejam uns mais em cima dos outros,
ou seja menos espaçados no tempo. O parasimpático actua de maneira oposta. O SNPS
vai hiperpolarizar as células ou aumenta-lhes o limiar de excitação de maneira a
diminuir a frequência cardíaca.
A frequência também pode variar por acção de um bloqueador dos canais de
cálcio. Neste caso, na presença de um bloqueador, o potencial fica mais lento, e portanto
diminui a frequência, e por outro lado como há menos cálcio a amplitude do potencial
de acção é menor.
Fisiologia de Sistemas II – 2º Módulo
Condutibilidade ou dromotropismo
O potencial de acção é conduzido no coração por fibras especiais. O impulso
resultante do nodo sinosal é conduzido pela aurícula para o nodo atrioventricular, que
por sua vez conduz o impulso para o feixe de His e daí para as fibras de Purkinje que
transmitem o impulso para o miocárdio ventricular.
A condutibilidade pode tornar-se mais rápida por acção do SNS, e mais lenta por
acção vagal, ou seja, por acção do SNPS, devido a anoxia (falta de oxigénio), por
intoxicação, inflamação etc.
Quando a condução deixa de se efectuar, por qualquer razão, num local do
coração diz-se que há um bloqueio. Como principais bloqueios temos o bloqueio
sinoauricular, entre o nodo SA e a aurícula, o bloqueio auriculoventricular, no nodo
AV e o bloqueio de ramo nas fibras de Purkinje que pode ser no ramo esquerdo ou no
ramo direito. Ambos podem ser completos ou incompletos, consoante deixem passar
algum estímulo ou nenhum. O bloqueio completo do ramo direito não é grave, mas do
ramo esquerdo já uma doença grave.
37:30WMP
Excitabilidade ou batmotropismo
O músculo cardíaco é excitável, ou seja contrai quando é estimulado por
determinados estímulos. Estes estímulos podem ser mecânicos, eléctricos, térmicos
químicos e outros...
Durante a sístole é totalmente refractário qualquer tipo de excitação, uma vez
que o músculo se encontra no período refractário absoluto. A excitabilidade é máxima
depois de várias oscilações e imediatamente a seguir ao período refractário relativo.
A intensidade mínima dos estímulos eficazes – estímulos passíveis de causar
excitação – varia, na razão inversa, com a excitabilidade. Existe um período latente de
excitação, quer isto dizer, que se faz a excitação num instante e o potencial de acção só
aparece passado algum tempo.
Leis da excitação do músculo cardíaco:
•
1ª Lei do tudo-ou-nada
No músculo esquelético estímulos de intensidade crescente vão originar no músculo
potenciais de acção cada vez maiores. No músculo cardíaco não se passa isto. Ou seja
no músculo cardíaco ou não acontece nada, o estimulo não atinge o limiar de excitação,
ou então se atinge o potencial de acção formado tem sempre a mesma amplitude e a
mesma intensidade, qualquer que seja a intensidade do estímulo.
•
2ª Lei da soma
No músculo cardíaco um estímulo sub limiar não origina um potencial de acção,
mas dois estímulos sub limiares com um intervalo de tempo muito pequeno entre eles
podem somar-se, atingir o limiar de excitação e originar potencial de acção.
•
3ª Fenómeno em escada (ND: a professora aqui não explica, mas calculo que
isto se passe no músculo esquelético e não no músculo cardíaco)
Estímulos de igual intensidade em intervalos de tempo relativamente curtos, temos
um 1º potencial normal, de seguida, devido a um 2º estimulo, obtemos um outro
Fisiologia de Sistemas II – 2º Módulo
potencial de acção maior, e de seguida, devido ao 3º estímulo, outro potencial ainda
maior.
•
4ª Potenciação
Quando o coração está a bater pode surgir uma extra sístole. Quando esta surge o
intervalo de tempo para o batimento seguinte é muito maior, e a amplitude da
contracção a seguir à extra sístole é muito maior. A isto chama-se potenciação pós extra
sistólica.
Contractilidade ou inotropismo
É uma medida da capacidade do músculo para realizar trabalho, para uma
determinada pré carga e uma determinada pós carga.
