Fisiologia do Intestino Delgado
1. Introdução
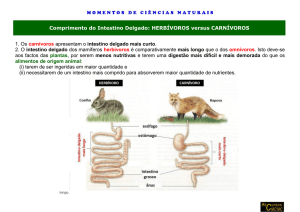
Nos mamíferos o intestino delgado é o sítio da digestão, designação genérica para a
transformação enzimática de algumas das substância ingeridas, e da absorção não só dos produtos
da digestão, mas também de elétrolitos e solutos orgânicos que, como as vitaminas, são absorvidos
sem transformação.
As enzimas luminais provêm do pâncreas exócrino. O intestino delgado secreta apenas uma
enzima, uma endopeptidase, que cliva o tripsinogênio, liberando a tripsina ativa. A digestão de
carboidratos, que começa pela alfa-amilase pancreática se completa por enzimas presas à membrana
apical ("bordo em escova") das células epiteliais. Quanto a proteínas, ocorre também digestao
intracelular de pequenos peptídios, com até quatro aminoácidos, por enzimas intracelulares, depois
da digestão luminal pelas proteases pancreáticas.
Os 5 m de extensão do intestino delgado não são homogêneos quanto à morfologia. O segmento
inicial de 25 cm, mais ou menos, é o duodeno. Segue-se o jejuno, com 40% do comprimento total. O
íleo é o segmento final, com 60% do comprimento. A válvula íleo-cecal separa o intestino delgado do
grosso.
O tempo completo do processamento, com absorção e transferência dos resíduos para o intestino
grosso, dura de 2 a 4 horas, conforme a quantidade e a natureza dos alimentos.
2. Motilidade
Movimento segmental, ou de segmentação rítmica, e peristalse por curtos segmentos são os
movimentos do intestino delgado no período digestivo. Os movimentos segmentais misturam o quimo
com as secreções pancreáticas, biliares e intestinais. Não são primariamente propulsivos. Os de
peristalse começam em regiões diversas, estimuladas pela presença do quimo, e estendem-se por
algumas dezenas de cm, antes da extinção. São movimentos que arrastam o quimo em direção ao
intestino grosso.
Ambos os tipos de movimento dependem da atividade dos marca-passos, que são células de tipo
miofibroblastos nas camadas musculares, e cuja atividade elétrica espontânea se espalha pelas
células musculares, acopladas por "gap junctions", e pode ser registrada como ondas lentas, também
denominadas ritmo elétrico basal. O padrão complexo dos movimentos é organizado pelo sistema
nervoso entérico e a atividade deste está modulada pelo sistema nervoso neurovegetativo, simpático
e parassimpático. O parassimpático colinérgico aumenta a motilidade e o simpático a inibe.
Hormônios produzidos por células diferenciadas na mucosa, como a secretina, a colecistocinina e
outros, modificam a motilidade. O estímulo para os movimentos é a presença do alimento que é
detectada pelos receptores de estiramento da parede e pelos quimioreceptores que analisam a
composição química do quimo.
A frequência das ondas lentas decresce ao longo do intestino, de 12/min no duodeno para 8/min no
íleo terminal. A diferença de frequência nos movimentos segmentais promove alguma propulsão do
alimento. A ampola duodenal se contrai com a mesma frequência do antro gástrico, o que impede o
refluxo de quimo duodenal para o estômago. A peristalse no íleo terminal abre o esfíncter íleo-cecal,
normalmente cerrado.
Nos períodos interdigestivos, a intervalos de 60 a 90 minutos, ondas peristálticas percorrem,
nascendo no estômago, o intestino delgado. São os complexos mioelétricos migratórios que arrastam
restos não digeridos para o intestino grosso.
Questões orientadoras do estudo
2.1. Discuta os movimentos do intestino delgado, dos esfíncteres e o controle deles pelo plexo
nervoso mioentérico, pelo sistema nervoso neurovegetativo e por hormônios.
3. Secreções no intestino delgado
Secreções pelo intestino
3.1- Estrutura da mucosa
3.2- Secreções intestinais: muco, solução de composição eletrolítica semelhante à do plasma,
enteropeptidase e células escamadas.
Secreções pancreáticas
4.1- Pâncreas exócrino e endócrino.
4.2- Estrutura do pâncreas exócrino: lóbulos, ácinos e dutos.
