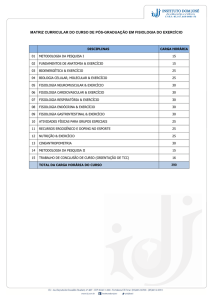
Fisiologia do Comportamento
Memória e Aprendizagem
Rodrigo Pavão
Laboratório de Neurociências e Comportamento
[email protected]
A memória pode ser definida como a capacidade de um organismo alterar seu
comportamento em decorrência de experiências prévias. Do ponto de vista fisiológico, essa
capacidade é resultado de modificações na circuitaria neural em função da interação do
indivíduo com o ambiente. Como já foi apresentado nos capítulos anteriores, o encéfalo
humano é composto por bilhões de neurônios, cada neurônio se projeta para centenas de
outros neurônios, e as regiões em que essas células se comunicam são denominadas
sinapses. A Figura 1 (esquerda) mostra um botão terminal do neurônio pré-sináptico “A”
sobrepondo-se ao corpo celular de um neurônio pós-sináptico; o primeiro é capaz de
modular a atividade do segundo. A formação de novas memórias envolve mudanças nas
sinapses existentes (como a do terminal “A“ com o neurônio pós-sináptico) ou a formação
de novas sinapses (como a do terminal axonal “B” sobre o terminal “A” – ver Figura 1,
direita); essas alterações levam à alteração e estabelecimento de circuitos neurais que
representam as memórias arquivadas.
Figura 1 – Sinapses axo-somática (esquerda) e axo-axônica (direita). A atividade do botão
axonal “B” pode modular a liberação de neurotransmissores do botão terminal “A” (modificado de
Carlson, 1998).
Esse
conhecimento
atual
resultou
do
trabalho
de
inúmeros
personagens;
destacaremos os principais em um breve histórico do estudo da memória. As primeiras
indagações de que se têm notícia na história da humanidade sobre a natureza da memória
foram formuladas pelos filósofos gregos e, posteriormente, reformuladas pelos pensadores
iluministas. No entanto, o estudo experimental da memória teve início no século XIX, com o
desenvolvimento do que hoje denominamos Psicologia Experimental. Hermann Ebbinghaus
(1880) realizou uma série de estudos (avaliando sua própria memória) envolvendo a
memorização de listas de sílabas sem sentido e a recordação das mesmas diferentes
períodos de tempo depois de sua apresentação. Suas principais observações são
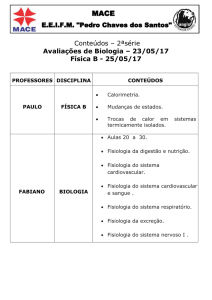
resumidas na Figura 2.
Fisiologia do Comportamento
Figura 2 – Resultados dos estudos de Ebbinghauss envolvendo memorização de listas
de sílabas. Foram descritas (1) a curva de recordação ao longo do tempo após a exposição às
sílabas (painel da esquerda) em que ocorre uma queda rápida da porcentagem de itens
recordados até cerca de duas horas; a partir de então, a porcentagem de itens recordados
permanece praticamente constante, (2) recordação em função da posição na série, em que a
recordação logo após a apresentação da seqüência de itens resulta numa maior lembrança
dos itens posicionados no início e no final da lista de itens, e (3) a curva de aprendizagem, em
que há necessidade de vários dias de treinamento para que a lembrança completa da lista
ocorra com menos treino (Ebbinghauss, 1885).
Müller e Pilzecker (1900), inspirados pelos trabalhos de Ebbinghaus, realizaram
testes que envolviam a apresentação de pares de sílabas que cuja lembrança deveria
ocorrer após um intervalo de tempo, oferecendo-se apenas um dos elementos de cada par;
uma lista distratora era oferecida para um segundo grupo de voluntários durante o intervalo
de tempo entre a lista apresentada e a lembrança da primeira lista. Os autores notaram que
os voluntários cuja atenção foi desviada do material estudado exibiram lembrança menor
do que o grupo de voluntários sem desvio da atenção; assim, enfatizaram a fragilidade das
memórias quando a atenção é desviada (Lechner e col.,1999). Esses autores descreveram
também o efeito de perseveração, em que testes posteriores eram afetados por testes
prévios. Os voluntários lembravam-se de pares de sílabas apresentadas em outro teste,
realizado semanas antes, resultando em erros, pois novas combinações deveriam ser
recordadas. A lembrança de combinações estudadas semanas antes evidencia que
atividade cerebral persevera após novo aprendizado (Lechner e col.,1999). Essa atividade
é resultante do processo de consolidação das memórias. No trabalho de Ebbinghauss
(1885) a estabilização da lembrança das sílabas várias horas após sua apresentação é
também resultado desse processo de consolidação.
