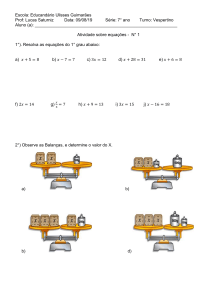
Síntese e degradação de bases púricas e pirimídicas - Rui Fontes
Síntese e degradação de bases púricas e pirimídicas
Índice
1-
A estrutura das bases púricas e das bases pirimídicas .......................................................................... 1
2-
A estrutura e a nomenclatura dos nucleosídeos púricos e pirimídicos ................................................. 1
3-
A estrutura e a nomenclatura dos nucleotídeos púricos e pirimídicos .................................................. 2
4-
A digestão dos ácidos nucleicos e a absorção e destino metabólico dos nucleosídeos da dieta ........... 2
5-
A síntese de novo das purinas ............................................................................................................... 2
6-
A síntese de AMP e GMP a partir do inosinato.................................................................................... 2
7-
Vias de salvação de bases púricas e de nucleosídeos ........................................................................... 3
8-
As cínases de nucleosídeos monofosfato e a cínase de nucleosídeos difosfato.................................... 3
9-
A redução dos ribonucleosídeos difosfato pelas formas reduzidas da tioredoxina e glutaredoxina ..... 4
10-
O catabolismo dos nucleotídeos púricos .......................................................................................... 4
11-
A conversão de AMP em ácido úrico ............................................................................................... 4
12-
A conversão da guanosina em ácido úrico ....................................................................................... 5
13-
Causas de hiperuricemia e a gota ..................................................................................................... 5
14-
A síntese de novo das pirimidinas .................................................................................................... 6
15-
A síntese de CTP, de UTP e do 2’-desoxiuridilato a partir de uridina difosfato .............................. 6
16-
A síntese do timidilato a partir de 2’-desoxiuridilato e o ciclo do dihidrofolato .............................. 7
17-
A fosforibosil-transférase do uracilo e as cínases de nucleosídeos pirimídicos ............................... 7
18-
O catabolismo dos nucleosídeos pirimídicos e das bases pirimídicas .............................................. 8
19-
A regulação da síntese de nucleotídeos púricos e pirimídicos.......................................................... 8
20-
A regulação da redútase dos ribonucleosídeos difosfato .................................................................. 9
21A importância da glutamina quer como substrato na síntese de nucleotídeos quer como nutriente
nas células em proliferação rápida................................................................................................................ 9
22-
Fármacos inibidores da síntese de nucleotídeos ............................................................................... 9
23-
Ação de enzimas do metabolismo dos nucleotídeos no efeito terapêutico do aciclovir ................... 9
1- A estrutura das bases púricas e das bases pirimídicas
Os nucleotídeos contêm resíduos de ácido fosfórico, de um açúcar (em geral uma pentose:
ribose ou 2'-desoxiribose) e de uma base púrica ou pirimídica. Quer as bases púricas, quer as
pirimídicas são anéis aromáticos heterocíclicos contendo átomos de azoto e carbono. As bases púricas
podem ser entendidas como constituídas por um anel pirimidina (anel com 6 átomos: 4C,2N) ligado a
um anel imidazol (anel com 5 átomos: 3C,2N). São bases púricas a adenina (6-aminopurina), a
guanina (2-amino-6-oxipurina), a hipoxantina (6-oxipurina) e a xantina (2,6-dioxipurina). São bases
pirimídicas a citosina (2-oxi-4-aminopirimidina), o uracilo (2,4-dioxipirimidina), a timina (2,4dioxi-5-metilpirimidina) e o ácido orótico (2,4-dioxi-6-carboxipirimidina).
2- A estrutura e a nomenclatura dos nucleosídeos púricos e pirimídicos
Por hidrólise dos nucleotídeos (saída dos resíduos fosfato) geram-se nucleosídeos púricos
(adenosina, guanosina, inosina, xantosina) ou pirimídicos (citidina, uridina, timidina e
orotidina) que contêm uma base e uma ose ligados por uma ligação glicosídica de tipo N. No caso
dos nucleosídeos púricos, as palavras que os designam terminam em “osina” e a ligação glicosídica
envolve o átomo N9 da base. A inosina é o nucleosídeo que contém hipoxantina; nos outros casos a
relação entre as designações da base púrica e do nucleosídeo respetivo é óbvia. No caso dos
Página 1 de 12
Síntese e degradação de bases púricas e pirimídicas - Rui Fontes
nucleosídeos pirimídicos as palavras que os designam terminam em “dina” e a ligação glicosídica
envolve o átomo N1 da base.
3- A estrutura e a nomenclatura dos nucleotídeos púricos e pirimídicos
Os nucleosídeos monofosfato (NMP) são os nucleotídeos mais simples e designam-se de
acordo com o nucleosídeo constituinte: adenilato (AMP), guanilato (GMP), inosinato (IMP),
xantinilato (XMP), citidilato (CMP), uridilato (UMP), timidilato (TMP) e orotidilato (OMP). Se
não se específica o contrário subentende-se que o fosfato está ligado (ligação fosfoéster) no hidroxilo
5’ da pentose.
