PLANOS GERAIS DE ORGANIZAÇÃO DOS SERES VIVOS
Teoria da Célula
A Teoria da Célula, ou Celular, foi proposta no século XIX, pouco antes da formulação
da Teoria da Evolução (1859), por Darwin, e da proposição das Leis da Hereditariedade por
Mendel (1865). A teoria da Célula foi formulada em 1838 por Theodor Schwann e Mathias
Schleiden. As principais conclusões de um livro escrito por Schwann, no ano seguinte, no qual
Schleiden não é citado, são:
A célula é a unidade de estrutura, fisiologia e organização de todas as coisas vivas.
A célula tem dupla existência, como entidade distinta e como elemento fundamental
de construção dos organismos.
As células se formam de material acelular, de maneira similar à formação dos
cristais (geração espontânea)
Por esta época os experimentos de Needham e Spallanzani, que levavam à hipótese de um
força vital, que gerava seres vivos da matéria não viva, não haviam sido contestados e a
teoria celular obviamente tinha de incorporar a geração espontânea. Se a célula era a
unidade de estrutura e fisiologia dos sere vivos e se estes podiam se originar da matéria não
viva, de fato quem se originava da matéria não viva eram as células
Em 1858 Rudolf Virchow corrigiu a proposta de Teoria Celular declarando “Omnis
cellula e cellula”: todas as células se originam de células pré-existentes. Em 1860 Pasteur, em
experimentos simples de esterilização, demonstrou que a geração espontânea de seres vivos
não ocorria, deitando por terra a hipótese de uma força vital.
De justiça, a Teoria Celular, é atribuída aos três, Shwann, Schleiden e Virchow.
Compõem-na, na formulação moderna, os seguintes princípios:
Todas as coisas vivas são formadas por células.
A célula é a unidade funcional & estrutural das coisas vivas.
Todas as células provêm de células pré-existentes, por divisão destas (a geração
espontânea não ocorre).
As células contém a informação hereditária, que é passada de célula a célula na
divisão celular.
Todas as células têm basicamente a mesma composição química.
Todo o fluxo de energia da vida (metabolismo e bioquímica) ocorre dentro das
células.
Embora a geração espontânea de células não ocorra nas condições atuais do planeta, ela
ocorreu em passado remoto, a cerca de 4 bilhões de anos e se estendeu por algo em torno de
2 bilhões de anos, para gerar os padrões celulares contemporâneos – procariótico e
eucariótico – que formam todos os seres vivos.
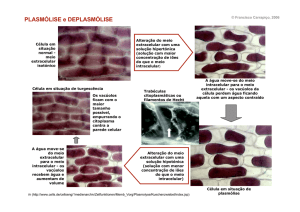
Padrões celulares contemporâneos.
Entre os seres vivos contemporâneos a diversidade fenotípica das células é enorme.
As células têm formas, tamanhos e funções muito variados. Contudo, há entre elas
semelhanças suficientes para classificá-las em dois grandes padrões: o procariótico e o
eucariótico. O padrão procariótico é de células pequenas, bastante menores, que as do
padrão eucariótico, como se vê na figura 1. As células procarióticas não têm organelas,
1
delimitadas por membranas. O material genético está em um cromossoma único, circular,
livre no citoplasma. Reações químicas vetorais, que ocorrem em membranas, se dão na
membrana celular. As células eucarióticas são maiores, mesmo as menores entre elas são
bem maiores que as células procarióticas. Caracteristicamente têm organelas, delimitadas
por membranas, com funções específicas. O material genético, reunido em múltiplos
cromossomas, está no núcleo, segregado do citoplasma pela membrana nuclear. No
protoplasma há mitocôndrias, cuja função precípua é a conversão de energia química, por
oxidação de compostos reduzidos, resultando, finalmente, na síntese de ATP. As
mitocôndrias ainda armazenam o Ca2+. Há ainda retículo endoplasmático, no qual se dá a
síntese de proteínas e armazenamentos, aparelho de Golgi, lisossomos, vesículas de
estocagem, e outras organelas.
Figura 1: Células procariótica, à esquerda, e eucariótica, à direita. Há, na eucariótica
variados tipos de organelas, entre elas o núcleo, que é a maior. Para se comparar
tamanhos, as escalas devem ser consideradas. As figuras são, ambas, micrografias
eletrônicas.
