EXISTE INFLUÊNCIA DA GRAVIDADE SOBRE A VELOCIDADE
DE CAPTURA DE PRESAS NA ARANHA CYCLOSA FILILINEATA
(ARANEAE, ARANEIDAE)?
Gustavo R. Mazão
INTRODUÇÃO
Entre as diversas estratégias de captura de presas
existentes está a estratégia de senta-e-espera (Viera
et al. 2007), que oferece benefícios aos predadores
devido ao pequeno gasto energético (Riechert &
Luczak 1982) e custos devido ao longo tempo de espera de suas presas (Wise 1993). Esse tipo de estratégia consiste no comportamento dos predadores de
aguardar suas presas em pontos estratégicos até que
estas passem, para neste momento apanhá-las, economizando o gasto energético da procura e da perseguição. As aranhas construtoras de teias provavelmente constituem o grupo de predadores que melhor ilustra essa estratégia de predação, merecendo
destaque em seu papel ecológico como predadores
(Foelix 1982).
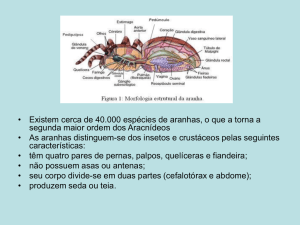
As aranhas podem ser ecologicamente classificadas
em dois grupos: aranhas errantes, que procuram
ativamente suas presas e aranhas sedentárias que,
em sua maioria, produzem teias para interceptar
suas presas (Gray 1978). As aranhas construtoras
de teias orbiculares, em geral, constróem teias
assimétricas, sendo na maior parte das vezes a região inferior maior que a superior (Sendoda et al.
2007). Uma hipótese para explicar essa assimetria
é que o deslocamento das aranhas é mais rápido na
descida da teia do que na subida, devido ao auxílio
da gravidade. Desse modo, a aranha investiria mais
na construção da parte inferior da teia, onde seu
deslocamento é facilitado pela gravidade. Portanto,
uma premissa dessa hipótese é que exista diferença
entre as velocidades de descida e subida das aranhas em suas teias, sendo a velocidade de descida
maior que a velocidade de subida.
As aranhas da espécie Cyclosa fililineata (Araneidae)
apresentam a parte inferior da teia maior que a superior, e constróem suas teias em um ângulo perpendicular ao chão e, portanto, são consideradas um
bom modelo de estudo para se testar a hipótese da
maior velocidade no deslocamento para baixo. O objetivo do presente trabalho foi testar se existe diferença entre as velocidades de subida e descida das
aranhas C. fililineata ao realizarem capturas de presas.
MATERIAIS E MÉTODOS
Foram coletados em uma área de restinga baixa no
Parque Estadual da Ilha do Cardoso (25°03’S;
47°53’W), 28 indivíduos (adultos e jovens) de C.
fililineata. As aranhas foram coletadas e mantidas
vivas durante três dias em cativeiro, onde foram
colocadas individualmente em galhos secos que simulavam a vegetação. No primeiro dia não foram
realizados testes com as aranhas para que estas se
estabelecessem e construíssem suas teias. Após o
período de estabelecimento e construção das teias,
as aranhas foram estimuladas a realizarem ataques
sobre pernilongos (Diptera: Culicidae) que eram introduzidos na teia com uma pinça. As presas eram
colocadas, uma em cada teia, em pontos do eixo central que se estende do ponto mais alto ao mais baixo
da teia, passando pelo centro. Depois que a presa
era colocada na teia, eram medidos a distância percorrida e o tempo gasto pela aranha até sua chegada
na presa. Cada aranha foi testada duas vezes, sendo
aguardado um período de 24 h entre os experimentos para que as aranhas reconstruíssem suas teias,
que haviam sofrido danos oriundos da introdução da
presa. A orientação no eixo (acima ou abaixo do centro) em que a presa iria ser colocada era sorteada
para cada aranha em cada um dos experimentos
antes do experimento. Os casos em que não houve
resposta por parte das aranhas foram descartados
da análise.
A velocidade média de cada aranha foi calculada através da fórmula Vm=Dd/Dt (Vm = velocidade média;
Dd = deslocamento; Dt = tempo). Para se comparar
a diferença entre as velocidades de subida e descida
das aranhas foi realizado um teste-t e, posteriormente, uma análise de regressão entre as velocidades
médias de subida e descida e as respectivas distâncias percorridas pelas aranhas.
RESULTADOS
Foram analisadas 28 aranhas, sendo obtido um total de 27 observações, 12 de subida e 15 de descida. A
1
diferença entre o número de aranhas e observações
se deu porque diversas vezes as aranhas não se deslocaram até suas presas. As médias e desvios padrões das velocidades de subida e descida foram 1,99
± 1,05 cm/s e 2,27 ± 1,09 cm/s, respectivamente (Fig.
1), não havendo diferença significativa entre essas
velocidades (t=0,67; g.l.=25; p=0,92).
Figura 1. Representação dos dados de velocidade média
para subida e descida das aranhas C. fililineata (? média; ? - erro padrão; I - desvio padrão; ? - pontos
extremos).