4- A digestão dos ácidos nucleicos e a absorção e destino metabólico dos nucleosídeos da
dieta
Os ácidos nucleicos constituintes da dieta são hidrolisados no lúmen intestinal por ação de
nucléases pancreáticas e fosfátases intestinais formando-se nucleosídeos que são absorvidos.
Contudo, a quase totalidade das purinas e a maior parte das pirimidinas que os constituem não são
incorporados nos ácidos nucleicos do organismo, mas antes degradadas na mucosa intestinal e no
fígado sendo os produtos do seu catabolismo (ácido úrico no caso das purinas e CO2 + ureia + β aminoisobutirato no caso das pirimidinas) excretados [1].
5- A síntese de novo das purinas
Na síntese de novo das purinas todas as enzimas são citoplasmáticas e todos os
intermediários contêm ribose-5-fosfato. O primeiro nucleotídeo formado é a inosina-5’-fosfato (IMP
ou inosinato) cuja base é a hipoxantina (6-oxipurina). Sendo uma via complexa destacamos apenas
alguns passos do processo.
Na primeira reação a ribose-5-fosfato, formada na via das pentoses-fosfato, funciona como
aceitador dos fosfatos β-γ do ATP que se ligam no carbono 1 da ribose gerando-se fosfo-ribosilpirofosfato (PRPP); esta reação é catalisada pela síntase do PRPP (ver Equação 1). Na segunda
reação ocorre rotura da ligação formada na primeira reação e transferência do azoto do grupo amida
da glutamina para o carbono 1 da ribose formando-se 5-fosforibosilamina; esta reação é catalisada
pela amido-fosforibosil-transférase da glutamina (ver Equação 2). O azoto N9 do anel purina
deriva assim da glutamina.
Equação 1
Equação 2
ribose-5-fosfato + ATP → PRPP + AMP
PRPP + glutamina → 5-fosforibosilamina + glutamato + PPi
Sucessivamente vão-se incorporando a glicina (que origina os átomos C4, C5 e N7 das
purinas), uma unidade monocarbonada cedida pelo N10-formil-H4-folato (C8), outro azoto do grupo
amida da glutamina (N3), o CO2 (C6), o grupo amina do aspartato (N1) e outra unidade
monocarbonada cedida pelo N10-formil-H4-folato (C2).
A equação geral que descreve o processo da síntese de novo das purinas entre ribose-5fosfato e IMP é a Equação 3. Essa equação evidencia que a formação de alguns dos intermediários
fosforibosilo da via metabólica está acoplada com a rotura de ligações fosfoanidrido do ATP, que os
carbonos do aspartato saem na forma de fumarato e que o N10-formil-H4-folato se converte em H4folato e a glutamina em glutamato.
Equação 3
ribose-5-fosfato + 5 ATP + 2 glutamina + glicina + aspartato + 2 N10-formil-H4-folato +
CO2 →
IMP + AMP + 4 ADP + PPi + 4 Pi + 2 glutamato + 2 H4-folato + fumarato
6- A síntese de AMP e GMP a partir do inosinato
É a partir do IMP que se formam quer o AMP (por aminação do carbono 6), quer o GMP
(oxidação dependente do NAD+ e posterior aminação do carbono 2).
O dador do grupo 6-amina do AMP é o aspartato. Na formação do AMP a partir do IMP
intervém a sintétase de adenilosuccinato (ver Equação 4); o adenilosuccinato formado sofre, de
seguida, a ação de uma líase desdobrando-se em AMP e fumarato (líase do adenilosuccinato; ver
Equação 5). A equação soma que descreve a síntese de AMP a partir de IMP é a Equação 6.
Página 2 de 12
Síntese e degradação de bases púricas e pirimídicas - Rui Fontes
Equação 4
Equação 5
Equação 6
IMP + aspartato + GTP → adenilosuccinato + GDP + Pi
adenilosuccinato → AMP + fumarato
IMP + aspartato + GTP → AMP + GDP + Pi + fumarato
O dador do grupo 2-amina do GMP é a glutamina; na formação do GMP a partir do IMP
intervém a desidrogénase do IMP que origina XMP (ver Equação 7); o intermediário XMP funciona,
de seguida, como aceitador do grupo amina da glutamina (sintétase do guanilato; ver Equação 8). A
equação soma que descreve a síntese de GMP a partir de IMP é a Equação 9.
Equação 7
Equação 8
Equação 9
IMP + NAD+ → XMP + NADH
XMP + glutamina + ATP → GMP + glutamato + AMP + PPi
IMP + NAD+ + glutamina + ATP → GMP + NADH + glutamato + AMP + PPi
Curiosamente, no processo de aminação (carbono 6) do IMP e consequente formação do
AMP consome-se “uma ligação rica em energia do GTP” (ver Equação 4 e Equação 6) enquanto no
processo de aminação (carbono 2) que origina o GMP se consomem “duas ligações ricas em energia
do ATP” (ver Equação 8 e Equação 9; notar que o PPi formado vai sofrer hidrólise subsequente).
7- Vias de salvação de bases púricas e de nucleosídeos
Para além de poderem ter origem na síntese de novo, os nucleotídeos púricos também podem
formar-se (1) a partir das bases guanina, hipoxantina e adenina ou (2) a partir dos nucleosídeos.