Todos os organismos pluricelulares (ou multicelulares), animais ou plantas, são
formados por células do tipo eucariótico. Não se sabe de um só que seja formado por células
do padrão procariótico. A tabela I contém informações sobre os vários reinos em que se
agrupam os seres vivos do planeta. De particular interesse ao que aqui se trata são os reinos
Plantae e Animalia. Nestes, células eucarióticas se diferenciam para formar tecidos, estes
formam órgãos, que formam sistemas de órgãos. Cada célula autossômica tem a informação
genética completa da espécie. A expressão diferenciada de genes gera o fenótipo celular, no
processo de diferenciação que leva à formação do organismo.
2
Tabela I: Reinos Biológicos
Tipo celular
Procariótico
Eucariótico
grandes
e
complexas.
Evoluíram
como
simbiontes
de
procariotos
Eucariótico
Organizado
em
tecidos, órgãos e
sistemas
Autotrófico
Quimiosintético
Reino Archea
Três linhagens,
todas
anaeróbicas
Nenhum reino
evoluiu com a
característica
Heterotrófico
Fotosintético
Reino Eubacteria
Bactérias
aeróbicas,
fotosintéticas.
Antigamente
chamdas
algas
azul-verdes
Reino Protista
Algas unicelulares
verdes, douradas,
vermelhas
e
marrons
Móveis
Reino Eubacteria
Espiroquetas
aeróbicas
Sésseis
Reino Eubactéria.
Bactérias aeróbicas
com esporos
Reino Protista
Protozoários,
amebas,
ciliados,
flagelados
Reino Protista
Esporozoários,
o plasmódio
Reino Plantae
Angiospermas,
ginospermas,
fungos, líquens
Reino Animalia
Espongiários,
celetenterados,
vermes chatos e
cilíndricos, moluscos
,
anelídos,
artrópodes,
equinodermatas,
cordados
Reino Fungi
ascomicetos,
basidiomicetos,
ficomicetos e fungos
imperfeitos
como
Reino Fungi
Levedura
Organização estrutural dos seres multicelulares
Se em meados do séculos XIX se estabelecia a teoria celular, estatuindo que a célula
é a unidade estrutural e funcional dos seres vivos e que o fluxo de energia e das reações
metabólicas ocorre nas células, a imediata conseqüência da concepção é que os organismos
pluricelulares são o resultado dos processos das suas muitas células, não raramente bilhões,
operando integradamente. Como os processos característicos de uma célula são o que se
pode chamar a vida, esta persiste enquanto os processos ocorrerem. Obviamente estes
mesmos processos – reações químicas – demandam suprimento contínuo de energia, que nos
seres heterotróficos, advêm da oxidação de compostos reduzidos, demandam os elementos
para a síntese das proteínas, que são os aminoácidos, etc, etc. Os mesmos processos
produzem gás carbônico, uréia e outros produtos. Portanto, as células que são a sede destes
processos, devem dispor de um volume adjacente de onde retirem os compostos para os seus
processos e onde despejem os que o metabolismo produziu.
Em 1848 Claude Bernard propôs o conceito de meio interno ou interior. Todos os
organismos multicelulares conteriam em si, além do volume das suas células, um volume
interno, cujas características químicas e físicas seriam mantidas relativamente constantes
pelas grandes funções fisiológicas.
A concepção sintética e contemporânea de um organismo multicelular é a da figura 2.
3
Figura 2: Representação do organismo. Os volumes somados das células forma o
compartimento intracelular. Entre as células está o espaço intersticial. O sistema
circulatório, um extenso conjunto de vasos, contém o plasma. Que constitui o
compartimento plsmático. O volume das células sangüíneas é contado como volume
intracelular. As bombas cardíacas movem o sangue, continuamente, entre os capilares
dos tecidos e as interfaces do organismo com o meio exterior: alvéolos pulmonares,
nefros, e epitélio do trato gastrintestinal.