Na análise dos dados de velocidade de subida das
aranhas, o modelo que melhor se ajustou aos dados
foi um polinomial quadrático (R2 = 0,89; Fig. 2A). Já
os dados de velocidade de descida não apresentaram
nenhuma relação significativa com a distância em
nenhum dos modelos de regressão (Fig. 2B), tanto o
linear (F=3,17; p=0,09) quanto o polinomial
quadrático (F=1,75; p=0,21).
Foi observado que, durante a execução do experimento, em alguns casos no qual a presa era
introduzida na parte superior da teia, as aranhas
sacudiam suas teias de modo que a presa se desprendia e caía para mais próximo de onde a aranha
se encontrava. Nos casos em que as presas eram
introduzidas na parte de baixo da teia as aranhas
muitas vezes realizavam o deslocamento com breves pausas, retardando o movimento de descida.
DISCUSSÃO
É comum encontrar assimetria nas teias orbiculares,
de forma que geralmente a parte inferior é maior
que a superior (Herberstein & Heiling 1999). A aranha Cyclosa fililinata não foge a este padrão (Sendoda
et al. 2007). Porém, a hipótese de que a velocidade
que as aranhas atingem na descida seria maior do
que na subida foi rejeitada neste trabalho. Como foi
observado, não há diferença significativa entre as
Figura 2. Relação entre os dados de velocidade média
(Vm=Dd/Dt) de subida (A) e descida (B) das aranhas
Cyclosa fililineata, em função da distância percorrida
até sua presa.
velocidades médias de subida e descida das aranhas
em suas teias.
Adicionalmente, existem diferentes padrões de comportamento de subida e descida em C. fililineata. O
padrão observado de subida das aranhas em suas
teias provavelmente decorre do fato dos estímulos
mais distantes indicarem a necessidade de um rápido deslocamento. Nesse caso, como a velocidade depende apenas da força empregada pela aranha (e não
da gravidade), os estímulos oriundos de presas na
periferia da teia devem estimular que a aranha se
desloque com maior velocidade. A ausência de padrão para os valores observados de velocidade de descida pode ser explicada pelo fato dos indivíduos agirem de forma a retardar o movimento, parando brevemente durante o movimento descendente. Como
no movimento de descida são somadas a força aplicada pela aranha e a força gravitacional, faz-se necessário que os indivíduos retardem seu movimento
para não perder o controle do movimento, e acabar
ultrapassando suas presas. O comportamento de
retardar a descida também pode estar associado à
economia de energia, tendo em vista que o movimento para subjugar as presas demanda um alto
gasto de energia, que não pode ser dispensada com
erros de ataque. De maneira análoga, o comportamento de sacudir a teia para que as presas captura2
das na parte de cima da teia caiam mais próximas
da aranha provavelmente está associado à economia
de energia durante a predação.
Sabendo que existe interferência da força
gravitacional sobre o deslocamento de C. fililineata,
sugiro a realização de trabalhos futuros para elucidar
a mecânica de tração de subida e descida dessas aranhas em suas teias. Tais trabalhos ajudariam a compreensão das diferentes respostas (subida e descida
na teia) da C. fililineata e sua relação com a força
gravitacional. Esses estudos são muito importantes
para fundamentar hipóteses explicativas sobre a
construção de teias assimétricas por aranhas.
M.O.; Santos, A.J. & Japyassú, H.F., eds.). Editora Interciência, Rio de Janeiro.
Wise, D.H. 1993. Spiders in ecological webs (Wise,
D.H., ed.). Cambridge University Press, Melbourne.
AGRADECIMENTOS
Agradeço a todos os coordenadores do Curso
de Campo por terem possibilitado minha presença
no curso e, conseqüentemente, a realização deste
trabalho. Também agradeço a todos os alunos do
curso que proporcionaram um excelente ambiente
de trabalho, em especial a Liliam, Tiago e Victor
que me ajudaram na coleta das presas utilizadas, e
também a Denise pelo auxílio na coleta das aranhas.
BIBLIOGRAFIA
Foelix, R.F. 1982. Lomomotion and Prey Capture,
pp148-171. In: Biology of spiders (Foelix, R.F. ed.).
Harvard University Press, Cambridge.
Gray, M.R. 1978. Spider webs, pp. 109-148. In:
Biology of spiders (Foelix, R.F. ed.). Harvard
University Press, Cambridge.
Herberstein, M.E. & Heiling, A.M. 1999. Asymmetry
in spider orb webs: a result of physical constraints?
Animal Behaviour 58: 1241-1246.
Ricklefs, R.E. 2003. A economia da natureza. Editora Guanabara Koogan, Rio de Janeiro.
Riechert, S.E. & Luczak, J. 1982. Spider foraging:
behavioral responses to prey, pp. 353-385. In:
Spider communication: mechanisms and
ecological significance (Witt, P.N. & Rovner, J.S.,
eds.). Princeton University Press, Princeton.
Sendoda, A., Pinotti, B., Mazão, G. & Pinto, L. Existe uma relação entre inclinação e simetria de teias em aranhas orbitelas? Livro curso de campo
“Ecologia da Mata Atlântica”. Universidade de São
Paulo e Universidade Estadual de Campinas.
Viera, C., Japyassú, H.F., Santos, A.J. & Gonzaga,
M.O. 2007. Teias e forrageamento, pp. 45-65. In:
Ecologia e comportamento de aranhas (Gonzaga,
3