Esses resultados sugeriram a existência de diferentes tipos de memória, incluindo (1)
uma memória que dura poucos segundos ou minutos, suscetível a interferências e não
consolidada, e (2) memória que dura dias ou semanas, robusta e resistente a
interferências, e consolidada.
Fisiologia do Comportamento
Em seu livro Principles of Psychology, William James (1890) denominou esses tipos
de memória como (1) memória consciente primária e (2) memória consciente secundária,
respectivamente. Além disso, esse autor mencionou também, em capítulos distintos,
“habilidades / hábitos”, envolvendo experiência não consciente. Curiosamente, essas idéias
foram ignoradas até a década de 1960.
No início do século XX, o fisiologista russo Ivan Pavlov e o psicólogo americano
Edward Thorndike, descreveram, respectivamente, o Condicionamento Clássico, em que
um animal aprende a associar dois estímulos (e.g., som de campainha a apresentação de
comida) pela sua apresentação contígua, e o Condicionamento Operante em que o animal
aprende a associar uma resposta motora a uma recompensa e uma segunda resposta a
uma punição.
Esses paradigmas estabelecidos por Pavlov e Thorndike influenciaram de modo
decisivo uma escola de pensamento denominada Behaviorismo, que almejava tornar a
psicologia uma “ciência objetiva”, baseada na observação de comportamentos,
desprezando conceitos como pensamento, imaginação, consciência ou mente, que eram
consideradas entidades subjetivas, não passíveis de abordagem experimental. A história
do Behaviorismo pode ser contada por seus conflitos com outras escolas de pensamento,
como sua disputa com a Psicologia Clínica, em que os behavioristas criticavam os
psicanalistas por uma suposta falta de controle experimental e de embasamento lógico e
científico. Os behavioristas defendiam que deve-se estudar as relações entre os estímulos
apresentados e as respostas geradas.
Uma escola alternativa de pensamento também baseada nos estudos iniciais de
Pavlov e Thorndike, denominada Cognitivismo, investigava não apenas como estímulos
geravam reações, mas também os processos não observáveis diretamente, mas que
intervêm entre o estímulo e a resposta. Essa escola de pensamento admite a flexibilidade
do comportamento animal, incluindo conceitos como representação, criação, inteligência,
memória e atenção, conceitos não admitidos pelo behaviorismo por não serem restritos à
relação entre estímulos e respostas.
O cognitivista Edward Tolman (1948) defendeu a idéia de que ratos arquivam em sua
memória uma representação espacial do ambiente, um mapa cognitivo, que permite a
orientação flexível no ambiente, inclusive encontrar atalhos nunca percorridos, mas
dedutíveis a partir do mapa cognitivo. Na esteira dos etologistas, Cooper e Zubek (1958)
realizaram estudos sobre as relações entre os comportamentos inatos e aprendidos (ver
Figura 5).
Fisiologia do Comportamento
Figura 5 – Ratos de uma mesma população inicial, que apresentavam diferentes níveis
de desempenho em uma tarefa envolvendo a aprendizagem de um labirinto foram cruzados
por gerações sucessivas, gerando uma linhagem “burra” e uma linhagem “brilhante” no
desempenho dessa tarefa. Em seguida, esses animais foram expostos a três condições
ambientais distintas, envolvendo (1) crescimento em ambiente empobrecido (gaiola com
animais isolados), (2) crescimento em ambiente padrão (caixa com um pequeno grupo de
animais), e (3) crescimento em ambiente enriquecido (caixa grande, com muitos animais,
brinquedos etc). Os animais das linhagens “burra” e “brilhante” crescidos em ambientes
empobrecido ou enriquecido exibiram desempenho equivalente. Diferentemente, animais
dessas duas linhagens crescidos em ambiente padrão exibiram marcada diferença de
desempenho; o desempenho dos animais da linhagem “brilhante” foi marcadamente melhor.