O organismo é formado por células, que se diferenciam em vários fenótipos. Estes
formam tecidos, os tecidos formam órgão que constituem os sistemas de órgãos. Cada célula,
individualizada pela sua membrana plasmática, está imersa em um pequeno volume de solução,
com o qual a célula troca continuamente espécies químicas. Em um tecido estes volumes
formam um labirinto, entre as células. O conjunto destes espaços adjacentes à célula
constitui nos organismos o comportimento ou volume intersticial. O volume intersticial
correspondente a cada célula, no organismo, é exíguo e a sua composição seria rapidamente
alterada pelas trocas com as células. No longo processo evolutivo dos seres vivos,
desenvolveu-se um sofisticado sistema circulatório, desenhado para prover um fluxo
contínuo de sangue pelos capilares que formam rede extensa, em contacto com o espaço
intersticial. Processos difusionais exclusivamente promovem a troca de compostos químicos
entre o sangue que circula nos capilares e o compartimento intersticial das células.
Impulsionado pelas bombas cardíacas, o sangue circula continuamente dos capilares dos
tecidos aos capilares dos sistemas fisiológicos que estabelecem um contato com o meio
externo do organismo. Os pulmões constituem uma dessas interfaces para a troca de gases.
Os rins formam a interface para a excreção controlada de produtos químicos não voláteis,
entre eles água, eletrólitos, ácidos, produtos do metabolismo do nitrogênio, etc. O trato
gastrointestinal é a interface sofisticada para a captação de nutrientes que, coletados
seletivamente no meio externo, são, alguns, transformados pelo processo digestivo que
ocorre na luz dos órgãos, ainda meio externo.
O espaço ocupado pelo plasma, no compartimento vascular, e o espaço intersticial
constituem o compartimento extracelular.
4
Os compartimento do organismo: barreiras.
Em síntese, há dois grandes compartimentos: o intracelular e o extracelular. A
arquitetura deles é complexa e não são contínuos. Todavia, considerá-los não se constitui em
mero exercício abstrato ou pedagógico. O compartimento extracelular se subdivide em
compartimento intersticial, adjacente às células, e o compartimento vascular ou plasmático.
Talvez se deva preferir a designação plasmático. No interior do sistema vascular estão, de
fato, o plasma e as células sangüíneas, eritrócitos, leucócitos e plaquetas. O volume destas
está computado como volume celular. Separando o compartimento celular do intersticial está
a membrana celular plasmática. Esta constitui estrutura funcionalmente complexa, com
permeabilidade difusional, carregadores, receptores, canais, propriedades elétricas, e
constitui principal tema deste curso. Entre os compartimentos plasmático e o intersticial,
separando-os, está o endotélio vascular. O endotélio é formado por um contínuo de células,
com poros, pelos quais moléculas inorgânicas e orgânicas de baixo peso molecular podem se
movimentar difusionalmente. Os poros restringem o fluxo de proteínas.
Há, no organismo, compartimentos especiais, delimitados por epitélios que secretam
os fluídos que preenchem estes compartimentos. São estes os humores do globo ocular, o
compartimento sinovial, das cartilagens, o espaço pleural e o pericárdico.
Entre o organismo e o meio externo, nas estruturas especiais para trocas, como nos
pulmões, rins e trato gastrintestinal, há um epitélio monoestraficado. Na superfície do
corpo, a pele separa o organismo do meio externo. Estruturalmente complexa, a pele limita a
perda de água e é um órgão de termoregulação.
O compartimento plasmático é o de acesso menos traumático, por punção de uma veia
periférica. tanto para a introdução no organismo de compostos com finalidade terapêutica
como para a análise de composição do meio interno, com propósitos de diagnose.
A figura 3 ilustra as barreiras entre os compartimentos.
Figura 3: Barreiras entre os compartimentos
intersticial e intracelular, que é a membrana celular
plasmática (acima) e entre o compartimento intersticial e
o plasmático, que é o endotélio vascular (à esquerda).
5
Os compartimentos do organismo: volume e composição.