É uma manifestação de todos os factores que influenciam as interacções entre as
proteínas de contracção. Os efeitos de um agente inotrópico podem ser positivos se
aumentam o inotropismo, e negativos se diminuem. O SNS aumenta o inotropismo, e o
SNPS faz o oposto. Também há agentes químicos que aumentam e que diminuem o
inotropismo
Tónus
É a eficiência mecânica, e é igual á energia mecânica de trabalho útil feito pelo
coração sobre a energia total de consumo de oxigénio.
O Coração como uma bomba
02:41
O coração é uma víscera oca com paredes visco elásticas, que constitui uma
bomba dupla: o coração direito que vai lançar o sangue para os pulmões para ser
oxigenado; o coração esquerdo que lança o sangue para a circulação sistémica.
A circulação pulmonar é uma circulação de baixas pressões, e a circulação
sistémica é uma circulação de altas pressões.
O potencial de acção sofre um ligeiro atraso quando passa pelo feixe
auriculoventricular, de maneira que as aurículas contraem ligeiramente antes que os
ventrículos, o que permite encher mais os ventrículos antes de eles expulsarem o
sangue.
O ventrículo direito vai lançar o sangue para uma circulação de baixas
pressões, portanto tem de vencer uma pressão baixa, ou seja, para expulsar o sangue do
seu interior, o ventrículo direito tem de vencer o sangue que está na artéria pulmonar a
uma pressão baixa. O ventrículo direito tem 3 tipos de contracção. (1ª) Uma contracção
em que a válvula tricúspide se aproxima da extremidade do coração, que é muito
evidente, mas não é a que expulsa mais sangue. (2ª) A que faz expulsar mais sangue é a
chamada acção de fole. A parede externa aproxima-se do septo auriculoventricular, e
ao mesmo tempo a contracção do ventrículo esquerdo puxa também a parede e faz
expulsar mais sangue, e o ventrículo direito expulsa o sangue como um fole para a
artéria pulmonar. Esta acção do ventrículo esquerdo sobre a parede do ventrículo
direito é tão importante que o coração pode funcionar não havendo potenciais de acção
no ventrículo direito. Só por contracção do ventrículo esquerdo a parede do ventrículo
direito é puxada fazendo expulsar sangue suficiente. Portanto uma isquémia no
ventrículo direito é muito menos grave do que uma isquémia no ventrículo esquerdo.
Fisiologia de Sistemas II – 2º Módulo
Isquémia é uma porção de músculo mal irrigada, e se ocorrer necrose, tem-se então um
enfarte. As paredes do ventrículo direito são relativamente delgadas uma vez que o
sangue que dele sai não precisa de estar a grandes pressões.
(ND: a professora disse que havia 3 tipos de contracção mas apenas falou em 2... ☺!!)
05:04
O ventrículo esquerdo tem a forma de um cilindro, de paredes muito espessas,
muito musculosas, e na extremidade desse cilindro tem um cone que forma o apex. No
ventrículo esquerdo também há um encurtamento da câmara, a válvula mitral aproximase da extremidade do coração, mas é sobretudo o movimento de contracção do cilindro
que vai fazer expulsar o sangue para a aorta. O sangue que está na aorta está a altas
pressões, e a oposição que o aparelho circulatório sistémico faz à passagem do sangue é
grande, de maneira que o ventrículo esquerdo tem de efectuar uma força maior para
expulsar o sangue do seu interior, por esta razão a parede é mais grossa.
O movimento de contracção do cilindro expulsa muito mais sangue do que o
movimento de contracção que faz a válvula mitral aproximar-se do apex, pois no
primeiro a variação do volume é proporcional à variação do quadrado do raio, enquanto
que no segundo a relação é linear entre o volume e o comprimento, a altura da câmara.
Válvulas cardíacas
As válvulas cardíacas são a tricúspide, a mitral, a aórtica e a pulmonar. As
válvulas auriculoventriculares – a tricúspide e a mitral – uma vez abertas permitem a
passagem do sangue da aurícula para o ventrículo. Quando estão fechadas impedem o
retrocesso do sangue dos ventrículos para as aurículas.
A válvula aórtica por exemplo durante a expulsão do sangue está aberta, mas
depois, durante a diástole está fechada impedindo o retorno do sangue das artérias para
os ventrículos.
