UNIVERSIDADE FEDERAL DO PARÁ
INSTITUTO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO
BIOLOGIA DE AGENTES INFECCIOSOS E PARASITÁRIOS
INTERAÇÃO PARASITO HOSPEDEIRO ENTRE METACESTÓIDES E Ageneiosus
ucayalensis CASTELNAU 1855 (PISCES SILURIFORMES) ORIUNDOS DA BAIA
DO GUAJARÁ-BELÉM- PARÁ: ASPECTOS MORFOLÓGICOS, MOLECULARES,
BIOLÓGICOS E ANGIOGÊNICOS
FABRISIA SILVA D’ENCARNAÇÃO
Belém-Pará
2013
FABRISIA SILVA D’ENCARNAÇÃO
INTERAÇÃO PARASITO HOSPEDEIRO ENTRE METACESTÓIDES E
Ageneiosus ucayalensis CASTELNAU 1855 (PISCES SILURIFORMES)
ORIUNDOS DA BAIA DO GUAJARÁ-BELÉM- PARÁ: ASPECTOS
MORFOLÓGICOS, MOLECULARES, BIOLÓGICOS E ANGIOGÊNICOS
Dissertação de Mestrado apresentado ao
Curso de Mestrado do Programa de PósGraduação em
Biologia de Agentes
Infecciosos e Parasitários do Instituto de
Ciências Biológicas da Universidade Federal
do Pará como requisito para obtenção do grau
de Mestre em Biologia de Agentes Infecciosos
e Parasitários.
Orientadora: Profa. Dra. Jeannie Nascimento
dos Santos.
Belém-Pará
2013
FABRISIA SILVA D’ENCARNAÇÃO
INTERAÇÃO PARASITO HOSPEDEIRO ENTRE METACESTÓIDES E Ageneiosus
ucayalensis CASTELNAU 1855 (PISCES SILURIFORMES) ORIUNDOS DA BAIA
DO GUAJARÁ-BELÉM- PARÁ: ASPECTOS MORFOLÓGICOS, MOLECULARES,
BIOLÓGICOS E ANGIOGÊNICOS
Dissertação de Mestrado apresentado ao Curso de Mestrado do Programa de PósGraduação em Biologia de Agentes Infecciosos e Parasitários do Instituto de
Ciências Biológicas da Universidade Federal do Pará como requisito para obtenção
do grau de Mestre em Biologia de Agentes Infecciosos e Parasitários.
Orientadora:
Profa. Dra. Jeannie Nascimento dos Santos
Laboratório de Biologia Celular e Helmintologia
ICB/UFPA
Banca Examinadora:
Prof. Dr. Arnaldo Maldonado Junior
Laboratório de Biologia e Parasitologia de Mamíferos
Silvestres e Reservatório, FIOCRUZ/RJ
Prof. Dr. Maurílio José Soares
Laboratório de Biologia Celular, Instituto Carlos
Chagas, FIOCRUZ/PR
Profa. Dra. Elane Guerreiro Giese
Laboratório de Histologia e Embriologia Animal,
ISPA – UFRA
Prof. Dr. Adriano Penha Furtado (Suplente)
Laboratório de Biologia Celular e Helmintologia,
ICB/UFPA
Belém, 06 de Dezembro de 2013
―A mente que se abre a uma nova ideia
jamais voltará ao seu tamanho original‖.
Albert Einstein (1879 – 1955)
Dedico este trabalho a Deus, criador de todas as
coisas inclusive os parasitos. Aos meus pais e a
todos os meus amigos pela presença constante e
encorajamento durante essa jornada.
Fabrisia D`Encarnação
AGRADECIMENTOS
Em primeiro lugar expresso meu agradecimento a Deus que me direcionou
em todos os momentos e colocou ao meu dispor todos que de alguma forma
contribuíram para a realização desse trabalho.
Aos meus familiares pelo encorajamento, compreensão e orações, mesmo de
longe, direcionadas ao meu favor.
Á minha orientadora Profa. Dra. Jeannie Nascimento dos Santos, pela
diligência e sabedoria em orientar e contornar todos os obstáculos que sobrevieram
durante a realização desse trabalho.
À Prof. Dra. Elane Guerreiro Giese, do Laboratório de Histologia e
Embriologia Animal, ISPA – UFRA pelas ajudas nunca negadas quando foram
necessárias e apeladas, durante a realização dessa monografia.
As Prof. Dr Adriano Penha Furtado do Laboratório de Biologia Celular e
Helmintologia ―Profa. Dra. Reinalda Marisa Lanfredi‖, pelas contribuições durante a
realização deste trabalho.
Ao Prof. Dr. Paulo Pimenta do Laboratório de Entomologia Médica do Instituto
Osvaldo Cruz de Belo Horizonte – Minas Gerais (FIOCRUZ-MG) que colocou o seu
laboratório à disposição para a realização de treinamentos e parte do procedimento
da Microscopia Eletrônica de Varredura e Transmissão.
À mestranda Luciana de Cássia Silva do Nascimento, pela paciência,
contribuição na realização dos experimentos e esclarecimento de dúvidas sempre
que foi necessário.
Ao Prof. Dr. Evonnildo Gonçalves do Laboratório de Tecnologia Biomolecular
do Instituto de Ciências Biológicas - UFPA, pelo fornecimento do espaço físico para
concretização de algumas etapas desse trabalho e pelas valiosas contribuições na
realização deste trabalho.
À Profa. Dra. Edilene Oliveira, responsável pelo Laboratório de Ultraestrutura
do Instituto de Ciências Biológicas da Universidade Federal do Pará pelo espaço
cedido para utilização de equipamentos necessários durante a realização de
técnicas histológicas.
Ao Prof. Dr. Antonio Vallinoto do Laboratório de Virologia do Instituto de
Ciências Biológicas - UFPA, pela disposição, opiniões cedidas e tentativas de ajuda
em algumas áreas do desenvolvimento dessa dissertação.
Aos mestrandos e doutorandos do laboratório de Biologia Celular e
Helmintologia ―Profa. Dra. Reinalda Marisa Lanfredi‖ pelo carinho, alegria, choro e
risadas que demos juntos nessa jornada e que muito nos acrescentou como
cidadãos.
A todos os estagiários do Laboratório de Biologia Celular e Helmintologia
―Profa. Dra. Reinalda Marisa Lanfredi‖, que de alguma forma deram sua contribuição
para realização desse trabalho.
Às minhas conterrâneas, Elisabeth Correia Rodrigues, Tânia Carlice dos Reis,
Núria Jussara Silva, Andreneida Letícia Santos, e à Kátia do Amorim pela paciência,
compreensão e palavras de conforto, demonstradas e proferidas em muitos
momentos difíceis.
À todos os meus Irmãos em Cristo que se fizeram presentes por meio de
orações ao meu favor e que contribuiu para o encorajamento e finalização deste
trabalho
SUMÁRIO
LISTA DE FIGURAS ................................................................................................... 8
RESUMO................................................................................................................... 11
ASTRACT ................................................................................................................. 12
1 INTRODUÇÃO ....................................................................................................... 13
1.1 Ageneyosus ucayalensis (CASTELNAU, 1855). ............................................... 15
1.1.1 Classificação Taxonômica ................................................................................ 15
1.1.2 Distribuição geográfica e hábitos de vida ......................................................... 16
1.1.3 Helmintos Parasitos de Ageneiosus ucayalensis ............................................. 17
1.2 CESTODA ........................................................................................................... 19
1.2.1 Características dos Cestodas da Família Proteocephalidae ............................ 25
1.2.2 Ciclo de vida dos Cestodas da família Proteocephalidae ........................... 26
1.3 PRODUTOS DE EXCREÇÃO E SECREÇÃO (E/S) DE CESTÓDEOS .............. 27
1.4.1 Fatores de Indução da Angiogênese ................................................................ 37
1.4.2 Helmintos e angiogênese ................................................................................. 40
1.4.3 Helmintos cestodas e angiogênese .................................................................. 45
1.5 OBJETIVOS ........................................................................................................ 48
1.5.1 Objetivo geral ................................................................................................... 48
1.5.2 Objetivos específicos........................................................................................ 48
2 MATERIAL E MÉTODOS ...................................................................................... 49
2.1 COLETA DE HOSPEDEIROS E AMOSTRAS DE PARASITOS CESTODAS .. 49
2.2 MICROSCOPIA DE LUZ ..................................................................................... 50
2.2.1 Análise taxonômica de Cestoda ....................................................................... 50
2.2.2 Análise histológica pelo método de inclusão em historesina® ......................... 50
2.2.3 Análise histológica pelo método de inclusão em parafina ................................ 51
2.3 BIOLOGIA MOLECULAR .................................................................................... 51
2.3.1 Extração de DNA e sequenciamento para Taxonomia e Filogenia .................. 51
2.4 MICROSCOPIA ELETRÔNICA DE TRANSMISSÃO .......................................... 53
2.5 MICROSCOPIA ELETRÔNICA DE VARREDURA (MEV)................................... 54
3 RESULTADOS ....................................................................................................... 55
3.1 AMOSTRAS DE CESTÓDEOS PARASITOS ..................................................... 55
3.2.ASPECTOS MORFOLÓGICOS DAS LARVAS ................................................... 58
3.2.1 Morfotipo I (Larvas Plerocercóides) .................................................................. 58
3.2.2 Morfotipo II (Larvas Merocercóides) ................................................................. 66
3.2.3 Morfotipo III (Larvas Merocercóides) ................................................................ 73
3.3
ANÁLISE
MOLECULAR
COMPARATIVA
ENTRE
OS
MORFOTIPOS
LARVARES I E II ....................................................................................................... 74
3.4 ANÁLISE DOS ASPECTOS ANGIOGÊNICOS DO PARASITISMO ................... 79
3.4.1 Descrição histológica........................................................................................ 79
3.4.2 Reconstrução tri- dimensional .......................................................................... 88
3.4.3 Microscopia Eletrônica de Varredura da massa cística .................................... 92
4 DISCUSSÂO ........................................................................................................ 103
4.1 ANÁLISE MORFOLÓGICA ............................................................................... 103
4.2 BIOLOGIA MOLECULAR .................................................................................. 106
4.3 CARACTERÍSTICAS MORFOLÓGICAS DA RELAÇÃO ENTRE AS LARVAS
METACESTÓIDES E A.ucayalensis ....................................................................... 107
4.3.1 Aspectos histológicos ..................................................................................... 107
4.3.2 Aspectos ultraestruturais ................................................................................ 108
4.3.3 Aspectos Angiogênicos .................................................................................. 111
4.4 ASPECTOS BIOLÓGICOS ............................................................................... 113
4.4.1 Prospecção Parcial do Ciclo Biológico ........................................................... 113
4.4.2 Ecologia do parasitismo ................................................................................. 115
5 CONCLUSÕES .................................................................................................... 117
REFERÊNCIAS ....................................................................................................... 119
LISTA DE FIGURAS
Figura 1: Características morfológicas de Ageneiosus ucayalensis ......................... 16
Figura 2: Mapa da distribuição geográfica de Ageneiosus ucayalensis ................... 17
Figura 3: Helmintos Parasitos de Ageneiosus ucayalensis visualizados pela
Microscopia Eletrônica de Varredura e Microscopia de Luz ...................................... 18
Figura 4: Representação esquemática e ilustrativa das estruturas externas
constituintes do corpo de cestódeos. ........................................................................ 20
Figura 5: Organização tegumentar de cestódeos). .................................................. 21
Figura 6: Representação esquemática das estruturas de excreção e
osmorregulação contituintes dos cestódeos.............................................................. 22
Figura 7: Representação esquemática de seis formas larvares de cestóides ou
metacestoides. .......................................................................................................... 23
Figura 8: Desenho ilustrativo das estruturas dos ovos de parasitos da classe
Cestoda. .................................................................................................................... 24
Figura 9: Processo de desenvolvimento de formas larvares dos parasitos da família
Proteocephalidae, em hospedeiros intermediários e definitivos. ............................... 26
Figura 10: Ciclo de vida de Cestoda da Família Proteocephalidae .......................... 27
Figura 11: Representação esquemática das estruturas que compõe o sistema
circulatório. ................................................................................................................ 31
Figura 12: Ilustração das camadas de revestimento dos vasos sanguíneos. .......... 32
Figura 13: Processo angiogênico desencadeado pela desestabilização do equilíbrio
das substâncias pró e anti-angiogênicas. ................................................................. 33
Figura 14: Esquema ilustrativo dos tipos de angiogênese que podem ocorrer a partir
de plexus capilares primitivos ou vasculogênese)..................................................... 34
Figura 15: Esquema ilustrativo do processo de migração e proliferação das células
endoteliais para formar novos vasos sanguíneos. .................................................... 36
Figura 16: processo de indução da proliferação celular e formação de
vascularização em eventos patogênicos. .................................................................. 37
Figura 17: Mecanismo de sinalização do VEGF nas células endoteliais). ............... 38
Figura 18: Mecanismo de liberação e ligação do fator de crescimento endotelial
vascular (VEGF) desencadeando a formação de novos vasos sanguíneos ............. 39
Figura 19: Variantes do fator de crescimento endotelial vascular e os possíveis
receptores, aos quais estas substâncias se ligam para desencadear eventos
angiogênico ............................................................................................................... 40
Figura 20: Esquema ilustrativo da angiogênese desencadeada por parasitos
helmintos. .................................................................................................................. 41
Figura 21: Estimulação das células endoteliais e formação de capilares ................. 42
Figura 22: Papel das galectinas 1 e 3 na imunomodulação do sistema imune do
hospedeiro e na indução da proliferação celular e angiogênese, desencadeado pelos
patógenos.................................................................................................................. 43
Figura 23: Hospedeiro Ageneiosus ucayalensis e a localização e disposição dos
cistos de cestóides e os vasos sanguíneos dentro do hospedeiro. ........................... 56
Figura 24: Desenho morfológico da fase larval Plerocercóide do Morfotipo I .......... 58
Figura 25: Características morfológicas das Larvas do Morfotipo I visualizadas por
meio de técnicas histológicas. ................................................................................... 60
Figura 26: Caracteristicas morfológicas da bolsa cistica e das formas larvares de
cestóides, observados pela Microscopia Eletrônica de Varredura.. .......................... 63
Figura 27: Características morfológicas das larvas plerocercóides de Cestódeos
encontrados parasitando Ageneiosus ucayalensis, observados pela Microscopia
Eletrônica de Varredura. ........................................................................................... 64
Figura 28: Desenho morfológico de larvas do Morfotipo II (Merocercóides). ........... 66
Figura 29: Características morfológicas das Larvas do Morfotipo II visualizadas por
meio de técnicas histológicas. ................................................................................... 68
Figura 30: Características morfológicas de larvas do Morfotipo II (fase larvar
Merocercóide). .......................................................................................................... 71
Figura 31: Desenho morfológico da larva do Morfotipo III (merocercóide) ............... 73
Figura 32: Árvore filogenética obtida pelo método da Máxima Parcimônia, avaliando
parte da Subunidade menor do DNA ribossomal de Cestódeos ............................... 76
Figura 33: Árvore filogenética feita utilizando o método da Inferência Baesiana, com
a finalidade de comparar parte da Subunidade menor do DNA ribossomal de
Cestódeos ................................................................................................................. 77
Figura 34: Análise de cortes histológicos de amostras embebidas em historesina® e
coradas pelo Azul de Toluidina. ................................................................................ 81
.Figura 35: imagens de secções histológicas semi-finas do tecido contendo cistos de
parasitos Cestódeos e vasos sanguineos ao redor ................................................... 83
Figura 36: Análise histológica de Cistos e vasos em amostras embebidas em
parafina e submetidos à coloração pela Hematoxilina – Eosina, Tricrômio de Mallory
e Orceína................................................................................................................... 84
Figura 37: Análise histológica de Cistos e vasos embebidos em parafina e
submetidos à coloração pelo Tricrômio de Gomori e pelo Ácido Periódico de Schiff
(PAS). ........................................................................................................................ 86
Figura 38: Reconstrução tri-dimensional dos cistos de cestóides e vasos
sanguíneos adjacentes a partir da somatória de 201 secções seriadas de material
embebido em Historesina®, obtido do parasitismo por cestóide em Ageneiosus
ucayalensis da Baia do Guaja, Belém-PA. ................................................................ 89
Figura 39: Diversos ângulos da reconstrução tri-dimensional obtidas a partir de
rotações no eixo X e Y, permitindo a visualização detalhada das estruturas
envolvidas no processo angiogênico induzida por larvas de Cestódeos em
Ageneiosus ucayalensis. ........................................................................................... 91
Figura 40: Características morfológicas do saco de tecido conjuntivo e vasos
sanguíneos que envolvem os cistos de Cestódeos, observados pela Microscopia
Eletrônica de Varredura. ........................................................................................... 93
Figura 41: Estruturas celulares e teciduais da bolsa cística ricamente vascularizada
e formada por larvas de Cestódeos em Ageneiosus ucayalensis observados através
da técnica de Microscopia Eletrônica de Transmissão. ............................................. 97
Figura 42: Aspectos da inter-relação vasos sanguíneos, cisto e, tegumento de
larvas de Cestódeos ou Metacestódeos observados através da técnica de
Microscopia Eletrônica de Transmissão. ................................................................... 99
Figura 43: Ultraestrutura do tegumento de larvas de Cestódeos ou Metacestódeos
observados através da técnica de Microscopia Eletrônica de Transmissão. .......... 101
Figura 44: Diagrama ilustrativa de possíveis formas de atuação de Ageneiosus
ucayalensis, no ciclo biológico de Metacestódeos Proteocephalídeos de Belém –PA.
................................................................................................................................ 113
RESUMO
Os helmintos parasitos interagem com seus hospedeiros modulando o sistema
imune e desta forma garantindo o seu desenvolvimento, crescimento e longevidade
nesses hospedeiros. Um dos meios encontrados para garantir esta permanência é a
formação de uma vascularização que atua no transporte de oxigênio e nutrientes do
hospedeiro para os parasitos e na excreção de metabólitos desses parasitos para o
hospedeiro. Este processo é bem caracterizado No parasitismo de alguns helmintos
como o Schistosoma mansoni, porém pouco se sabe a respeito desse aspecto em
relação a outros helmintos parasitos. Assim, o presente trabalho tem como objetivos
caracterizar os aspectos estruturais e moleculares básicos de larvas de Cestoda da
família Proteocephalidae parasitos de Ageneiosus ucayalensis, assim como
caracterizar aspectos da biologia e do parasitismo dessas larvas, incluindo processo
angiogênico no hospedeiro. Exemplares de Ageneiosus ucayalensis da Baía do
Guajará foram adquiridos ainda vivos de pescadores ribeirinhos e as larvas de
Cestoda encistadas foram retiradas e submetidas à identificação através das
técnicas de taxonomia clássica e biologia molecular. Técnicas histológicas e de
microscopia eletrônica foram aplicadas para descrição dos aspectos celulares e
estruturais do parasitismo por estas larvas. Entre as larvas de Cestoda, três
morfotipos distintos, possivelmente pertencentes a gêneros e espécies distintas, em
diferentes fases (Plerocercóide e Merocercóide) foram identificados. Os cistos
estabelecem contato íntimo com a superfície dos vasos sanguíneos e os parasitos
mostram aspectos ultraestruturais característicos de atividades de troca de
metabólitos entre tecido do hospedeiro e parasito. No entanto são necessários
estudos futuros sobre as moléculas envolvidas no estabelecimento do quadro deste
parasitismo bem como a caracterização de fatores angiogênicos.
Palavras- chave: Metacestódeos, Ageneiosus ucayalensis, Baia do Guajará, Belém,
Angiogênese
ASTRACT
The helminth parasites interact with their host by modulating the immune system and
ensuring the growth, longevity and their development in these hosts. One way found
to guarantee the stay inside the host is the formation of a vasculature that mediate
the transport of oxygen and nutrients from the host to the parasites and excretion of
metabolites of these parasites to the host. This process is well characterized in some
parasitic helminths such as Schistosoma mansoni, but little is known about those
aspects to other helminth parasites. Thus, the present study has the objective to
characterize the structural and molecular aspects of Proteocephalidae metacestoes
parasites of Ageneiosus ucayalensis, as well as characterize the biology and
parasitism of the larvae, including the angiogenic process in the host. Samples of live
Ageneiosus ucayalensis from Guajará Bay were acquired by local fishermen and
encysted metacestode were removed and identified using classical taxonomy and
molecular biology techniques. Histological and electron microscopy techniques were
applied to describe the cellular and structural aspects of parasitism by these larvae.