As soluções que preenchem os compartimentos têm a água como solvente e são
relativamente diluídas, com osmolaridade de cerca de 290 mOsmóis/l. O compartimento
intracelular, com concentração mais alta de proteínas, algumas delas formando o
citoesqueleto, tem estrutura que limita os processos difusionais. Rigorosamente, não se pode
referir à solução do compartimento intracelular como homogênea, não só, mas também, pela
sub-compartimentalização dele pelas organelas. Como são soluções diluídas as que preenchem
os compartimentos, a concentração da água neles é elevada e o volume deles é pouco maior
que o volume da água neles contido. Considerando-se um indivíduo de tamanho médio, com
peso de 70kg, o volume total de água deste organismo é de 42 L. Destes, 24 L estão no
compartimento intracelular. No compartimento intersticial estão 13 L, dos 2 L estão nos
ossos, 3 L no tecido conjuntivo denso, e 8 L nos demais tecidos. A água dos ossos e do tecido
conjuntivo denso é trocada mais lentamente com a água plasmática, dada a estrutura densa
destes tecidos. Para certas considerações, a subdivisão do volume intersticial é conveniente.
Em um organismo de porte do que está sob consideração, o volume total de sangue, contido
no sistema vascular, é de 5 L. Para um hematócrito de 40% (volume de células/volume de
snague), o volume de plasma é de 3 L.
A soma dos volumes dos compartimentos transcelulares é de 1 L.
Tabela 2: Composição dos compartimentos
Soluto
Plasma
+
Na
mM
142
K+
mM
4,4
2+
Ca
mM
1,2 (ionizado)
2,5 (total)
2+
Mg
mM
0,6 (ionizado
0,9 (total)
ClmM
102
HCO3
mM
22
H2PO4-/HPO42- mM
0,7 (ionizado
1,4 (total)
Proteínas
7 g/dl
1 mmol/l
14 mEq/l
Glicose
mM
5,5
pH
7,4
Osmolalidade
290
mOsmol/kg
Interstício
145
4,7
1,2
Célula
15
120
10-4 (ionizado)
0,55
116
25
0,8
1 (ionizado)
18 (total)
29
15
0,7 (ionizado)
1 g/dl
30 g/dl
5,5
7,4
290
Baixa
~7,2
290
Na tabela 2 tem-se a composiç ão dos compartimentos. A tabela contém os valores
normais para os solutos quantitativamente mais importantes. Obviamente as soluções são
muito mais complexas, tanto em solutos orgânicos como em eletrólitosl. O Na + é o eletrólito
predominante no compartimento extracelular, enquanto o K+ o é no intracelular. O Na+ e o K+
são metais alcalinos, em períodos contíguos na tabela periódica. No compartimento
extracelular Ca2+ e Mg2+ são o segundo e o terceiro eletrólitos catiônicos, em ordem
6
quantitativa. Estes são dois metais alcalinos terrosos, também em períodos contíguos na
tabela periódica. Dados os raios cristalinos e a dupla carga positiva destes íons, eles ligam-se
eletrostaticamente a carbonilas das proteínas. Portanto, a concentração do íon livre
(ionizado) é uma fração da concentração total, e representa um condição de equilíbrio com o
íon ligado às cadeias peptídicas. A concentração intracelular livre do Ca 2+ é cerca de 10000
vezes menor que a extracelular. Pequenos influxos de Ca 2+ para o citosol promovem
significativo aumento na concentração do íon que, por sua afinidade elevada com as
proteínas, liga-se a elas, alterando-lhes a estrutura terciária. O Ca2+ , como se verá, é um
sinalizador intracelular de modificações fisiológicas. Nas organelas intracelulares, como
retículo endoplasmático e mitocôndria, a concentração do Ca 2+ é muito mais elevada que no
citosol.
O ânion predominante em quantidade no compartimento extracelular é o Cl -, um
halogênio, seguido pelo bicarbonato, que provêm da dissociação do ácido carbônico, que se
forma pela hidratação do CO2. O sistema CO2/HCO-3 é o principal tampão do plasma. A
concentração intracelular de Cl- é mais baixa que a extracelular. Tanto as soluções
extracelulares como as intracelulares são eletricamente neutras de um ponto de vista
macroscópico. Isto é, a soma das cargas positivas é igual à soma das negativas. Um pequeno
desbalanceamento gera a diferença de potencial elétrico entre os compartimentos
intracelular e extracelular, a diferença de potencial elétrico de membrana, da ordem de
poucas dezenas de mV, que é característico de todas as células.
