Guia de Estudos:
Fisiologia Geral- GMV 107
Tecnologias de Informação e Comunicação na Educação
Professores: Dr. Luciano José Pereira
Dr. Márcio Gilberto Zangeronimo
Tutor: Edna Lopes
Lavras/MG
2011
1 | Página
Ficha catalográfica preparada pela Divisão de
Processos
Técnicos da Biblioteca Central da UFLA
Espaço a ser preenchido pela biblioteca
[A ser preenchido posteriormente]
Espaço a ser preenchido pelo CEAD
______________Digite o Título do Documento______________
Índice
UNIDADE 6 ............................................................................... 5
6.1 Sistema Endócrino ......................................................... 6
6.1.1 Hormônio ................................................................... 6
6.1.2 Principais mecanismos de ação hormonal ....................... 7
6.1.3 Hipotálamo – Hipófise ................................................. 11
6.1.3.1 Tabela Hormônios Hipotalâmicos ............................. 12
6.1.3.2 Hormônios da hipófise anterior (adeno-hipófise) .... 13
6.1.3.3 Hormônios da Hipófise posterior (Neuro-hipófise) .. 21
6.1.4 Glândula Pineal .......................................................... 27
6.1.5 Pâncreas Endócrino .................................................... 28
6.1.5.1 Tabela Regulação da secreção da insulina ............... 29
6.1.6 Paratireóide............................................................... 35
6.2 Bibliografia .................................................................. 38
4 | Página
______________Digite o Título do Documento______________
UNIDADE 6
OBJETIVO: Rever e Discutir os conceitos básicos sobre as teorias do
Sistema Endócrino
5 | Página
______________Digite o Título do Documento______________
6.1 Sistema Endócrino
6.1.1 Hormônio
Substância química secretada para os líquidos
corporais por uma célula ou grupo de células, exercendo
um efeito fisiológico de controle sobre outras células do
corpo. Sendo que, essas últimas precisam possuir
receptores específicos para receber a informação.
Existem três tipos de hormônios diferentes, dependendo
de sua composição química. São eles:
-Esteróides: derivados do colesterol (córtex adrenal,
gônadas e placenta).
-Derivados do aminoácido tirosina: tireóide e medula
adrenal
-Proteínas
ou
Peptídeos:
sintetizados
a
partir
de
aminoácidos
Os tipos de ação dos hormônios também podem variar,
de acordo com suas atuações nas células:
Autócrina: atua na própria célula
que
secretou o
hormônio.
Parácrina: atua em célula adjacente.
Endócrina: atua em célula localizada em tecido distante,
havendo passagem pela corrente sanguínea.
Para conseguir atuar nas diferentes células do
corpo, os hormônios precisam se ligar a receptores. Os
receptores hormonais são grandes proteínas, e cada
célula tem de 2.000 até 100.000 receptores.
A sensibilidade de uma célula a um hormônio pode ser
alterada pela diminuição ou aumento do número de
receptores ou da afinidade dos receptores ao hormônio.
Um tipo de regulação é aquela que reduz a resposta
hormonal,
chamada
down-regulation.
Isso
ocorre
6 | Página
______________Digite o Título do Documento______________
normalmente
após
a
permanência
de
uma
alta
quantidade de hormônio em contato com os receptores,
levando a uma resposta forte, inicialmente. Porém, após
um certo período essa situação gera diminuição da
densidade dos receptores ou de sua
causando
uma
diminuição
da
sensibilidade,
resposta.
Exemplo:
aumento de T3 diminui o número de receptores de TRH
na hipófise.
Outro tipo de regulação é aquela que aumenta a
resposta,
sendo
circunstância
chamada
gera
o
de
up-regulation.
aumento
da
densidade
Essa
dos
receptores e da sensibilidade, causando dessa vez, um
aumento
da
resposta.
Exemplos:
GH,
prolactina,
estrogênio que aumentam o número de seus próprios
receptores nos tecidos-alvo.
6.1.2 Principais mecanismos de ação hormonal
-Alteração da permeabilidade da membrana - Receptores
ionotrópicos: canal através do qual passam íons (como o
Na+ e K+). O canal é constituído por subunidades
proteicas organizadas em círculo.
-Ativação
de
enzimas
intracelulares
(receptores
metabotrópicos – Proteína G)
-Ativação
dos
genes
pela
fixação
a
receptores
intracelulares (hormônios esteróides)
Receptores acoplados a Proteína G - Mecanismos
de segundos mensageiros
As células respondem ao seu ambiente recebendo
informações de hormônios e outros sinais químicos
externos.
A
interação
destes
sinais
químicos
extracelulares, os chamados mensageiros primários, com
7 | Página
______________Digite o Título do Documento______________
os receptores na superfície pode levar à produção de
mensageiros secundários dentro da célula.
Os receptores metabotrópicos não possuem canais, mas
estão
ligados
a
proteínas
G
no
lado
interno
da
membrana celular que é responsável por passar a
mensagem ao interior da célula. Exemplo de receptor
metabotrópico: receptores adrenérgicos.
A proteína G possui três subunidades, α, β e γ, que
estão
localizadas
no
lado
citosólico
da
membrana
plasmática. A proteína G inativa apresenta a subunidade
α unida ao GDP (difosfato de guanosina). Quando ocorre
a união hormônio-receptor, ocorre a troca do GDP por
GTP, tornando a proteína ativada. A subunidade α
dissocia-se das subunidades β e γ. A subunidade α,
unida ao GTP, se desloca na membrana, desde o
receptor
até
uma
molécula
de
adenilciclase
ou
fosfolipase C (dependendo do tecido), ocorrendo sua
ativação e produção de segundos mensageiros: AMPc ou
DAG/IP3 respectivamente. Após sua ação, a subunidade
α cliva o GTP de volta a GDP retornando a sua posição
inicial. As subunidades β e γ também podem ter
atividade
intrínseca,
após
a
separação,
ativando
estruturas de membrana, como por exemplo modulando
a atividade de canal iônico de forma indireta.
Mecanismo de adenililciclase (enzima) e AMPc (2º
mensageiro)
O AMP cíclico é um mensageiro secundário, capaz
de
modular
diversos
processos
fisiológicos.
Este
mensageiro secundário é formado a partir do ATP em
uma reação catalisada pela adenililciclase, uma enzima
associada com a face interna da membrana plasmática.
8 | Página
______________Digite o Título do Documento______________
O hormônio, uma vez ligado a um receptor específico
localizado na membrana celular de uma célula-alvo,
provoca
a
ativação
de
uma
enzima
intracelular
(adenilciclase). Esta enzima converte parte do ATP
intracelular em AMP-cíclico. O AMP-cíclico, enquanto
presente no interior da célula, ativa a proteína quinase A
(dependente de AMP-cíclico), que fosforila uma série de
outras proteínas, ativando-as ou inibindo-as. Assim, o
AMP-cíclico, direta ou indiretamente, executa na célula
uma série de alterações fisiológicas como: ativação de
enzimas; alterações da permeabilidade da membrana
celular; modificações do grau de contração de músculo
liso; ativação de síntese protéica; aumento na secreção
celular. Exemplo: hormônio ADH (para receptor V2).