Among the metacestode found, we observed three distinct morphotypes, possibly
belonging to different genera and species, in different phases (Plerocercoid and
Merocercoid). The cysts establish intimate contact with the surface of blood vessels
and ultrastructural aspects of trading activities of metabolites between the host tissue
and parasites were observed. However, future studies are necessary to understand
the molecules involved in the larval estabilishment as well as to characterize the
possible angiogenic factors.
Keywords:
Angiogenesis
Metacestodes,
Ageneiosus
ucayalensis,
Guajará
Bay,
Belem,
13
1 INTRODUÇÃO
A interação entre o sistema imunológico de vertebrados e alguns dos
principais grupos de helmintos ocorre desde o início da co-evolução entre
hospedeiro e parasito (Jackson et al., 2008). A capacidade que estes parasitos têm
de modular o sistema imunológico aumenta sua longevidade no hospedeiro
vertebrado. A base molecular dessa imunomodulação realizada por helmintos é de
grande interesse científico uma vez que irá refletir a história de vida, estratégias de
transmissão e nichos fisiológicos de Nematoda, Cestoda e Trematoda (Hewitson et
al., 2009).
Para se evadir do sistema imunológico do hospedeiro, os parasitos produzem
uma série de enzimas e proteínas que atuam inibindo ou alterando o funcionamento
do sistema imune em favor dos parasitos, e isso os possibilita um ambiente estável
para o seu desenvolvimento, crescimento e permanência no hospedeiro. Assim, o
equilíbrio entre as respostas pró e anti-inflamatórias determinam o destino do
parasito e as imunopatologias induzidas no hospedeiro (Klaus, 2007).
Várias pesquisas em Biologia Molecular da interação parasito-hospedeiro têm
demonstrado a ação dessas substâncias protéicas e enzimáticas, como por
exemplo: 1) na inibição da proliferação de células T e produção de óxido nítrico por
macrófagos, desencadeado por Heligmosomoides polygyrus (Rzepecka et al., 2006);
2) na indução da resposta Th2 e inibição da migração de neutrófilos por
Equinococcus granulosus (Siracusano et al., 2008); 3) na promoção da invasão
parasitária e infeção de hospedeiros mamíferos por Fasciola hepatica (Robinson et
al., 2009); 4) na inibição do processamento de antígenos e modulação da resposta
imune antígeno específico por Nipostrongilus brasiliensis (Dainichi et al., 2001); 5)
na inibição da sinalização de receptores Toll Like por Schistosoma mansoni
(Brannstrom et al., 2009); 6) na inibição da proliferação de células TCD4+ e células B
por Acanthocheilonema viteae (Marshall et al., 2005), entre outros. Estes e demais
estudos têm contribuído para a comprovação e elucidação desses mecanismos
imunomoduladores (Hewitson et al., 2009).
A análise sistemática de produtos de Excreção/Secreção (E/S) de helmintos
só se tornou possível com os avanços das técnicas de Biologia Molecular,
favorecendo a compreensão do parasitismo e da interação existente entre parasitos
e hospedeiros, assim como o fornecimento de dados relevantes para a aplicação
14
terapêutica, como o desenho de novas drogas anti-helmínticas, vacinas e
diagnósticos (Brehm et al., 2006).
Além da imunomodulação da resposta do hospedeiro induzida pelos
helmintos, também há evidências de uma possível relação entre infecções
helmínticas e doenças alérgicas. As infecções por helmintos constituem um dos
fatores ambientais que estão associados ao risco de desenvolvimento ou não de
alergias (Flohr et al., 2008).
Klaus (2007) estabeleceu a hipótese de bloqueio de imunoglobulina E, onde
se acredita que as IgEs não específicas (policlonais) induzida pela infecção por
helmintos saturam os sítios de ligação da IgE impedindo a ligação de alérgenos IgE
específicos, mostraram que esta interação pode suprimir a resposta imunológica
contra alergia.
A hipótese mencionada anteriormente não foi constatada por Wohlleben et al.
(2004) quando estes utilizaram dois grupos de camundongos, um infectado por
Nipostrongylus brasiliensis e imunizado com ovalbumina e outro apenas imunizado
com ovalbumina, e avaliaram o grau de competição das IgEs de N. brasiliensis, e
IgEs de ovalbumina pelos sítios de ligação de IgEs. Após os experimentos
observaram que não houve diferença significativa na expressão alérgica pelos dois
grupos. Este tipo de interação pode se mostrar positiva ou negativa dependendo da
espécie estudada, o tempo e a intensidade da infecção, e são de grande relevancia,
pois podem trazer novidades quanto ao desenvolvimento de drogas derivados de
parasitas não só para o tratamento como, também, para a prevenção de doenças
alérgicas.
Outro aspecto da relação helminto-hospedeiro é a indução da proliferação
celular verificada na associação entre o parasitismo por helminto e fibrose hepática,
assim como entre o parasitismo por helminto e a angiogênese. O quadro de fibrose
no organismo se caracteriza pelo acúmulo de matriz extracelular e exibe tanto
neovascularização, quanto a proliferação de células secretoras de matriz (Lemos et
al., 2010).
Dentre os diversos fatores que podem desencadear a fibrose hepática,
encontram-se as infecções helmínticas provocadas principalmente por Schistosoma
mansoni Silva, 1908 e Capillaria hepatica Bancroft, 1893. Em ambas patologias a
fibrose ocorre devido ao acúmulo de ovos desses parasitos no fígado do hospedeiro,
causando obstrução com o comprometimento da fisiologia hepática e esclerose.
15
Segundo os experimentos de Kanse et al. (2005) e Lemos et al. (2010), os ovos são
as únicas formas de vida de S.mansoni, capazes de secretar/excretar substâncias
que estimulam a proliferação e a migração das células endoteliais, e células dos
músculos lisos dos vasos.
A progressão da doença para o estágio de fibrose na capilaríase hepática
envolve a proliferação de vasos sanguíneos. Nos dois casos, a presença dessas
formas parasitárias no tecido, estimula uma resposta inflamatória que culmina na
ativação de leucócitos, fibroblastos e células endoteliais desencadeando uma reação
de cicatrização da área lesionada. Esta reação se traduz inicialmente em um
processo angiogênico e, posteriormente, em uma fibrose nesse local.
Os mecanismos para elucidação da origem desses estímulos, do hospedeiro
ou do parasito, são alvos dos pesquisadores no quesito interação ParasitoHospedeiro.
1.1
Ageneyosus ucayalensis (CASTELNAU, 1855).
1.1.1 Classificação Taxonômica
Ageneiosus ucayalensis, peixe siluriforme conhecido popularmente como
mandubé, é um integrante da Familia Auchenipteridae e se caracteriza,
principalmente, por apresentar corpo não revestido por escamas, cabeça afilada na
sua extremidade anterior, fenda bucal larga e ampla, olhos situados lateralmente no
prolongamento da boca, dentes diminutos, viliformes e localizados em placas. Os
barbilhões maxilares são geralmente reduzidos e os mentonianos, ausentes (Fig. 1).
Apresentam dimorfismo sexual em que os machos têm barbilhões maxilares e os
espinhos da nadadeira dorsal rígidos, hipertrofiados e providos de ganchos. São
todos carnívoros pelágicos e ocorrem geralmente no leito dos rios. Além desta
espécie, existem outras três espécies que ocorrem no Brasil: Ageneiosus brevifilis
Valenciennes, 1840 e Ageneiosus dentatus Kner, 1858 de maior valor comercial e
Ageneiosus inermis Linnaeus, 1766 (Santos et al., 2006).
16
Figura 1: Características morfológicas de Ageneiosus ucayalensis, evidenciando o
corpo sem escamas, a cabeça afilada na estremidade anterior e olhos laterais em
relação ao prolongamento da boca. Barra = 3 cm. Fonte: Laboratório de Biologia
Celular e Helmintologia (2012).
1.1.2 Distribuição geográfica e hábitos de vida
Os peixes ocupam a maioria dos níveis tróficos, desde herbívoros até
carnívoros secundários, podendo ser generalistas, sem especificidade alimentar, ou
especialistas, fazendo uso de um número restrito de tipos de alimentos. Estas
informações permitem estabelecer a dinâmica de uma determinada comunidade, e
as relações que podem influenciar a estrutura e interações ecológicas como
predação e competição (Ribeiro et al., 2003). Neste estudo os autores constataram
que A. ucayalensis era um consumidor ―de topo‖ de cadeia e mais tarde Froese &
Pauly (2010) o caracterizaram como uma espécie carnívora, podendo alimentar-se
de pequenos peixes e camarões.
A espécie A. ucayalensis encontra-se distribuída na Amazônia e bacias do
Rio Orinoco mais especificamente, na América do Sul desde a bacia do Amazonas e
Orinoco, até a bacia do alto Paraná (Fig. 2) (Froese & Pauly, 2010).
17
Figura 2: Mapa da distribuição geográfica de Ageneiosus ucayalensis, indicado pelo
círculo em vermelho. Adaptado de www.fishwise.co.za › Search › Species Detail
Page, acessado em 12 de agosto de 2012.
1.1.3 Helmintos Parasitos de Ageneiosus ucayalensis
Relatos de helmintos parasitando espécies de peixes tem se tornado
frequente na literatura. O interesse nesse tipo de parasitismo deve-se ao fato da
importação de cardápios com peixes crus da culinária oriental, destacando o
potencial zoonótico como um problema de saúde pública, uma vez que muitos
peixes servem como hospedeiros intermediários ou paratênicos de helmintos e
muitos desses parasitos tem o homem como hospedeiro definitivo (Oliveira, 2005).
Giese (2010), em sua tese de doutorado, realizou a primeira descrição da
helmintofauna de A. ucayalensis da Foz do rio Guamá e Baia do Guajará em Belém,
Pará relatando a presença de parasitos dos filos Platyhelminthes, Acanthocephala e
Nematoda. Para o filo Platyhelminthes foram descritos parasitos das classes
Cestoda (larvas plerocercóides da família Proteocephalidae e adultos representantes
do gênero Gibsoniela) e Monogenea (integrantes da família Dactylogyridae). Como
representantes
do
filo
Acantocephala
apenas
exemplares
do
gênero
Neoechinorhynchus foram encontrados, e do filo Nematoda foram observados
18
integrantes dos gêneros Anisakis, Pseudoproleptus, Cucullanus e Procamallanus
(Fig. 3).
Figura 3: Helmintos Parasitos de Ageneiosus ucayalensis visualizados pela
Microscopia Eletrônica de Varredura e Microscopia de Luz: A) Cucullanus
ageneiosus com 2 pares de papilas cefálicas (cp), e anfídios (am). Barra = 15 µm.
B) Pseudoproleptus sp com colar cefálico (cl), e anfídio (an). Barra = 10 µm. C)
Procamallanus (Spirocamallanus) belenensis com abertura oral (ma), deirídio (d) e
poro excretor (ep). Barra = 50 µm. D) Anisakis sp com dente larvar (dl), boca (b),
papilas cefálicas (pc), anfídio (an) e poro excretor (pe). Barra = 4 µm. E)
Neoechinorhynchus sp mostrando Proboscide com ganchos grandes (g) e pequenos
(gp). Barra = 10 µm. F) Gibsoniela sp mostrando escólex com ventosas triloculadas
(Vtl). Barra = 50 µm. G) Monogenea Dactylogiridae da ordem Ancyrocephalinae,
mostrando a regiao cefálica (lb), e opistohaptor (oh). Barra = 10 µm. H) Larva
Plerocercoide da família Proteocephalídea com órgão apical (oap) e ventosa circular
(v). Barra = 10 µm. Fonte: Adaptado de Giese (2010); Giese et al. (2009) e Giese et
al. (2010.
19
1.2 CESTODA
Os cestódeos formam um grupo de helmintos que se distinguem da classe
Trematoda e do filo Nematoda por dois fatores morfológicos: a ausência de um canal
alimentar que determina a fisiologia desses parasitos, e por possuírem um corpo
bastante alongado. O parasito adulto tem preferência por habitats com níveis
nutricionais elevados como o tubo digestivo, vesícula biliar e dutos pancreáticos;
enquanto que as larvas têm seus habitats em sítios mais inespecíficos e por esta
razão são de difícil visualização em órgãos dos hospedeiros vertebrados e
invertebrados (Smyth, 1994; Schmidt et al., 2009 ).
A classe Cestoda subdivide-se em duas subclasses: Cestodaria (cestódeos
cujo corpo não se divide em segmentos e que possuem apenas um tipo de órgão
reprodutor) e Eucestoda (cestódeos com corpo dividido em segmentos ou
proglótides e que possuem ambos os órgãos reprodutores feminino e masculino). Os
parasitos pertencentes à Subclasse Eucestoda possuem as três formas evolutivas
(helmintos adultos, ovos e larvas) e têm como principais características a presença
de escólex na extremidade anterior, corpo ou estróbilo dividido em proglotes, o
hermafroditismo e ovos com embrião hexacanto (Schmidt et al., 2009).
As formas adultas são parasitos de vertebrados, geralmente achatados dorsoventralmente e não apresentam cavidade geral nem tubo digestivo. São constituídos
por uma extremidade anterior denominada escólex, onde se situam os órgãos de
fixação, seguido do pescoço ou colo, região que une o escólex ao restante do corpo
do parasito, e do estróbilo onde ocorre a segmentação dos proglotes e o
desenvolvimento do parasito (Fig. 4). Os proglotes podem ser encontrados nas fases
imaturas, sem total desenvolvimento dos órgãos sexuais, maturas, com os órgãos
sexuais masculinos e femininos bem desenvolvidos, e gravídicos, repletos de ovos
embrionados (Fig. 4). Nesta fase final, esses proglotes podem se destacar do
restante do corpo do parasita e possuem movimentos próprios. Localizado no
excólex, está o sistema nervoso composto por gânglios cerebrais que inervam o
mesmo assim como toda a extensão do corpo do parasito (Schmidt et al., 2009).
20
Figura 4: Representação esquemática e ilustrativa das estruturas externas
constituintes do corpo de cestódeos: A) Cabeça ou escólice com os órgãos de
fixação; B) Pescoço ou colo; C) Proglotes imaturas, com os órgãos sexuais pouco
desenvolvidos e sem ovos; D) Proglotes maduros, com órgãos sexuais masculino e
feminino bem desenvolvidos; E) Proglotes gravídicos repletos de ovos, incorporadas
ao estróbilo e soltas. Fonte: Adaptado de Fonseca et al. (2002).
O corpo do parasito é revestido por uma camada homogênea elástica de
tecido vivo com alta capacidade metabólica denominada tegumento. Este se
constitui de uma camada distal anucleada (citoplasma distal) contínua, isto é, sem
intervenção da membrana celular, o que faz com que o tegumento seja do tipo
sincicial. O citoplasma distal comunica-se com as células corporais nucleadas que
se encontram abaixo da camada muscular superficial por meio de canais
citoplasmáticos (Fig. 5) (Schmidt et al., 2009).
Na superfície do parasito observam-se numerosas projeções denominadas
microtríquias cuja finalidade é aumentar a área absortiva do tegumento. As
microtríquias possuem uma porção distal densa que se separa da base por uma
camada multilaminar. A base é composta por arranjos de microfilamentos (Fig. 5)
(Schmidt et al., 2009).
21
Figura 5: Organização tegumentar de cestódeos. A) Estruturas constituintes do
tegumento sincicial: B) Detalhes da contituição das microtríquias na parte superficial
do tegumento onde se observa uma região apical pontiaguda (c), uma região basal
(b), separadas por placas basais multilaminares (bp) e microfilamentos basais (mf).
Fonte: Adaptado de Schmidt et al., (2009).
A excreção de metabólitos ocorre por meio das células flama ou solenócitos,
que se externalizam por meio de canais eferentes localizados nas bordas laterais de
cada proglote. Normalmente dois pares de canais, um ventrolateral e outro
dorsolateral, percorrem o comprimento do parasito e um terceiro canal transversal
une o canal ventral à margem posterior da proglote. As células flamas se conectam
a ductos coletores que desembocam dentro desses canais e, neste aparato, ainda
se encontram flagelos que fornecem a força motriz para os fluidos do sistema (Fig.
6). A função desse sistema inclui o transporte ativo e regulação iônica de resíduos
excretórios (Schmidt et al., 2009).
22
Figura 6: Representação esquemática das estruturas de excreção e
osmorregulação contituintes dos cestódeos. Fonte: Adaptado de Schmidt et al.
(2009).
De acordo com Smyth et al. (1999), as formas larvares de parasitos de
vertebrados ou invertebrados, se desenvolvem no intestino do hospedeiro
intermediário ou definitivo; porém, o desenvolvimento em forma adulta ocorre
apenas no hospedeiro definitivo quando este, ingere um hospedeiro intermediário
infectado por este parasito. De acordo com estes autores o termo metacestóide
pode ser usado para classificar todos os estágios larvais dos cestódeos.
Chervy (2002) fez uma revisão sobre a nomenclatura de tais formas larvais e
baseando-se nas características morfológicas classificou e denominou como mostra
a Figura 7.
23
Figura 7: Representação esquemática de seis formas larvares de cestóides ou
metacestoides: A) Procercóide (parasito sem escólex, geralmente caudado, com
lacuna primária, e só desenvolve quando ingerido por um segundo hospedeiro
intermediário); B) Plerocercóide (possui desenvolvimento parcial do escólex e este
apresenta evaginado, sem lacuna, com ou sem sarcômero e infectante para
hospedeiro definitivo); C) Merocercóide (presença de escólex invaginado, sem
lacuna primária, caudado ou não); D) Plerocerco (lacuna primária ausente, escólex
retraído, caudado, com ou sem bexiga); E) Cisticercóide (presença de lacuna
primária, escólex retraído, com sarcômero evidente ou reduzido; F) Cisticerco
(lacuna primária presente, escólex invaginado, com ou sem sarcômero e bexiga).
Fonte: Adaptado de Chervy (2002).
24
Dependendo do grupo de Cestoda e da espécie, o processo de formação dos
ovos podem ser diferentes devido à diversidade da forma uterina, mas de um modo
geral resultam em uma oncosfera ou embrião hexacanto dotado de três pares de
ganchos, o embrióforo, membrana que envolve a oncosfera, e a cápsula que reveste
externamente o ovo. Em algumas espécies observa-se a formação de três
membranas (Fig. 8) (Schmidt et al., 2009).
Figura 8: Desenho ilustrativo das estruturas dos ovos de parasitos da classe
Cestoda. 1 – Cápsula; 2 - Casca externa; 3 – Envoltório externo; 4 - Envoltório
interno 5 – Embrióforo; 6 – Oncosfera com seis ganchos. Adaptado de Schmidt et
al., 2009.
A Subclasse Eucestoda é composta por cinco Ordens, onde duas possuem o
maior
número
de
representantes
parasitos
de
peixes:
Trypanoryncha
e
Proteocephalidea. A Ordem Proteocephalidea possui duas famílias: Monticellidae e
Proteocephalidae. Esta última, com número representativo de espécies parasitas de
peixes já descritas, se encontra subdividida em seis Subfamílias, com 18 gêneros e
77 espécies. O gênero Proteocephalus é o maior representante, em número de
espécies descritas (Chambrier & Scholz, 2008).
25
Até a década de 30, menos de 50 espécies de proteocefalídeos eram
conhecidos na América do Sul. Com o passar dos anos, este conhecimento
aumentou, resultando em cerca de 100 espécies de proteocefalídeos descritos em
aproximadamente 70 espécies hospedeiras de peixe (Rego, 2003).
Rego (2003) considerou que a classificação sulamericana de proteocefalídeos
tem sido controversa, devido às diversificações morfológicas e o enquadramento das
espécies nas respectivas Famílias Proteocephalidae e Monticellidae. Em estudos de
filogenia, este autor questionou a validade da Família Monticellidae, sugerindo a
eliminação da mesma e de suas Subfamílias, ficando apenas a Família
Proteocephalidae dentro da Ordem Proteocephalidea. Os parasitos que pertenciam
aos gêneros da Família Monticellidae foram transferidos para as Subfamílias
Corallobothriinae ou Proteocephalinae, dependendo da presença ou não de um
metascólex.
Assim considerando esta classificação de Rego (2003), o parasito que
constitui o objeto desse estudo pertence ao:
Filo – Platyhelminthes Gegenbaur, 1859.
Classe – Cestoda Monticelli, 1892.
Sub-classe – Eucestoda Southwall, 1930.
Ordem - Proteocephalidea Mola, 1929.