O compartimento plasmático contém 7g/dl de proteínas. A albumina é a mais
abundante. Como o endotélio dos capilares é quase impermeável a grandes peptídios, a
concentração intersticial deles é baixa. Proteínas que escapem do vaso, atravessando o
endotélio são recolhidas pelos vasos linfáticos. A concentração de proteínas intracelulares é
elevada, e a variedade delas é imensa. No pH plasmático, a carga elétrica das proteínas é
predominantemente negativa. Como as proteínas se restrigem ao compartimento vascular, a
carga elétrica delas, por equilíbrio de Donnan, determina pequenas diferenças nas
concentrações de cátions ânions eletrolíticos entre os compartimentos plasmático e
intersticial.
O pH extracelular é ligeiramente alcalino. O intracelular quase neutro.
As concentrações dos solutos e volume do compartimento extracelular são
finamente reguladas pelas grandes funções fisiológicas, associadas à homeostase. Por
exemplo, as concentrações plasmáticas de Na+ ,K+, Ca2+ e Mg2+ dependem do balanço entre a
absorção intestinal e excreção renal. As concentrações de Ca 2+ e Mg2+ livres plasma
dependem também da deposição e remoção destes metais dos ossos. A glicemia depende de
balanço entre o consumo pelas células do organismo e a liberação hepática do
monossacarídeo.
A água no organismo
A vida, tal qual ocorre no planeta, dependeu e depende da existência prévia da água.
Este solvente dos seres vivos tem propriedades físico-químicas muito peculiares que serão
oportunamente discutidas. Pode-se adiantar que entre elas estão a sua natureza de dipolo,
que permite à molécula interação eletrostática, sem ter carga, e a sua capacidade por
formar pontes de H.
7
A água está em equilíbrio osmótico entre os compartimentos intracelular e
intersticial. Equilíbrio osmótico pressupõe
que a pressão osmótica e, portanto, a
osmolaridade das soluções, nos dois compartimentos, são as mesmas, e que a membrana
celular plasmática é permeável à água. A osmolaridade das soluções é de 290 mOsmóis/l, e a
pressão osmótica delas, pela equação de van’t Hoff é de 6,5 atm. A membrana plasmática é
permeável à água por causa da permeabilidade da bicamada e das aquaporinas, canais para
água formados por proteínas geneticamente codificados. A conseqüência do equilíbrio
osmótico entre os compartimentos intersticial e intracelular é que modificações primárias na
osmolalidade daquele, refletem-se no volume deste.
O endotélio dos capilares, entre os compartimentos plasmático e intersticial, é
francamente permeável aos solutos de baixo peso molecular. Excluídas as proteínas, a
osmolaridade dos dois compartimentos é praticamente a mesma. Porém, a pressão
hidrostática nos capilares, dada pelo trabalho cardíaco, para impulsionar o sangue, é maior no
interior dos capilares, algo da ordem de 25 mmHg acima da atmosférica. A intersticial é
atmosférica. Portanto, há uma diferença de pressão hidrostática de 25 mmHg, que força o
movimento de água do vaso para o interstício. Este fluxo de água, dado que as soluções são
diluídas e que o volume molar parcial do solvente predomina, corresponderia a fluxo de
volume. Todavia, a presença exclusiva de proteínas no compartimento plasmático produz uma
pressão osmótica da ordem de 25 mmHg, que, igual à pressão hidrostática, anula o fluxo de
volume. A pressão osmótica dada por macromoléculas é denominada pressão oncótico ou
coloidosmótica. As duas forças operando nos capilares foram denominas forças de Starling.
O desbalanceamento delas, por exemplo por um aumento de pressão hidrostática ou redução
da concentração de proteínas, leva à transferência de volume para o interstício,
característico do edema.
O equilíbrio osmótico entre os compartimentos tem conseqüências para o organismo
na higidez e na doença. Considere, por exemplo, que o NaCl, dissociando-se em Na+ e Cl-, é o
soluto predominante no compartimento extracelular. Contribui com cerca de 240 dos 290
mOsmóis/l neste compartimento. Suponha-se que por um certo intervalo de tempo a
absorçao intestinal do NaCl ingerido exceda a excreção renal do sal. Como a distribuição
dele é principalmente extracelular, a osmolaridade deste compartimento tenderá aumentar.