A síntese a partir das bases envolve transferência de ribose-5-fosfato do PRPP para as bases:
base púrica + PRPP → NMP + PPi. Nos mamíferos existem duas enzimas com este tipo de atividade:
a fosforibosil-transférase da guanina e da hipoxantina (ver Equação 10) e a fosforibosiltransférase da adenina (ver Equação 11).
Equação 10 guanina ou hipoxantina + PRPP → GMP ou IMP + PPi
Equação 11 adenina + PRPP → AMP + PPi
A síntese de nucleotídeos a partir dos nucleosídeos envolve a ação de diversas cínases dos
nucleosídeos que catalisam a transferência do fosfato terminal do ATP para o carbono 5’ do resíduo
de ribose (ou desoxiribose) dos diversos nucleosídeos levando à formação de nucleosídeos
monofosfato (ver Equação 12).
As reações catalisadas pelas fosforibosil-transférases e pelas cínases de nucleosídeos também
se designam por “vias de salvação” (ou “de recuperação”) pois permitem recuperar, respetivamente,
bases e nucleosídeos a nucleotídeos. Na ausência destas “vias de salvação” as bases guanina,
hipoxantina e xantina assim como todos os nucleosídeos (resultantes de nucleotídeos) continuariam o
seu processo catabólico com formação de ácido úrico que se perde na urina. Embora o processo de
síntese de novo de nucleotídeos púricos seja pouco ativo, os processos que levam à degradação destes
a nucleosídeos e às bases, assim como as "vias de salvação" são, em muitos tecidos, processos
importantes e com atividade elevada.
Equação 12 ATP + nucleosídeo → ADP + NMP
8- As cínases de nucleosídeos monofosfato e a cínase de nucleosídeos difosfato
Quer no caso dos nucleotídeos púricos quer no caso dos pirimídicos, a conversão dos
nucleotídeos que contêm um resíduo de fosfato (nucleosídeos monofosfato; NMP) em nucleosídeos
difosfato (NDP) e destes em nucleosídeos trifosfato (NTP) envolve a ação de cínases que catalisam a
transferência do fosfato terminal do ATP para os substratos aceitadores. A cínase de nucleosídeos
difosfato (ver Equação 13) é muito inespecífica quer relativamente à base, quer ao açúcar mas, na
conversão dos nucleosídeos monofosfato em nucleosídeos difosfato (ver Equação 14) intervêm
diversas cínases com diferentes especificidades; a cínase do adenilato (ver Equação 15) é um
exemplo deste tipo de cínases.
Página 3 de 12
Síntese e degradação de bases púricas e pirimídicas - Rui Fontes
Equação 13 ATP + NDP ↔ ADP + NTP
Equação 14 ATP + NMP ↔ ADP + NDP
Equação 15 ATP + AMP ↔ 2 ADP
Na síntese de ATP a partir de ADP tem particular relevância a síntase de ATP da membrana
interna da mitocôndria.
A diminuição do número de fosfatos dos nucleotídeos pode envolver múltiplas enzimas como,
por exemplo, fosfátases específicas e inespecíficas. A hidrólise dos nucleosídeos monofosfato é
catalisada por fosfátases que se designam de 5’-nucleotídases (ver Equação 16): a ação das 5’nucleotídases leva à formação de nucleosídeos.
Equação 16 NMP + H2O → nucleosídeo + Pi
9- A redução dos ribonucleosídeos difosfato pelas formas reduzidas da tioredoxina e
glutaredoxina
Os derivados 2’-desoxinucleotídeos (quer púricos quer pirimídicos) formam-se por ação da
redútase dos ribonucleosídeos difosfato e os agentes redutores diretos são duas proteínas: as formas
reduzidas da tioredoxina e da glutaredoxina (ver Equação 17).
A manutenção da tioredoxina e da glutaredoxina nas formas reduzidas depende, em última
instância, do NADPH e da ação catalítica de oxi-redútases específicas. Na ação da redútase da
tioredoxina o redutor direto é o NADPH (ver Equação 18) mas, na ação da redútase da
glutaredoxina, o redutor direto é o glutatião (ver Equação 19). A redução do GSSG formado aquando
da ação da redútase da glutaredoxina é catalisada pela redútase do glutatião, uma enzima em que o
agente redutor é o NADPH (ver Equação 20).
Equação 17 NDP + tioredoxina reduzida ou glutaredoxina reduzida →
2’-dNDP + tioredoxina oxidada ou glutaredoxina oxidada
Equação 18 tioredoxina oxidada + NADPH → tioredoxina reduzida + NADP+
Equação 19 glutaredoxina oxidada + 2 GSH → glutaredoxina reduzida + GSSG
Equação 20 GSSG + NADPH → 2 GSH + NADP+
A redução dos NDPs ocorre no hidroxilo 2’. São substratos da redútase dos ribonucleosídeos
difosfato o ADP, o GDP, o UDP e o CDP. Como será explicado adiante, os nucleotídeos da timina
(TMP, TDP e TTP) formam-se a partir do 2’-dUDP ou do 2’-dCDP. Quando não se especifica o
contrário subentende-se que a pentose da timidina e dos nucleotídeos da timina é a 2’-desoxiribose;
nos outros casos subentende-se que é a ribose.