Mecanismo da fosfolipase C e DAG/IP3/Ca²+
A fosfolipase C é uma enzima hormônio-sensível
presente
na
membrana
plasmática,
que
atua
especificamente sobre o fosfotidil-inositol-4,5-difosfato
(fosfolipídio de membrana), catalisando sua hidrólise em
diacilglicerol
(DAG)
e
inositol-1,4,5-trifosfato
(IP3).
Esses dois compostos podem agir como segundos
mensageiros da ação hormonal. O diacilglicerol é um
ativador de proteínas-quinases C, fosforilando proteínas.
O
IP3
libera
endoplasmático.
íons
cálcio
Este
armazenados
mecanismo
tem
no
um
retículo
papel
importante na contração de músculo liso e de outros
processos fisiológicos. O TRH, ACTH, LH, angiotensina II,
serotonina e vasopressina, são alguns dos hormônios
que atuam por este mecanismo.
9 | Página
______________Digite o Título do Documento______________
Mecanismo dos hormônios esteróides (derivados do
colesterol)
1) Atravessam a membrana
2) Interagem com os receptores no citoplasma (ou até
mesmo dentro da carioteca)
3) Migram para dentro do núcleo
4) Modificam DNA (promovem aumento ou inibição da
transcrição de genes).
Ex: testosterona penetra na célula e se liga ao receptor,
então o complexo hormônio-receptor passa através da
carioteca e se liga em regiões específicas do DNA
promovendo
estímulo
da
transcrição
de
genes
específicos (como por exemplo, estímulo da síntese
protéica).
10 | P á g i n a
______________Digite o Título do Documento______________
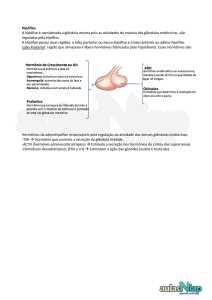
6.1.3 Hipotálamo – Hipófise
A glândula hipófise está localizada na sela túrcica e
consiste em dois lobos principais: anterior e posterior. O
lobo anterior é também chamado de adeno-hipófise,
sendo
constituído
posterior,
de
denominado
tecido
glandular.
neuro-hipófise
Já
é
o
lobo
constituído
exclusivamente por tecido neural. Funcionalmente o
hipotálamo controla a glândula hipófise por mecanismos
neurais e hormonais.
A
hipófise
anterior
secreta
seis
hormônios
peptídicos: hormônio tiroestimulante (TSH), hormônio
folículo estimulante (FSH), hormônio luteinizante (LH),
hormônio do crescimento (GH), prolactina (PRL) e
hormônio adrenocorticotropico (ACTH). Cada hormônio
do lobo anterior é sintetizado e secretado por tipo celular
distinto, exceto para o FSH e LH, que são secretados
pelo mesmo tipo celular. Já a hipófise posterior secreta
dois hormônios, o ADH e a ocitocina.
A natureza da relação entre o hipotálamo e a
hipófise anterior é tanto neural,
quanto endócrina
(contrastando com a do lobo posterior, exclusivamente
neural).
O
hipotálamo
e
a
hipófise
anterior
são
conectados diretamente, pelos vasos sanguíneos portahipotalamo-hipofisários, responsáveis pela maior parte
do suprimento sanguíneo para o lobo anterior da
hipófise.
O
suprimento
sanguíneo
para
a
hipófise
anterior é em sua maior parte de sangue venoso, vindo
do hipotálamo, levado pelos vasos porta longos e curtos.
Os hormônios liberadores de hormônios e os hormônios
inibidores da liberação são sintetizados nos corpos
celulares
de
neurônios
hipotalâmicos
e
estes
são
11 | P á g i n a
______________Digite o Título do Documento______________
transportados ao longo dos axônios desses neurônios até
a eminência mediana do hipotálamo. Quando esses
neurônios são estimulados, os hormônios são secretados
para o tecido hipotalâmico circundante, entrando no
plexo capilar vizinho. O sangue desses capilares drena
para os vasos porta hipofisários e é levado diretamente
para o lobo anterior da hipófise, onde os hormônios
hipotalâmicos agem sobre as células do lobo anterior,
estimulando ou inibindo a liberação dos hormônios da
hipófise anterior.
Os hormônios hipotalâmicos são levados para a
hipófise anterior diretamente, e em altas concentrações,
não
aparecendo
na
circulação
sistêmica
em
concentrações elevadas. Por conseguinte, as células da
hipófise anterior são as únicas células do corpo que
recebem
concentrações
elevadas
dos
hormônios
hipotalâmicos. Abaixo estão descritos os hormônios
hipotalâmicos com suas respectivas ações:
6.1.3.1 Tabela Hormônios Hipotalâmicos
Hormônios
Ação
Hipotalâmicos
TRH
Libera TSH e
PRL
CRH
Libera ACTH
GHRH
Libera GH
GHIH
Inibe GH
(somatostatina)
GnRH
Libera LH e
FSH
PIH (dopamina)
Inibe PRL
12 | P á g i n a
______________Digite o Título do Documento______________
6.1.3.2 Hormônios da hipófise anterior (adenohipófise)
Cada um dos hormônios da hipófise anterior é um
peptídeo. O
hormônio
é
armazenado
em grânulos
secretores revestidos por membrana para liberação. A
secreção da hipófise anterior é modulada por hormônio
hipotalâmico de liberação ou por hormônio inibidor de
liberação.
Hormônio do crescimento
É secretado durante toda a vida. Exerce efeitos
profundos sobre o metabolismo das proteínas, dos
carboidratos e dos lipídios. A sua síntese é estimulada
pelo GHRH, seu hormônio hipotalâmico de liberação.
O GH é liberado segundo padrão pulsátil, com surtos de
secreção ocorrendo, aproximadamente a cada duas
horas. Sua intensidade secretória não é constante
durante toda a vida. Aumenta continuamente até a
primeira infância. Na puberdade ocorre aumento abrupto
provocado no sexo feminino pelo estrogênio e no sexo
masculino pela testosterona (surto de crescimento da
puberdade).
Após
a
puberdade,
a
intensidade
da
secreção do hormônio declina até valor estável. Por fim,
na
idade
senil,
as
intensidades
secretórias
e
a
pulsabilidade declinam até seus níveis mais baixos.
A secreção do hormônio do crescimento, pela
hipófise
anterior
hipotalâmicas,
uma
é
controlada
estimuladora
por
duas
(GHRH)
e
vias
outra
inibitória (somatostatina).