Como há equilíbrio osmótico, imediatamente água do compartimento intracelular move-se
para o extracelular, e as células murcham. Sobrevem a sede, ingere-se água, o volume celular
é recuperado. Mas o organismo terá ganho algum NaCl e água, com expansão do
compartimento extracelular e, eventualmente, hipertensão.
Sinopse das funções da membrana celular plasmática.
Embora em alguns tecidos células possam estabelecer conexões entre si por meio de
canais juncionais (“gap junction”) pelos quais se propagam sinais elétricos de célula para
célula, ou mesmo sinais químicos legados por pequenas moléculas, que passam de célula para
célula, a membrana celular plasmática é uma fase predominantemente hidrofóbica, contínua,
a separar o compartimento celular do intersticial. Para entrar ou sair das células as espécies
químicas têm de atravessar esta fase. Como os compartimentos separados pela membrana
são de composição diferente, o transporte nela é seletivo.
8
Como a matriz da membrana é lipídica, substâncias com algum grau de
lipossolubilidade atravessam-na por difusão. É o caso dos gases, N2, O2 e CO2. Para as
espécies químicas hidrossolúveis a matriz da membrana celular é uma barreira. Se estiverem
implicadas na fisiologia da célula, haverá nas membranas sistemas de transportes, formados
por proteínas geneticamente codificadas. Estas proteínas podem formar canais,
relativamente hidrofílicos e seletivos. Por eles o fluxo é passivo. Servem, na sua maioria, à
transferência de íons inôrganicos. Os canais podem ser modulados de vários formas, por
voltagem ou por ligantes químicos, e geram sinais elétricos na superfície das células,
associados à condificação ou à transferência de informação ao longo da superfície de uma
célula. Outras proteínas formam carregadores, nos quais o processo de transporte depende
de alterações conformacionais da molécula. Estes carregadores podem levar uma única
espécie química, como é o caso dos transportadores de glicose, GLUT. Podem, transferir em
um ciclo duas ou mais espécies químicas, caso em que, sem violar a termodinâmica, uma das
espécies pode ser transportada contra uma diferença de energia livre. Outros carregadores,
as bombas, usando de energia metabólica. levam especíes químicas contra gradientes de
energia livre.
Outras espécies químicas, proteínas principalmente, produzidas e armazenadas pelas
células em vesículas podem ser exportadas quando estas vesículas fundem-se à membrana
superficial e liberam extracelularmente o conteúdo. O processo é de exocitose.
Um conjunto de células é um organismo quando o funcionamento delas é coordenado.
A coordenação depende de sinais químicos, moléculas que são produzidas por células
especializadas e lançadas no meio interno. Alguns destes sinais químicos podem cruzar as
membranas celulares e agir no compartimento intracelular. Outros ligam-se a receptores na
membrana celular plasmática e suscitam cascatas enzimáticas com produção de mensageiros
intracelulares que modificam a fisiologia das células.
A análise destes processos na membrana celular é o propósito deste curso.
SISTEMAS DE REGULAÇÃO – HOMEOSTASE
“De fato, regulação em um organismo é o problema central em fisiologia”. W. B. Cannon, 1929.
Regulação, um dos substantivos que mais freqüentemente aparece em textos de
fisiologia, é um conjunto de processos pelos quais variáveis químicas e físicas dos sistemas
biológicos são mantidas não constantes, mas dentro de limites, limites estes que podem ser
redefinidos, em resposta a atitudes comportamentais do organismo, ou oscilar de forma
periódicas, adapatando o organismo aos ciclos da sua existência. Os exemplos são muitos,
entre eles a regulação da pressão arterial, a regulação da freqüência cardíaca, regulação da
temperatura, regulação do volume extracelular, regulação do volume celular, regulação da
secreção gastrintestinal, etc. Defeitos em qualquer parte do sistema de regulação provocam
a deriva da variável para valores fora dos limites considerados populacionalmente normais.
Estas variações ocorrem em estados mórbidos, como na hipertensão, hipertermia,
hipernatremia, acidose, etc.