10- O catabolismo dos nucleotídeos púricos
No processo catabólico, os nucleosídeos monofosfato (NMP) sofrem hidrólise na ligação
fosfoéster (5’-nucleotídase) formando-se os respetivos nucleosídeos (ver Equação 16); a partir do
AMP forma-se adenosina e a partir do GMP, guanosina. As vias catabólicas seguidas pela adenosina e
pela guanosina são algo distintas mas, em ambos os casos, o produto final é o urato, uma substância
em que o anel purina se mantém intacto e que é excretada na urina.
11- A conversão de AMP em ácido úrico
A adenosina formada pela ação da 5’-nucleotídase é desaminada a inosina por ação catalítica
de uma hidrólase: a desamínase da adenosina (ver Equação 21). De facto, a conversão do AMP em
inosina também pode seguir uma sequência em que o primeiro passo é a desaminação (catalisada pela
desamínase do AMP; ver Equação 22) e o segundo a hidrólise (ver Equação 23) do fosfoéster do IMP,
o nucleotídeo formado pela desamínase do AMP. Ou seja, na conversão de AMP em inosina, a
sequência de reações pode seguir um caminho em que primeiro ocorre a hidrólise do grupo fosfato
(ver Equação 16) e depois a desaminação hidrolítica do grupo 6-amina (ver Equação 21) ou o inverso
(ver Equação 22 e Equação 23).
Página 4 de 12
Síntese e degradação de bases púricas e pirimídicas - Rui Fontes
Equação 21 adenosina + H2O → inosina + NH4+
Equação 22 AMP + H2O → IMP + NH4+
Equação 23 IMP + H2O → inosina + Pi
A inosina formada perde a pentose e gera hipoxantina por ação de uma fosforílase de
nucleosídeos; ou seja, a fosforílase catalisa a fosforólise da inosina ficando o fosfato que reagiu
ligado ao carbono 1 da ribose e libertando-se a base hipoxantina (ver Equação 24).
Equação 24 inosina + Pi → hipoxantina + ribose-1-fosfato
As oxidações da hipoxantina a xantina e da xantina a ácido úrico são da responsabilidade de
uma mesma enzima: a oxiredútase da xantina. De facto, o produto do gene codificador da enzima é
uma desidrogénase dependente do NAD+ (desidrogénase da xantina; ver Equação 25 e Equação 26)
que se pode converter numa oxídase (oxídase da xantina; ver Equação 27 e Equação 28).
Aparentemente, in vivo, podem coexistir as duas formas da enzima [2].
Equação 25
Equação 26
Equação 27
Equação 28
hipoxantina + NAD+ → xantina + NADH
xantina + NAD+ → urato + NADH
hipoxantina + O2 → xantina + H2O2
xantina + O2 → urato + H2O2
A ribose-1-fosfato formada aquando da ação da fosforílase pode sofrer isomerização a ribose5-fosfato (ver Equação 29).
Equação 29 ribose-1-fosfato ↔ ribose-5-fosfato
Assim, a formação de urato a partir de AMP pode ocorrer via
AMP → adenosina → inosina → hipoxantina → xantina → urato ou via
AMP → IMP → inosina → hipoxantina → xantina → urato.
12- A conversão da guanosina em ácido úrico
No processo catabólico da guanosina (formada pela ação da 5’-nucleotídase; ver Equação
16) há, relativamente ao caso da adenosina, uma inversão na sequência das reações e forma-se
diretamente xantina em vez de hipoxantina. Aqui é a própria guanosina que sofre fosforólise gerando
a guanina (ver Equação 30) e é a base (e não a guanosina ou o GMP) que sofre desaminação
hidrolítica (guanase ou desamínase da guanina; ver Equação 31). Da ação da guanase (ver Equação
31) resulta a formação da xantina que, por ação da oxiredútase da xantina, origina o urato (ver
Equação 26 e Equação 28). Assim, a formação de urato a partir de GMP ocorre via GMP →
guanosina → guanina → xantina → urato.
Equação 30 guanosina + Pi → guanina + ribose-1-fosfato
Equação 31 guanina + H2O → xantina + NH4+
13- Causas de hiperuricemia e a gota
O ácido úrico é excretado na urina1. À condição em que, quer por excesso de formação, quer
por défice de excreção (mais frequente) a sua concentração aumenta no plasma dá-se o nome de
hiperuricemia.
Uma das causas possíveis de hiperuricemia é o aumento da velocidade de destruição celular
como a que ocorre em neoplasias durante a quimioterapia ou a radioterapia. Uma causa rara de
hiperuricemia é o síndrome de Lesch-Nyhan que se deve a alterações no gene que codifica a
1
O ácido úrico constitui sempre uma pequena percentagem do azoto excretado. A quantidade de ácido úrico
excretado é de cerca de 4 mmol/dia (o que corresponde a 16 mmol de N /dia). Se admitirmos balanço azotado nulo e
uma ingestão de 100 g diárias de proteínas a massa de azoto excretado será de 16 g/dia (pouco mais de 1 mole de N)
e a percentagem do azoto total excretado que sai na forma de ácido úrico seria inferior a 2 % [0,016 mol / (16g/14g)
mol = 1,4 %].