13 | P á g i n a
______________Digite o Título do Documento______________
O GHRH atua diretamente sobre os somatotrofos
da hipófise anterior estimulando tanto a síntese como a
secreção do GH. A somatostatina também é secretada
pelo hipotálamo e atua sobre os somatotrofos para inibir
a secreção de hormônios do crescimento.
A
secreção do
GH
é
regulada
por
feedback
negativo. O GHRH inibe sua própria secreção pelo
hipotálamo por meio de uma alça ultra-curta. As
somatomedinas, substâncias produzidas pela ação do GH
sobre os tecidos-alvo (especialmente fígado), inibem a
secreção do hormônio do crescimento pela hipófise
anterior. Tanto o GH como as somatomedinas estimulam
a secreção de somatostatina pelo hipotálamo. O efeito
global
dessa
somatostatina
estimulação
inibe
a
é
inibitório,
secreção
do
por
que
hormônio
a
do
crescimento pela hipófise anterior.
O GH exerce múltiplas ações metabólicas sobre o
fígado, o músculo, o tecido adiposo e o osso, bem como
ações promotoras do crescimento em virtualmente todos
os órgãos. Algumas das ações do GH resultam de efeito
direto
sobre
os
tecidos-alvo,
como
o
músculo
esquelético, o fígado e o tecido adiposo. Outras ações
são mediadas indiretamente por meio da produção das
somatomedinas (IGF) pelo fígado. As somatomedinas
atuam
nos
tecidos-alvo
por
meio
de
receptores
semelhantes aos receptores de insulina, recebendo por
este motivo a denominação de IGF que significa insulinlike growth factor.
O GH promove resistência à insulina, reduzindo a
captação e utilização de glicose causando aumento da
concentração
sanguínea
de
glicose
(Efeito
diabetogênico). Este hormônio promove ainda aumento
14 | P á g i n a
______________Digite o Título do Documento______________
da
síntese
protéica
e
do
crescimento
dos
órgãos
(aumento da captação de aminoácidos e síntese de DNA,
de RNA e de proteínas). Esses efeitos explicam as ações
promotoras de crescimento, aumento da massa corporal
magra e maior tamanho dos órgãos. O aumento do
crescimento
linear
é
principalmente
somatomedinas,
alterando
cartilagens.
ossos
Nos
em
o
mediado
pelas
metabolismo
das
crescimento,
as
placas
epifisárias se alargam e mais osso é depositado nas
extremidades dos ossos longos.
TSH (Hormônio Tireoestimulante)
A síntese e a secreção dos hormônios tireóideos
são
controladas
pelo
eixo
hipotálamo-hipofise.
O
hormônio liberador de tireotropina (TRH) é secretado
pelo hipotálamo e atua sobre os tirotrofos da hipófise
anterior
para
promover
a
secreção
do
hormônio
Tireoestimulante (TSH) (e prolactina). O TSH então atua
sobre a glândula tireóide para estimular a síntese e
secreção dos hormônios da tireóide (T3 e T4).
O TRH atua sobre os tirotrofos da hipófise anterior
para estimular tanto a transcrição do gene do TSH, como
a secreção de TSH. A função do TSH é regular o
crescimento e o metabolismo da glândula tireóide.
A secreção de TSH é regulada por dois fatores
recíprocos: (1) o TRH do hipotálamo estimula a secreção
de TSH e (2) os hormônios tireóideos inibem a secreção
de TSH por regulação para baixo do receptor para TRH,
nos tirotrofos, o que diminui sua sensibilidade (down
regulation) à estimulação pelo TRH.
O TSH exerce dois tipos de ações sobre a glândula
tireóide: (1) aumenta a síntese e a secreção dos
15 | P á g i n a
______________Digite o Título do Documento______________
hormônios tireóideos por estimular cada etapa da via
Biosintética e (2) O TSH exerce ação trófica sobre a
glândula tireóide. Quando os níveis de TSH estão
aumentados por período prolongado, ocorre hipertrofia e
hiperplasia das células foliculares da tireóide e aumento
do fluxo sanguíneo para a glândula tireóide.
Síntese de Hormônios Tireoidianos
Os hormônios tireoidianos necessitam da presença
de iodo para serem produzidos. O Iodo entra no tireócito
por mecanismo de co-tranporte sódio-iodo (sequestro do
iodo). A célula tiroidiana sintetiza uma proteína chamada
tireoglobulina, que contém os radicais tirosina, que são
unidos por oxidação, ao iodo da dieta, por meio de uma
enzima chamada peroxidase.
A formação do T3 (triiodotironina) ocorre pela
união de uma monoiodotirosina (um iodo e uma tirosina)
com uma Diiodotirosina (dois iodos e uma tirosina), e a
formação
do
T4
(tiroxina)
pela
adesão
de
duas
Diiodotirosinas, sendo esta última a maior parcela da
produção hormonal, mas também a menos ativa. Após a
produção, T3 e T4 são armazenados no colóide.
O TSH, além de estimular todas as etapas da
produção,
também
estimula
a
liberação
desses
hormônios na corrente sanguínea. Para que isso ocorra,
os
constituintes
nos
lisossomas
são
fragmentados,
liberando-os das tereoglobulinas. Dessa forma, chegam
aos seus órgãos-alvo, ligam aos seus receptores e
exercem suas diversas funções metabólicas. Como por
exemplo, aceleram o metabolismo por meio da indução
da síntese de Bomba Na/K, que aumenta o consumo de
ATP (maior gasto energético). Também, atuam na
16 | P á g i n a
______________Digite o Título do Documento______________
termogênese, aumentando consumo de O2 e a contração
muscular, produzindo calor. Portanto o efeito global
desses hormônios é catabólico.
Prolactina
É o principal hormônio responsável pela produção
de leite, participando também do desenvolvimento das
mamas. Nas fêmeas não gestantes e não lactantes, bem
como nos machos, os níveis de prolactina são baixos,
contudo durante a gestação e a lactação os níveis
sanguíneos de prolactina aumentam. A prolactina é
produzida pelos lactotrofos da hipófise anterior. O
número dos lactotrofos aumenta durante a gestação e a
lactação. O TRH é o estimulador da secreção de
prolactina,
aumentando
a
transcrição
do
gene
da
prolactina. A dopamina é o inibidor da secreção de
prolactina
e
reduz
a
transcrição
desse
gene.
Em
indivíduos que não estejam lactando, a secreção de
prolactina é tonicamente inibida pela dopamina no
hipotálamo.
A gestação e a amamentação são os estímulos
mais importantes para a secreção de prolactina. Por
exemplo, durante a amamentação os níveis séricos de
prolactina podem aumentar em até 10 vezes o valor
basal.