Família – Proteocephalidae Mola 1929.
1.2.1 Características dos Cestodas da Família Proteocephalidae
De acordo com Smyth (1994) os membros da Família Proteocephalidae são
parasitos de peixes de água doce, anfíbios ou répteis e são de pequena ou
nenhuma importância econômica. Os helmintos adultos se caracterizam por
possuírem escólex com quatro ventosas e, algumas vezes, apresentam um órgão
apical.Os proglótides são semelhantes aos observados na Ordem Tetraphyllidea
com folículos vitelínicos localizados nas margens laterais dos proglótides. Possuem
poro genital lateral, com ovários localizados posteriormente e os numerosos
testículos preenchendo a região anterior (Scholz, 1999).
Baseando-se nas características larvares dos cestóides apresentados
anteriormente na Figura 4, os proteocefalídeos podem ter dois tipos de formas
26
larvais: plerocercóides ou merocercoides. As características comumente observadas
nas formas larvares da Família Proteocephalidae é a presença do Sarcômero, mais
especificamente em espécies do gênero Proteocephalus, que pode permanecer
ligado ao metacestóide ou separar-se, até três dias após a infecção dos hospedeiros
intermediários (Fig. 9). Também é característico a invaginação do escólex por estes
parasitos (Scholz, 1999).
Figura 9: Processo de desenvolvimento de formas larvares dos parasitos da família
Proteocephalidae, em hospedeiros intermediários e definitivos. A) Ovo liberado
pelos adultos no ambiente, contendo o embrião hexacanto; B) Quando ingerido pelo
hospedeiro intermediário, a casca do ovo rompe e o embrião é liberado; C) Embrião
desenvolve para as fases larvares seguintes, denominados de metacestóides.
Primeiramente origina a larva plerocercóide com ou sem sarcômero e este
posteriormente evolui para larva merocercóide ainda dentro do hospedeiro
intermediário. Nesta fase, o crescimento larvar se estaguina, até este encontrar um
hospedeiro definitivo onde continua o desenvolvimento até a fase adulta,
completando o ciclo evolutivo. Fonte: Adaptado de Chervy (2002).
1.2.2 Ciclo de vida dos Cestodas da família Proteocephalidae
O ciclo de vida dos parasitos desta Família envolve os crustáceos como hospedeiros
intermediários. A infecção inicia quando o hospedeiro intermediário ingere ovos
infectantes que contêm a larva oncosfera multicelular (Fig. 10). No intestino os ovos
eclodem e liberam as oncosferas que penetram a barreira intestinal, e passam por um
processo de metamorfose para as fases larvares seguintes, chamados de
metacestóides em órgãos-alvo específicos (Fig. 10). A transição segue-se para a fase
de desenvolvimento do protoescólex (uma forma primitiva de escólex do adulto) que
27
eventualmente é retomada por hospedeiros definitivos. Estas transições sofrem
influência do hospedeiro e são eventos chave no desenvolvimento do cestóide (Brehm
et al., 2006).
Figura 10: Ciclo de vida de Cestoda da Família Proteocephalidae. Os adultos
habitam o intestino delgado de peixes e liberam ovos nas fezes que são
embrionados no ambiente aquático. Os copépodes ingerem o ovo contendo uma
larva na forma de oncosfera que eclode na cavidade intestinal e libera a larva
plerocercóide. O copépode pode ser ingerido por um hospedeiro definitivo, onde
continua o desenvolvimento da larva até a fase adulta e completa o ciclo, ou por um
hospedeiro paratênico, onde o parasita não se desenvolve. Neste último caso o ciclo
se completa quando um predador (grandes peixes) se alimenta do hospedeiro
paratênico infectado. Fonte: Laboratório de Biologia Celular e Helmintologia (2013)
1.3 PRODUTOS DE EXCREÇÃO E SECREÇÃO (E/S) DE CESTÓDEOS
Os cestódeos interagem com o ambiente em que vivem (hospedeiro
intermediário ou definitivo) através do tegumento. Este contém uma variedade de
proteínas e enzimas estruturais, algumas produzidas pelo parasito a exemplo de
actina, tubulina, queratina, colágeno e outras adsorvidas dos fluidos do hospedeiro
como colesterol, glucose, glicerol, todas envolvidas na evasão do sistema imune do
hospedeiro pelo parasito, uma vez que o sistema imune reconhece e distingue o
parasito, do hospedeiro, por meio dessas moléculas. Esta evasão pode ser
explicada através de proteínas do glicocálice e da membrana das microtríquias
capazes de proteger a digestão do parasita pelas enzimas proteolíticas do
28
hospedeiro ou pela internalização, conversão e exposição de proteínas na superfície
dos parasitos que mimetizam as proteínas do hospedeiro, fazendo com que o
organismo os reconheça como próprias (Marr et al., 2003).
Apesar dos cestódeos constituírem a terceira maior Classe de helmintos,
estudos relacionados aos seus ciclos de vida e dos produtos de secreção e
excreção são relativamente poucos. No entanto, substâncias excretadas e
secretadas, capazes de mimetizar ou modificar a função das substâncias endógenas
dos hospedeiros são as armas principais de evasão do sistema imune pelos
parasitos. Johnston et al. (2009) reuniram em um quadro os principais cestódeos
descritos na literatura cujos produtos de excreção/secreção condicionam a
imunomodulação, bem como as atividades biológicas desses produtos nas células
dos hospedeiros (Quadro 1).
Além destes, outros trabalhos têm mostrado a relação entre parasitismo por
cestódeos e os locais alvos de seus produtos de E/S com função imunomoduladora.
Como evidenciado por Miura et al. (2000), produtos de E/S de larvas plerocercóides
de Spirometra erinaceieuropaei, possivelmente uma proteína, suprime a expressão
do gene TNF-α e Lipopolissacarídeos (LPS) em macrófagos (Miura et al., 2000).
Carmena et al. (2005), analisaram 12 amostras sorológicas de cães
naturalmente infectados com Equinococus granulosus e identificaram sete
componentes de antígenos de excreção/secreção. Mais tarde, Maity et al.(2007), ao
realizar experimentos com isolados de E. granulosus parasitando búfalos na India
constatou a presença de 7 peptídeos de flúidos do cisto que possuíam homologias
com antígeno B de vertebrados. A Família de antígenos B influencia na migração de
neutrófilos (Siracusano et al., 2008).
Outro aspecto que merece ser considerado é o fato desses parasitos
possuírem sistemas de sinalização semelhantes aos do hospedeiro, e secretam
substâncias com funções idênticas dos mamíferos podendo haver uma comunicação
crucial entre as citocinas secretadas e os receptores correspondentes, tanto do
hospedeiro quanto do parasita, contribuindo para atividades imunomoduladoras ou
favorecendo o tropismo dos parasitos para os órgãos. Um exemplo disso é
demonstrado na presença da secreção de citocinas do tipo EGF e de receptores
para EGF (Spiliotis et al., 2003), filogeneticamente homólogas a outros organismos,
expressados em metacestóides e protoescólex de E. multiloculares. Apesar de ainda
não existir uma comprovação de acoplamento ligante-receptor entre essas
29
moléculas, sabe-se que o gene codificante de EGF é expresso cerca de 10 vezes
mais em vesículas de metacestóide em condições que promovem o crescimento e
diferenciação do parasita (Spiliotis et al., 2003).
Também há relatos de que E. multilocularis expressa receptores sinalizantes
da família de insulina (IGF), que regulam processos metabólicos, controlam o
crescimento, reprodução e envelhecimento (Geer et al., 1994) e evidências para a
sinalização de TGF-ß que atuam no controle da proliferação, diferenciação,
apoptose e regulação imune (Zavala et al., 2003).
30
Quadro 1: Exemplos de moléculas imunomoduladoras de cestódeos e suas funções biológicas em sítios alvos do hospedeiro.
Cestódeo
Produtos E/S
Echinococcus
granulosus
Taenia
multiceps
Taenia
taeniaeformis
Taenia
crassiceps
Taenia solium
Referência
Rakha et al. (1991)
Glicoesfingolipídeos de
metacestóides
Modifica função de macrófagos e inibe a proliferação de
linfócitos in vitro.
Inibe a proliferação de PBMC; diminui a expressão do receptor
de IL-2.
Proteinas 14-3-3 em produtos
de E/S
Inibe a produção de óxido nítrico induzida por LPS em
macrófagos de ratos.
Andrade et al. (2004)
Produto de E/S
Echinococcus
multilocularis
Bioatividade
E4Gala1, 4Galglicanode
extrato de protoscolesces
Antigeno inibidor da
proteaseBno fluido
dahidatidose
Inibe a proliferação induzida por mitogenicos; aumenta IL-10
Persat et al. (1996)
Dematteis et al. (2001)
Inibe recrutamento de neutófilo in vitro; aumenta a produção de
IL-4 e IL-13 pelas PBMCs humanas;
Shepherd et al. (1991)
Rigano et al. (2001, 2007)
Lectinade glicose nosfluidos
císticos
Pode ter função semelhante aos produtos de E/S de
E.multilocularis
Judson et al. (1987)
Proteinase (19,5 kDa)
Inibidor de extrato somático; inibe a proliferação de
esplenócitos em resposta a estímulos mitogênicos e antigênica
Leid et al.(1984, 1986)
Suquet et al.(1984)
Produtos de E/S
Sequestra complemento; diminui a proliferação da produção de
IL-2 induzida por esplenócitos mitogénicos
Aumenta a proliferação e produção de IFNC e IL-10 induzida
por esplenócitos mitógenos; aumenta a produção de óxido
nítrico por macrófagos
Rikishia et al. (1985)
Burger et al. (1986)
Spolski et al. (2002)
Inibe a resposta e a proliferação de antigenos de Salmonella;
inibe a resposta inflamatória a antigenos de metacestóide e
causa diminuição da IL-2, IFNC e IL-4; produção de TNFα
reduzida por macrófagos após a estimulação com LPS
Arechavaleta et al. (1998)
Interferon-c análogo a p66 em
produtos E/S
Peptídeos de RNA em extratos
de metacestóides
Fonte: Adaptado de Johnston et al .(2009).
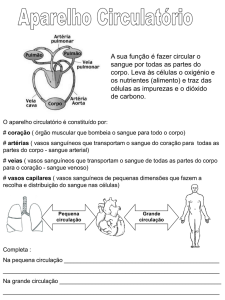
31
1.4 ANGIOGÊNESE
Os vasos sanguíneos constituem uma rede de tubos que transportam o
sangue por todo o organismo distribuindo oxigênio e nutrientes, transportando
hormônios e outros compostos e coletando resíduos metabólicos e materiais tóxicos
para os órgãos excretores (Dias et al., 2003).
Morfológica e funcionalmente, os vasos sanguíneos podem ser divididos em
artérias que são vasos sanguíneos elásticos que transportam o sangue do coração
para os tecidos e, nesta direção, ramificam-se cada vez mais em estruturas de
menor calibre até atingirem o nível capilar e veias que transportam o sangue
proveniente de vários tecidos em direção ao coração, formando ramos de calibre
cada vez maior (Fig.11). As artérias apresentam um espessamento da parede em
relação às veias, em decorrência da sua função pulsatória do sangue, enquanto que
nas veias observa-se um fluxo sanguíneo lento (Dias et al., 2003).
Figura 11: Representação esquemática das estruturas que compõe o sistema
circulatório: A) Veia com camada delgada; B) Arteria com espessamento da
camada; C) Capilar que é a etrutura que faz ligação entre veia e artéria. Fonte:
http://www.cabuloso.xpg.com.br/Anatomia Humana/SistemaCardiovascular/VasosSanguineos.htm, acesso em 05 de dezembro de 2012.
Estruturalmente os vasos são organizados em três camadas: uma túnica
externa ou adventícia composta de tecido conjuntivo e pequenos filetes nervosos e
vasculares; uma túnica média que constitui a camada intermediária composta por
fibras musculares lisas e pequenas quantidades de tecido conjuntivo elástica; e uma
32
túnica íntima ou endotélio, com uma camada única de células endoteliais
interconectadas que reveste a superfície luminal dos vasos revestida pela matriz
extracelular e por camadas de células com função de suporte (Fig. 12) (Dias et al.,
2002).
Figura 12: Ilustração das camadas de revestimento dos vasos sanguíneos. Fonte:
Adaptado de http://www.auladeanatomia.com/cardiovascular/vasos.htm, acesso em
05 de dezembro de 2012.
Quando intacto, o endotélio controla a permeabilidade vascular regulando o
tráfego de moléculas pequenas e grandes e/ou células. Em resposta a fatores ou
sinais específicos, as células endoteliais e as células musculares lisas apresentam
um crescimento desenfreado para formar novos vasos sanguíneos em um processo
conhecido como angiogênese que pode sofrer alterações e desencadear doenças
como a psoríase, cegueira em casos de diabete, artrite e câncer (Dias et al., 2002).
A angiogênese representa uma complexa formação de novos vasos a partir
de células endoteliais diferenciadas de um vaso pré-existente (Yoshida, 2005). Em
condições normais, existe um equilíbrio entre moléculas pro-angiogênicas e antiangiogênicas com favorecimento das moléculas anti-angiogênicas. Contudo quando
o equilíbrio é invertido, devido à ação de fatores metabólicos, mecânicos, mutações
genéticas
e
reações
imuno-inflamatórias,
angiogênese (Fig. 13) (Folkman et al., 2002).
têm-se
o
processo
denominado
33
Figura 13: Processo angiogênico desencadeado pela desestabilização do equilíbrio
das substâncias pró e anti-angiogênicas: A – equilíbrio das substâncias com
favorecimento das anti-angiogênicas verificado em condições normais do organismo;
B – cascata de eventos desencadeado a partir da quebra do equilíbrio entre essas
substâncias com favorecimento das pró-angiogênicas. Fonte: Adaptado de
Quintavalle et al. (2011).
De acordo com Rissaw (1997), durante a embriogênese de vertebrados a
formação
dos
plexos
vasculares
primários
(vasculogênese)
origina-se
do
mesoderma pela diferenciação de angioblastos e subsequente produção dos vasos
sanguíneos primitivos. Estes angioblastos expressam os receptores do fator de
crescimento endotelial vascular (VEGF-R), enquanto que este é produzido e
secretado pelas células do endoderma. A ativação desses receptores pelo VEGF dá
início à diferenciação de angioblastos cuja sobrevivência e manutenção está
condicionada aos níveis de VEGF. Um precursor bipotencial, induz o mesoderma a
expressar o receptor de VEGF do tipo 2 (VEGF-R2) que influencia o processo de
formação do haemangioblasto (desenvolvimento tanto da linhagem hematopoiética
quanto da linhagem angioblástica), levando a formação dos primeiros vasos
sanguíneos (Fig. 14).
34
Figura 14: Esquema ilustrativo dos tipos de angiogênese que podem ocorrer a partir
de plexus capilares primitivos ou vasculogênese. Fonte: Adaptado de Rissaw (1997).
Após a formação do plexo vascular primário, muitas células endoteliais são
regeneradas e podem formar novos capilares através de duas fases: de brotamento
e de maturação. O brotamento pode ocorrer através do processo denominado
angiogênese verdadeira (formação de capilares de vasos pré-existentes), ou pela
junção de dois vasos que se romperam, formando um só. Posteriormente, ocorre a
remodelagem do plexo vascular formado, determinada por sinais que suprimem o
VEGF, fator de sobrevivência das células endoteliais, acarretando a morte dessas
células. Em seguida ocorre a fase de maturação, originando um sistema vascular
maduro (Rissaw, 1997; Yoshida, 2005).
A angiogênese normal ocorre durante o desenvolvimento embrionário e é
observada, ao longo da vida adulta, em alguns eventos fisiológicos, tais como:
ovulação, ciclo endometrial, desenvolvimento da placenta, reparação de feridas,
constituindo
assim
um
importante
papel
nos
processos
de
reprodução,
desenvolvimento e reparação tecidual (Fechine-Jamacaru et al., 2005).
A angiogênese também pode ser observada em eventos patológicos como no
câncer, no desenvolvimento da placa de ateroma e em doenças cujas células
35
endoteliais têm participação importante, como na psoríase, retinopatia diabética,
degeneração macular, artrite reumatóide, angiofibroma, hemangioma, glaucoma
vascular (Malucelli et al., 2002).
Recentemente a angiogênese também vem sendo relatada em infecções por
helmintos como, por exemplo, em infeções por S.mansoni (Loeffler et al., 2002;
Kanse et al., 2005; Lemos et al., 2010; Dennis et al., 2011), filárias (Esterre et al.,
2005), Spirocerca lupi (Dvir et al., 2010), Taenia solium (Sikasunge et al., 2009)
entre outros.
A angiogênese em vertebrados na fase pós-embrionária, pode ser resumida
nas seguintes etapas: a) aumento da permeabilidade vascular e depósito de fibrina
extra-vascular; b) desestruturação da parede vascular, com remoção dos pericitos;
c) desestruturação da membrana basal e da matriz extracelular; d) migração da
célula endotelial através da matriz extracelular remodelada; e) proliferação da célula
endotelial; f) formação de estruturas tubulares; g) inibição da proliferação e migração
da célula endotelial; h) reconstituição da membrana basal; i) maturação dos
complexos juncionais; j) construção da parede vascular, através do recrutamento e
diferenciação de células murais, pericitos e células de músculo liso e k)
estabelecimento de fluxo sanguíneo no novo vaso (Fig. 15) (Lemos et al., 2010).
Nesse caso, a estimulação desse processo é decorrente de outros fatores,
diferentes daqueles da embriogênese, como é o caso da hipóxia e da presença de
integrantes da família do fator de crescimento endotelial.
36
Figura 15: Esquema ilustrativo do processo de migração e proliferação das células endoteliais
para formar novos vasos sanguíneos. A) Mecanismos de estimulação das células endoteliais
através da produção de fatores de crescimento; B) Processo de alongamento e direcionamento
das células endoteliais (EC); C) Formação e maturação do novo vaso. Fonte: Adaptado de
Carmeliet et al. (2011).
37
1.4.1 Fatores de Indução da Angiogênese
Diferentemente da embriogênese, o processo angiogênico em indivíduos
maduros surge em decorrência da liberação de diversas proteínas como: fator de
crescimento fibroblástico básico (bFGF), fator de crescimento endotelial vascular
(VEGF) e enzima ciclo-oxigenase-2 (COX-2), que são desencadeados por fatores
locais estimulantes como a hipóxia e a elevação de CO2 ou óxido nítrico (Fig. 16)
(Pinho, 2005).
Figura 16: processo de indução da proliferação celular e formação de
vascularização em eventos patogênicos, mediado por diversos fatores em resposta
a fatores locas estimulantes, a exemplo da hipóxia. COX-2 = Ciclo Oxigenase 2;
VEGF = Fator de Crescimento Endotelial vascular; FGF = Fator de Crescimento
Fibroblasto; CO2 = Dióxido de Carbono; NO = Óxido Nítrico. Fonte: Adaptado de
Bikfalvi (2012).
Moraes (2008), ao realizar experimentos com células endoteliais de aorta de
coelho, elucidou o papel do óxido nítrico no processo da angiogênese. O óxido
nítrico, substância mediadora da inflamação, fosforila os receptores EGFR
provocando a liberação de VEGF. Além disso, esses autores observaram que a
bradicinina aumenta a fosforilação em resíduos de tirosina de EGFR similarmente à
produção do óxido nítrico.
A hipóxia em resposta às mudanças do meio ambiente, aciona mecanismos
de adaptação que levam a alterações ao nível sistêmico tecidual e celular que
tendem a reduzir o consumo de oxigênio e melhorar a oferta deste. Entre essas
adaptações alternativas adotadas pelo organismo encontra-se a angiogênese, pela
38
indução do VEGF. A hipóxia leva a um aumento da transcrição do VEGF ao nível de
RNA mensageiro por meio da estimulação da atividade do fator 1 de indução de
hipóxia (HIF-1) , que se liga a promotores de VEGF e forma um complexo que
resulta na transcrição do gene de VEGF (Fig.17) (Levy et al., 1995).