Página 5 de 12
Síntese e degradação de bases púricas e pirimídicas - Rui Fontes
fosforibosil-transférase da hipoxantina e guanina (ver Equação 10). Nesta doença, a baixa atividade
da enzima prejudica a via de salvação das bases púricas hipoxantina e guanina havendo excesso de
produção de ácido úrico e alterações neurológicas de patogenia desconhecida (que incluem atraso
mental e comportamentos anormais de automutilação).
Na ausência de qualquer patologia, a concentração normal de urato no plasma encontra-se
perto do limite de solubilidade. Quando, devido a concentração elevada, o ácido úrico precipita na
sinovial de articulações pode haver uma resposta inflamatória que provoca dor; esta condição
patológica denomina-se gota. O tratamento da gota e das hiperuricemias de qualquer causa pode
incluir a administração de alopurinol, um fármaco que inibe a oxiredútase da xantina (ver Equações
25-28). Aquando da administração de alopurinol, as concentrações plasmáticas de hipoxantina e de
xantina aumentam anormalmente mas, porque são mais solúveis que o urato, não precipitam.
14- A síntese de novo das pirimidinas
A via da síntese de novo dos nucleotídeos pirimídicos envolve, como primeiro passo, uma
sintétase de carbamil-fosfato citoplasmática (sintétase de carbamil-fosfato II) em que o dador de
azoto é a glutamina (ver Equação 32). A segunda reação é catalisada pela transcarbamílase do
aspartato (ver Equação 33). O carbamil-aspartato sofre a ação de uma desidrátase formando o
dihidro-orotato que é o primeiro composto cíclico da via metabólica (ver Equação 34).
O dihidro-orotato é substrato da desidrogénase do dihidro-orotato que catalisa a sua
oxidação a orotato (ver Equação 35). A desidrogénase do dihidro-orotato é uma enzima que contém
FMN como grupo prostético e que se situa na face externa da membrana interna da mitocôndria tendo
o centro ativo voltado para o espaço intermembranar [3]. Os eletrões que resultam da oxidação do
dihidro-orotato, via FMN, reduzem a ubiquinona a ubiquinol e via complexos III e IV da cadeia
respiratória acabam por reduzir o oxigénio a H2O.
O orotato formado vai reagir com o PRPP gerando o primeiro nucleotídeo desta via
metabólica: o orotidilato (OMP); a reação é catalisada pela fosforibosil-transférase do orotato (ver
Equação 36). O OMP por descarboxilação gera o UMP (ver Equação 37).
Equação 32
Equação 33
Equação 34
Equação 35
Equação 36
Equação 37
CO2 + glutamina + 2 ATP → carbamil-fosfato + 2 ADP + Pi + glutamato
carbamil-fosfato + aspartato → carbamil-aspartato + Pi
carbamil-aspartato → dihidro-orotato + H2O
dihidro-orotato + ubiquinona → orotato + ubiquinol
orotato + PRPP → OMP + PPi
OMP → UMP + CO2
Por ação catalítica de uma cínase o UMP pode ser fosforilado a UDP (ver Equação 14) que
está na origem quer do CTP, quer do TMP.
Com exceção da desidrogénase do dihidro-orotato todas as enzimas da via de síntese dos
nucleotídeos pirimídicos se situam no citoplasma das células.
Os átomos N1, C4, C5 e C6 do anel pirimidina têm origem no aspartato; o átomo N3 tem
origem no grupo amida da glutamina e o átomo C2, no CO2. A equação soma relativa à síntese do
UDP (síntese de PRPP incluída; ver Equação 1) é a Equação 38. As 4 moléculas de ATP que se
consomem na síntese de uma molécula de UDP a partir de ribose-5-fosfato, CO2, glutamina e
aspartato são utilizadas nas ações catalíticas da síntase do PRPP (ver Equação 1), da sintétase de
carbamil-fosfato II (ver Equação 32; 2 ATPs) e da cínase do UMP (ver Equação 14). (Uma molécula
de CO2 é consumida na ação da sintétase de carbamil-fosfato II, mas forma-se outra na
descarboxilação do OMP).
Equação 38 ribose-5-fosfato + glutamina + aspartato + 4 ATP + ubiquinona →
UDP + glutamato + AMP + PPi + 3 ADP + 2 Pi + ubiquinol
15- A síntese de CTP, de UTP e do 2’-desoxiuridilato a partir de uridina difosfato
(i) O CTP forma-se por aminação do carbono 4 do UTP por ação da sintétase do CTP (o
dador do grupo amina é a glutamina que sai como glutamato; ver Equação 39). O UTP, que é
substrato desta sintétase, forma-se por fosforilação do UDP (ação da cínase de nucleosídeos difosfato;
ver Equação 13).
Página 6 de 12
Síntese e degradação de bases púricas e pirimídicas - Rui Fontes
(ii) O TMP forma-se por metilação do carbono 5 do 2’-dUMP e o 2’-dUMP também tem
origem no UDP. Por ação da redútase dos ribonucleosídeos difosfato (ver Equação 17) o UDP
converte-se em 2’-dUDP que, por ação da cínase de nucleosídeos difosfato, origina o 2’-dUTP
(Equação 13). O 2’-dUMP pode formar-se a partir do 2’-dUTP por ação catalítica de uma hidrólase
específica que atua na ligação entre os fosfatos α e β do 2’-dUTP (ver Equação 40).