Ações da Prolactina
A
prolactina
estrogênio
e
em
a
função
sustentadora
progesterona,
com
estimula
o
o
desenvolvimento mamário, promove a secreção de leite
pelas mamas durante a lactação e suprime a ovulação.
17 | P á g i n a
______________Digite o Título do Documento______________
Desenvolvimento
mamário:
Na
puberdade,
a
prolactina, junto com o estrogênio e a progesterona,
estimula a proliferação dos ductos mamários. Durante a
gestação, a prolactina e os dois hormônios estimulam o
crescimento e o desenvolvimento dos alvéolos mamários
que vão produzir leite após a parturição.
Lactogênese: Produção de leite em resposta ao ato
de mamar. (Não precisa haver gestação). O mecanismo
de ação da prolactina na mama depende de sua fixação
a receptor na membrana celular e por meio de segundo
mensageiro
transcrição
ainda
dos
não
genes
identificado
para
as
e
promove
enzimas
das
a
vias
biosintéticas para a lactose, a caseína e os lipídios.
Embora durante a gestação os níveis de prolactina sejam
muito altos, não ocorre lactação por que os níveis
elevados de estrogênio e de progesterona bloqueiam a
ação da prolactina sobre a mama e também devido ao
fato
de
que
os
níveis
de
ocitocina
–
hormônio
responsável pela ejeção do leite estão baixos. Na
parturição, os níveis de estrogênio e progesterona caem
acentuadamente, removendo suas ações inibitórias; a
prolactina
pode
então
estimular
a
lactogênese,
ocorrendo a lactação. Aliado a isso, o lactente ao sugar o
teto, estimula a liberação de ocitocina pela neurohipófise causando a ejeção do leite.
Inibição da ovulação: Nas fêmeas, a prolactina
inibe a ovulação, por inibir a síntese e a liberação do
hormônio liberador de gonadotropinas (GnRH). A inibição
da secreção de GnRH e secundariamente a inibição da
ovulação explica a fertilidade reduzida durante o período
de lactação.
18 | P á g i n a
______________Digite o Título do Documento______________
ACTH
A síntese e a secreção dos hormônios esteróides,
pelo córtex supra-renal, dependem de estimulação pelo
ACTH. A zona fascicular, secretora dos glicocorticóides,
está
sob
controle
exclusivo
do
eixo
hipotálamo-
hipofisário. O hormônio hipotalâmico é o hormônio
liberador de corticotropina (CRH) e o hormônio da
hipófise anterior é o ACTH. A zona glomerular secretora
dos mineralocorticóides, também depende do ACTH para
a etapa inicial da biosíntese dos esteróides, mas depois
disso, é controlada separadamente por meio do sistema
renina-angiotensina-aldosterona.
O CRH é secretado pelo hipotálamo e atua sobre os
corticotrofos da hipófise anterior, para produzir ACTH.
Por sua vez, o ACTH atua sobre as células do córtex
supra-renal, para estimular a síntese e a secreção dos
hormônios adrenocorticais.
Como outros hormônios hipotalâmicos que atuam
sobre a hipófise anterior, o CRH atinge a hipófise por
meio do sangue porta hipotálamo-hipofisário. No lobo
anterior, ele atua sobre os corticotrofos, para promover
a secreção de ACTH para a corrente sanguínea. O ACTH,
o hormônio da hipófise anterior exerce efeitos diversos
sobre o córtex supra-renal. O ACTH apresenta padrão
secretório pulsátil e diurno, o que determina um padrão
paralelo,
para
a
secreção
de
cortisol
(principal
glicocorticóide).
O controle por feedback negativo do eixo CRHACTH
é
exercido
pelo
cortisol,
tanto
no
nível
hipotalâmico, como no hipofisário anterior. O cortisol
19 | P á g i n a
______________Digite o Título do Documento______________
inibe diretamente a secreção de CRH pelo hipotálamo; e
indiretamente inibe a secreção de CRH por meio de seus
efeitos
sobre
neurônios
hipocâmpicos,
que
fazem
sinapses no hipotálamo inibindo a ação do CRH sobre a
hipófise anterior, resultando na inibição da secreção de
ACTH. Desse modo, a deficiência crônica de cortisol leva
à estimulação do eixo CRH-ACTH e o excesso crônico de
cortisol leva à inibição do eixo CRH-ACTH.
Naturalmente, o ACTH permanece essencial para a
secreção de aldosterona, contudo, a regulação primária
da secreção de aldosterona é controlada pelas variações
do volume do LEC, por meio do sistema reninaangiotensina-aldosterona e pelos níveis séricos de K e
não pelo ACTH.
O cortisol tem importante papel na regulação do
ciclo circadiano (maiores níveis durante o dia e menores
durante a noite), nas reações inflamatórias (potente
agente antiinflamatório) e nas reações de estresse
(aumentando a disponibilidade de energia, aminoácidos,
etc. na corrente sanguínea e favorecendo também a
liberação das catecolaminas).
FSH e LH (Hormônio Folículo-estimulante e Hormônio
Luteinizante)
O evento primário na puberdade é o início da
secreção pulsátil de GnRH (Hormônio liberador de
gonadotrofinas). Esse padrão de secreção desencadeia
paralelamente secreção também pulsátil de FSH e de LH
pelo lobo anterior da hipófise.
A secreção pulsátil de FSH e LH estimula a
secreção
de
hormônios
esteróides
gonádicos,
testosterona e estradiol. Os níveis circulantes elevados
20 | P á g i n a
______________Digite o Título do Documento______________
dos
hormônios
responsáveis
pelo
esteróides
sexuais
aparecimento
das
são
então
características
sexuais secundárias.
As duas funções dos testículos, espermatogênese e
secreção de testosterona são controladas pelo eixo
hipotálamo-hipofisário. O hormônio hipotalâmico é o
hormônio liberador de gonadotropina (GnRH) e os
hormônios da hipófise anterior são o hormônio folículoestimulante (FSH) e o hormônio luteinizante (LH). O
GnRH é secretado para o sangue porta-hipotálamohipofisário e levado, em alta concentração, diretamente
para o lobo anterior da hipófise.
6.1.3.3 Hormônios da Hipófise posterior (Neurohipófise)
As conexões entre o hipotálamo e o lobo posterior
da hipófise são neurais. Na verdade, a hipófise posterior
é uma coleção de axônios, cujos corpos celulares ficam
situados no hipotálamo. Desse modo, os hormônios
secretados pelo lobo posterior (ADH e ocitocina) são
peptídeos liberados por neurônios ou neuropeptídeos.
Uma vez sintetizados nos corpos celulares, os
neuropeptídeos são transportados ao longo dos axônios,
em
vesículas
neurosecretórias,
e
armazenados
nas
terminações bulbosas, na hipófise posterior. Quando o
corpo celular é estimulado, as vesículas neurosecretórias
são liberadas das terminações nervosas por exocitose; o
hormônio entra nos capilares fenestrados próximos. O
sangue venoso da hipófise posterior, chega à circulação
sistêmica, que os leva aos tecidos-alvo.