Em outras palavras, a diferença existe apenas condição padrão de criação. Os autores
concluíram que tanto fatores inatos como ambientais influenciam o comportamento
(modificado de Cooper e Zubek, 1958).
E os mecanismos fisiológicos subjacentes a esses processos?
Gold e colaboradores (1970) expuseram ratos a uma câmara clara de uma caixa
conectada, por uma porta tipo guilhotina, a uma câmara escura cujo assoalho é constituído
de barras metálicas eletrificáveis. Os ratos rapidamente entram na câmara escura; no
entanto, ao entrarem nessa câmara, levam um choque nas patas. Em tentativa posterior
(teste), realizada 24 horas depois, os animais inseridos na câmara clara não entram na
câmara escura (ver a barra vermelha da Figura 6). Animais de um grupo controle, que não
receberam choque nas patas no dia anterior, entram rapidamente na câmara escura (ver
barra verde da Figura 6). Em experimentos adicionais, a intervalos de tempo variáveis
depois do treinamento com choque nas patas, aplica-se uma corrente elétrica no sistema
nervoso dos animais, um choque eletro-convulsivo (ver Figura 6 - esquerda). Observa-se
que quanto menor o intervalo de tempo entre o choque nas patas e o choque eletro-
Fisiologia do Comportamento
convulsivo, maior é o prejuízo de memória aversiva sobre o ambiente escuro. Porém, a
medida que esse intervalo de tempo aumenta, menor é o efeito, como se o choque eletroconvulsivo perdesse sua efetividade para “apagar” a memória (ver Figura 6 – direita, barras
de cor laranja).
Figura 6 – Experimento ratos e choques. A organização temporal dos eventos e os resultados
estão apresentados à esquerda; os resultados obtidos estão à direita. Modificado de Xavier (2004) e
Gold (1970).
Shashoua (síntese publicada em 1985) prendeu um flutuador nas nadadeiras
peitorais de peixinhos dourados de modo que os animais ficam em posição invertida. Após
longo esforço de cerca de 3 horas, alguns peixes conseguem volta à posição normal,
apesar do flutuador (Figura 7, treino inicial representado pela curva verde). Se o flutuador
for removido e recolocado três dias depois, os animais que aprenderam a tarefa mais
rapidamente; i.e., os peixes conseguem voltar à posição normal em apenas 15 minutos, o
que indica que eles aprenderam e retiveram a solução desse desafio (curva azul) (para
detalhes sobre esses experimentos, ver Helene e Xavier, 2007a).
Figura 7 – Experimentos de Shashoua (1985) envolvendo aprendizagem em peixes dourados
(para detalhes ver o texto) (modificado de Shashoua, 1985, e Xavier, 2004).
Em outro teste, Shashoua (1985) injetou valina marcada com hidrogênio radioativo
(valina-H*) no ventrículo encefálico de animais que ficaram por 4h com o flutuador, ou
valina marcada com carbono radioativo (valina-C*) no ventrículo de animais que não foram
Fisiologia do Comportamento
treinados.
Os
encéfalos
dos
animais
dos
dois
grupos
foram
homogeneizados
conjuntamente e as proteínas foram separadas por peso molecular. A maioria das
proteínas presentes estava marcada tanto com valina-H* quando com valina-C*; porém,
algumas delas estavam mais marcadas com valina-H*, indicando que elas se originaram no
cérebro dos animais que aprenderam a tarefa; essas proteínas foram denominadas
ependiminas. Num terceiro teste, as ependiminas foram isoladas e injetadas em coelhos
para producão de anticorpos específicos contra as ependiminas. Então, os anticorpos
foram injetados no ventrículo encefálico de peixes que tinham acabado de aprender a
tarefa de nadar com o flutuador; no teste de memória realizado 3 dias depois, esses peixes
demoraram cerca de 3h para voltar à posição normal (Figura 7, curva vermelha). Ou seja,
esses animais comportam-se como se nunca tivessem sido submetidos ao treinamento.
Presentemente, as ependiminas são denominadas “moléculas de adesão celular” e estão
diretamente relacionadas com o fortalecimento e formação de sinapses.
Em conjunto, os resultados dos experimentos envolvendo choques eletro-convulsivos
e síntese de proteínas sugerem que há dois processos envolvidos na manutenção da
memória. Um deles, mais instável, é prejudicado pelo choque eletro-convulsivo, estando
relacionado ao padrão de atividade eletrofisiológica dos neurônios (freqüência de disparos,
por exemplo). O outro, associado com produção de proteínas, parece envolver alterações
estruturais nas sinapses gerando circuitos alterados no sistema nervoso. Num certo
sentido, esses dois tipos de processos parecem sobrepor-se aos descritos por James
(1890).