Uma outra via metabólica que também se inicia no UDP e origina 2’-dUMP tem como
intermediário o CTP que foi formado na ação catalítica da sintétase do CTP (ver Equação 39). O CTP
é desfosforilado a CDP, o CDP é reduzido a 2’-dCDP pela ação da redútase dos ribonucleosídeos
difosfato (ver Equação 17) e o 2’-dCDP é desfosforilado a 2’-dCMP que, por ação da desamínase do
2’-dCMP, sofre desaminação hidrolítica no carbono 4 originando o 2’-dUMP (ver Equação 41).
Equação 39 UTP + glutamina + ATP → CTP + glutamato + ADP + Pi
Equação 40 2’-dUTP + H2O → 2’-dUMP + PPi
Equação 41 2’-dCMP + H2O → 2’-dUMP + NH4+
Assim, dependendo das enzimas envolvidas, as sequências que levam à formação de 2’dUMP a partir de UDP podem ser
UDP → 2’-dUDP → 2’-dUTP → 2’-dUMP ou
UDP → UTP → CTP → CDP → 2’-dCDP → 2’-dCMP → 2’-dUMP.
16- A síntese do timidilato a partir de 2’-desoxiuridilato e o ciclo do dihidrofolato
A reação de conversão do 2’-dUMP em TMP é catalisada pela síntase do timidilato (ou
síntase do TMP) e consiste na metilação do 2’-dUMP pelo N5,N10-metileno-H4-folato (ver Equação
42). Nesta reação, ao contrário do que acontece na generalidade das reações de transferência de
unidades monocarbonadas que envolvem o ácido fólico, o produto da reação não é o H4-folato, mas o
dihidrofolato (H2-folato).
Ao contrário do H4-folato, o H2-folato não é aceitador de unidades monocarbonadas; por
isso, a manutenção do processo metabólico exige a redução do H2-folato a H4-folato que ocorre à
custa da oxidação do NADPH e é catalisada pela redútase do dihidrofolato (ver Equação 43). O
N5,N10-metileno-H4-folato, dador do grupo metilo na síntese do TMP, pode formar-se por ação de
diversas enzimas onde se destacam a hidroxi-metil-transférase da serina (ver Equação 44) e a enzima
de clivagem da glicina (ver Equação 45). É costume agrupar as reações em que o N5,N10-metilenoH4-folato se converte em H2-folato (e o 2’-dUMP gera timidilato, ver Equação 42), o H2-folato se
reduz a H4-folato (ver Equação 43) e este último é novamente metilado a N5,N10-metileno-H4-folato
(ver Equação 44 e Equação 45) sob a denominação de ciclo do dihidrofolato (N5,N10-metileno-H4folato → H2-folato → H4-folato → N5,N10-metileno-H4-folato).
Equação 42
Equação 43
Equação 44
Equação 45
2’-dUMP + N5,N10-metileno-H4-folato → TMP + H2-folato
H2-folato + NADPH → H4-folato + NADP+
serina + H4-folato ↔ glicina + N5,N10-metileno-H4-folato
glicina + NAD+ + H4-folato → NH3 + CO2 + N5,N10-metileno-H4-folato + NADH
17- A fosforibosil-transférase do uracilo e as cínases de nucleosídeos pirimídicos
Tal como no caso dos nucleosídeos das purinas também os nucleosídeos das pirimidinas
(contendo diferentes bases e ribose ou desoxiribose) podem ser “salvos” por ação de cínases de
nucleosídeos (ver Equação 12).
A “salvação” das bases pirimidinas (não ligadas à ribose ou à desoxiribose) é um processo
pouco ativo mas, à semelhança do que acontece no caso das bases púricas adenina, guanina e
hipoxantina, também o uracilo pode ser “salvo” por ação da fosforibosil-transférase do uracilo (ver
Equação 46). (O orotato é, como já referido, um intermediário da via da síntese de novo das
pirimidinas e a fosforibosil-transférase do orotato participa nessa via; ver Equação 36).
Equação 46 uracilo + PRPP → UMP + PPi
Página 7 de 12
Síntese e degradação de bases púricas e pirimídicas - Rui Fontes
18- O catabolismo dos nucleosídeos pirimídicos e das bases pirimídicas
O catabolismo dos nucleotídeos pirimídicos envolve a prévia libertação das bases (citosina,
uracilo e timina) por ação sequenciada de fosfátases que atuam nos nucleotídeos (5’-nucleotídase
incluída; ver Equação 16) formando os nucleosídeos correspondentes e de fosforílases que atuam nos
(2’-desoxi)nucleosídeos catalisando a sua fosforólise (com libertação das bases correspondentes e de
(2-desoxi)ribose-1-fosfato; ver Equação 47).
Equação 47 citidina ou uridina ou timidina + Pi →
citosina ou uracilo ou timina + ribose-1-fosfato ou 2’-desoxiribose-1-fosfato
No caso da citidina e da citosina o catabolismo envolve a desaminação hidrolítica formandose, respetivamente, uridina e uracilo (ver Equação 48).