As válvulas auriculoventriculares são constituídas por folhetos delicados, que
têm o mesmo peso específico do sangue de modo que qualquer pequeno fluxo de
sangue faz mover os folhetos das válvulas. Além disso, para fecharem bem, as válvulas
auriculoventriculares estão ligadas aos músculos papilares por meio das cordas
tendinosas que impedem que a válvula não abra para o interior da aurícula quando está
o ventrículo a contrair.
Ciclo cardíaco (do ventrículo esquerdo)
O ciclo cardíaco é a sucessão de acontecimentos que se passam desde que se
forma um potencial de acção no nodo sinosal até à contracção auricular seguinte.
10:19
Quando duas cavidades estão em comunicação a pressão do sangue em ambas as
cavidades é praticamente a mesma, há apenas um pequeno gradiente que é o que faz
mover o sangue em geral.
Onda P – Contracção auricular, a pressão na aurícula é ligeiramente superior à
do ventrículo, e o sangue passa da aurícula para o ventrículo. Cerca de 20% do sangue
que enche o ventrículo é passado na contracção auricular.
Onda QRS - o ventrículo começa a contrair, havendo uma tendência para o
refluxo de sangue para a aurícula, que ajuda a fechar a válvula auriculoventricular. As
Fisiologia de Sistemas II – 2º Módulo
cordas tendinosas fecham a
válvulas, e para além disso a
pressão no ventrículo já está igual
à pressão na aurícula, e a válvula
fecha mesmo. A válvula aórtica
está fechada.
Quando
a
válvula
auriculoventricular está fechada
dá-se início à sístole. Ao contrair o
ventrículo vai ter uma contracção
isovolumétrica, dado que as
válvulas auriculoventricular e a
semilunar estão fechadas, ou seja
não se altera o seu volume. O
ventrículo altera a sua forma, há
alteração do comprimento das
fibras musculares, mas como está
cheio
de
sangue
(que
é
incompressível), não há alteração
do volume.
O volume do ventrículo
mantém-se constante até a pressão
no interior do ventrículo ser
superior à pressão na aorta. Nesta
altura devido às diferenças de
pressão a válvula aórtica abre e
começa a expulsão do sangue para
a artéria aorta.
A velocidade de contracção
das fibras é máxima antes da
abertura da válvula aórtica.
O sangue é expulso em
duas fases. Tem uma fase de
expulsão rápida onde 2/3 do
volume sistólico de sangue é lançado, seguida por uma fase mais lenta. Durante a fase
de expulsão rápida o volume do ventrículo diminui muito rapidamente. Quanto à
aurícula durante a contracção isovolumétrica a válvula auriculoventricular sofre
protusão, o ventrículo comprime o sangue e este empurra os folhetos da válvula para
cima, aumentando um pouco a pressão nesta cavidade. Quando ocorre a expulsão do
sangue pelo ventrículo a pressão na aurícula desce, e fica inferior à das veias, o que
conduz à entrada de sangue nas aurículas, ao mesmo tempo que ocorre a expulsão de
sangue pelos ventrículos.
A expulsão é muito rápida até que começa a onda T do electrocardiograma –
ECG. Portanto começa a repolarização, inicia-se o relaxamento. Nesta altura a aorta está
cheia de sangue, e por essa razão está muito distendida, mas devido à sua parede
elástica, a aorta exerce pressão sobre o sangue, o que impede a entrada de mais sangue
vindo do ventrículo. Aqui a pressão na aorta começa a ficar superior à pressão no
ventrículo, o que torna a expulsão de sangue mais lenta.
Inicia-se então a diástole, com uma fase chamada a protodiástole, que é uma fase
muito curta, em que embora a válvula aórtica esteja aberta não há fluxo de sangue, pois
Fisiologia de Sistemas II – 2º Módulo
a pressão é igual na aorta e no ventrículo. Isto acontece até que devido a um
relaxamento do ventrículo a pressão diminui nesta cavidade, ficando inferior à pressão
na aorta, e a válvula aórtica fecha-se. Portanto a diástole começa não quando a válvula
aórtica fecha mas sim no início da protodiástole que é um pouco vaga, e que não se
sabe muito bem onde começa e onde acaba.
Seguidamente se olharmos para a curva de pressão da aorta vemos uma inflexão
da curva, um aumento na pressão a que se chama insizura.