Resumindo, parece haver (1) uma Memória de Curta Duração, baseada na atividade
elétrica dos neurônios e, assim, um tanto suscetível a interferências e (2) uma Memória de
Longa Duração, representada por alterações estruturais dos neurônios, particularmente nas
sinapses com outros neurônios, robusta e resistente a interferências.
Onde esses traços de memória estão no sistema nervoso? Eles estariam localizados
em áreas discretas ou estariam espalhadas pelo sistema nervoso? Este tipo de
investigação ficou conhecida como “a busca pelo engrama (= traços de memória)”.
Franz Gall, fundador da Frenologia, no século XIX, defendia que quando uma pessoa
usa muito uma determinada região do cérebro, esta se hipertrofiaria (de modo similar a um
músculo) e, assim, deformava a caixa craniana, gerando um “calombo”; por outro lado, se a
região não fosse usada, ela atrofiaria, gerando uma “depressão”. Seguindo esta
concepção, Gall investigava o formato da caixa craniana de pessoas inteligentes,
engraçadas, egoístas, loucas etc. e propôs mapas sobre a localização das funções mentais
(publicados
em
revistas
especializadas
como
a
American
Phrenology
Journal,
http://www.phrenology.com/americanphrenology.html). Esta proposta gozou de grande
reputação durante o século XIX, mas foi totalmente abandonada posteriormente.
Fisiologia do Comportamento
Figura 8 – Franz Gall e um mapa frenológico.
Na década de 1920, Karl Lashley tentou localizar, em ratos, o engrama, ou seja, os
traços da memória, responsáveis pelo aprendizado do percurso para se orientar num
labirinto. Para testar essa idéia ele fez incisões no córtex antes ou depois do animal
aprender a tarefa; então, avaliava o desempenho do animal, tentando correlacionar a
extensão das lesões, com seu desempenho. O autor descreveu que os prejuízos de
aprendizagem e memória se correlacionam com a extensão da lesão, mas não com sua
localização (ver Bear, 2002; Helene e Xavier, 2007b). Esse resultado favoreceu a hipótese
de que os engramas estão espalhados pelo sistema nervoso e não dispostos em áreas
específicas como propunham, por exemplo, os frenologistas.
No entanto, as memórias parecem não estar totalmente espalhadas pelo encéfalo
como sugerem os resultados de Lashley. Pensa-se, atualmente, que os ratos solucionam a
tarefa valendo-se de diferentes modalidades sensoriais (visão, propriocepção, tato, olfato
etc.) e estratégias (orientação alocêntrica, egocêntrica etc.); quando as lesões são
pequenas, os ratos podem aprender e lembrar a solução usando as modalidades
preservadas; quando as lesões são extensas, o rato é incapaz de aprender ou lembrar do
labirinto.
Essa interpretação vai ao encontro de uma idéia interessante, denominada modelo de
“cell assembly” (de agrupamento de células) de Donald Hebb (1949). De acordo com essa
proposta, o engrama estaria representado em uma rede neural distribuída como
apresentada na Figura 9.
Figura 9 – Esquema representativo de redes neurais de Hebb. Os pontos pretos são os
neurônios e as linhas são as conexões. A rede tem uma organização inicial como representado em
(A); ao receber um estímulo, é ativada (B); esse estímulo pode ser apresentado repetidas vezes, ou
pode ter reverberado nessa rede, de modo que as conexões entre os neurônios são fortalecidas (C e
Fisiologia do Comportamento
D); então, um estímulo mais fraco ou mesmo incompleto, mas que mantenha algumas das
características do inicial (D) é capaz de ativar a rede fortalecida (E) (modificado de Bear, 2002, e de
Helene e Xavier, 2007b).
A perspectiva de que o engrama da memória esteja representado em circuitos
neurais que funcionam de maneira cooperativa e que diferentes regiões nervosas podem
contribuir para esse processo, estimulou os neurocientistas a se debruçassem sobre a
tentativa de localizar os sítios da memória em nosso encéfalo.