21 | P á g i n a
______________Digite o Título do Documento______________
Síntese e secreção de ADH e ocitocina
Os neurônios secretores de ADH têm seus corpos
celulares, principalmente, nos núcleos supra-ópticos do
hipotálamo. Os neurônios secretores de ocitocina têm
seus corpos celulares nos núcleos paraventriculares do
hipotálamo.
Hormônio Anti-diurético
O ADH ou vasopressina é o principal hormônio
relacionado à regulação da osmolaridade dos líquidos
corporais. O ADH é secretado pela hipófise posterior em
resposta ao aumento da osmolaridade sérica. O ADH,
então, atua sobre as células principais do túbulo distal
final e ductos coletores, para aumentar a reabsorção de
água e, dessa forma, diminuindo a osmolaridade dos
líquidos corporais até seu valor normal.
Adicionalmente, o ADH atua nos vasos sanguíneos
exercendo
efeito
de
vasocontrição,
aumentando
a
pressão arterial.
Regulação da secreção de ADH
O
aumento
da
osmolaridade
sanguínea
é
o
estímulo fisiológico mais importante para a regulação da
secreção de ADH. Por exemplo, quando um indivíduo
está em privação de água, a osmolaridade sérica
aumenta, o que é detectado pelos osmorreceptores do
hipotálamo anterior. São gerados potenciais de ação nos
corpos celulares dos neurônios secretores de ADH,
situados próximos, que se propagam pelo axônio,
promovendo a secreção do ADH pelas terminações
nervosas, na hipófise posterior. De modo inverso, a
22 | P á g i n a
______________Digite o Título do Documento______________
diminuição
da
osmolaridade
sérica
sinaliza
os
osmorrecptores para inibir a secreção de ADH.
A
hipovolemia
(por
exemplo,
decorrente
de
hemorragia) também é estímulo potente para a secreção
de ADH. A redução do volume do líquido estracelular em
10% ou mais provoca baixa da pressão arterial que é
detectada pelos baroceptores, no átrio esquerdo, na
artéria carótida e no arco aórtico. Essa informação sobre
a pressão arterial é transmitida por meio dos nervos
vagos, para o hipotálamo, que promove a secreção de
ADH. O ADH então estimula a reabsorção de água, nos
ductos coletores, na tentativa de normalizar o volume do
LEC.
Etanol
e
antagonistas
alfa-adrenérgicos
e
o
peptídeo natriurético atrial inibem a secreção de ADH.
Ações do ADH
O ADH tem duas ações, uma no rim e a outra no
músculo liso vascular. Essas ações são mediadas por
diferentes receptores
- Aumento da permeabilidade à água: A principal ação
do ADH é a de aumentar a permeabilidade à água das
células
principais
do
túbulo
distal
final
e
ductos
coletores. A permeabilidade aumentada à água, pelas
células principais permite que a água seja reabsorvida
pelos ductos coletores, tornando a urina concentrada ou
hiperosmótica. O ADH aumenta a reabsorção de água do
filtrado glomerular através da inserção de canais de
água (aquaporinas) na membrana luminal das células do
ducto coletor. A aquaporina 2 (AQP2) é o canal de água
regulado pelo ADH nos ductos coletores renais (receptor
V2 – AMPc).
23 | P á g i n a
______________Digite o Título do Documento______________
-
Contração
do
músculo
liso
vascular:
efeito
vasoconstritor (receptor V1 – ligado a DAG/IP3).
Ocitocina
As ações fisiológicas da ocitocina são exercidas
principalmente sobre a musculatura uterina e sobre a
musculatura lisa que reveste os alvéolos da mama.
Através
desses
mecanismos,
a
ocitocina
participa
respectivamente do mecanismo do parto e da ejeção do
leite durante a lactação.
Ação sobre o útero
A elevação dos níveis séricos de ocitocina leva a
um aumento da freqüência e duração dos trens de
potenciais de ação na musculatura uterina, ações que
(1)
iniciam
contração
na
musculatura
uterina
previamente inativa e (2) aumentam a freqüência, força
e duração das contrações em músculos já ativos. A
administração de estrógenos a animais imaturos traz o
potencial de membrana das células uterinas a níveis
mais próximos do seu limiar de disparo. Nesse sentido, o
potencial
de
membrana
dessas
células
declina
gradualmente da metade até o término da gestação,
quando
os
níveis
de
estrógenos
estão
bastantes
elevados. Essa queda do potencial de membrana facilita
a ação da ocitocina sobre o útero que, por aumentar a
excitabilidade do miométrio e por facilitar a condução
dos potenciais de ação, ativa as células uterinas que
estavam quiescentes, aumentando assim o número de
células participantes e a força de cada contração.
Acredita-se que a ocitocina exerça esses efeitos por meio
de um aumento generalizado da permeabilidade iônica
24 | P á g i n a
______________Digite o Título do Documento______________
da membrana celular. A ocitocina, entre outras ações, é
responsável
(1)
pelo
aumento
do
número
de
transportadores de sódio no sarcolema, durante a fase
de potencial em espícula, e (2) pela mobilização de
cálcio dos estoques intra e extracelulares.
Ação da ocitocina no parto
A ação da ocitocina no parto está bem definida,
tendo em vista as seguintes evidências clínicas: (1)
aumento da secreção de ocitocina durante o parto; (2)
correlação positiva entre a concentração plasmática de
ocitocina e o prosseguimento do trabalho de parto; (3) o
trabalho
de
parto
hipofisectomizadas,
com
é
difícil
secção
em
cirúrgica
pacientes
da
haste
hipofisária ou com bloqueio da liberação de ocitocina.
Em algumas espécies, como rato e coelho, a ocitocina é
o fator desencadeante do parto, enquanto nas mulheres,
a ocitocina apenas contribui de forma importante para o
desenvolvimento do trabalho de parto e expulsão fetal.
Dessa forma, embora na espécie humana, a ocitocina
não
seja
indispensável
para
o
parto,
sua
ação
estimulante é altamente vantajosa.
A secreção de ocitocina, durante o trabalho de
parto, é decorrente de um reflexo neuroendócrino
desencadeado por estimulação mecânica de estruturas
componentes do trato genital inferior (cérvix e vagina).
A distensão da cérvix uterina provocada pelas primeiras
contrações uterinas, as quais independem da ocitocina
na espécie humana, leva à estimulação dos receptores
de estiramento aí localizados; esses potenciais de ação
propagam-se
por
fibras
aferentes
específicas
que
chegam hipotálamo – via medula espinal e tronco
25 | P á g i n a
______________Digite o Título do Documento______________
encefálico
–
onde
fazem
sinapse
com
neurônios
ocitocinérgicos. O resultado desse processo é o aumento
da secreção de ocitocina que, atuando na musculatura
uterina, induz novas contrações e realimenta o processo
que leva à sua secreção por um mecanismo de feedback
positivo, esse mecanismo perdura até a expulsão do
feto.