Figura 17: Mecanismo de sinalização do VEGF nas células endoteliais. Em
condições de injúria tecidual como a hipóxia, o gene de VEGF é expresso e
transcrito. Após transcrição, este se liga aos seus respectivos receptores e estimula
a migração das células endoteliais e irrigação vascular no sítio afetado. HIF-1α =
Fator de Indução da Hipóxia 1 alfa; VEGF = Fator de Crescimento Endotelial
Vascular; sFIt = gene de transcrição do receptor solúvel 1 de VEGF; VEGFR1 (Fit1)
= receptor do Fator de Crescimento Endotelial Vascular 1; VEGFR2 (KDR) =
receptor do Fator de Crescimento Endotelial Vascular 2; PLCy = Phospholipase Cgamma; PKC = Proteina Kinase C; MAPK = Proteína Kinase ativado por Mitógeno;
Fonte: Adaptado de Bikfalvi (2012).
Além disso, a hipóxia pode também mudar a abundância de micro RNAs em
células endoteliais, como por exemplo a produção de miR-210, e promover a
migração de células endoteliais e formação de capilares e estruturas semelhantes.
MIR-210 tem atividade pro-angiogênica devido seu direcionamento de ephrin A3 que
é dependente do VEGF (Quintavalle et al., 2011).
39
O VEGF é o fator principal no processo angiogênico, e um regulador crucial
do desenvolvimento vascular durante embriogênese (vasculogênese) e formação
dos vasos sanguíneos (angiogênese) nos adultos (Fig. 18) (Olsson et al., 2006).
Esta molécula foi isolada pela primeira vez do líquido ascítico de cobaias e
primeiramente foi denominado como fator de permeabilidade vascular (VPF) (Senger
et al., 1986). Hoje se sabe que o VEGF é liberado por células endoteliais,
fibroblastos, plaquetas, neutrófilos e macrófagos, e tem expressão acentuada em
situações de hipóxia (Bao et al., 2008).
Figura 18: Mecanismo de liberação e ligação do fator de crescimento endotelial
vascular (VEGF) desencadeando a formação de novos vasos sanguíneos.Fonte:
Adaptado de https://www.Vascular_endothelial_growth, acessado em março de
2013.
A biologia do VEGF é complexa, pois existem seis variantes deste fator de
crescimento: VEGFA, B, C, D, E e o fator de crescimento placentário (PLGF). Estas
variantes podem interagir com um dos três receptores tirosino-kinase específicos
para VEGF e conduzir a angiogênese. O VEGFA, B e PLGF ligam-se ao receptor
VEGFR1; VEGFA e E ligam-se ao receptor VEGFR2, e VEGFC e D ao receptor
VEGFR3 (Fig. 19) (Ferrara, 2004; Takahashi et al., 2005).
40
Figura 19: Variantes do fator de crescimento endotelial vascular e os possíveis
receptores, aos quais estas substâncias se ligam para desencadear eventos
angiogênico. Fonte: Adaptado de Cao (2009).
Além da atividade angiogênica, o VEGF também apresenta atividade próinflamatória e neuroprotetora. Suas funções incluem a quimiotaxia para leucócitos e
indução da permeabilidade vascular, promovendo a desorganização das zônulas de
oclusão e retração das células endoteliais (Ferrara et al., 1996; Bao et al., 2008).
1.4.2 Helmintos e angiogênese
Muitos helmintos parasitos estimulam a formação de vascularização no
hospedeiro (angiogênese), como um dos pré-requisitos para o sucesso no
crescimento parasitário destacando a maturidade e a reprodução. Este fenômeno
pode ocorrer nos tecidos do hospedeiro tanto por um mecanismo de reparo de dano
tecidual causado pelo parasito, quanto por moléculas pró-angiogênicas derivados
dos parasitos para benefício próprio (Fig. 20). No sistema vascular este processo
pode ocorrer através de uma co-adaptação, de forma ativa ou passiva, do sistema
vascular do hospedeiro na região intra-vascular e ou linfático (Dennis et al., 2011).
41
Figura 20: Esquema ilustrativo da angiogênese desencadeada por parasitos
helmintos. A) Fatores de crescimento endotelial vascular A ou C (VEGF-A-C)
derivados de helmintos ou fatores proangiogênicos do hospedeiro; B) Ativação de
células endoteliais (CEs) por esses fatores; C) Formação da vascularização
parasitária através da penetração da membrana basal, formação de uma matriz
provisória, remodelação da matriz extracelular, ativação do sistema plasminogênio–
plasmina, proliferação, migração, brotamento e ramificação das células endoteliais
(CE) produzindo um sistema vascular imaturo e instável; D) Interação entre as
células endoteliais e os pericitos, estabilizando os vasos e auxiliando na maturação
da vascularização parasitária. Fonte: Adaptado de Dennis et al. (2011).
Um dos processos angiogênicos induzidos por helmintos mais bem estudados
é o processo desencadeado pelo S.mansoni. Durante as etapas do ciclo evolutivo, o
parasito passa por diferentes tecidos e vai adquirindo alterações morfológicas e
bioquímicas significantes (Souza, 2011).
O estabelecimento da patogenia da esquistossomose está ligado a vários
fatores como espécie do parasito, estado nutricional, idade e resposta imunológica
do hospedeiro e a carga parasitária adquirida que é mensurada pela contagem de
ovos por grama de fezes. Os ovos são os elementos fundamentais para a patogenia
da esquistossomose, pois em números elevados atingem o fígado e podem provocar
alterações como hemorragias, edemas de submucosa e fenômenos degenerativos
que são reparadas através da reconstituição da integridade dos tecidos. Por outro
lado a resposta imunológica do paciente frente aos antígenos dos esquistossômulos
e dos ovos leva a alterações cutâneas e hepáticas (Rey, 2005).
Segundo Loeffler et al. (2002) e Kanse et al. (2005) os ovos de S.mansoni
secretam
fatores
angiogênicos
que
ativam
células
endoteliais.
Em
seus
experimentos in vitro, antígenos solúveis de ovos de S.mansoni, desprovidos de
lipopolissacarídeos, adicionados a cultura de células endoteliais de veia umbilical
42
humana contendo soro bovino fetal, apresentaram uma proliferação celular
significativamente maior do que cultura de células endoteliais de veia umbilical
humana contendo apenas soro bovino fetal. Ao substituirem esses antígenos por
fatores de crescimento, a proliferação celular continuou. Estes achados mostraram
que os produtos de secreção do ovo de S.mansoni podem contribuir para a ativação
e proliferação de células endoteliais e, subsequentemente, promoção da resposta
inflamatória, formação de granuloma e vascularização através de um mecanismo de
liberação de fatores de crescimento.
A liberação desses fatores pró-angiogênicos foi verificado no trabalho de Kanse et
al. (2005) os quais demonstraram que a presença de ovos vivos de S.mansoni em
cultura de células endoteliais marcadas com microcarregadores, estimula a
neovascularização, da mesma forma que cultura de células tratadas com
substâncias estimuladores de crescimento endotelial (Fig. 21).
Figura 21: Estimulação das células endoteliais e formação de capilares. A)
Estimulação por ovos vivos de S.mansoni; B) Estimulação por uma solução
contendo fatores de crescimento endotelial. Fonte: Adaptado de Kanse et al. (2005).
Shariati et al. (2011), também constataram a presença de VEGF em amostras
sorológicas de pacientes com diagnóstico de esquistossomose, assim como a
produção de VEGF e FGF2 por macrófagos alveolares estimulados pelos antígenos
de cercária de S.mansoni. Estes autores também verificaram a redução de vermes,
ovos e granulomas no fígado de ratos infectados com S.mansoni após estes serem
tratados com endostatina, um inibidor da angiogênese.
Estimulação e proliferação de células endoteliais também pode ocorrer por
mecanismos secundários, através da super expressão de substâncias endógenas
como integrina ß8 e galectinas 1 e 3 por macrófagos ativados com extrato somático
43
de ovos de Schistosoma (Zaccone et al., 2013). Dependendo do grau de expressão,
a integrina ß8 tem papel comprovado na angiogênese ou na capacidade de invasão
de células tumorais (Tchaicha et al., 2011) e as galectinas, além de modularem a
resposta imune contra os parasitos, estimulam a migração das células endoteliais
(Fig. 22) e, consequentemente, a angiogênese (Alves, 2009).
Figura 22: Papel das galectinas 1 e 3 na imunomodulação do sistema imune do
hospedeiro e na indução da proliferação celular e angiogênese, desencadeado pelos
patógenos; a-d eventos desencadeados pela galectina 1 e e-i são eventos
desencadeados pela galactina 3 ambos nos compartimentos intra e extracelular: a)
associação de galectina 1 a H-RAS e indução de crecimento tumoral no espaço
intracelular; b) indução de apoptose de células do sistema imune como Th1 e Th17;
c) inibição de migração transendotelial de células Th1; d) liberação de galectina 1
pelas células endoteliais e promoção de angiogênese na luz dos vasos sanguíneos;
e) associação de galectina 3 a K-RAS e indução de P13K que por sua vez induz
crescimento tumoral no espaço intracelular; f) inibição de apoptose pela galectina 3
no espaço intracelular; g) indução de Cyclin D1, c-MYC e crescimento tumoral por
galectina 3 associada a β-Catenin; h) inibição de galectina e crescimento tumoral e
indução de apoptose pelo p53 i) indução da angiogênese por galectina 3 na luz de
vasos sanguíneos. Fonte: Adaptado de Yang et al. (2008).
44
Acredita-se que os pericitos (células mesoteliais que envolvem as células do
endotélio capilar e vênulas) estão envolvidos no processo angiogênico verificado na
esquistossomose, não só pela sua capacidade contrátil e plasticidade na formação
de fibrose, mas também como reguladores obrigatórios do desenvolvimento
vascular, estabilização da maturação e remodelação (Andrade et al., 2010).
Além do Schistosoma, processos de formação de vasos sanguíneos ou
produção de substâncias pelos parasitas, como os fatores de crescimento e
proteínas, com papel importante no desenvolvimento deste processo vem sendo
relatado em outras espécies de helmintos e tem ganhado relevância não só pelos
danos causados no hospedeiro, mas também como forma de inibição das funções
vitais do parasito através do entendimento dos mecanismos de liberação de
substâncias pro-angiogênicas e desenvolvimento de drogas que culminam na morte
do parasita.
Segundo Esterre et al. (2005), VEGF-A (responsável pelo extravasamento do
sangue e linfa para os tecidos circundantes) e VEGF-C (responsável pela dilatação
linfática) podem ser importantes fatores angiogênicos no desenvolvimento de
linfedema e hidrocele em filariose linfática e a associação destas tem efeito sinérgico
na indução de angiogênese.Isto foi comprovado por Debrah et al. (2007), quando
verificaram que a presença ou a morte natural de vermes adultos de filária na região
escrotal de pessoas infectadas leva a liberação de antígenos de filária e Wolbachia,
que induzem a super-expressão de VEGF-A e VEGF-C. Estes seriam os
responsáveis pelo extravasamento e acúmulo de líquidos a partir dos vasos
sanguíneos e linfáticos, levando à formação de hidrocele.
Jo et al. (2010) descobriram que timosina b4, uma proteína multifuncional
anti-inflamatória expressada em locais inflamatórios, induz a expressão de VEGF
pela crescente estabilidade da proteína HIF-1. Um ano mais tarde Kang et al. (2011),
realizaram infecção experimental de camundongos com Triquinela spiralis e apesar
de não constatarem a ocorrência de hipóxia nas fibras musculares infectadas
verificaram uma expressão elevada de timosina b4 na fase tardia da infecção
sugerindo que a infecção por T.spiralis induz a expressão desta proteína que
estimula VEGF levando a angiogênese.
Dvir et al. (2010), avaliou a expressão de fatores de crescimento em
espirocercose canina (Spirocerca lupi) onde constatou a expressão de VEGF, FGF e
Fator de Crescimento derivado de Plaquetas (PDGF) e, apesar de não esclarecer se
45
estas proteínas eram produzidas nos nódulos ou vinculadas aos receptores após
serem secretadas por células inflamatórias, endotélio ou pelo próprio parasita,
concluiu que a expressão desses fatores está associada aos nódulos de
espirocercos.
1.4.3 Helmintos cestodas e angiogênese
Os cestódeos e helmintos em geral, necessitam de um sistema que lhes
ofereça oxigênio, nutrientes, hormônios e fatores de crescimento necessários para a
sua colonização no hospedeiro parasitado. Além disso, este sistema também auxilia
na remoção de resíduos metabólicos para o meio externo ao parasita. Para isso
estes seres desenvolvem mecanismos para o estabelecimento de plexos capilares a
partir de outros já existentes nesses hospedeiros, e que podem o acompanhar
durante toda a sua existência (Dennis et al., 2011).
Estudos sistemáticos de cestódeos relacionados com a formação de vasos
sanguíneos nos tecidos dos hospedeiros são relativamente escassos. Os primeiros
relatos da literatura foram voltados para a fase larval metacestóide de
Mesocestoides corti presente em camundongos (hospedeiro intermediário deste
parasita) e de cisticercos de Taenia solium, infectante tanto para porcos quanto para
os seres humanos (Dennis et al., 2011).
Mesocestoides corti tem aspectos do ciclo de vida semelhante a T. solium, em
que ambos necessitam de hospedeiros intermediários para o desenvolvimento de
suas fases larvares e estas, posteriormente, quando ingeridas por um hospedeiro
definitivo desenvolvem-se em tênias intestinais capazes de reproduzir, liberar ovos e
manter o ciclo. Apesar de não haver relatos de Mesocestoides corti desenvolvendo
a infecção no sistema nervoso central dos seus hospedeiros, quando a infecção é
contraída pela via fecal-oral, tal como ocorre com T. solium, este parasito tem
servido como modelo de estudo laboratorial para compreensão dos diferentes
aspectos da interação parasito-hospedeiro através da injeção direta do parasito no
cérebro de camundongos (Alvarez et al., 2009).
Na maioria das vezes, metacestódeos de M. corti encontram-se envoltos por
tecido conjuntivo, e o seu crescimento está associado com a formação de novos
vasos sanguíneos ao redor do parasito, importantes interfaces para a captação de
46
fatores nutricionais, bem como mediadores de sinais que regulam a relação parasitohospedeiro (Stadelmann et al., 2010).
Alvarez et al. (2002), ao analisar o tecido nervoso humano na proximidade de
lesões granulomatosas induzidas por metacestóides de T. solium, em cinco regiões
cerebrais diferentes, mostrou que as células cerebrais adjacentes exibiram
moléculas pró-inflamatórias, como IFNу e IL-18, com atividade associada a células
da microglia/macrófagos e componentes da barreira hematoencefálica (células
endoteliais e / ou pericitos). Além disso, destacaram o efeito imunomodulador
através da produção de citocinas anti-inflamatórias (TGF e IL-10) e uma atividade
de reparo tecidual promovido pelo bFGF (substância vasoativa e profibrótica) devido
à presença de uma cicatriz glial e deposição de colágeno, evidenciando a ocorrência
do processo angiogênico.
Sikasunge et al. (2009), mostraram que o tecido cerebral de 30 suínos
infectados com cisticercos de T. solium apresentaram astrogliose (aumento de
astrócitos em número e tamanho), extensa formação de cicatriz glial e fibrose e dano
neuronal e axonal a partir de fibras nervosas dilatadas, aumento do número de
vasos sanguíneos e da permeabilidade da barreira hematoencefálica. Estes
aspectos evidenciaram claramente a angiogênese nesses tecidos.
Segundo Gottstein & Hemphill (2008), os metacestóides de Equinococus
multilocularis possuem dois fatores distintos, ambos sintetizados na camada
germinativa, que podem ser os principais agentes envolvidos no processo de evasão
do sistema imune: a camada acelular altamente glicosilada e laminada formando um
glicocálix estendida sobre a superfície do parasita e produtos de excreção/secreção
(E/S) dos parasitos encontrados no líquido das vesículas ou no ambiente exterior e
que podem influenciar a resposta imune do hospedeiro e / ou processos fisiológicos.
Mais tarde, Stadelmann et al. (2010) realizaram cultivo in vitro de
metacestóides de E.multilocularis, extraíram células das vesículas do parasito, e
identificaram uma proteína de E/S denominado EmPGI (fosfoglicose isomerase de
E.multilocularis) que atua na proliferação celular e / ou diferenciação tecidual. Além
disso, foi detectado expressão de RNA para EmPGI em metacestóides, protoescólex
e em culturas primárias de células da camada germinativa do parasito.
Não existem relatos na literatura do desenvolvimento do processo
angiogênico por cestóides em peixes. Porém estudos realizados por Fouquet et al.
(1997), Pichler et al. (2003), Lam et al. (2006) e Tobia et al. (2011) com embriões de
47
Danio rerio (peixe-zebra), um teleósteo da Ordem Siluriforme, mostram que este
peixe tem grande potencial para ser usado futuramente como um modelo
angiogênico de câncer humano.
Assim o uso de peixes como D.rerio representa uma ferramenta interessante
para testes de bioensaio de moléculas pequenas e dissecção das vias biológicas
como alternativa a outros modelos de vertebrados, devido as suas inúmeras
vantagens, como a facilidade de experimentação e a administração de fármaco bem
como a receptividade na manipulação in vivo.
O
entendimento
imunomoduladoras
e
dos
princípios
indutoras
de
básicos
processo
de
ação
angiogênico,
de
substâncias
associado
aos
mecanismos de liberação dessas substâncias em órgãos-alvos poderão contribuir
para o desenvolvimento de novos agentes terapêuticos.
Deste modo, pesquisas básicas sobre a relação parasito-hospedeiro entre
helmintos e peixes podem contribuir com informações importantes nos processos
patológicos em demais vertebrados e em humanos.
Em virtude da constatação macroscópica da presença de cistos de cestódeos
da Família Proteocephalidae associada com intensa formação de vasos sanguíneos
ao seu redor e por entre esses cistos, na cavidade celomática de Ageneiosus
ucayalensis e, da escassez de informações sobre a interação de parasitos
cestódeos
com
seus
hospedeiros,
objetivou-se
caracterizar
morfológica
e
ultraestruturalmente o processo angiogênico visualizado na interação parasito
hospedeiro entre estes parasitos e Ageneiosus ucayalensis.
48
1.5 OBJETIVOS
1.5.1 Objetivo geral
Caracterizar os aspectos estruturais e moleculares elementares de larvas de
Cestoda da família Proteocephalidae parasitos de Ageneiosus ucayalensis e os
aspectos do parasitismo destas larvas.
1.5.2 Objetivos específicos
a)
Caracterizar
por
morfologia
e
sequências
moleculares
os
metacestódeos envolvidos no parasitismo de Ageneiosus ucayalensis
b)
Descrever por meio de técnicas histológicas e de ultraestrutura o
processo angiogênico da relação de metacestódeos proteocephalídeos e A.
ucayalensis.
c)
Relacionar as estruturas teciduais neo-formadas à presença das larvas
de Cestódeos.
49
2.
MATERIAL E MÉTODOS
2.1
COLETA DE HOSPEDEIROS E AMOSTRAS DE PARASITOS CESTODAS
Um total de 40 exemplares de Ageneiosus ucayalensis foram adquiridos ainda
vivos, através da pesca artesanal com isca e anzol, de pescadores ribeirinhos da
baia do Guajará no Município de Belém, Estado do Pará (01º24'S e 48º28'W),
conduzidos ao Laboratório de Biologia Celular e Helmintologia do Instituto de
Ciências Biológicas da Universidade Federal do Pará, para realização da necropsia
e pesquisa de parasitos cestódeos. Durante a necropsia, tecidos e órgãos foram
removidos para placas de Petri com solução de tampão fosfato salino (PBS), a pH
7,4 e analisados através de estereomicroscópio (Carl Zeiss modelo 475032) para
retirada dos helmintos e vasos sanguíneos associados. A partir daí, os
procedimentos para fixação, processamento e posterior análise de acordo com as
metodologias empregadas decorreram como mostra o organograma a seguir:
50
2.2 MICROSCOPIA DE LUZ
2.2.1 Análise taxonômica de Cestoda
As larvas de Cestoda fixadas com uma solução de ácido acético glacial,
formol a 37% e álcool etílico a 70% (AFA) sob compressão entre lâmina e lamínula
foram utilizadas para análise direta ao microscópio de luz. Após a fixação,
desidratação em série etanólica de concentrações crescentes (Etanol 70%, Etanol
80%, Etanol 90%, Etanol 100%) por 10 minutos em cada etapa, os cistos foram
corados com Carmim alcoolico, clarificados com salicilato de metila e montados com
Entellan® entre lâmina e lamínula segundo Moraes Neto et al. (1998).
Posteriormente foram analisados em Microscópio Olympus BX 41 equipado com
câmara clara (sem ajuste de Zoom) no LBCH – ICB – UFPA, para realização dos
desenhos e identificação dos caracteres taxonômicos.