Equação 48 citidina (ou citosina) + H2O → uridina (ou uracilo) + NH4+
No catabolismo das bases pirimídicas, ao contrário do que acontece no caso das bases púricas,
o anel sofre rotura formando-se CO2, amónio e β-aminoácidos que podem sofrer a ação de enzimas
catabólicas (convertendo-se em CO2, amónio e/ou ureia) ou serem parcialmente excretados intactos
na urina; no caso da timina o β-aminoácido formado é o β -aminoisobutirato. A Equação 49 e a
Equação 50 são equações soma relativas aos catabolismos do uracilo e da timina. É de notar que,
embora se tratem de vias catabólicas, ocorre, durante o processo, um passo redutor em que se
consome NADPH. O amónio é transformado em ureia mas, dada a baixa taxa no metabolismo dos
nucleotídeos relativamente à dos aminoácidos, o seu contributo para esta formação é relativamente
pequeno.
Equação 49 uracilo + NADPH + 2 H2O → NADP+ + alanina-β + CO2 + NH4+
Equação 50 timina + NADPH + 2 H2O → NADP+ + β-aminoisobutirato + CO2 + NH4+
19- A regulação da síntese de nucleotídeos púricos e pirimídicos
As atividades das enzimas que catalisam as vias de síntese de novo e de recuperação
(“salvação”) das bases púricas e pirimídicas têm mecanismos de regulação de que os mais estudados
são os mecanismos alostéricos.
(i) Uma reação comum às vias metabólicas de síntese de novo dos nucleotídeos púricos e
pirimídicos é a síntese do PRPP (ver Equação 1). Esta reação é um dos pontos de controlo da
velocidade de síntese de novo dos nucleotídeos púricos e pirimídicos, mas também nas vias de
“salvação” que envolvem fosforibosil-transférases (o PRPP é o substrato dador de ribose-5-fosfato na
ação destas enzimas). A síntase do PRPP é inibida por nucleotídeos quer púricos, quer pirimídicos.
(ii) Na via de síntese de novo das purinas existem outros passos de regulação de que
destacamos os catalisados pela amido-fosforibosil-transférase da glutamina (ver Equação 2), pela
sintétase de adenilosuccinato (Equação 4) e pela desidrogénase do IMP (ver Equação 7).
A amido-fosforibosil-transférase da glutamina, a primeira enzima específica do processo de
síntese das purinas, é inibida pelos nucleotídeos púricos e estimulada pelo PRPP. O PRPP é substrato
da enzima, mas o valor do seu Km é superior ao das suas concentrações fisiológicas; por isso, a
amido-fosforibosil-transférase da glutamina é sensível às variações fisiológicas da concentração do
PRPP.
No processo de síntese de AMP a partir de IMP a “primeira” enzima (a sintétase de
adenilosuccinato; ver Equação 4) é inibida pelo produto final, o AMP. De modo semelhante, no
processo de síntese de GMP a partir de IMP, a “primeira” enzima (a desidrogénase do IMP; ver
Equação 7) é inibida pelo GMP.
(iii) Na via da síntese das pirimidinas são de destacar os passos catalisados pela sintétase de
carbamil-fosfato II (ver Equação 32) e pela transcarbamílase do aspartato (ver Equação 33). A
sintétase de carbamil-fosfato II (ver Equação 32) é ativada pelo PRPP e inibida pelo UTP e a
transcarbamílase do aspartato (ver Equação 33) é ativada pelo ATP e inibida pelo CTP.
Página 8 de 12
Síntese e degradação de bases púricas e pirimídicas - Rui Fontes
20- A regulação da redútase dos ribonucleosídeos difosfato
A redútase dos ribonucleosídeos difosfato só está ativa aquando da fase S do ciclo celular
(síntese de DNA). A sua atividade é regulada de forma muito complexa envolvendo ações inibidoras
e ativadoras dos diversos nucleotídeos (quer contendo ribose, quer já contendo desoxiribose)
relativamente às reações de redução dos diferentes nucleosídeos difosfato que são substratos da
enzima. Esta regulação faz com que o consumo dos diferentes 2’-desoxiribonucleosídeos trifosfato
(2’-dNTP) na síntese de DNA seja compensada pela síntese e que não haja nem excesso nem défice
de nenhum dos 2’-dNTPs.
21- A importância da glutamina quer como substrato na síntese de nucleotídeos quer como
nutriente nas células em proliferação rápida
As células em multiplicação rápida como são as células das criptas das vilosidades intestinais,
os linfócitos em fase de proliferação (em resposta a desafios de natureza imunológica), as células
precursoras dos eritrócitos e dos leucócitos na medula óssea, assim como as células da raiz dos
cabelos têm atividades de síntese de nucleotídeos púricos e pirimídicos muito elevadas. Estas sínteses
sustentam as atividades das polimérases de DNA e RNA. Nestas células a síntese proteica também é
elevada e tudo isto contribui para explicar a sua elevada despesa energética.
Nestes tipos celulares, os principais nutrientes são a glutamina2 e a glicose3 que sustentam o
alto consumo energético. No entanto, algumas das moléculas de glicose e de glutamina que são
consumidas nestas células têm por destino a síntese de nucleotídeos. A glicose, através da via das
pentoses, leva à formação de ribose-5-fosfato que é usada na síntese de PRPP (ver Equação 1) e a
glutamina é substrato de várias enzimas, quer na síntese de nucleotídeos púricos (ver Equação 2 e
Equação 8), quer na de nucleotídeos pirimídicos (ver Equação 32 e Equação 39).