Ação sobre a glândula mamária
As ações da ocitocina sobre a glândula mamária
estão relacionadas com o processo de ejeção do leite dos
alvéolos e ductos galactóforos menores. Estas estruturas
são
envolvidas
por
células
mioepiteliais,
alvos
da
ocitocina; a contração dessas células leva à ejeção do
leite armazenado. Este processo também é regulado por
um mecanismo reflexo, conhecido como reflexo de
ejeção do leite, desencadeado em resposta à sucção do
mamilo. O reflexo envolve a estimulação de terminações
nervosas presentes no mamilo (mecanoceptores), de
onde potenciais de ação são transmitidos à medula
espinal, tronco encefálico e hipotálamo, onde então
atingem os neurônios ocitocinérgicos. A ativação desses
neurônios leva à secreção de ocitocina e contração das
células mioepiteliais; segue-se um aumento da pressão
intramamária
e
ejeção
do
leite
para
os
ductos
galactóforos maiores e cisternas, de onde pode ser
obtido passivamente por sucção. Esse processo não deve
ser confundido com o de secreção de leite, o qual é
controlado
pela
prolactina.
O
aleitamento
depende
totalmente desse reflexo que, na espécie humana, pode
ser condicionado ao choro de um bebê.
26 | P á g i n a
______________Digite o Título do Documento______________
Regulação da Secreção de Ocitocina
A estimulação de mecanoceptores localizados na
cérvix uterina, canal vaginal e mamilo leva a um
aumento
da
liberação
experimentais
sugerem
de
ocitocina.
que
o
As
evidências
neurotransmissor
envolvido nesses mecanismos seja a acetilcolina. Por
outro lado, as catecolaminas exercem efeitos inibitórios
sobre a liberação de ocitocina. Isto pode ser evidenciado
em situações de estresse, onde ocorre inibição da
secreção de leite. A administração de ocitocina nessa
condição é capaz de restabelecer o fornecimento de
leite, sugerindo
que
essa inibição
decorra
de um
bloqueio central da liberação de ocitocina.
Existem evidências de que a inibição da ejeção de
leite
causada
pelo
estresse
também
possa
ser
o
resultado da constrição de vasos sanguíneos da glândula
mamária, o que dificultaria o acesso da ocitocina ao seu
sítio
de
ação.
Nesse
sentido,
verificou-se
que
a
adrenalina é capaz de inibir a resposta contrátil de fatias
isoladas de tecido mamário, por ação direta nas células
mioepiteliais. Assim, acredita-se que a adrenalina ou a
noradrenalina cause inibição do reflexo de ejeção de leite
agindo centralmente, por bloqueio da liberação de
ocitocina,
e
em
menor
grau,
perifericamente,
dificultando ou inibindo a ação desse hormônio na
glândula mamária.
6.1.4 Glândula Pineal
Produz o hormônio melatonina no período noturno
(pico
das
20:00h
às
08:00h).
Esse
hormônio
é
responsável pelo controle do ritmo sazonal dia/noite e
pelo controle do processo reprodutivo, modulando as
27 | P á g i n a
______________Digite o Título do Documento______________
épocas de acasalamento em diferentes espécies, por
meio da indução ou inibição do cio. Por exemplo:
- Felinos e Equinos: Positivamente influenciados pelo
aumento da luz. Efeito antigonadotrópico: acasalam
primavera/verão.
- Caprino e Ovinos: Positivamente influenciados pela
redução
da
luz.
Efeito
indutivo:
acasalam
outono/inverno.
6.1.5 Pâncreas Endócrino
O pâncreas é um órgão responsável por dois tipos
de mecanismos secretores: Endócrino e exócrino. O
pâncreas exócrino secreta aproximadamente 1 Litro de
secreção no duodeno contendo água, bicarbonato e
enzimas para a digestão de carboidratos, proteínas e
lipídios. O pâncreas exócrino é composto de ácinos e
contempla 90% do órgão. O pâncreas endócrino, as
ilhotas de Langerhans, contém quatro tipos de células:
beta - secretam insulina; alfa - secretam glucagon; delta
- secretam somatostatina e células PP que secretam
polipeptídeo pancreático. Os neurônios simpáticos e
parassimpáticos possuem terminais nervosos nas ilhotas
por intermédio dos quais o sistema nervoso pode
influenciar o metabolismo. A proporção entre insulina e
glucagon determina a direção do metabolismo.
Insulina
É um hormônio polipeptídico constituído de duas
cadeias contendo 21 e 30 aminoácidos ligadas por duas
pontes de dissulfeto. A insulina é sintetizada como prépro-insulina no retículo endoplasmático rugoso e no
aparelho
de
Golgi,
sofre
clivagens
proteolíticas
28 | P á g i n a
______________Digite o Título do Documento______________
sucessivas em pró-insulina e a seguir em insulina e
peptídeo C que são armazenados nos grânulos das
células beta e secretados em quantidades equimolares.
Regulação da secreção de Insulina
A ingestão de glicose é o estímulo mais potente
para a secreção de insulina. A glicose oral estimula a
secreção de GIP (Peptídeo insulinotrópico dependente da
glicose) que tem efeito estimulador independente sobre
a secreção de insulina. A glicose ou os aminoácidos no
lúmen do intestino delgado liberam GIP a partir das
células endócrinas sendo levado pela circulação pelas
células
beta
chegando
antes
da
primeira
glicose
absorvida. A liberação antecipatória de insulina impede
um surgimento súbito de glicose no plasma quando a
refeição é absorvida. Este efeito é reforçado pela
estimulação colinérgica parassimpática que acompanha a
digestão.
6.1.5.1 Tabela Regulação da secreção da insulina
ESTIMULADORES
INIBIDORES
Aumento da glicose
Diminuição da
glicose
Aumento de
Jejum
aminoácidos
Aumento de AG e
Exercício
cetoácidos
Glucagon
Somatostatina
Cortisol
Agonistas αadrenérgicos
K+
Obesidade
29 | P á g i n a
______________Digite o Título do Documento______________
Após a ingestão de glicose e aumento da glicemia,
a glicose penetra nas células beta por difusão facilitada,
através de um receptor de membrana denominado Glut2. Dentro da célula a glicose é fosforilada em glicose-6fosfato pela glicocinase e a seguir esta é oxidada. Um
dos produtos desta fase oxidativa é o ATP. O ATP fecha
os canais de K na membrana da célula beta, o que
despolariza a membrana da célula. A despolarização
abre os canais de Cálcio-voltagem dependentes. O Cálcio
flui para o interior da célula beta a favor do seu
gradiente
eletroquímico
e
provoca
exocitose
dos
grânulos secretores contendo insulina.