2.2.2 Análise histológica pelo método de inclusão em historesina®
Algumas amostras de cistos contendo formas larvares de Cestoda e vasos
adjacentes foram fixados em solução de ácido acético glacial, formol a 37% e álcool
etílico a 70% (AFA). Após período mínimo de 24 horas de fixação, foram
desidratadas em série etanólica crescente (30%, 50%, 70%, 80%, 90%, e 100%) por
30 minutos em cada etapa.
Após a desidratação realizou-se a infiltração em resina, utilizando EtanolHistoresina® em diferentes proporções: 5:1, 4:1, 3:1, 2:1, 1:1 e Historesina® pura.
Cada etapa de infiltração teve uma duração de 24 horas.Para inclusão, os cistos
contendo as formas larvares foram inseridos em placas de nove poços, em molde de
inclusão preenchidos com uma solução de Historesina®-polimerizador em uma
proporção 15:1, de acordo com o protocolo do fabricante LEICA®.
Após 2 horas de repouso em temperatura ambiente até o início da
polimerização da resina, os moldes foram levados a uma estufa a 40 ºC por três
dias, para a finalização da polimerização.
Os blocos polimerizados foram então seccionados de forma seriada, com
cortes de 3 μm de espessura, com navalhas de vidro, em ultra micrótomo LEICA EM
UC6 no Laboratório de Biologia Estrutural – ICB – UFPA. Os cortes seriados foram
capturados um a um e então colocados ordenada e sequencialmente, em lâminas de
51
microscopia; corados diretamente com Azul de Toluidina 1% levemente aquecido;
lavados em água corrente, secados, montados em Entellan®, analisados e
fotografados serialmente em microscópio Olympus BX 53 equipado com sistema
capturador de imagem.
As imagens capturadas foram processadas e alinhadas no programa
RECONSTRUCT® Versão 1.1.0.0 1996-2009, desenvolvido pelo Institutes of Health
and the Human Brain Project.
2.2.3 Análise histológica pelo método de inclusão em parafina
Após a fixação prévia em AFA, as amostras foram lavadas e desidratadas em
concentrações crescentes da bateria etanólica: 70%, 80%, 90% e 100%, por 30
minutos cada, seguida da diafanização em três sucessivos banhos de Xileno durante
20 minutos cada. A infiltração foi realizada com banhos de parafina líquida em estufa
a 60 ºC e a inclusão ocorreu à temperatura ambiente. Cortes obtidos pela
microtomia foram desparafinizados em xileno, hidratados em concentrações
decrescentes da bateria alcoólica (100%, 90%, 80% 70% 50% e 30%) e lavados
com água destilada. Em seguida foram corados para análise histológica e também
processados para análise histoquímico, através das colorações de Orceína,
Tricrômios de Malllory e Gomori e pelo Ácido Periódico de Schiff.
2.3 BIOLOGIA MOLECULAR
2.3.1 Extração de DNA e sequenciamento para Taxonomia e Filogenia
O DNA genômico total foi extraído a partir de pools de larvas pequenas e
pools de larvas grandes, previamente preservada em etanol a 70% a – 4 ºC, pela
técnica do fenolclorofórmio segundo o protocolo descrito por Sambrook et al. (1989).
Seguindo este protocolo, os parasitos foram lisados em 300 µL de tampão de
homogeinização, 300 µL de tampão de lise celular, 15 µL de Proteinase K (10
ng/mL), 15 µL de RNase (10µg/mL) e incubados a 56 ºC durante 1 hora.Após
completa digestão, as proteínas foram separadas do restante do material a partir da
adição de 600 µL de fenolclorofórmio alcool - isoamil (25 : 24 : 1) e lavagem com 600
µL de clorofórmio álcool isopropílico (24:1).O DNA foi precipitado com 100µL de
acetato de sódio (3M, pH 4,8) e 500 µL de álcool isopropanol sob -20 ºC durante 15
52
minutos. Após congelamento, as amostras foram centrifugadas por 10 minutos, o
sobrenadante descartado e o pellet de DNA lavado com 500 µL de etanol a 70%.O
DNA resuspenso em 50 µL de tampão de eluição (TE) foi armazenado a -20 ºC até a
reação em cadeia da polimerase.
A subunidade menor do rDNA (18S) foi amplificada através da reação em
cadeia
da
polimerase,
utilizando
o
par
de
primers
Ces1
(5-’
CCAGCAGCCGCGGTAACTCCA-3') e Ces2 (5'- CCCCCGCCTGTCTCTTTTGAT-3')
que amplificam um fragmento de aproximadamente 400 bp. O volume final da
reação foi de 25 µL, contendo 6mM de MgCL2, tampão 10X (20 mM de Tris HCL pH
8.4 e 50mM de KCL), 0.2 pmol/µL de dNTP, 1U de taq polimerase, 0,4 pmol/µL de
cada primer e 10ng de DNA. A reação ocorreu sob as seguintes condições:
desnaturação do DNA a 95 ºC durante 2 min e 30 ciclos de amplificação (95 °C, por
1 min, 65 °C por 1 min, 72 °C por 2 min) e uma etapa de extensao final a 72 ºC por 5
min.
Os produtos de PCR foram purificados com Exosap - IT® (Affymetrix®),
clonados no vetor pGEM®-T (Promega®) e introduzidos em Escherichia coli Top 10
(Invitrogen®). O sequenciamento ocorreu com ambos os iniciadores forward (Ces 1)
e reverse (Ces 2), cujas variações térmicas consistiram em desnaturação inicial de 2
min a 95 ºC, seguido de 35 ciclos de 1min a 95 ºC, 1 min a 50 ºC e 2 min a 72 ºC e
uma fase final de extensão a 72 ºC por 5 min, com auxílio de um analisador
automático DNA modelo ABI 3130 (Applied Biosystem ®) em conjugação com BigDye
Terminator V3.1 Kit, de acordo com as instruções do fabricante.As sequências
obtidas foram editadas no BioEdit (Hall, 2007).
Para a análise filogenética, a distância P não corrigidao foi avaliada pelo
programa PAUP (Swofford, 2002) e a análise de Máxima Parcimônia (MP) baseada
em um algorítimo de busca heurística foi realizada no MEGA com 1000 réplicas de
bootstrap para avaliar o nível de confiança dos nós definidos por esta análise. A
inferência Baesiana (BI) Mr Bayes 3.1.2 (Ronquist & Huelsenbeck, 2003) foi
realizada com o banco de dados rodados utilizando 5.000.000 gerações com
freqüência de amostragem de uma árvore a cada 1000 em duas corridas
independentes com quatro cadeias cada e os resultados das primeiras 1000 árvores
descartadas como burn-in. As demais árvores foram utilizadas pelo programa Mr
Bayes para estimar a probabilidade posterior de cada nó na reconstrução
filogenética.
53
2.4 MICROSCOPIA ELETRÔNICA DE TRANSMISSÃO
Após a fixação inicial em Glutaraldeido 2,5% em tampão Cacodilato de Sódio
0,1 M a p.H 7.2, as amostras de cistos foram lavadas em tampão e pós-fixadas em
tetróxido de ósmio (OsO4) por 2 horas.Em seguida foram lavadas com tampão e
desidratados em concentrações crescentes de acetona (30%, 50%, 70%, 80%, 90%,
e 100%) com uma duração de 30 minutos em cada etapa.
Após a desidratação foi realizada a fase de infiltração em resina epóxi,
utilizando o kit Epon® (Luft, 1961). Os cistos foram infiltrados em concentrações
crescentes de epon, iniciando em solução acetona + epon, na proporção 5:1 até
epon puro por 24 horas em cada etapa. A inclusão ocorreu à temperatura ambiente
e a polimerização em estufa à temperatura de 60 ºC por 72 horas.
Os blocos polimerizados foram então cortados em secções semi-finas de 1
µm de espessura, com navalhas de vidro, para análise prévia e secções ultrafinas de
70 nm foram feitas em ultra micrótomo LEICA EM UC6 no Laboratório de Biologia
Estrutural – ICB – UFPA.Os cortes ultrafinos foram capturados por grades de 300
mesh, contrastados com uranila, lavados em solução de citrato de chumbo e
analisados em microscópio eletrônico de transmissão para identificação dos
constituintes celulares.
2.4.1 Citoquímica
Para a análise citoquímica, foi empregada a técnica de Vermelho de Rutênio
para localização de componentes ácidos, associados à superfície celular e à matriz
extracelular. As amostras previamente fixadas em Glutaraldeido 2,5% em tampão
cacodilato de sódio 0,1 M a p.H 7.2, contendo 0,5mg/mL de Vermelho de Rutênio,
foram lavadas em tampão Cacodilato de Sódio 0,1M adicionado de 0,5mg/mL de
Vermelho de Rutênio, pós-fixadas em OsO4 1% com tampão Cacodilato de Sódio
0,1M acrescido de 0,5mg/mL de Vermelho de Rutênio e lavadas novamente no
mesmo tampão contendo 0,5mg/mL de Vermelho de Rutênio para retirar o excesso
do segundo fixador (OsO4). Ambas as lavagens no tampão foram realizadas por
duas vezes com duração de 10 minutos em cada etapa.Após esse procedimento,
seguiu-se as etapas de desidratação, infiltração, inclusão e microtomia, conforme
descrito no procedimento de rotina e a visualização ao Microscópio Eletrônico de
Transmissão, sem contrastação prévia.
54
2.5 MICROSCOPIA ELETRÔNICA DE VARREDURA (MEV)
Os cistos e fragmentos de tecidos parasitados com cistos contendo formas
larvares de Cestódeos e vasos sanguíneos foram fixados em AFA, pós-fixados em
OsO4 a 1%, e
lavados em tampão fosfato 0,1M pH 7,4.Em seguida foram
submetidos à desidratação em serie crescente de Acetona a partir de Acetona 30%,
até o ponto crítico do CO2. Os helmintos desidratados foram então montados
cuidadosamente em ―stubs‖, metalizados com ouro e analisados ao Microscópio
Eletrônico de Varredura JEOL JSM 5600.
55
3 RESULTADOS
3.1 AMOSTRAS DE CESTÓDEOS PARASITOS
A prevalência do parasitismo foi observada em 93,3% dos peixes coletados.
Durante a necropsia foi possível observar uma massa de cistos, delimitada por um
saco de tecido conjuntivo aqui denominado de ―bolsa cística‖, com os parasitos
ainda vivos e distribuídos na cavidade abdominal de Ageneiosus ucayalensis.
Macroscopicamente esta bolsa cística não mostrou evidências de alterações
fisiológicas e, nem danos anatômicos ao hospedeiro, podendo passar despercebido
por possuir textura semelhante aos órgãos deste hospedeiro.
A bolsa cística apresentava vários cistos agrupados, com formato esférico a
ovalado, escólex invaginados e tamanhos diferentes, característico de larvas
Merocercóides segundo critérios de classificação estabelecidos por Chervy (2002).
Vasos sanguíneos de pequeno e médio calibre se estendem e se
entremeiam por toda a extensão da massa cística, circundando, interconectando e
irrigando os cistos, assim como o tecido que os envolve. Esses pequenos e médios
vasos se conectam com um vaso de grande calibre que emerge de órgãos como o
fígado, ou outro órgão altamente irrigados e que estejam nas proximidades onde a
massa cística se instalou após a infecção (Fig. 23).
56
Figura 23: Hospedeiro Ageneiosus ucayalensis e a localização e disposição dos
cistos de cestóides e os vasos sanguíneos dentro do hospedeiro.
Figura 23A: Visão geral de exemplar da espécie Ageneiosus ucayalensis
Barra = 3 cm.
Figura 23B: Ageneiosus ucayalensis á necropsia mostrando a cavidade
abdominal e a disposição dos órgãos internos (OI) antes de serem removidos
para a placa de Petri (seta). Barra = 2 cm.
Figura 23C: Detalhe da conectividade da bolsa cística (Bc) com o fígado (F)
através de um pedúnculo composto de grandes vasos (Gv) que emergem do
fígado e pequenos vasos (Pv) que ramificam para dentro da bolsa cística e
irriga os cistos. Barra = 3 cm.
Figura 23D: Detalhe da bolsa cística (Bc), observado ao estereomicroscópio,
repleto de cistos de cestódeos, com um grande vaso (Gv) penetrando a bolsa e
ramificando em vasos pequenos (Pv) ao redor dos cistos. Barra = 30 µm.
Figura 23E: Detalhes da intensa vascularização da bolsa cística e disposição
dos vasos sanguíneos (Vs) envolvendo os cistos.Ainda se observa cistos
pequenos (Cp) intercalados por cistos maiores (Cg). Barra = 10 µm.
57
58
3.2.ASPECTOS MORFOLÓGICOS DAS LARVAS
As formas larvares de Cestódeos observadas pertencem à Famlia
Proteocephalidae. De acordo com as características morfológicas, e morfométricas,
agrupamos os parasitos em três morfotipos larvares, em diferentes fases conforme
as descrições seguintes:
3.2.1 Morfotipo I (Larvas Plerocercóides)
3.2.1.1 Análise Taxonômica
Este grupo de larvas possuem as seguintes características: corpo ovoide sem
segmentação, medindo 0,2801 mm ± 0,0736983 de comprimento por 0,1836 mm ±
0,0303975 de largura, com escólex bem desenvolvido mensurando 0,2070 mm ±
0,0533623
x 0,131µ± 0,0248527. Este pode ser encontrado invaginado, ou
evaginado. Essas larvas apresentam escólex provido de quatro ventosas
musculares com formato redondo, medindo 0,0474mm ± 0,0078255 x 0,0383mm ±
0,0064356, e um órgão apical desarmado dividido em lóbulos com 0,0515mm ±
0,0099263 de comprimento e 0,0498 mm ± 0,0114957 de largura. Na cavidade e nos
bordos das ventosas se observam estruturas filamentosas semelhantes a
microtríquias volumosas (Fig. 24). Além disso, não se observou a lacuna primária e o
sarcômero.
Figura 24: Desenho morfológico da fase larval Plerocercóide do Morfotipo I,
mostrando o escólex curto evaginado, corpo alongado e ausência de pescoço ou
colo. Barra = 100 µm.
59
3.2.1.2 Análise histológica das larvas do Morfotipo I
Nos exemplares com escólex evaginado é possível observar escólex pequeno
e redondo, com quatro ventosas musculares sendo duas láteroventrais e duas
laterodorsais e uma quinta ventosa apical circular característica do órgão apical. O
escólex se liga ao corpo do parasito por uma pequena constrição, o que caracteriza
colo ou pescoço ausente para esses parasitos. O corpo é delimitado por uma
membrana hialina espessa, que caracteriza o tegumento do parasita (Fig. 25A). Este
se mostrou altamente basofílico pela coloração com azul de toluidina.
Pela observação de cortes histológicos, o corpo desses parasitos encontra-se
preenchido por células vacuolizadas e o tegumento bastante espesso é interrompido
por uma pequena abertura, por onde o escólex se invagina. Estruturas celulares
alongadas e basofílicas são observadas circundando toda a superfície do escólex
(Fig. 25B).
As ventosas asseptadas apresentam padrões de bandas musculares e
algumas células basofílicas nos bordos. Nas cavidades e no ápice das ventosas são
observadas estruturas pontiagudas enfileiradas, característico de microtríquias. As
ventosas se conectam umas as outras e com o órgão apical por meio de canalículos
(Figs. 25C, D e E).
O órgão apical apresenta-se lobado e rodeado de células basofílicas. A
glândula apical esférica é perceptível no ápice do órgão apical emitindo raios que se
espalham por toda a extensão deste órgão conferindo-lhe a característica de lobos
(Fig. 25F).
60
Figura 25: Características morfológicas das Larvas do Morfotipo I visualizadas por
meio de técnicas histológicas.
Figura 25A: Visão geral da larva após evaginação do escólex. Barra = 100
µm.
Figura 25B: Larva com escólex invaginado, apresentando o corpo coberto
por células vacuolizadas e estruturas celulares ao redor do escólex e do
órgão apical. Barra = 100 µm.
Figura 25C: Visão das estruturas constituintes do escólex. Detalhe das quatro
ventosas musculares arredondadas e do órgão apical protuberante dividido
em lobos. Barra = 50 µm.
Figura 25D: Detalhe da conectividade das ventosas por meio de um
canalículo e dos espinhos pontiagudos nos bordos das ventosas.Células
basofílicas são observadas ao redor das ventosas e por toda a extensão do
escólex. Barra = 20 µm.
Figura 25E: Detalhe da ventosa muscular com a cavidade repleta de
espinhos ou volumosas microtríquias e células basofílicas ao redor.Barra = 50
µm.
Figura 25F: Glândula apical esférica no ápice do órgão apical com radiações
conferindo aspecto lobado deste órgão. Barra = 20 µm.
61
62
3.2.1.3 Microscopia Eletrônica de Varredura de larvas do Morfotipo I
A análise pela Microscopia Eletrônica de Varredura mostrou os cistos
envolvidos por um saco de tecido conjuntivo de forma irregular e aspecto bastante
enrugado (Fig. 26A). Quando livres do saco de tecido conjuntivo, os cistos
apresentaram superfície lisa com formato oval (Figs. 26B e 26C). Pequenas
estriações foram observadas neste cisto, indicativo de elasticidade, e que
provavelmente estão associado aos movimentos de contração e relaxamento deste
parasito. Fora da cavidade cística, o parasito apresentou tegumento desprovido de
espinhos, papilas, ganchos ou qualquer outra estrutura sensorial ou de fixação. Além
disso, o tegumento apresentou estruturas semelhantes a soluções de continuidade
indicando a alta atividade absortiva pelo mesmo (Fig. 26D).
Larvas desses parasitos com escólex evaginado foram observados fora da
cavidade cística (Figs. 26E e 27A). Estes apresentaram corpo alongado, escólex
curto e arredondado e ausência de pescoço ou colo, apresentando uma pequena
constrição que faz separação entre o corpo e o escólex.
O escólex é provido de quatro ventosas arredondadas que constituem os
órgãos de fixação, com espinhos pontiagudos na cavidade, nos bordos e em todo o
contorno das ventosas (Fig. 27B). Os espinhos apresentam-se enfileirados de forma
ordenada com convergência dos bordos para a região central da cavidade. Nestas
regiões, os espinhos são maiores e mais pontiagudos que os encontrados nos
contornos das ventosas, que vão se tornando mais truncadas quanto maior a
distância dos bordos das ventosas (Fig. 27C). Nos espaços entre as ventosas o
tegumento apresenta-se liso, semelhante ao observado no corpo destas formas
larvares.
A região apical do escólex apresenta levemente protuberante, característico
do órgão apical.Este se encontra coberto por pequenos filamentos em forma de
bastão, diferentes dos espinhos das ventosas (Fig. 27D).
O tegumento desses parasitos não apresenta especializações como espinhos
e papilas (Fig. 27E).
63
Figura 26: Caracteristicas morfológicas da bolsa cistica e das formas larvares de
cestóides, observados pela Microscopia Eletrônica de Varredura. A: Visão geral de
um grupo de cistos envoltos pelo saco de tecido conjuntivo. Barra=100 µm. B:
Detalhe parcial do cisto quando livre do saco de tecido conjuntivo apresentando a
musculatura lisa (seta). Barra: 50 µm. C: Cisto inteiramente livre do saco de tecido
conjuntivo apresentando formato oval e estriações externas que garantem a
elasticidade do parasito. Barra: 100 µm. D: Detalhe do tegumento rugoso do corpo
do parasito quando retirado do cisto (seta). Barra: 20 µm. E: Forma larval
apresentando escólex e o corpo do parasito e as aberturas das ventosas no escólex.
Barra: 200 µm.
64
Figura 27:
Figura 27: Características morfológicas das larvas plerocercóides de Cestódeos
encontrados parasitando Ageneiosus ucayalensis, observados pela Microscopia
Eletrônica de Varredura.
Figura 27A: Visão geral de uma larva plerocercóide, apresentando corpo
alongado e escólex arredondado e curto. Barra = 30 µm.
Figura 27B: Destaque para o escólex mostrando as ventosas com formato
arredondado, repleto de microtríquias volumosas na cavidade e na parte
externa. Barra = 10 µm.
Figura 27C: Detalhe das microtíquias volumosas enfileiradas, confluindo para
o centro da cavidade cística. Observa-se que são maiores e mais pontiagudas
nas cavidades e nos bordos do que na parte externa das ventosas. Barra = 3
µm.
Figura 27D: Região apical do escólex com estruturas filamentosas
semelhantes a volumosas microtríquias. Barra = 7 µm.