O estudo do caso do paciente H.M. muito contribuiu para o desenvolvimento dessa
área. Esse paciente sofria de epilepsia intratável (na ocasião); o foco epiléptico situava-se
no lobo temporal medial, bilateralmente. Então, na tentativa de ajudar o paciente, removeuse essa estrutura cirurgicamente; isso resultou na remoção dos 2/3 anteriores do
hipocampo e da amígdala, além de outras porções corticais (Scoville e Milner, 1957) (ver
Figura 10, esquerda). Como esperado, H.M. foi curado da epilepsia; porém, exibiu uma
perda de memória. A amnésia de H.M. era anterógrada (o paciente era incapaz de formar
novas memórias) e também retrógrada; porém, neste último caso a amnésia era
temporalmente graduada (ver Figura 10 direita). O prejuízo cognitivo de H.M. estava restrito
à aquisição de memórias de longa duração; suas capacidades perceptuais se mantiveram,
assim como seu QI, sua personalidade e a memória de curta duração; ou seja, estes
últimos, estavam todos preservados (Scoville e Milner, 1957).
Vale ressaltar aqui que mais uma vez foi mostrada a distinção entre memória de curta
duração (associada à atividade elétrica) e memória de longa duração (associada à
estrutura neural) proposta por James um século antes.
Figura 10 – O paciente H.M. teve parte do lobo temporal medial removido bilateralmente
(porção cortical, amígdala e hipocampo). A amnésia exibida por H.M. era anterógrada (ele era
incpaz de formar novas memórias) e retrógrada, neste último caso, temporalmente graduada
(lembranças da juventude e de eventos ocorridos até 2 anos antes da cirurgia foram preservados,
mas as lembranças são gradualmente prejudicadas até o momento da cirurgia (modificado de Bear,
2002 e Xavier, 2004).
Fisiologia do Comportamento
No entanto, H.M. conseguia adquirir e reter diversas informações. Por exemplo,
aprendeu a ler palavras invertidas, como se apresentadas por meio de um espelho, após a
cirurgia e também novas habilidades motoras e cognitivas (ver Helene e Xavier, 2007a,
para detalhes). Mesmo assim, se consultado sobre seu treinamento prévio nessas tarefas,
alegava nunca ter feito isso; mesmo assim, seu desempenho nessas tarefas treinadas era
proficiente.
Aparentemente, o hipocampo (e outras estruturas do lobo temporal medial) é
fundamental para a reverberação da atividade neural, que leva ao arquivamento de
informações sobre eventos experienciados. Essa reverberação seria essencial para o
arquivamento das informações sobre “o que” ocorreu, mas não sobre “como” desempenhar
uma tarefa percepto-motora. Na aquisição de uma habilidade, por exemplo, “como” andar
de bicicleta, a aquisição envolveria o treinamento repetitivo e envolveria regiões nervosas
intactas no paciente H.M. Assim, embora o paciente adquira essa habilidade não é capaz
de se recordar “que” já a praticou. Em outras palavras, a natureza da informação “saber
que” é diferente da natureza da informação sobre “saber como” (ver Helene e Xavier,
2007a, para detalhes). As memórias “saber como” são atualmente denominadas memórias
implícitas (o que faz bastante sentido, pois é muito difícil declarar como se anda de
bicicleta) e “saber que” são denominadas memórias explícitas.
Pacientes com Doença de Parkinson (que exibem disfunções em estruturas nervosas
denominadas gânglios da base) exibem um quadro oposto ao dos amnésicos (que, como
vimos, tem lesão no lobo temporal medial). Os pacientes com disfunções nos gânglios da
base exibem memória explícita preservada e prejuízo da memória implícita; esse prejuízo
pode envolver tanto aprendizagem e desempenho de respostas motoras, como perceptuais
(pacientes com Parkinson, por exemplo, exibem prejuízo na aprendizagem da habilidade
de leitura de palavras invertidas) (Knowlton e col., 1996; Perretta e col., 2005).
A memória de curta duração, preservada em amnésicos e parkinsonianos, usada
corriqueiramente para guardar, por exemplo, um número de telefone obtido numa lista (e
quando terminamos de teclá-lo já não somos mais capazes de declará-lo), está associada
ao funcionamento dos córtices frontal e parietal (Baddeley e Warrington, 1970). A memória
de curta duração é frequentemente denominada memória operacional.