A
insulina
é
secretada
no
sangue
venoso
pancreático direto para a circulação porta. A insulina
liga-se a receptor específico na superfície de suas
células-alvo. Esse receptor, chamado de receptor ligado
à
atividade
tirosina-quinase,
é
um
complexo
transmembrana composto de duas subunidades alfa e
duas subunidades beta ligadas por pontes dissulfeto. As
subunidades alfa são extra-celulares e possuem sítios de
ligação da insulina. As subunidades beta possuem
atividade tirosina-quinase. A ligação da insulina provoca
alteração conformacional ativando a atividade tirosinaquinase das subunidades beta, que atuam uma sobre a
outra (autofosforilação incestuosa) bem como sobre
outras proteínas alvo (IRS – substrato do receptor de
insulina).
complexos
vesículas,
Após
sua
interação
insulina-receptor
resultando
em
com
são
a
insulina,
internalizados
regulação
decrescente.
os
em
A
insulina é degrada por lisossomos e o receptor pode ser
reciclado. As proteínas de substrato do receptor de
insulina (IRS- insulin receptor substrate) sofrem rápida
30 | P á g i n a
______________Digite o Título do Documento______________
fosforilação da tirosina em resposta à insulina e ao fator
de crescimento semelhante à insulina 1 (IGF-1). O
substrato mais bem caracterizado é o substrato IRS-1,
este interage com proteínas que contém um denominado
domínio SH2 (sequencia de reconhecimento do substrato
de insulina) passando adiante o sinal da insulina. A
ativação do IRS-fosforilado com o domínio SH2 da
fosfatidilinositol-3-quinase
exerce
vários
efeitos
importantes, incluindo recrutamento de transportadores
GLUT-4 de glicose para a membrana plasmática nas
células musculares e adipócitos.
Ações da Insulina
Ações sobre o fígado, músculo e gorduras. Seu
efeito consiste em conservar os combustíveis energéticos
ao facilitar a captação e o armazenamento da glicose,
dos aminoácidos e dos lipídios. Reduz o nível de
glicemia. Estes nutrientes estarão disponíveis durante os
períodos de jejum mantendo a oferta de glicose ao
cérebro, músculos e outros órgãos. (1) A insulina
aumenta o transporte de glicose no músculo e tecido
adiposo por comandar a inserção de transportadores de
glicose (GLUT 4) nas membranas celulares. Quando a
glicose entra nas células, diminui a glicemia. (2) A
insulina promove a formação de glicogênio a partir da
glicose no fígado e no músculo e ao mesmo tempo inibe
a glicogenólise (degradação do glicogênio). (3) A insulina
inibe a gliconeogênese (síntese de glicose de fontes
diferentes
dos
carboidratos),
os
substratos
são
direcionados para longe da formação de glicose. (4)
Diminuição da concentração sanguínea de ácidos graxos
e cetoácidos e aumenta o armazenamento destes. Inibe
31 | P á g i n a
______________Digite o Título do Documento______________
a lipólise por inativação das lipases. A insulina também
provoca lipogênese no fígado. (5) A insulina aumenta a
captação de aminoácidos e de proteínas e inibe sua
degradação (músculo). Inibe a oxidação dos aminoácidos
no fígado.
A
longo
prazo,
é
um
importante
hormônio
anabólico, particularmente durante o desenvolvimento
fetal. Estimula a proliferação celular e está implicada no
crescimento e desenvolvimento somáticos e viscerais. No
crescimento, age junto ao GH. Também tem ação no
centro da saciedade do hipotálamo, informando que o
organismo pode parar de ingerir alimentos.
Patofisiologia da Insulina
A diabetes mellitus é causada pela diminuição da
produção de insulina ou pela resistência a insulina. Isso
causa aumento da glicemia, mobilização acentuada das
gorduras e deposição de colesterol nas paredes arteriais
causando aterosclerose e depleção das proteínas nos
tecidos do corpo. Sempre que a quantidade de glicose
que entra nos túbulos renais, no filtrado glomerular sobe
acima de um nível crítico (acima de 180 mg/dl), uma
proporção
significativa
do
excesso
não
pose
ser
reabsorvida e extravasa na urina (glicosúria). A pressão
osmótica aumentada nos líquidos extracelulares causa a
transferência osmótica da água para fora das células.
Além disso, a glicose diminui a reabsorção tubular de
líquido causando desidratação do líquido extracelular que
por sua vez causa desidratação compensatória do líquido
intracelular
e
isso
pode
contribuir
para
o
choque
circulatório. A poliúria é devido ao efeito diurético
osmótico da glicose nos túbulos renais. Por sua vez a
32 | P á g i n a
______________Digite o Título do Documento______________
polidipsia é devida à desidratação resultante da poliúria.
O colapso do metabolismo da glicose (e da proteína)
causa a perda de peso e uma tendência a polifagia. A
falta de energia (astenia) é causada pela perda da
proteína
corporal
e
pela
utilização
diminuída
dos
carboidratos para energia.
O desvio do metabolismo dos carboidratos para o
das gorduras aumenta o nível de cetoácidos na corrente
sanguínea (ácido acetoacetico e beta-hidroxibutírico).
Esses ácidos são excretados com o Na+ derivado do
líquido extracelular. Consequentemente, a concentração
de Na+ diminui e parte do Na+ é substituída por íons
Hidrogênio aumentando a acidose.
Diabetes
Diabetes Melito Tipo I é causado pela secreção
insuficiente de insulina e
tipo
II é resultante da
resistência a insulina. O diabetes tipo I é causado pela
destruição das células beta, (autoimune) por interação
de fatores genéticos e ambientais. Em geral os pacientes
são jovens e não obesos, quando os sintomas aparecem
pela primeira vez. Existe uma predisposição hereditária
com uma incidência 10 vezes maior em parentes de
primeiro grau. Indivíduos geneticamente predispostos
também devem ser expostos a um fator ambiental,
como infecção viral (p.ex. vírus Coxksackie ou Echo).
O diabetes tipo II é com frequência associado a
obesidade. É causado pela regulação para menos dos
receptores de insulina nos tecidos-alvo e resistência a
insulina. A insulina é secretada normalmente pelas
células beta, mas em concentrações normais ela não
pode ativar seus receptores no músculo, no fígado e no
33 | P á g i n a
______________Digite o Título do Documento______________
tecido adiposo. De modo típico a hiperglicemia está
presente em jejum e no período pós-prandial.
Glucagon
Polipeptídeo
de
cadeia
simples
com
29
aminoácidos. É o hormônio da inanição, da desnutrição e
do jejum. Sintetizado pelas células alfa das ilhotas de
Langerhans.