Figura 27E: Visualização da superfície do tegumento do corpo do parasito,
sem estruturas evidentes na superfície. Barra = 3 µm.
65
66
3.2.2 Morfotipo II (Larvas Merocercóides)
3.2.2.1 Análise taxonômica
Morfologicamente, essas larvas foram caracterizadas da seguinte forma:
corpo ovóide sem segmentação, medindo 1.033 mm ± 0.191 de comprimento por
0.468 mm
± 0.047 de largura, com escólex invaginado bem desenvolvido
mensurando 0,4070 mm ± 0,093 x 0,24 mm ± 0,045. Essas larvas são
monocefálicas, acetabuladas e apresentam escólex provido de quatro ventosas
musculares com formato redondo medindo 0,057 mm ± 0,0085 de comprimento por
0,049 mm ± 0,0063 de largura, e um órgão apical desarmado medindo 0,053 mm ±
0,0094 x 0,05 mm ± 0,013. Na cavidade e nos bordos das ventosas se observam
estruturas pontiagudas semelhantes a espinhos ou microtríquias longas. Além disso,
observa-se a ausência da lacuna primária e do sarcômero.
Figura 28: Desenho morfológico de larvas do Morfotipo II (Merocercóides),
evidenciando ocorpo ovóide, escólex invaginado, órgão apical grande e as ventosas
musculares arredondadas. A: Visão geral da larva e escólex invaginado contraído.
Barra = 150 µm; B: Visão geral da larva e escólex invaginado relaxado. Barra = 100
µm.
67
3.2.2.2 Análise histológica de larvas do Morfotipo II
A análise histológica de parasitos deste moroftipo mostrou-se semelhante ao
observado na análise do Morfotipo I. Os parasitos encontravam encistados, sendo
estas larvas e os cistos maiores que os do Morfotipo I.
O corpo desses parasitos é preenchido internamente por células em formato
de vacúolos acidofílicos ao azul de toluidina e externamente por um tegumento
espesso altamente basofílico ao mesmo corante (Fig. 29A). O tegumento é
interrompido por uma pequena abertura, por onde o escólex se invagina para dentro
do corpo do parasito (Fig. 29B).
O escólex apresenta tegumento composto de estruturas celulares basofílicas
alongadas e o conteúdo interno acidofílico quando corados com o azul de toluidina.
Além disso, estruturas especializadas do escólex, como as ventosas e o órgão
apical se fazem presentes.
As ventosas são asseptadas, uniloculares e apresentam padrões de bandas
musculares na região de circunferência das bordas, além de células basofílicas.
Estruturas pontiagudas, semelhantes a espinhos são observados nos bordos das
ventosas e na região de conexão destes com o órgão apical, porém não foram
observadas nas cavidades (Fig. 29C). As ventosas estabelecem conexões umas
com as outras e com o órgão apical por meio de estruturas semelhantes a
canalículos.
O órgão apical é bastante protuberante e apresenta-se com aspecto
granulado, também rodeado de células basofílicas. A glândula apical não foi
observada neste órgão (Fig. 29D).
68
Figura 29: Características morfológicas das Larvas do Morfotipo II visualizadas por
meio de técnicas histológicas.
Figura 29A: Visão geral do cisto e da larva apresentando escólex invaginado
e órgão apical protuberante. Barra = 200 µm.
Figura 29B: Detalhe da ligação do escólex invaginado com o tegumento
externo do corpo do parasito através de um orifício (seta). Notam-se
estruturas celulares basofílicas alongadas ao redor do escólex. Barra = 100
µm.
Figura 29C: Ventosas musculares com corpos celulares basofílicos e
microtríquias volumosas nas bordas. Barra = 50 µm.
Figura
29D:
Detalhe
do
órgão
apical
protuberante,
granuloso.Glândula apical não observada. Barra = 50 µm.
com
aspecto
69
70
3.2.2.3 Microscopia Eletrônica de Varredura de larvas do Morfotipo II
A análise desses parasitos pela Microscopia eletrônica de Varredura mostrou
que estes possuem corpo arredondado, com escólex invaginado. A superfície
corporal apresenta-se lisa e não foram observadas estruturas especializadas como
espinhos, microtríquias e papilas (Fig. 30A). Em uma das extremidades observou-se
o orifício por onde o escólex se invagina para dentro do corpo do parasito.Este
possui inúmeras pregas ao redor (Figs. 30B e 30C).
O escólex e suas estruturas constituintes foram observados quando estes
parasitos tiveram o corpo rompido. Este apresentou formato arredondado e curto, e
o pescoço ou colo ausente (Fig. 30D).
As ventosas apresentaram formato arredondado e séssil, constituindo assim
os órgãos de fixação desses parasitos. Espinhos pontiagudos são observados nas
cavidades e nos bordos. Estes estão presentes na região apical dessas ventosas
confluíndo para o centro da cavidade das ventosas. Na parte basal dessas ventosas
não se verifica padrões de espinhos (Figs. 30E e 30F).
O órgão apical não foi visível por esta técnica nestes parasitos.
71
Figura 30: Características morfológicas de larvas do Morfotipo II (fase larvar
Merocercóide) encontrados parasitando Ageneiosus ucayalensis, observados pela
Microscopia Eletrônica de Varredura.
Figura 30A: Visão geral do formato corpóreo da larva com escólex
invaginado. Barra = 100 µm.
Figura 30B: Presença do orifício de invaginação do escólex em uma das
extremidades do corpo do parasito. Barra = 30 µm.
Figura 30C: Detalhes do orifício, com abertura pequena circundada por
pregas. Barra = 10 µm.
Figura 30D: Disposição do escólex dentro do corpo deste parasito,
mostrando as ventosas arredondadas. Barra = 50 µm.
Figura 30E: Visão em maior aumento de uma das ventosas, apresentando
microtríquias volumosas do centro da cavidade para o ápice e ausência
destes na outra parte da ventosa. Barra = 5 µm.
Figura 30F: Detalhe das microtríquias volumosas nas regiões central e apical
da ventosa, sendo estes pontiagudos e dispostos em fileiras. Barra = 5 µm.
72
73
3.2.3 Morfotipo III (Larvas Merocercóides)
3.2.3.1 Análise Taxonômica
As formas larvares deste grupo apresentaram as seguintes características: corpo
alongado a subesférico, sem segmentação afinando posteriormente mensurando
0,904 mm ± 0,163 x 0,572mm ± 0,062. Escólex invaginado bem desenvolvido com
órgão apical e quatro acetábullos mensurando 0,440 mm ± 0,061 x 0,397mm ±
0,034; acetábulos em forma de ventosas. Órgão apical desarmado e pequeno
0,056mm ± 0,027 x 0,052mm ± 0,022; ventosas redondas a oblongas, semelhante a
botrídias, sésseis e uniloculadas medindo 0,086 mm ± 0,005 x 0,101mm ± 0.004. Na
cavidade e nos bordos das ventosas se observam estruturas pontiagudas
semelhantes a microtríquias volumosas. Não foram observadas lacuna primária e
sarcômero.
Figura 31: Desenho morfológico da larva do Morfotipo III (merocercóide),
evidenciando o corpo alongado, escólex invaginado, órgão apical pequeno e as
ventosas musculares arredondadas, com a base em formato cônico, confluindo para
o centro do órgão apical. A: Visão geral do formato das larvas. Barra = 150 µm; B:
Detalhe das ventosas e do órgão apical. Barra = 100 µm
74
3.3
ANÁLISE
MOLECULAR
COMPARATIVA
ENTRE
OS
MORFOTIPOS
LARVARES I E II
Para a análise filogenética foram obtidas 544 nucleotídeos da subunidade
menor do rDNA do morfotipo I e 635 nucleotídios do Morfotipo II. Estes nucleotídeos
foram blastados e alinhados com as sequências de Cestódeos Proteocephalídeos
neotropicais previamente depositadas no GenBank conforme mostra o Quadro 2.
Após a comparação das sequências pelo programa Blast, o Morfotipo I apresentou
99% de identidade com 68% de cobertura com as espécies Endorchis piraeeba e
Amphoteromorphus parkamoo enquanto o Morfotipo II apresentou a mesma taxa de
similaridade com 69% de cobertura com as espcies Proteocephalus brooksi e
Megathylacus brooksi.
A análise filogenética pelo método de Máxima Parcimônia e pela Inferência
Baesiana demonstrou que as fases larvares do Morfotipo I e do Morfotipo II
pertencem a gêneros distintos, pois o Morfotipo I formou um clado monofilético com
as espécies Amphoteromorphus parkamoo, Endorchis piraeeba e Choanoscolex
abscisus, enquanto que o morfotipo II se agrupou em outro clado com a espécie
Ageneiella brevifilis. Os valores de bootstrap e da probabilidade posterior foram
superiores a 30 e 0,60 respectivamente (Figs. 28 e 29).
Pela Inferência Baesiana, apenas as espécies de Proteocephalus ficaram
completamente separadas das demais espécies ao se agruparem em um clado com
valor de probabilidade posterior 0,9.
A distância ―P‖ inferida pelo PAUP, corroborou com os dados obtidos
anteriormente mostrando que o Morfotipo I e o Morofotipo II pertencem a gêneros
diferentes.No entanto por este parâmetro, o Morfotipo I mostrou semelhanças com a
espécie Endorchis piraeeba, e o morfotipo II mostrou-se diferente de todos os
demais gêneros analisados (Tabela 1).
75
Quadro 2: Espécies de parasitos neotropicais depositadas no genbank e utilizadas
neste trabalho para construção filogenética juntamente com os parasitos em estudo.
Hospedeiro
Origem
Genbank
Morfotipo I –larvas cestódeos
Ageneiosus
ucayalensis
Pará - brasil
________
Morfotipo II –larvas cestódeos
Ageneiosus
ucayalensis
Pará - brasil
________
Endorchis piraeeba
Brachyplatistoma
filamentosum
Amazonas Brasil
AY551107
Amphoteromorphus parkamoo
Paulícea luetkeni
Amazonas Brasil
AY551103
Nomimoscolex sudobim
Pseudoplatystoma
fasciatum
Amazonas Brasil
AY551117
Choanoscolex abscisus
Pseudoplatistoma
coruscans
Paraguai
Proteocephalus hemioliopteri
Phractocephalus
hemioliopterus
Amazonas –
Brasil
AY551129
Proteocephalus pirarara
Phractocephalus
hemioliopterus
Amazonas –
Brasil
AY551131
Goezeella siluri
Pinirampus
pirinampu
Amazonas –
Brasil
AY551110
Ageneiella brevifilis
Ageneiosus
breviffilis
Paraguai
Gibsoniela meursaulti
Ageneiosus
breviffilis
Paraguai
MesoCestódeos corti
_______
_______
Parasitos
AY551105
AY551102
AY551109
AF286984
76
Figura 32: Árvore filogenética obtida pelo método da Máxima Parcimônia, avaliando
parte da Subunidade menor do DNA ribossomal de Cestódeos. O círculo preto indica
a amostra dos parasitos do Morfotipo II e os círculos brancos representam as
amostras dos parasitos do Morfotipo I, ambos fases larvares de Cestódeos
encontrados parasitando Ageneiosus ucayalensis. As demais sequências foram
obtidas do GenBank e compõem diversos gêneros e espécies Neotropicais da
Família Proteocephalidae. São elas: Anphoteromorphus Parkamoo ( A parkam),
Endorchis Piraeeba (E Piraeba), Coanoscolex absiscis (Ch abscis), Nomimosclex
sudobim (N sudobim), Anphoteromorphus piraeeba (A piraeba), Gibsoniela
meursatus (G meursa), Ageneiela brevifilis (A brevif), Goezela siluri (G siluri),
Proteocephalus pirarara (P pirar), Proteocephalus hemioliopteri (P hemio).
Mesocestides corti (M corti) foi utilizado como grupo externo, para esta análise.
77
Figura 33: Árvore filogenética feita utilizando o método da Inferência Baesiana, com
a finalidade de comparar parte da Subunidade menor do DNA ribossomal de
Cestódeos. Os Morfotipos I (plerocercóides) e II (merocercóides) são fases larvares
de
Cestódeos
encontrados
parasitando
Ageneiosus ucayalensis.As
demais
sequências foram obtidas do GenBank e compõem diversos gêneros e espécies
Neotropicais da Família Proteocephalidae. São elas: Anphoteromorphus Parkamoo (
A parkam), Endorchis Piraeeba (E Piraeba), Coanoscolex absiscis (Ch abscis),
Nomimosclex sudobim (N sudobim), Anphoteromorphus piraeeba (A piraeba),
Gibsoniela meursatus (G meursa), Ageneiela brevifilis (A brevif), Goezela siluri (G
siluri), Proteocephalus pirarara (P pirar), Proteocephalus hemioliopteri (P hemio).
Mesocestides corti (M corti) foi utilizado como grupo externo, para esta análise.
78
Tabela 1: Distância ―p‖, obtida utilizando as sequências das fases larvares dos Morfotipos I e II e as sequências de Proteocephalídeos
Neotropicais disponíveis no GenBank.
Morfotipo Morfotipo E_piraeba A_parkam A_piraeba Ch_abscis G_siluri N_sudobim G_meursa A_brevif P_hemio M_corti
I
II
Morfotipo I
0.0000
Morfotipo II
0.0114
E_piraeba
0.0000
0.0115
A_parkam
0.0022
0.0137
0.0022
A_piraeba
0.0088
0.0162
0.0088
0.0110
Ch_abscis
0.0088
0.0206
0.0088
0.0110
0.0176
G_siluri
0.0198
0.0232
0.0197
0.0221
0.0198
0.0286
N_sudobim
0.0242
0.0319
0.0242
0.0264
0.0286
0.0286
0.0330
G_meursa
0.0220
0.0209
0.0220
0.0242
0.0264
0.0307
0.0286
0.0418
A_brevif
0.0330
0.0276
0.0330
0.0330
0.0330
0.0374
0.0396
0.0484
0.0417
P_hemio
0.0844
0.0843
0.0844
0.0869
0.0844
0.0911
0.0777
0.0914
0.0887
0.0933
P_pirar
0.0505
0.0531
0.0505
0.0529
0.0549
0.0593
0.0549
0.0615
0.0637
0.0703
0.0822
M_corti
0.1185
0.1216
0.1187
0.1211
0.1185
0.1184
0.1184
0.1319
0.1226
0.1225
0.1419
0.1319
79
3.4 ANÁLISE DOS ASPECTOS ANGIOGÊNICOS DO PARASITISMO
3.4.1 Descrição histológica
Os cortes histológicos do tecido contendo os cistos mostram um tecido
conjuntivo com poucas fibras e abundantes células ao redor dos cistos,
característico de tecido conjuntivo frouxo (Fig. 34A). Pequenos vasos sanguíneos
são visualizados compondo este tecido, circunvizinho aos cistos e, apresentam
circundados por uma substância de consistência viscosa, característico de
substância fundamental. Esta substância também é observada ocupando a maior
parte da cavidade cística, ao redor dos parasitos, o que faz com que esta representa
o maior volume observado neste tecido (Fig. 34B).
Neste tecido ainda é possível observar diferentes tipos celulares basofílicos
como células epiteliais alongadas ou fusiformes, com núcleo e nucléolo evidentes,
característico de fibroblastos e células granulocíticas agrupadas, formando
pequenos inchaços típicos de infiltrados inflamatórios.Hemácias nucleadas podem
ser vistas na luz dos vasos sanguíneos (Fig. 34B).
Os parasitos encontram-se envoltos por uma bolsa cística, cuja membrana é
fina, irregular e com ondulações típicas de tecido fibroelástico. Esta última
característica também foi observado no tegumento do parasito que apresentou
levemente metacromático ao azul de toluidina. Células basofílicas se apresentam
em grande abundância ao redor do escólex, principalmente na região muscular das
ventosas e nas redondezas do órgão apical (Fig. 34C).
Mais externamente ao agrupamento de cistos, os pequenos vasos
sanguíneos vão se convergindo em vasos de calibre cada vez maior, até a fusão
completa em um vaso de grande calibre (Fig. 34D) que normalmente é recrutado de
órgãos do hospedeiro altamente irrigados, como fígado e estômago. Estes vasos
estão sustentados por um tecido conjuntivo com muitas fibras e poucas células,
caracterizando possivelmente tecido conjuntivo denso.
Em alguns casos foram observados cistos destes psrasitos no fígado de
Ageneiosus ucayalensis e estes apresentaram uma pequena área de necrose,
caracterizado por poucas células (hepatócitos) ao redor e rico em fibras (Figs. 34E e
34F).
Secções semi-finas coradas com azul de toluidina mostraram vasos de médio
calibre no tecido conjuntivo ao redor dos cistos e a íntima relação entre pequenos
80
vasos e a membrana cística (Figs. 35A e 35B). Detalhes das estruturas internas do
parasito referentes ao escólex também foram evidenciadas tais como a musculatura
da parte apical arredondado das ventosas e as células basofílicas alongadas ao
redor do escólex (Fig. 35B). Vasos de grande calibre foram observados mais
externamente ao agrupamento cístico, com a luz repleta de hemácias nucleadas e
tecido fibroso na constituição da parede (Fig. 35C).
Colorações mais específicas, tais como a Hematoxilina-Eosina (HE) e os
Tricrômios de Mallory e Gomori, objetivando melhor caracterização das estruturas
celulares e o tipo de tecido conjuntivo envolvido neste processo foram utilizadas.
A coloração pela HE evidenciou células extremamente basofílicas na região
do escólex, principalmente ao redor das ventosas, do órgão apical e na bainha de
invaginação do escólex (Figs. 36A e 36B), além de células teciduais e hemácias no
interior dos vasos sanguíneos presentes no tecido conjuntivo que albergam os
cistos.Em contrapartida, o tecido conjuntivo ao redor dos cistos mostrou-se
altamente acidofílico (Fig. 36A).
A coloração pela orceína denunciou a presença de fibras elásticas como o
principal constituinte fibroso do escólex, membrana cística e tegumento do parasito
(Fig. 36C). Fibras colágenas também foram evidenciadas como constituintes desses
tecidos, pela coloração com tricrômio de Mallory (Fig. 36D).
Os vasos sanguíneos assim como o tecido conjuntivo ao redor dos cístos
também mostraram presença de fibras elásticas e colágenas em suas constituições,
porém com menor intensidade (Figs. 36C e 36E).
Fibras musculares na região apical do escólex e células basofílicas ao redor
deste órgão foram visualizadas pela coloração com os tricrômios de Mallory (Fig.
36F) e Gomori. Vestígios nucleares corados em preto se fizeram presentes no
escólex (Figs. 37A e 37 B).
A reação ao Ácido Periódico de Schiff mostrou-se intensamente positiva para
as regiões do escólex, tegumento, e conteúdo cístico ao redor dos parasitos
evidenciando a presença de compostos glicídicos nessas regiões. Estes compostos
também se fizeram presentes nas membranas dos vasos sanguíneos, porém com
menor intensidade (Figs. 37 C e 37D). Corte tratado apenas com o reagente de
Schiff foi tomado como controle negativo (Fig. 37E).
81
Figura 34: Análise de cortes histológicos de amostras embebidas em historesina® e
coradas pelo Azul de Toluidina.
Figura 34A: Visão geral do tecido contendo cistos de Cestódeos e vasos
sanguíneos, com destaque para os tecidos conjuntivos frouxo (tcf) e denso
(tcd) presentes nessa estrutura ricamente vascularizada. Pequenos vasos são
vistos por todo o tecido (setas brancas). Barra = 100 µm.
Figura 34B: Detalhe de vasos sanguíneos (V) na proximidade dos cistos;
cavidade cística (Cc) com ausência de parasito; parasito dentro do cisto com
células basofílicas na região do escólex (seta) circundando as ventosas e o
órgão apical; e infiltrado inflamatório (Inf) ao redor dos cistos. Barra = 20 µm.
Figura 34C: Visão geral da disposição de vasos e cistos, com destaque para
vaso de grande calibre (Vgc) de onde ramificam os vasos de médio calibre
(Vmc) que entrelaçam os cistos, caracterizando a rica vascularização nesse
tecido. Barra = 100 µm.
Figura 34D: Detalhe de vasos sanguíneos grandes (Vg) e pequenos, com
destaque para o pequeno infiltrado inflamatório (inf) e células epiteliais
fusiformes (setas), observados no epitélio e na luz dos vasos (seta). Barra =
10 µm.
Figura 34E: Fragmento de fígado do hospedeiro (F), com um parasito
encistado em seu interior (C). Barra = 100 µm.