4.3- Água e eletrólitos: os principais e relação entre concentração e ritmo de secreção. Secreção
nos ácinos e nos dutos e, quanto a estes, diferenças entre os vários segmentos. Mecanismos
celulares da secreção e modulação desta por secretina e pela colecistocinina (CCK).
4.4- Enzimas. As enzimas são secretadas pelos ácinos das glândulas pancreáticas. A colecistocina
intestinal é o estímulo hormonal importante, que é potencializado pela secretina. A gastrina ocupa o
receptor da CCK, pois as moléculas são semelhantes, mas o efeito estimulatório é pouco. A inervação
parassimpática colinérgica estimula a secreção das proteínas.
4.4.1- Proteases. As proteases, por razões óbvias, são secretadas na forma de pró-enzimas,
inativas. A enteroquinase secretada pelo intestino delgado hidrolisa o tripsinogênio a tripsina que,
uma vez produzida, cliva mais tripsinogênio e os precursores da quimiotripsina, carboxipeptidase A e
B e elastase. Sob a catálise por estas enzimas as proteínas são convertidas a aminoácidos e pequenos
peptídios. Os aminoácidos e peptídos com até quatro aminoácidos são transportados por
carregadores da membrana apical. Os peptídios são hidrolisados por proteases em organelas
intracelulares. Pequenos peptídios inibidores das proteases são também produzidos pelas células
acinares. Estes se ligam com afinidade elevada às proteases respectivas e as mantém inativas.
Previnem ação proteolítica por ativação precoce das proteases ainda nos dutos.
4.4.2- -1,4-amilase. Em tudo semelhante à salivar.
4.4.3- Lipases. A triacilglicerol hidrolase hidrolisa as ligações éster 1 e 1' de triglicerídios,
produzindo monoglicerídios. São lipases que agem na interface lipídio-água e devem ser protegidas
dos ácidos biliares pela colipase, de secreção também pancreática. A emulsificação das gorduras é
fundamental para ação eficiente da lipase. A hidrolase de ésteres do colesterol produz colesterol e
ácidos graxos. Finalmente, a fosfolipase A2 hidrolisa a ligação éster dos fosfolipídios, produzindo
ácido graxo e lisofosfatídio.
4.4.4- RNAase e DNAase
4.5- Controle da secreção. Fases cefálica, gástrica e intestinal.
4.5.1- Ácinos: Gastrina, CCK ACh e subst. P estimulam a secreção via InsP 3 e DAG como
mensageiros. A insulina têm sobre os ácinos efeito trófico e estimulante da síntese e secreção dos
grânulos. A somatostina tem efeito inibitório.
4.5.2- Dutos: Secretina e VIP estimulam a secreção. O estímulo é amplificado pela ação
simultânea da CCK. A ACh não tem efeitos enquanto a somatostatina, o glucagon e o peptídio
pancreático têm efeito inibitório.
4.5.3- Secretina e CCK são hormônios produzidos pela mucosa do intestino delgado, nas regiões
do duodeno e jejuno superior. O primeiro é liberado quando o pH do quimo que entra no duodeno é
ácido, inferior a 5. A colecistocinina é liberada quando na luz do intestino delgado há gorduras ou
peptídios ou aminoácidos.
Fígado e secreção biliar.
A bile é uma solução a excretar que, pelas propriedades físico-quimicas dos sais biliares e de
fosfolipidios, tem importância na digestão de absorção de lipídios. As propriedades mencionadas são
a capacidade de emulsificação das gorduras e a formação de micelas que agregam os produtos da
digestão e lhes facilita a absorção.
5.2- A bile.
5.2.1- Os solutos orgânicos mais importantes são os sais biliares, o colesterol, a lecitina, e os
pigmentos biliares. Os eletrólitos são o Na+ , o K+, Cl- e bicarbonato, que a faz alcalina.
5.2.1- Secreção pelos hepatócitos e pelos dutos. Os hepatócitos estimula-os a CCK e os dutos
são estimulados pela secretina. Os sais biliares reabsorvidos no íleo são secretados de novo. Uns 10
% são excretados nas fezes e substituídos por síntese hepática. Coleréticos.
5.2.2- Concentração na vesícula biliar. Cálculos biliares. Esvaziamento vesical: parassimpático e
colecistocinina.
Questôes orientadoras do estudo.
Discuta a secreção pancreática, orgânica e inorgânica:
a. Componentes e ação no processo de digestão.
b. Mecanismos celulares da secreção.
c. Controle neural e endócrino dos processos de secreção.