Assim, memória vem sendo classificada em (1) memória de curta duração ou
memória operacional e (2) memória de longa duração. Por sua vez, a memória de longa
duração pode ser subdividida em (2a) memória explícita e (2b) memória implícita.
Fisiologia do Comportamento
Memória de Curta Duração (Memória Operacional)
Memória Explícita
Memória Implícita
ex. lembrar número da lista telefônica
ex. lembranças
ex. habilidades e hábitos
mantida em amnésicos
prejudicada em amnésicos,
mantida em amnésicos
mantida em parkinsonianos
especialmente para
prejudicada em parkinsonianos
prejudicada em pacientes com danos frontais
eventos recentes.
mantida em pacientes frontais
mantida em parkinsonianos
mantida em pacientes frontais
dura poucos segundos ou minutos
dura semanas ou anos
suscetível a interferências
é resistente a interferências
não consolidada (representada no padrão de atividade
consolidada (representada na estrutura das redes neurais; e.g.,
eletrofisiológica das redes neurais; e.g., freqüência
ependiminas) – memória implícita pelo treino repetitivo,
de disparos)
memória explícita pela reverberação (hipocampo)
É possível fazer uma comparação, que poderia ser interpretada como provocação,
entre a Frenologia do século XIX e o modelo de memória atual. Apesar de um pouco
agressiva, essa comparação é útil, pois estimula a interpretação de que os modelos são
aproximações
incompletas
que
nos
ajudam
entender
a
realidade
(ver
http://fisio.ib.usp.br/fisioteorica). De fato, algumas limitações do modelo de memória podem
ser apontadas, como não levar em conta a dramática plasticidade do sistema nervoso e a
clara inspiração nos equipamentos eletrônicos.
O primeiro aspecto pode ser evidenciado pelo estudo realizado por Leah Krubitzer
(1998) sobre a estrutura cortical de gambás. O córtex de um gambá adulto normal exibe
uma estrutura como a representada na Figura 11 (esquerda); se nos estágios fetais o
animal é submetido à remoção parcial do córtex, seu córtex adulto exibirá estrutura
bastante diferente da do gambá normal (Figura 11, direita). Isso mostra que estruturas
relacionadas com determinados tipos de processamento podem assumir funções distintas
(o animal lesado apresenta uma reorganização generalizada do sistema, não limitado a
prejuízo no processamento visual). Assim, uma interpretação alternativa a dos correlatos
anátomo-funcionais obtidos dos estudos envolvendo lesões é de que o sistema lesado
funcione de modo distinto, não limitado ao prejuízo naquela função.
Figura 11 – Organização cortical de gambás adultos. À esquerda o córtex normal e à direita o
córtex re-organizado após uma lesão fetal (modificado de Krubitzer, 1998).
O outro aspecto é o uso de analogia entre funcionamento de equipamentos
Fisiologia do Comportamento
eletrônicos e o funcionamento dos sistemas de memória. Isso, em princípio, não é um
problema; é simplesmente uma estratégia de estudo. A evolução dos modelos de memória
parecem corresponder a evolução dos equipamentos eletrônicos, e.g., (1) modelo de
conexões estímulo-resposta inspirada nas centrais telefônicas do início do século XX, (2)
os modelos sobre tipos de memória, estocagem e recuperação da informação inspirados
nos computadores dos anos 50-80 que também sofreram grande avanço e (3)
computadores atuais estão muito mais flexíveis, com grande interação entre hardware e
software. Um exemplo dessa questão, que pode ser apresentada como uma restrição ao
entendimento do sistema nervoso ao avanço tecnológico dos computadores, é evidente
nas palavras de Baddeley (1998): “por que não desenvolver computadores que são
baseados em processamento paralelo, e estudar as capacidades desse sistema para
aprender, lembrar e pensar?”. Talvez a analogia tenha assumido um outro papel que não
inspirar / facilitar a comunicação, tornando-se uma “camisa-de-força” ao restringir o
entendimento do fenômeno às características do sistema descrito na analogia.