Figura 34F: Detalhe do cisto no fígado do hospedeiro, mostrando a pequena
área fibrosada (f) com poucos hepatócitos ao redor. Barra = 20 µm.
82
83
.Figura 35: imagens de secções histológicas semi-finas do tecido contendo cistos de
parasitos Cestódeos e vasos sanguineos ao redor. A- cistos com parasito e vaso de
médio calibre no tecido conjuntivo. Barra: 50 µm. B – detalhe de estruturas do
escólex invaginado e pequenos vasos próximos à membrana cística. Barra: 50 µm.
C – vasos de grande calibre no tecido mais distante do cisto. Barra: 50 µm.
84
Figura 36: Análise histológica de Cistos e vasos em amostras embebidas em
parafina e submetidos à coloração pela Hematoxilina – Eosina, Tricrômio de Mallory
e Orceína.
Figura 36A: Visão geral de cistos, vasos e tecido adjacente corados com
hematoxilina eosina, destacando tecido basofílico na região do escólex,
acidofilia do tecido conjuntivo e vasos sanguíneos ao redor dos cistos. Barra =
200 µm.
Figura 36B: Detalhe da parte interna do corpo do parasito com destaque para
as células basofílicas ao redor das ventosas e do órgão apical; o tecido entre
as ventosas e na região central do órgão apical mostraram-se acidofílicos.
Barra = 20 µm.
Figura 36C: Visão geral da coloração do tecido com cistos de cestóides
pequenos (Cp) e grande (Cg) e vasos sanguíneos pelo tricrômio de Mallory.
Barra = 100 µm.
Figura 36D: Detalhe de cisto corado por tricrômio de Mallory, mostrando as
fibras musculares (Fm) do escólex, mais precisamente na região das
ventosas e as fibras colágenas (Fc) presentes no corpo do parasito e na
região tegumentar. Barra = 20 µm.
Figura 36E: Visão geral do tecido com cistos e vasos mostrando as fibras
elásticas em marrom, após coloração com Orceína. A região do escólex,
tegumento do parasito e tecido em volta destes mostraram maior afinidade
por este corante. Barra = 100 µm.
Figura 36F: Detalhe da região do escólex rico em fibras elásticas (marrom),
após coloração pela Orceína. Barra = 20 µm.
85
86
Figura 37: Análise histológica de Cistos e vasos embebidos em parafina e
submetidos à coloração pelo Tricrômio de Gomori e pelo Ácido Periódico de Schiff
(PAS).
Figura 37A: Visão geral da coloração do tecido conjuntivo e cistos de
cestóides pelo Tricrômio de Gomori. Barra = 100 µm.
Figura 37B: Detalhe de cisto corado por Tricrômio de Gomori, mostrando as
fibras musculares (seta) e as células basofílicas ao redor do escólex
(asterisco). Barra = 20 µm.
Figura 37C: Visão geral de cistos de Cestódeos e vasos sanguíneos do
tecido conjuntivo com reação positiva para a coloração do P.A.S. Barra = 100
µm.
Figura 37D: Detalhe da região do escólex, mais especificamente a região das
ventosas e do órgão apical, do tegumento do parasito e do conteúdo cístico
intensamente positivos pelo P.A.S. Barra = 20 µm.
Figura 37E: Visão geral do controle negativo da massa cística, tratada pela
reação do P.A.S. Note a ausência de reação desse tecido ao Ácido Periódico
de Schiff. Barra = 100 µm.
87
88
3.4.2 Reconstrução tri- dimensional
A reconstrução Tri-dimensional foi realizada para reconstruir a complexa
estrutura e a relação entre os vasos sanguíneos de diferentes calibres, desde a
conexão com órgãos do hospedeiro até as ramificações em vasos menores
circundando os cistos.(Figs. 38A, 38 B e 38C).
A disposição e a sobreposição dos cistos com parasitos maiores e menores
(Fig. 38D); a espessura dos vasos sanguíneos e o seus direcionamentos ao longo
da bolsa cística formando assim uma espécie de rede de conexão (Fig. 38E) e a
íntima relação entre os cistos contendo os parasitas e os vasos sanguíneos (Fig.
38F) foram observadas.
Através de rotações no eixo X em diferentes ângulos (Fig. 39), é possível
visualizar detalhes da relação cisto e vaso sanguíneo, desde as primeiras camadas
da área de reconstrução, até à última secção.
89
Figura 38: Reconstrução tri-dimensional dos cistos de cestóides e vasos
sanguíneos adjacentes a partir da somatória de 201 secções seriadas de material
embebido em Historesina®, obtido do parasitismo por cestóide em Ageneiosus
ucayalensis da Baia do Guaja, Belém-PA.
Figura 38A: Área do corte histológico, tomado para obtenção da
reconstrução tri-dimensional através do alinhamento em série de 201
cortes. Barra = 200 µm.
Figura 38B: Demonstração do contorno dos cistos, que foram seguidos
ordenada e sequencialmente até o término das 201 secções, para
obtenção da estrutura tri-dimensional dos mesmos. Barra = 200 µm.
Figura 38C: Demonstração do contorno dos vasos sanguíneos que foram
seguidos ordenada e sequencialmente até o término das 201 secções para
obtenção da estrutura tri-dimensional dos mesmos. Barra = 200 µm.
Figura 38D: Estrutura tri-dimensional dos cistos originado após o
alinhamento e contorno em software Reconstruct® Versão 1.1.0.0 19962009.
Figura 38E: Estrutura tri-dimensional dos cistos originado após o
alinhamento e contorno em software Reconstruct® Versão 1.1.0.0 19962009.
Figura 38F: Estrutura tridimensional dos cistos e vasos sanguíneos,
mostrando a íntima relação entre estes.
90
91
Figura 39: Diversos ângulos da reconstrução tri-dimensional obtidas a partir de
rotações no eixo X e Y, permitindo a visualização detalhada das estruturas
envolvidas no processo angiogênico induzida por larvas de Cestódeos em
Ageneiosus ucayalensis. A- Visão basal do aglomerado de cistos e vasos
sanguíneos entralaçados entre estes. B-Rotação no eixo do X em um ângulo de
180º levógero, mostrando a fase inicial da reconstrução, expessura e dimensão dos
cistos e vasos. C-Rotação no eixo do Y em um ângulo de 270º sentido descendente
com destaque para dimensão lateral e o entrelaçamento dos vasos com os cistos. D
– Rotação no eixo do Y, em um ângulo de 90º sentido descendente, mostrando a
massa de cistos bem mais ampla e pequenos vasos entre eles.
92
3.4.3 Microscopia Eletrônica de Varredura da massa cística
A análise do tecido contendo os parasitos e os vasos sanguíneos, que juntos
caracterizam um tecido ricamente vascularizado encontrado em Ageneiosus
ucayalensis. Pela técnica de Microscopia Eletrônica de Varredura observou-se um
saco cístico mostrou que o saco cístico, formado de tecido conjuntivo conectado
com um pedúnculo de vasos sanguíneos emergentes de órgãos do hospedeiro, a
exemplo do fígado (Fig. 40A).
Dentro deste saco é possível observar outros pequenos sacos albergando
grupos de cistos, assim como cistos de tamanhos diferentes, maiores e menores,
(Fig. 40B), que caracteriza as formas larvares distintas, descritas anteriormente, nas
diferentes fases de vida.
Ainda neste saco podemos observar uma intensa rede de vasos sanguíneos,
composto de vasos de pequeno e médio calibre que se ramificam em todas as
direções, envolvendo toda a massa cística (Figs. 40C e 40D).
Estes vasos se difundem pelo tecido conjuntivo, e apresentam uma íntima
relação com os cistos em particular. (Figs. 40E e 40F).
93
Figura 40: Características morfológicas do saco de tecido conjuntivo e vasos
sanguíneos que envolvem os cistos de Cestódeos, observados pela Microscopia
Eletrônica de Varredura.
Figura 40A: Visão geral do saco de tecido conjuntivo (stc) envolvendo os
cistos e o pedúnculo de vasos (p) ligado ao fígado (f). Barra = 1 mm.
Figura 40B: Detalhe da diferença de tamanho dos cistos, podendo ser
grandes (cg) e pequenos (cp) extremamente irrigados por uma rede de vasos
sanguíneos. Barra = 500 µm
Figura 40C e 40D: Detalhe da intensa rede de vasos sanguíneos de pequeno
e médio calibre multidirecionadas, de acordo com a disposição dos cistos.
Barra = 100 µm e 200 µm respectivamente.
Figura 40E: Em maior aumento observa-se diversas ramificações dos vasos,
contornando os cistos, direcionando desta forma o sentido do crescimento e a
formação de uma rede desordenada de vasos. Barra = 200 µm.
Figura 40F: Detalhe da conexão de um vaso de médio calibre com um grupo
de cistos dentro de um pequeno saco de tecido conjuntivo. Barra = 100 µm.
94
95
3.4.4 Microscopia Eletrônica de Transmissão da massa cística
A análise pela Microscopia Eletrônica de Transmissão evidenciou estruturas
teciduais e celulares na interface da relação entre os parasitos, vasos sanguíneos e
o tecido conjuntivo formado durante o processo angiogênico verificado no
parasitismo de larvas de Cestódeos em Ageneiosus ucayalensis.
No tecido conjuntivo circunvizinho aos cistos dos parasitos, agregados
filamentosos frouxamente arranjados foram observadas, característico de fibrilas ou
microfibrilas
de
colágeno,
constituinte
desse
tecido
como
demonstrado
anteriormente pelas colorações de orceína e tricrômio de Mallory (Figs. 41A e 41B).
Além disso, células com núcleo esférico e excêntrico, possuindo uma extensa área
de heterocromatina periférica que alterna com áreas de eucromatina, característico
de células plasmocíticas foram visualizadas nesse tecido (Fig. 41C). Também foi
possível observar estruturas apresentando aspecto proliferativo para formação de
membranas celulares (Fig. 41D ).
Para o estabelecimento do processo angiogênico, vasos sanguíneos migram
pelo tecido em direção aos cistos contendo os parasitos (Fig. 42A) e se estendem
por toda a circunferência dos cistos. Pela técnica de vermelho de rutênio evidenciouse as estruturas membranosas, fazendo separação nítida entre a membrana cística
e a membrana epitelial dos vasos (Fig. 42B). Regiões eletrondensas características
de íntima associação entre membranas sugerem conexão entre a membrana cística
e a membrana epitelial dos vasos sanguíneos (Fig. 42C).
Densa camada de aspecto fibrilar eletrondenso é observada no cisto, entre
os vasos de pequeno calibre e a superfície do parasito (Fig. 42D).
Uma camada eletronlúcida preenchida por estruturas vesiculares separa a
limitação externa do citoplasma distal do tegumento do parasito e a interface fibrosa
do cisto. Na região cortical próximo á superfície do tegumento do parasito, existem
numerosas vesículas de aspecto caveolar característico de vesículas pinocíticas
(Fig. 42F). Ainda nessa região de interface foi possível observar estruturas celulares
características de figuras mielínicas (Fig. 42G).
Os parasitos apresentam tegumento do tipo sincicial composto pelo
citoplasma distal anucleado, lâmina basal e células tegumentares perinucleares
situados abaixo da camada muscular do tegumento (Fig. 43A).
96
O citoplasma distal é composto quase na sua totalidade por vesículas
eletronlúcidas.
Corpos
eletrondensos
são
visualizados
em
algumas
áreas
intercalando com as vesículas. Externamente o citoplasma distal é limitado por
microtríquias filamentosas que se estendem por toda a superfície corpórea, sendo
estas melhor visualizadas em áreas de depressão tegumentar (Figs. 43B e 43C).
Estas microtíquias apresentam duas áreas distintas: uma capa eletrondensa, cujo
centro é também homogeneamente eletrondenso e o ápice composto por glicocálix,
e uma base cujo centro é eletronlúcido. Esta última é inserida na superfície do
citoplasma distal por uma membrana eletrondensa (fig. 43D).
A lâmina Basal se subdivide em duas camadas, sendo uma camada
eletrondensa que parece intimamente ligada ao citoplasma distal e uma camada
fibrosa que se conecta com o restante do aparato tegumentar. Bandas musculares
circulares e longitudinais são observadas abaixo da lâmina basal (Fig. 43E).
Processos internunciais, fazem conexão entre o citoplasma distal e o
proximal, conferindo característica de sincício observado neste tegumento. Estes
são visualizados em dois formatos diferentes: um com espessura fina, formando
pequenos canalículos entre o citoplasma distal e o proximal e outro largo, mais
elaborado, formado pela invaginação da membrana basal. Este cresce em direção
ao citoplasma proximal, atravessando a lâmina basal e a camada muscular (Fig. 43E
e 43 F).
97
Figura 41: Estruturas celulares e teciduais da bolsa cística ricamente vascularizada
e formada por larvas de Cestódeos em Ageneiosus ucayalensis observados através
da técnica de Microscopia Eletrônica de Transmissão.
Figura 41A: Arranjos fibrosos do tecido conjuntivo ao redor dos cistos. Barra
= 2µm.
Figura 41B: Detalhe da estrutura filamentosa das fibras organizadas em
feixes longitudinais (setas longas) e ou em secções aleatorias (setas curtas).
Barra =1 µm.
Figura 41C: Detalhe de célula do tecido conjuntivo com destaque para o
núcleo com extensa área de heterocromatina periférica e eucromatina central
eletronlúcida. Barra = 2µm.
98
99
Figura 42: Aspectos da inter-relação vasos sanguíneos, cisto e, tegumento de
larvas de Cestódeos ou Metacestódeos observados através da técnica de
Microscopia Eletrônica de Transmissão.
Figura 42A: Detalhe de vasos sanguíneos (v) que se estendem por toda a
superfície dos cistos (c). Barra = 1µm.
Figura 42B: Pontos de conexão entre os vasos, e a camada fibrilar, disposta
na superfície dos parasitos (setas). Barra = o,5µm
Figura 42C: Visão geral da associação dos vasos sanguíneos, membrana
cística fibrosa, tegumento do parasito e o espaço comunicante entre o
parasita e os constituintes do cisto. Detalhe da área cortical da larva repleto
de grânulos. Barra = 5µm.
Figura 42D: Detalhe da interface fibrilar disposta na superfície do parasito e
do espaço eletronlúcido com inúmeras vesículas próximo ao tegumento do
parasito (p). Barra = 2µm.
Figura 42E: Detalhe da região cortical do parasito com intensa atividade
pinocítica caracterizada pela formação de cavéolos (setas). Barra = 1µm.
Figura 42F: Figura mielínica observada dentro do espaço cístico, na interface
entre o parasito e o meio exterior a este. Barra = 0,5µm.
100
101
Figura 43: Ultraestrutura do tegumento de larvas de Cestódeos ou Metacestódeos
observados através da técnica de Microscopia Eletrônica de Transmissão.
Figura 43A: Detalhe do Citoplasma apical (cd) cheio de grânulos, do
tegumento de larvas de Cestódeos com células eletronlúcidas, lâmina basal
(Lb), bandas musculares (m) e células tegumentares do citoplasma proximal
(cp). Barra = 1µm.
Figura 43B: Detalhe de microtríquias por toda a extensão da superfície do
tegumento da larva (setas). Barra = 2µm.
Figura 43C: Detalhe das microtríquias filamentosas (mf) na região de
depressão tegumentar. Abaixo das microtríquias observa-se vesículas
absortivas. Barra = 0,5µm.
Figura 43D: Detalhe de uma microtríquia filamentosa eletrondensa, composta
pela capa (ca) eletrondensa e a base (b) eletronlúcida. Barra = 0,5µm.
Figura 43E: Detalhe da invaginação do citoplasma distal (setas) em direção
ao citoplasma proximal
e das camadas de músculos circulares (mc) e
longitudinais (ml), situados abaixo da lâmina basal. Barra = 2µm.
Figura 43F: Detalhe dos processos internunciais largo (seta branca) e finos
(setas pretas) fazendo conexão entre o citoplasma distal e o proximal. Barra =
1µm.
102
103
4 DISCUSSÂO
4.1 ANÁLISE MORFOLÓGICA
As formas larvares deste estudo pertencem à Ordem Proteocephalidea,
Família Proteocephalidae, pois se encontravam encapsuladas, possuindo um
escólex com quatro ventosas musculares, acetabulares e uniloculadas, órgão apical,
e são parasitos de cavidade celomática de Ageneiosus ucayalensis. De acordo com
a chave taxonômica para fases larvares de Cestódeos proposta por Rego et al.
(1999) estas são as principais características de diagnose para formas larvares da
Ordem Proteocephalidea, Família Proteocephalidae.
Todas as formas larvares deste estudo apresentaram escólex invaginado e
bem desenvolvido pelo que foram classificados inicialmente como sendo larvas
Merocercóides da Família Proteocephalidae, pois segundo Chervy (2002), os
parasitos pertencentes a esta Família possuem duas fases larvares: a fase
Plerocercóide cuja característica principal é a presença do escólex evaginado, e a
fase Merocercóide que apresenta escólex invaginado.
Pelas características morfológicas, morfométricas e biológicas, encontramos
três morfotipos de larvas de Cestódeos: o Morfotipo I com tamanho pequeno e
excólex com órgão apical grande; o Morfotipo II com tamanho grande e escólex
possuindo órgão apical grande; e o Morfotipo III com tamanho relativamente grande
e apresentando o órgão apical muito pequeno.
As larvas do Morfotipo I e do Morfotipo II são idênticas morfologicamente,
diferindo apenas em morofmetria e prevalência, onde o Morfotipo I (menor) é mais
abundante que o Morfotipo II (mais volumoso), aparentando assim, tratar-se de um
mesmo tipo, podendo a diferença de tamanho estar relacionada com o tempo de
infecção do hospedeiro Ageneiosus ucayalensis. Contudo, de acordo com Schoolz
(1991), os platelmintos não são capazes de desenvolver dentro de seus hospedeiros
de transporte ou paratênicos. No entanto, podem permanecer vivos durante muito
tempo. Diante disso, investigamos a hipótese de que as formas larvares menores
(Morfotipo I) poderiam ser do tipo Plerocercóide, enquanto que as maiores
(Morfotipo II) seriam do tipo Merocercóide, uma vez que é comum a invaginação do
escólex pelos Plerocercóides Proteocephalídeos, visando a proteção do parasito
frente às condições do meio em que está exposto. Esta hipótese foi fundamentada
104
ao observamos a evaginação de excólex por alguns exemplares do Morfotipo I
quando estes foram expostos a uma solução tampão fosfato salino (PBS); reação
que não foi observada no Morfotipo II. Assim, a invaginação do escólex por esta
forma larvar pode ser explicado pelo fato de Ageneiosus ucayalensis entrar no ciclo
de vida como hospedeiro paratênico, e, portanto não possui as características físicoquímicas adequadas para o crescimento e desenvolvimento desta larva.
Anikieva (1983) relatou a invaginação do escólex de larvas de Proteocephalus
logo após a ingestão por seus hospedeiros definitivos, como forma de proteção
contra a acidez do estômago e, quando em meios alcalinos, os parasitos
evaginariam o escólex. Em contrapartida, Freeman (1964) observou ocasionalmente
a invaginação do escólex de metacestóides em meio salino, mas não observou in
situ. Agusti et al., (2005) também observou evaginação do escólex de
Phyllobothrium delphini e Monorygma grimaldii, ambas as espécies pertencentes à
Ordem Tetraphyllidea, após exposição dos parasitos em meio salino.
No entanto pelas características moleculares, concluímos que estas formas
larvares se tratam de três morfotipos larvares em estágio diferente, estando o
Morfotipo I na fase Plerocercóide e os Morfotipos II e III na fase Merocercóide. Estes
dados levantam uma questão importante do ponto de vista ecológico onde
Ageneiosus ucayalensis seria tanto hospedeiro intermediário quanto paratênico para
diferentes fases larvares da Família Proteocephalidae.
Os três morfotipos larvares não possuem metascólex, aspecto característico
de espécies dos gêneros Zygobothrium, Brayela, Chambriella, Gibsoniela,
Harriscolex,
Houssayela,
Travassiella,
Crepidobothrium,
Proteocephalus,
Cangatiella, Nomimoscolex, Nupelia, Monticellia e Ophiotaenia de acordo com Rego
et al. (1999). Scholz et al. (1998) e Rego (1999) consideram que a presença ou não
de metascólex, as característricas de escólex referentes a glândulas frontais, órgão
apical, apêndices das ventosas, presença de espínhos ou microtríquias, associadas
à descrição da disposição das gônadas e vitelarias em relação à musculatura interna
longitudinal, forneceriam informações mais precisas na separação de subfamílias e
gêneros dentro do grupo Proteocephalidea. No entanto, de acordo com Rego (1999)
esses caracteres têm sido negligenciados pelos autores na classificação de
Proteocephalídeos da América do Sul que consideram apenas as características dos
órgãos reprodutivos. Além disso, Rego (1999) afirma que as características de
105
escólex de formas larvares de Proteocephalídeos não sofrem muitas modificações
quando este se desenvolve para a forma adulta.