Os processos de regulação, nos quais determinada variável é mantida dentro de
limites de valor, envolvem um esquema circular de retroalimentação, palavra que bem traduz
9
“feed-back”. Se o esquema mantém a variável entre limites, envolve retroalimentação
negativa. Um mecanismo de regulação, operando em retroalimentação, envolve os elementos
básicos, mostrados no esquema da figura.
K
F
Sensor
Y=f(x)
x(t)
10
Y-K
9
8
7
6
w
5
Series2
4
3
2
1
0
0
20
40
60
80
100
120
Efetor
Z=f(y-K)
Figura 4: Esquema de um mecanismo de retroalimentação. O sensor gera um sinal, y,
relacionado pelo valor da variável x(t). O centro de controle compara o valor de y ao valor
programado K. Se forem iguais o sinal na saída no centro é nulo. Se houver discrepância o
sinal de saída, proporcional ao erro, determina que o efetor atue no sentido de recuperar o
valor da variável x(t).
Um sensor faz medidas da variável em um série temporal contínua. O sinal deste
sensor é enviado a um centro de controle, que o compara continuamente ao valor K,
programado para a variável. Se o valor medido da variável diverge do valor programado o
centro de controle aciona um efetor que atua sobre a variável e promove correções que a
aproximam o valor programa. O valor programado, para muitas das variáveis, pode ser
modificado para uma maior flexibilidade adaptativa dos organismos.
Um sistema de retroalimentação promove correção dos desvios da variável. As
amplitudes das oscilações dela dependerão de características dos sistema, que definem-lhe o
ganho. Em um sistema de ganho elevado, a amplitude das oscilações é menor.
10
Analise-se para maior concretude, a regulação da pressão arterial. Sensores, que são
os receptores de estiramento na parede das artérias enviam ao tronco cerebral salvas de
potenciais de ação (oscilações breves, ms, do potencial de membrana) cuja freqüência está
univocamente relacionado à pressão do sangue no interior da artéria. Núcleos de neurônios
no tronco cerebral respondem às informações que lhe chegam, providenciando, se houver
desvios, as correções modificando a fisiologia dos efetores, que, no caso, são o débito
cardíaco (volume de sangue injetado por segundo na aorta) e a resistência periférica ao
fluxo, determinada principalmente pelo calibre das pequenas artérias, a montante dos
capilares.
Questões Orientadoras do Estudo
1.
2.
3.
4.
5.
6.
Faça uma análise comparativa extensa dos padrões celulares contemporâneos, o
procariótico e o eucariótico.
Reveja a compartimentalização dos organismos metazoários, investigando as
barreiras que os separam, diferenças na composição e na “função”.
Suponha que um indivíduo ingira 200g de NaCl (peso molecular 58,44). Considere,
como uma aproximação, que a distribuição do sal é exclusivamente extracelular, que
os volumes extracelular e intracelular sejam de, respectivamente, 12 e 25 l e que a
osmolaridade dos compartimentos seja de 290 mOsmóis/l. Que alterações ocorrerão
nos volumes relativos dos compartimentos e qual será a osmolaridade final deles.
Considere um bloqueador de canais para Ca, usado para tratamento de hipertensão. O
endotélio vascular é permeável à droga, mas a membrana celular não o é. Suponha que
a concentração para um efetivo bloqueio dos canais, que produz vasodilatação
significativa, é de 1 microg/l. Supondo que a absorção da droga é rápida e que a sua
excreção é lenta, calcule a quantidade, em g, que se deve administrar. Suponha,
agora, que a a droga se ligue à albumina plasmática: na concentração de 1 microg/l,
cada g de albumina se liga a 0,6 microg do bloqueador. Recalcule a quantidade da
droga a ser administrada. (volume de sangue = 5l, hematócrito 40%, volume
intersticial 13l, concentração plasmática de albumina = 70 g/l.
Considere uma queda na concentração plasmática de proteína, de 7 para 3 g/dl. A
alteração poderia ocorrer por doença renal, em que ocorre proteinúria. Que
alterações relativas ocorreriam nos volumes dos compartimentos extracelulares.
Escolha uma das variáveis do meio interno submetidas a rígido controle
homeostático. Tente, com sua cultura fisiológica atual, montar esquema de
retroalimentação negativa que mantém o seu valor.
11