Promove
utilização
e
mobilização
de
combustíveis metabólicos. O principal estimulador da
secreção de glucagon é a concentração sanguínea de
glicose diminuída.
A atividade nervosa simpática e a
adrenalina circulante estimulam a liberação de glucagon
através
dos
receptores
beta
adrenérgicos.
A
somatostatina inibe a sua liberação.
Mecanismo de ação do Glucagon
As principais ações são no fígado. O hormônio se
liga a um receptor da membrana celular acoplado a
adenililciclase por meio de proteína Gs. O segundo
mensageiro é o AMPc que ativa proteína-quinases que
por sua vez fosforilam várias enzimas; a seguir as
enzimas fosforiladas medeiam as ações fisiológicas do
glucagon.
Ações do Glucagon
A ação do Glucagon se opõe a da insulina. O
glucagon aumenta a concentração sanguínea de glicose,
pois estimula a glicogenólise e degradação de glicogênio.
Aumenta a gliconeogênese por diminuir a produção de
frutose 2,6-bifosfato. Assim o substrato é direcionado
para
a
formação
de
glicose.
Os
aminoácidos
são
utilizados para a gliconeogênese e os grupamentos
34 | P á g i n a
______________Digite o Título do Documento______________
amino resultantes são incorporados na uréia. O glucagon
também aumenta a lipólise e desvia os substratos para a
gliconeogênese. Portanto, causa lipólise no fígado e nos
adipócitos (aumento da gliconeogênese) e o catabolismo
das proteínas no músculo.
Somatostatina
Polipeptídeo com 14 aminoácidos secretado pelas
células delta. A secreção é estimulada por todas as
formas de ingestão de nutrientes, pelo glucagon e pelos
agonistas beta-adrenérgicos. A secreção é inibida pela
insulina por meio de mecanismo parácrino. Ações: inibe
a secreção de insulina e glucagon por meio de ações
parácrinas nas células alfa e beta. Sua função parece ser
limitar ou modular as respostas da insulina e do
glucagon ao alimento ingerido.
6.1.6 Paratireóide
As glândulas paratireóides são quatro ou mais
glândulas pequenas localizadas na superfície posterior da
tireóide e possuem células densamente agrupadas. A
função principal das paratireóides é manter o nível de
cálcio
no
sangue
dentro
do
limite
apropriado
ao
funcionamento dos sistemas nervoso e muscular.
Metabolismo do Cálcio e Fosfato
O Cálcio tem importantes papéis na contração
muscular, formação de ossos, coagulação sanguínea,
homeostasia. A homeostasia do cálcio proveniente da
dieta depende da ação integrada e coordenada dos ossos
(estoque), rins (reabsorção) e intestino (absorção).
35 | P á g i n a
______________Digite o Título do Documento______________
Existem três hormônios participando desse controle. O
paratormônio, que é o mais importante, a calcitonina
que é um refinamento do paratormônio (efeito oposto ao
paratormônio) e a vitamina D, um esteróide.
Paratormônio (PHT)
É
produzido
nas
paratireóides
e
regula
a
concentração plasmática de Ca++. Para isso, aumenta a
concentração de Cálcio no LEC, caso essa se reduza a
níveis inferiores a 10mg/dl. Dessa forma, maiores
concentrações de Cálcio, diminuem a secreção de PHT e
menores concentrações de Cálcio, estimulam a secreção
do PHT.
Quando ocorre o aumento da concentração de
cálcio no LEC, ocorre ativação de receptor ligado a
proteína G na membrana da paratireóide e o segundo
mensageiro resultante inibe a secreção do PTH.
Já as
baixas concentrações de cálcio, inativam o receptor,
diminuindo
a
produção
do
segundo
mensageiro,
estimulando a secreção do PTH.
Para aumentar a concentração de cálcio, o PTH age
nos tecidos ósseo, renal e intestino. Nos ossos, estimula
a atividade dos osteoclastos. Este reabsorve Ca++,
formando fosfato de cálcio. Nos rins, o PTH favorece a
reabsorção do cálcio e negligencia a reabsorção de
fosfato. No intestino, estimula a ação da vitamina D, que
aumenta a absorção intestinal de cálcio, o que se traduz
num incremento rápido e sustentado da quantidade de
cálcio no sangue.
Calcitonina
36 | P á g i n a
______________Digite o Título do Documento______________
Produzida nas células parafoliculares da tireóide.
Ação oposta ao do PHT, ou seja, diminui a secreção de
cálcio no sangue, inibe a reabsorção óssea e diminui
formação e liberação de Cálcio.
Vitamina D
Também chamada de colecalciferol (forma inativa).
Importante na mineralização óssea e na presença do
PTH aumenta as concentrações plasmáticas de cálcio.
Diferente do PTH, a vitamina D favorece o incremento
dos níveis séricos de Ca++ e fosfato. A vitamina D pode
ser produzida nos queratinócitos da pele estimulados
pelos raios UV do sol a partir do colesterol vindo da dieta
ou então pode ser adquirida pela ingestão do próprio
colecalciferol.
Existem duas formas do colecalciferol, a forma
biologicamente ativa que é o 1,25-di-hidroxicalciferol
(calcitriol)
e
a
hidroxicalciferol.
forma
Então
inativa
para
se
que
é
tornar
o
24,25ativo
o
colecalciferol sofre duas hidroxilações, uma no fígado e
outra no rim. O que define se será a forma ativa ou
inativa resultante é a concentração de cálcio. Sendo que
a baixa concentração de cálcio, de fosfato e o PTH
favorecem a forma ativa.
Ações da vitamina D
Na membrana intestinal a vitamina D se liga a
receptores intracelulares, e dentro das células estimula a
síntese da proteína Calbinidina, causando incremento da
absorção intestinal de Ca++ por atuar como um tampão.
Nos ossos, estimula a remodelação óssea (ação de
37 | P á g i n a
______________Digite o Título do Documento______________
osteoclastos). E nos rins, aumenta a reabsorção de
cálcio e fosfato.
6.2 Bibliografia
COSTANZO, L.S. Fisiologia.
Guanabara Koogan, 2006.
3.ed.
Rio
de
Janeiro:
CUNNINGHAM J.G. Tratado de fisiologia veterinária. Rio
de Janeiro: Guanabara, 454p, 1999.
GUYTON, A. C. & HALL, J. E. Tratado de Fisiologia
Médica. 9.ed. Rio de Janeiro: Guanabara Koogan, 1996.
SILVERTHORN, D. E.; OBER, W. C.; GARRISON, C. W.;
SILVERTHORN, A. C. Fisiologia humana: uma abordagem
integrada. 2.ed. São Paulo: Manole. cap 15, p., 2003.
SWENSON, M. J., AND W. O. Reece. Dukes-Fisiologia dos
animais domésticos. 11.ed. Guanabara, Rio de Janeiro,
Brasil, 1996.
38 | P á g i n a