22- Fármacos inibidores da síntese de nucleotídeos
As células em multiplicação rápida são sensíveis a fármacos que interferem na multiplicação
celular; por isso, estes fármacos são usados na quimioterapia de neoplasias ou como
imunossupressores. Nalguns destes fármacos o seu mecanismo de ação é a interferência no
metabolismo das purinas e das pirimidinas.
(i) O mais conhecido é o metotrexato cuja ação é a inibição da redútase do dihidrofolato (ver
Equação 43) desta forma bloqueando o processo de reaproveitamento do dihidrofolato para a síntese
do TMP. O metotrexato é um análogo estrutural do N5,N10-metileno-H4-folato e liga-se à redútase do
dihidrofolato no centro ativo da enzima.
(ii) Um outro exemplo é a 6-mercaptopurina que, de facto, é um pró-fármaco: só tem ação
depois de convertida no respetivo nucleosídeo monofosfato por ação da fosforibosil-transférase da
hipoxantina e guanina (ver Equação 10). O ribonucleosídeo monofosfato da 6-mercaptopurina é um
potente inibidor de, pelo menos, três enzimas da via de síntese das purinas nomeadamente: a amidofosforibosil-transférase da glutamina (ver Equação 2), a sintétase de adenilosuccinato (Equação 4) e a
desidrogénase do IMP (ver Equação 7). O ribonucleotídeo da 6-mercapotopurina tem analogias
estruturais com substratos destas enzimas: o PRPP no caso da amido-fosforibosil-transférase da
glutamina e o IMP nos outros dois casos.
23- Ação de enzimas do metabolismo dos nucleotídeos no efeito terapêutico do aciclovir
Alguns antivirais também são pró-fármacos. O aciclovir, usado no tratamento do herpes, é
um análogo da guanosina em que o resíduo de (desoxi)ribose está substituído por um outro resíduo
que não têm um grupo hidroxilo numa posição correspondente ao 3’-OH da (desoxi)ribose.
2
No caso da glutamina a oxidação pode ser incompleta envolvendo a sua conversão em aspartato, em alanina ou em
lactato. Em todos estes casos o primeiro passo no catabolismo da glutamínase consiste na ação da glutamínase que
catalisa a formação do glutamato. Outros passos comuns envolvem a ação da desidrogénase do glutamato (ou de
uma transamínase), da desidrogénase α-cetoglutarato, da sintétase do succinil-CoA, da desidrogénase do succinato,
da fumárase e da desidrogénase do malato o que explica a síntese de ATP (via cadeia respiratória e síntase de ATP)
e a produção de CO2. O aspartato pode formar-se por ação da transamínase do aspartato usando como substratos
glutamato e oxalacetato (produto da ação da desidrogénase do malato). A formação de piruvato resulta ou da ação
da enzima málica no malato ou das ações sequenciadas da carboxicínase do fosfoenolpiruvato (no oxalacetato) e da
cínase do piruvato (no fosfoenolpiruvato). A alanina pode formar-se por ação da transamínase do piruvato e o
lactato por ação da desidrogénase do lactato: em ambos há conversão do piruvato formado.
3
Nestas células grande parte da glicólise é de tipo anaeróbica havendo, por isso, uma produção elevada de lactato.
Página 9 de 12
Síntese e degradação de bases púricas e pirimídicas - Rui Fontes
A ação terapêutica do aciclovir só se pode exercer depois deste composto sofrer fosforilação a
aciclovir monofosfato e esta fosforilação só ocorre nas células infetadas pelo vírus: a cínase que
catalisa esta reação é a cínase de timidina codificada pelo genoma viral. O processo só ocorre nas
células infetadas porque a cínase de timidina do hospedeiro não reconhece o aciclovir.
O aciclovir monofosfato formado é, de seguida, fosforilado por cínases do hospedeiro em
aciclovir trifosfato que se incorpora no DNA do vírus mas, porque o aciclovir não tem um hidroxilo
na posição 3’, esta incorporação provoca a terminação da síntese do DNA viral.
1. Berthold, H. K., Crain, P. F., Gouni, I., Reeds, P. J. & Klein, P. D. (1995) Evidence for incorporation of intact dietary
pyrimidine (but not purine) nucleosides into hepatic RNA, Proc Natl Acad Sci U S A. 92, 10123-7.
2. Nishino, T., Okamoto, K., Eger, B. T. & Pai, E. F. (2008) Mammalian xanthine oxidoreductase - mechanism of transition
from xanthine dehydrogenase to xanthine oxidase, Febs J. 275, 3278-89.
3. Rawls, J., Knecht, W., Diekert, K., Lill, R. & Loffler, M. (2000) Requirements for the mitochondrial import and localization
of dihydroorotate dehydrogenase, Eur J Biochem. 267, 2079-87.
Página 10 de 12
Síntese e degradação de bases púricas e pirimídicas - Rui Fontes
Página 11 de 12
Síntese e degradação de bases púricas e pirimídicas - Rui Fontes
Página 12 de 12