Agora que estão as duas válvulas fechadas, a mitral e a aórtica, ocorre o
relaxamento isovolumétrico do ventrículo. O ventrículo relaxa sempre com o mesmo
volume pois no seu interior está a mesma quantidade de sangue. O ventrículo relaxa até
ter uma pressão inferior à da aurícula. A pressão no ventrículo ao ficar inferior à da
aurícula leva à abertura da válvula mitral, e ao seu enchimento.
O enchimento ocorre em 3 fases: enchimento rápido , enchimento lento ou
diastase e contracção auricular. Na primeira fase o gradiente de pressão é muito grande
e o ventrículo como que suga o sangue para o seu interior, na segunda já a pressão está
quase equilibrada, e daí ao sangue já não entrar tão depressa, e de seguida a contracção
auricular que finaliza o enchimento do ventrículo.
(...)
Ciclo cardíaco (do ventrículo direito)
No ventrículo direito as pressões são bastantes mais baixas (circulação de baixas
pressões), para além disso o máximo de pressão está na parte inicial da expulsão,
enquanto que no ventrículo esquerdo está a meio da expulsão. E existe uma ligeira
dessincronia entre o encerramento e a abertura das válvulas dos dois lados do coração,
por exemplo a aórtica e a pulmonar não fecham ao mesmo tempo.
Índice de inotropismo
23:55
A contractilidade do coração é muito importante, e é necessário saber como
quantificá-la e como melhorá-la, pois, por muitos doentes têm uma má contractilidade e
é necessário melhorar esse aspecto.
Portanto estabeleceram-se índices de inotropismo.
Há índices de inotropismo da fase de contracção isovolumétrica como a derivada
máxima da pressão. Se este valor for pequeno a contracção é lenta, e o inotropismo está
mau, se for grande a contracção é rápida e o inotropismo é muito bom. Também se
utiliza a derivada máxima da pressão sobre a pressão, que está normalizada.
Mas estes índices de inotropismo dependem da pré carga. A pré carga no
coração é a pressão ou o volume de sangue existente no final da diástole, mesmo antes
da contracção, e a pós carga é a pressão na aorta ou o impedimento que toda a
circulação faz à passagem do sangue. Estes dois índices dependem da pré carga, mas
são muitas vezes(??) utilizados, pois é difícil encontrar índices que não dependam da
pré carga.
Como outro índice temos o índice de inotropismo durante a expulsão, como a
velocidade inicial de ejecção que é a inclinação da curva de débito aórtico, ou da curva
de pressão aórtica.
A fracção de ejecção é o volume sistólico, ou seja o volume de sangue expulso
durante a sístole, sobre o volume no final da diástole. A velocidade de ejecção sistólica
normalizada média é a fracção de ejecção sobre o tempo de ejecção.
Fisiologia de Sistemas II – 2º Módulo
25:30
Todos estes índices dependem da carga, quer seja da pré carga ou da pós carga,
e por isso durante muito tempo procurou-se sistematicamente um índice que dependesse
muito pouco da carga.
Num diagrama de trabalho do coração (work diagram of the heart), temos a pressão
ventricular em função do volume do ventrículo. De A para D (A → D), temos a
contracção isovolumétrica, de D → S temos a expulsão, de S → V temos o relaxamento
isovolumétrico e de V → A enchimento.
Se aumentarmos a pré
carga, ou seja o volume no final do
enchimento, a curva com os pontos
A-D-S-V-A translada toda para a
direita ao longo da curva de
pressão/volume passiva.
Se aumentarmos a pós
carga, ou seja a pressão aórtica, a
válvula aórtica ( ponto D do
diagrama) não abre enquanto a
pressão no ventrículo não aumentar
suficientemente. Portanto a pressão
em vez de subir apenas até ao ponto
D vai subir até um novo ponto Dt , e
sucede uma fase transitória Dt → St
→ VÜ de reequilibrio onde o volume
sistólico diminui enquanto a pressão aumenta. De seguida o começo contracção
isovolumétrica translada para a direita ao longo da curva de pressão/volume passiva até
Fisiologia de Sistemas II – 2º Módulo
ao ponto A2. Nesta altura já o volume sistólico (SV)
normalizou (SV = SV2), apesar do aumento da pressão
sistólica (D2), que resulta num aumento relativamente grande
do volume residual (ESV2= ESV – End - systolic volume).