Apesar dessas limitações, é inegável que esse modelo é útil e pode gerar
conseqüências práticas. Sabe-se que pacientes com a doença de Parkinson exibem sérias
dificuldades em suas atividades rotineiras, em decorrência do prejuízo da memória
implícita. Piemonte (2000), partindo do conhecimento de que pacientes com a doença de
Parkinson exibem prejuízo da memória implícita, mas memória explícita preservada, treinou
esses pacientes a realizarem suas atividades cotidianas como andar, levantar-se da cama
ou vestir uma camisa, com base em seqüências de instruções memorizadas explicitamente
sobre como executar cada uma dessas tarefas; isto é, cada uma dessas ações foi
subdividida em sub-componentes de movimentos, por exemplo, levantar a perna, virar o
tronco, empurrar a cama, que foram memorizados explicitamente pelos pacientes. Então,
essas memórias declarativas eram utilizadas no momento do desempenho da atividade. O
resultado foi um aumento na velocidade e precisão dos movimentos por parte dos
pacientes, com melhora substancial de sua qualidade de vida. Isso ressalta que modelos,
apesar de muitas vezes incompletos, podem gerar conseqüências práticas e também
contribuir para o avanço do conhecimento numa área.
Referências Bibliográficas
Baddeley AD (1998). Human Memory: theory and practice. p. 258-259.
Baddeley AD, Warrington EK (1970). Amnesia and the distinction between long-and short- term
memory. Journal of Verbal Learning and Verbal Behavior. v. 9, p. 176-189.
Bear MF, Connors BW & Paradiso MA (2002). Neurociências – Desvendando o Sistema Nervoso.
Porto Alegre 2ed, Artmed Editora.
Carlson NR (1998). Physiology of behavior. Boston : Allyn and Bacon. 6a ed.
Fisiologia do Comportamento
Cooper RM & Zubek JP (1958). Effects of enriched and restricted early environments on the learning
ability of bright and dull rats. Canadian Journal of Psychology, 12:159–164.
Ebbinghaus H (1985). Memory: A contribution to experimental psychology (Henry A. Ruger & Clara
E. Bussenius, Trad.). Original publicado em New York: Teachers College, Columbia
University. (Original em alemão “Über das Gedächtnis” publicado em 1885).
Gold PE, Farrell W & King RA (1971). Retrograde amnesia after localized brain shock in passiveavoidance learning. Physiology & Behavior, 7(5):709-712.
Hebb DO (1949). The organization of behavior: a neuropsychological theory. New York:J.Wiley Sons.
Helene AF, Xavier GF (2007a). Memória (e a elaboração da) percepção, imaginação, inconsciente e
consciência. Em Landeira-Fernandes J, Silva MTA (Eds.), Intersecções entre psicologia e
neurociências, MedBook Editora Científica Ltda.
Helene AF, Xavier GF (2007b). Interfaces entre a psicologia e as ciências biológicas: um olhar a
partir da biologia. Em Bastos AVB, Rocha NMD (Org.). Psicologia. Novas direções no diálogo
com outros campos de saber. Casa do Psicólogo.
James W (1890). The Principles of Psychology.
Knowlton BJ, Mangels JA, Squire LR (1996) A neostriatal habit learning system in humans. Science
273, 1399-402.
Krubitzer L (1998). Constructing a neocortex: influences on the pattern of organization in mammals.
In: Brain and Mind: evolutionary perspectives. Ed. Gazzaniga M e Altman J. (HFSP: Strabourg)
Lashley KS (1929). Brain mechanisms and intelligence. Chicago: University of Chicago.
Lechner HA, Squire LR, Byrne JH (1999). 100 Years of Consolidation - Remembering Müller and
Pilzecker. Learning & Memory 6(2), 77-87.
Pavlov, IP (1927). Conditioned reflexes: An investigation of the physiological activity of the cerebral
cortex (G. V. Anrep, Trad.).
Perretta JG, Pari G, Beninger RJ (2005). Effects of Parkinson disease on two putative nondeclarative
learning tasks: probabilistic classification and gambling. Cogn Behav Neurol 18, 185-92.
Piemonte MEP (1998). Uma nova proposta fisioterápica para pacientes com doença de Parkinson.
Dissertação de Mestrado. Instituto de Psicologia, Universidade de São Paulo. São Paulo.
Scoville WB, Milner B (1957). Loss of recent memory after bilateral hippocampal lesion. Journal do
Neurology, Neurosurgery and Psychiatry; 20:11-21.
Shashoua V. (1985). The role of extracellular proteins in learning and memory. American Scientist,
73, 364-370.
Thorndike, EL (1911). Animal intelligence.
Tolman EC (1948). Cognitive Maps in Rats and Men. The Psychological Review 55(4), 189-208.