Começou então a determinar-se a chamada elastância
máxima ou rigidez da câmara máxima, neste ponto (??) (ND:
lamento imenso mas não consegui perceber a que ponto a
professora se refere, mas julgo ser o ponto M), que é o ponto
de elastância máxima, traçou-se então uma recta que vai para
a um ponto VD (??) que é o chamado volume morto, e
quando o inotropismo aumenta, temos uma recta que vai
parar ao mesmo ponto mas com uma inclinação muito maior,
e esse índice não depende nem da pré carga nem da pós
carga, e é portanto o melhor índice de inotropismo que existe.
Relaxamento isovolumétrico.
30:36
O relaxamento, a queda de tensão é exponencial no tempo. Se se impedir a
válvula mitral de abrir, ou seja, se não houver enchimento, o ventrículo continua a
relaxar do mesmo modo, até um valor de pressão assimptótico que é negativo. Isto
explica-se pelo o facto de existir uma força de recuo que faz relaxar o ventrículo para
uma pressão negativa.
Por altura do encerramento da válvula aórtica há um mínimo na derivada da
curva da pressão do ventrículo, e a seguir a este momento continua o relaxamento
isovolumétrico. Isto é um índice para o relaxamento isovolumétrico na fase inicial do
relaxamento. Para uma fase mais avançada do relaxamento utiliza-se o simétrico do
inverso do índice para o relaxamento isovolumétrico.
Evidentemente o relaxamento isovolumétrico convém ser muito rápido, para o
coração ter tempo de encher. Se o relaxamento isovolumétrico é mau, o coração não tem
tempo de encher a cavidade. Verificou-se que os problemas de relaxamento no coração
antecediam os problemas de contracção, ou seja antes de surgirem os problemas de
contracção, que são as que afligem mais as pessoas, aparecem deficiências de
relaxamento.
Além disso no músculo cardíaco como material visco elástico a reacção, o seu
comportamento depende da história da carga, ou seja depende do que se passou durante
muitos ciclos anteriores.
Enchimento diastólico
Num gráfico em que temos a
pressão diastólica em função do volume
diastólico, define-se como rigidez da
câmara a derivada dP/dV. A medição
deste valor só se efectua depois do
primeiro mínimo de pressão, ou seja
depois de ter passado o enchimento
rápido. Este valor para a derivada
depende das dimensões da câmara, e do
estado em que se encontra o miocárdio. A rigidez varia linearmente com a pressão, e
Fisiologia de Sistemas II – 2º Módulo
por isso à constante de linearidade que estabelece a recta dP/dV em da pressão chama-se
constante de rigidez da câmara. O inverso da rigidez, ou seja, dV/dP é a expansibilidade
da câmara.
Mas por vezes o coração funciona mal, ou porque está hipertrofiado, ou devido à
forma da câmara ser anormal, ou pelo miocárdio não estar em bom estado, e é preciso
determinar o módulo de elasticidade do miocárdio , e portanto tem que se ir calcular a
tensão na parede do ventrículo, e ver a deformação. Estabelecem-se então modelos do
ventrículo, e este modelo serve para
calcular o valor da tensão em função da
pressão intracavitária, e de seguida
traça-se as curvas de tensão/deformação
para o miocárdio, que dão o módulo de
elasticidade do miocárdio, que se chama
em medicina rigidez do miocárdio. O
inverso da rigidez do miocárdio chamase susceptibilidade do miocárdio.
Bibliografia:
Sangue e as suas propriedades visco elásticas – Folhas de Apoio Fisiologia II
Músculo Cardíaco – Tortora; Color Atlas of Physiology
O Coração como uma bomba – Color Atlas of Physiology
Índices de Inotropismo Physiology
Folhas de Apoio Fisiologia II; Color Atlas of
Peço desculpa pelos momentos de menor coerência linguística que estou certo
que irão encontrar, principalmente, segundo a minha opinião, na parte final sobre índice
de inotropismo, que de facto está pouco documentada, e que foi muito difícil por do
discurso da professora para um texto mais ou menos legível. De qualquer forma foi o
melhor que se conseguiu.
O desgravador:
Armando Granate