Deste modo, foi observado em A. ucayalensis que ambos os Morfotipos I, II e
III apresentam escólex pequeno, arredondado, com quatro ventosas musculares
sem apêndices e repletas de microtríquias volumosas semelhantes a pequenos
espinhos nos bordos e nas cavidades, glândula apical na região apical do escólex e
um órgão apical característico e desarmado. Este último é bem protuberante nos
Morfotipo I e II, enquanto que no Morfotipo III se apresenta de forma sutil. Segundo
Rego, 1999, nas espécies de Cestódeos Neotropicais descritas, o órgão apical é
frequentemente
encontrada
nas
espécies
dos
gêneros
Proteocephalus
e
Ophiotaenia, podendo ser bastante notável ou em alguns casos de difícil
observação. Por outro lado, Yamaguti (1959) utiliza como características de
diagnose do gênero Ophiotaenia a presença do órgão apical vestigial.
A glândula apical, também conhecida como glândula frontal é mais frequente
em espécies dos gêneros Proteocephalus, Ophiotaenia e Nomimoscólex. Já os
espinhos podem ser encontrados recobrindo toda a superfície das ventosas em
algumas espécies dos gêneros Monticellia, como M. Spinulífera e M. Lenha, e
Nomimoscolex, a exemplo de N. piraeeba, N. Sudobim, N. Lenha e N. Semenasae
(Rego, 1999). Por outro lado, Yamagutti (1959) considera que o gênero Monticellia
não possui órgão apical, podendo este estar ou não presente no gênero
Nomimoscolex. Estes dados nos dão indício de que as larvas do Morfotipo I e II
possuem algumas características que se assemelham a espécies do gênero
Proteocephalus e outras que tendem para espécies do gênero Nomimoscolex;
enquanto que as características do Morfotipo III direciona este parasito para o
gênero Nomimoscólex.
Rego (1999) acrescenta que as espécies de Proteocephalídeos Neotropicais
não
necessitam
de
estudos
minuciosos
dos
sistemas
reprodutivos
para
diferenciações morfológicas espécies-específicas, como acontece nas espécies de
Proteocephalídeos da região paleártica, podendo a identificação ser feita pelas
características morfológicas de escólex e metascólex. Em nossa opinião, para a
identificação taxonômica clássica de espécies de proteocephalídeos, ainda se faz
necessário o estudo dos órgãos reprodutores através da análise de gônadas e
vitelárias em cortes histológicos de proglotes maturas, o que não é possível para
este estudo, pois os parasitos em questão tratam-se de fases larvares da Família
106
Proteocephalidae. Para esclarecer este impasse buscou-se técnicas de Biologia
molecular na tentativa de esclarecer os gêneros e mesmo espécies aos quais
pertencem os parasitos dos Morfotipos I e II. O emprego desta técnica para o
Morfotipo III não foi possível, por falta de exemplares.
4.2 BIOLOGIA MOLECULAR
Os resultados obtidos pelas técnicas de biologia molecular inferiram uma
separação em nível de gêneros das fases larvares dos Morfotipos I e II, uma vez que
estes se separaram em clados diferentes, agrupando com diferentes espécies. Pelos
dados da distância ―P‖, o Morfotipo I mostrou similaridade com a espécie Endorchis
piraeeba, podendo se tratar da mesma espécie. O Morfotipo II mostrou distância
significativa com todos os gêneros analisados, inclusive o Morfotipo I. No entanto,
estes dados não são suficientes para afirmar com precisão a quais gêneros esses
parasitos pertencem. Isto provavelmente se deve ao fato das sequências obtidas
oferecerem poucas informações nucleotídicas para garantir a separação desses
parasitos, uma vez que apenas cerca de 400 pares de bases da região 18S foram
utilizados para análise.
Outro aspecto a ser considerado é o fato de que a região gênica 18S é
bastante conservada, sendo necessário um estudo mais criterioso utilizando regiões
com maiores especificidades para a separação de espécies, a exemplo da região
ITS. Este fato foi comprovado por Hypsa et al. (2004), que ao analisarem espécies
de proteocephalídeos das regiões neotropical e paleártico, não obtiveram resultados
satisfatórios apenas com a região 18S, sendo isto possível após a combinação
dessa região com as regiões ITS, 16S e 28S para se obter um suporte significativo
dos ramos filogenéticos. Além disso, estes autores observaram que destas
sequências de proteocephaíideos analisadas, a região 18S foi a que apresentou
maior nível de conservação, enquanto a região ITS foi o que apresentou maiores
informatições. Estes autores realçaram ainda que a região ITS pode ser útil
futuramente para estudos mais complexos.
Assim, acreditamos que a princípio o gene 18S é útil para separação e/ou
distinção entre os morfotipos larvares I e II encontrados em A. ucayalensis; no
entanto, é necessário a utilização de outros genes para se investigar a identificação
de gêneros e, por que não, espécies destes dois Morfotipos.
107
4.3 CARACTERÍSTICAS MORFOLÓGICAS DA RELAÇÃO ENTRE AS LARVAS
METACESTÓIDES E A.ucayalensis
4.3.1 Aspectos histológicos
O intenso parasitismo culmina com a formação de uma massa individualizada
composta por centenas de cistos e complexa irrigação sanguínea a partir de um
tronco venoso que se origina de órgãos parenquimatosos ou não. As numerosas
larvas agregam-se por meio de tecido conjuntivo tipo frouxo, com fibras escassas,
frouxamente arranjadas e abundantes células de diferentes tipos. Segundo Ross et
al. ( 2008) estes são os principais ítens que caracterizam um tecido conjuntivo
frouxo.
A matriz extracelular também está composta por material amorfo, viscoso e
abundante, ocupando maior volume que as fibras. A complexa rede de vasos
sanguíneos ao redor dos cistos provavelmente atua no transporte de oxigênio e
nutrientes para o tecido conjuntivo frouxo e os parasitos, e ainda na condução de
produtos de excreção/secreção dos parasitos. De acordo com Ross et al. (2008) a
substância fundamental dos tecidos conjuntivos frouxos circunda os pequenos vasos
sanguíneos e tem papel importante na difusão de oxigênio e nutrientes a partir dos
pequenos vasos assim como na difusão de dióxido de carbono e resíduos
metabólicos até os vasos. Assim, a partir deste conceito é possível inferir sobre a
necessidade dos parasitos de estabelecer a relação com seu hospedeiro, visando à
manutenção de sua permanência e sobrevivência em A. ucayalensis.
A presença de fibroblastos na massa cística sugere atividade de síntese de
colágeno, fibras elásticas e reticulares e os complexos carboidratos da substância
fundamental. Além disso, a presença de outros tipos celulares pode caracterizar
discretas manifestações de reações inflamatórias e imunes, por migração de células
transitórias dos vasos sanguíneos em resposta a estímulos específicos. Estes
aspectos são similares aos padrões conceituais de tecido conjuntivo frouxo descritos
em vertebrados por Ross et al. (2008).
Os vasos sanguíneos de diferentes calibres que irrigam este tecido e os cistos
dos parasitos e convergem posteriormente ou ramificam-se a partir de um único
vaso de calibre maior sugerem um processo angiogênico induzido pelo parasitismo.
O tecido conjuntivo que envolve os vasos maiores fornece a força significativa para
resistir ao estiramento e à distensão excessiva do tecido conjuntivo frouxo,
108
provocada pela movimentação das larvas dentro dos cistos o que justifica a
presença de abundantes fibras e escassez de células caracterizando o tecido
conjuntivo denso.
As fibras elásticas e colágenas evidenciados neste tecido são condizentes
com os tipos de fibras que compõem os tecidos conjuntivos frouxo e denso descrito
por Ross et al. ( 2008), o que reforça mais uma vez a correta caracterização desses
tecidos. As fibras elásticas e colágenas permitem que o tecido responda
dinamicamente aos movimentos dos parasitos e mesmo a possíveis agregações
posteriores de novas larvas infectivas.
Os compostos glicídicos evidenciados pela reação do P.A.S. nas regiões do
escólex, tegumento, e conteúdo cístico ao redor dos parasitos sugerem que estes
parasitos possuem uma alta taxa metabólica, podendo estes compostos atuarem
ainda como uma barreira de reconhecimento deste parasito pelo sistema imune do
hospedeiro. Coggins et al. (1979) relatou a presença de um componente no órgão
apical de larvas Plerocercóides de Proteocephalus ambloplitis positivo para P.A.S.,
ao contrário do que tinha sido relatado anteriormente por Wood (1965) em órgão
apical de Ophiotaenia filaroide, quando submetido à coloração pelo P.A.S. Ainda
segundo Coggins et al. (1979) estes achados sugerem diferenças químicas e
morfológicas entre órgãos apicais de Cestódeos Proteocephalídeos. Contudo,
acreditamos que nas larvas parasitas de A. ucayalensis, podem atuar como reserva
energética, além de fonte para manutenção de glicocálice e mesmo para produção
de componentes de matriz extracelular e outros produtos de secreção.
4.3.2 Aspectos ultraestruturais
Aspectos ultraestruturais mostram íntima associação entre a membrana
cística e a membrana das células endoteliais dos vasos sanguíneos dando origem
em alguns pontos de junções, o que sugere que estas membranas estão em estreito
contato, permitindo e facilitando o transporte de líquidos e eletrólitos.
Ross et al. (2008), destacou a importância das junções, fundamentais nos
tecidos em que a atividade de células adjacentes devem ser coordenadas pela troca
de íons, moléculas reguladoras e pequenos metabólitos.
Os compostos glicídicos do conteúdo cístico, como mostrado na histoquímica
do P.A.S., podem ser visualizados como uma extensa e espessa área de
109
microfibrilas eletrondensas que recobre o corpo do parasito. Esta espessa estrutura
que cria uma interface entre a larva e o tecido conjuntivo do hospedeiro encerra a
larva em um cisto individual e pode ser produzida por ambos (cisto e hospedeiro),
atuando ainda como um filtro protetor do parasito.
A presença de vesículas no espaço entre o parasito e a ―capa‖ cística fibrosa
indica grande atividade metabólica na superfície do parasito, através do transporte
vesicular. Este aspecto também é bastante evidente abaixo da membrana do
tegumento, no córtex do citoplasma sincicial apical. Mais uma vez representando
alta atividade metabólica caracterizada por vesículas pinocíticas caveolares.
Atividades pinocíticas no tegumento de larvas de cestódeos foram relatadas por
Hopkins et al. (1978), Threadgold et al. (1981), Threadgold et al. (1983) e
Schramlová et al. (1988). Segundo Schramlová et al. (1988), os cestódeos não
possuem sistema digestivo e todos os transportes de substâncias realizados através
do tegumento são mediados não apenas pela difusão mas também por microporos
presentes na membrana, pelos quais ocorre a endocitose. Estes autores ainda
ressaltam que a atividade de pinocitose ocorre apenas no tegumento corpóreo e não
no tegumento do escólex de Cisticercus bovinos, sendo notório aproximadamente
quatro semanas após a infecção.
A finalidade pela qual estes parasitos internalizam vesículas por meio de
pinocitose não é bem entendida. Threadgold et al. (1981), acreditam que a
pinocitose no tegumento tem caráter nutricional, porém não descartam a
possibilidade de outra função das macromoléculas internalizadas. Por outro lado,
Beffus (1977), Threadgold et al. (1983) e Schramlová et al. (1988), sugerem que a
pinocitose dessas moléculas possui papel importante na degradação de enzimas e
anticorpos que aderem na superfície dos parasitos, protegendo os parasitos contra a
destruição pelas células imunes do hospedeiro. A degradação de tais substâncias
ocorreria nas células tegumentares do citoplasma proximal, pela localização de
heterolisossomos nestas células. Porém em alguns casos estas macromoléculas
não seriam degradadas, mas sim modificadas e transportadas novamente para o
citoplasma distal onde contribuiriam para mascarar o parasito e este sobreviver
contra o sistema imune do hospedeiro (Schramlová et al., 1988).
Acredita-se que ambas as funções (nutritiva e protetora) podem ocorrer
através da internalização vesicular nesses organismos, tal como ocorre no
tegumento de Schistosoma. Segundo Hellemond et al. (2006), a superfície do
110
tegumento de schistosoma é a única estrutura de membrana com função dupla de
modulação da resposta imune do hospedeiro e sobrevivência parasitária.
Este é o primeiro relato de pinocitose no tegumento de larvas de Cestódeos
proteocephalídeos.
Os parasitos de A.ucayalensis apresentam tegumento sincicial dividido em
três partes: citoplasma distal, lâmina basal e citoplasma proximal. Este aparato
tegumentar é comum em Cestódeos como observado por Jones et al. (1998); Palm
et al. (1998); Levron et al. (2008) e Dey et al. (2009) e em Trematódeos de acordo
com Dorsey et al. (2002) & Cavalcanti et al. (2009). A presença de vesículas
eletronlúcidas preenchendo toda a estensão do citoplasma distal também foi
verificada nos trabalhos de Palm et al. (1998), e Levron et al. (2008); porém elas se
mostram muito mais numerosas no presente estudo. Diferente do pressuposto
anteriormente, Jones et al. (1998) sugeriram que estas vesículas são produzidas
pelo tegumento sincicial dos Cestódeos e transportadas para o citoplasma distal
onde realizam a manutenção da superfície do parasito e sintetizam novas estruturas
apicais. Ainda segundo estes autores, a natureza do conteúdo dessas vesículas é
pouco entendida, mas poderiam estar envolvidas na formação de microtríquias.
Microtríquias filamentosas preenchem a superfície externa do citoplasma
distal do tegumento desses parasitos. Diferentes tipos de microtríquias podem ser
vistas por toda a extensão do corpo de Cestódeos. Microtríquias filamentosas são
comumente encontradas na região do escólex, mais precisamente no órgão apical e
rostelo, e nas proglotes imaturas do corpo de adultos de Proteocephalus tidswelli de
acordo com Thompson et al. (1980). Estas estruturas atuam ampliando a área
superficial de digestão e absorção de nutrientes através da agitação do microhabitat
adjacente onde estão depositados os nutrientes o que resulta em um movimento
passivo dos filamentos e ocorre em simultâneo com a contração e o relaxamento do
parasito, de acordo com Thompson et al. (1980) e Jones et al. (1998). Além dessas
funções, elas também atuam na ancoragem e na função mecanoreceptora de
receptores sensoriais segundo Palm et al. (1998).
Nas larvas metacestóides parasitos de A.ucayalensis, a lâmina basal
encontra-se dividida em duas regiões distintas. Uma região eletrondensa
intimamente ligada ao citoplasma distal e uma região fibrosa que se estende para o
citoplasma proximal. Segundo Jones et al. (1998), a lâmina basal é geralmente
observada abaixo do citoplasma apical e nunca abaixo das células tegumentares do
111
citoplasma proximal, pois a matriz basal tem a função de manter a polaridade e a
integridade estrutural do citoplasma distal.
Os dois tipos de processos internunciais observados neste estudo,
caracterizados como estreitos e expessos estão intimamente associados à troca de
metabólitos entre o parasito e o meio externo onde os processos estreitos carreiam
informações do citoplasma proximal para o distal e os processos espessos atuariam
no sentido inverso.
Os processos internunciais estreitos têm sido relatados como uma extensão
citoplasmática progressiva de células tegumentares que culminam no citoplasma
distal por Chappell (1980), Coil (1991), Smyth (1994), Roberts et al. (2000) e Dey et
al. (2009); carreando informações do citoplasma proximal para o distal de acordo
com Dey et al. (2009). Os processos espessos foram descritos pela primeira vez por
Dey et al. (2009) que mostrou sua função contrária aos processos finos.
Ainda segundo Dey et al. (2009) os processos espessos estariam atuando no
transporte vesicular por estes serem mais elaborados e com diâmetro superior a de
uma vesícula; ao contrário dos processos estreitos que apresentam diâmetros curtos
e não são, portanto suficientemente largos para a passagem de uma vesícula.
Maiores estudos de ultraestrutura são necessários para melhor entendimento
da relação parasito hospedeiro entre esses metacestódeos e A.ucayalensis.
4.3.3 Aspectos Angiogênicos
A formação de granuloma cística, neoformação de tecido conjuntivo com
estruturas celulares típicas, e vasos sanguíneos ao redor dos parasitos caracteriza o
processo angiogênico à presença de larvas de Cestódeos no peritônio visceral de A.
ucayalensis. Esses eventos podem ocorrer de diferentes modos e nas infecções por
parasitos das diferentes classes como Cestódeos, Trematódeos e Nematódeos.
Tecido granuloso e remodelação tecidual, com proliferação vascular e
hiperplasia endotelial, associado ao aumento de colágenos tipos I e III e expressão
de Fator de Crescimento Fibroblástico básico foram observados por Alvarez et al.
(2002) e Alvarez et al. (2002) em Cisticercose suína e Neurocisticercose humana
induzidas por Metacestóides de Taenia solium. A presença de citocinas como
Interleucina 10 (IL-10) e Fator de Crescimento Tumoral beta (TGF-β) e ausência das
112
Interleucinas 4 e 13 (IL-4 e IL13), presentes na resposta imune do tipo Th2, em
Neurocisticercose humana caracterizaram atividade imunomodulatória nesse tecido.
Estes eventos também são observados e bem caracterizados em infecções
por S. mansoni (Trematódeo conhecido por desenvolver angiogênese e fibrose
hepática em humanos), como mostra os trabalhos desenvolvidos por Lenzi et al.
(2002), Sobky et al. (2002), Baptista et al. (2005), Andrade (2009), Andrade et al.
(2010), Lemos et al. (2010), Shariati et al. (2011), e Raso et al. (2012). Além disso,
estudos de ultraestrutura de células teciduais vêm sendo correlacionados quanto a
suas funções no estabelecimento desses processos. Weinberg et al. (1992)
observaram intensa atividade angiogênica associada a atividade de plasmócitos no
peritônio de ratos infectados por Schistosoma mansoni e segundo estes autores, os
plasmócitos estão associados à produção de anticorpos diretamente no peritônio
para controle específico deste local e com formação de estruturas particulares para
produção em larga escala de outros plasmócitos maturos. Lenzi et al. (2002) atribui
a síntese de fibras a fibroblastos presentes em nódulos fibrovasculares causados
por S.mansoni.
De semelhante modo, processos fibróticos e formação de angiogênese e
linfangiogênese são relatados em infecções por Nematódeos como mostra os
trabalhos de Attout et al. (2009), Shariati et al. (2010), Gaban et al. (2010) e Lemos
et al. (2010). O organismo que melhor desencadeia este quadro é a Capillaria
hepática que semelhante a S. mansoni ocasiona quandros fibróticos e angiogênese
no fígado.
O
processo
angiogênico
verificado
em
A.ucayalensis
é
induzido
provavelmente, pela secreção/excreção de substâncias angiogênicas pelas larvas
Metacestódeos visando garantir a nutrição, desenvolvimento, e sobrevivência nesse
hospedeiro. De acordo com Dennis et al. (2011), durante o desenvolvimento tecidual
dos parasitos helmintos, ocorrem interações diretas e indiretas entre estes e o
sistema vascular do hospedeiro, e os parasitos podem ter a capacidade de iniciar
eventos pro-angiogênicos.
Tais substâncias angiogênicas podem ser similares a Fatores de Crescimento
Vascular e Fibroblasto com ação direta na formação de vasos sanguíneos como
verificado em Ceanorabditis elegans (Tarsiano et al., 2006) e Equinococcus
multilocularis (Stadelman et al., 2010) ou podem ter ação secundária na indução da
angiogênese de acordo com Higazi et al. (2003), Kanse et al. (2005), Maity et al.,
113
(2007), Kreider et al. (2007), Gottstein et al., (2008), Shariati et al., (2010) entre
outros. Contudo estudos de expressão gênica e de Proteômica desses parasitos se
fazem necessários para elucidar as moléculas e os genes envolvidos na formação
de vasos sanguíneos e proliferação celular em A. ucayalensis.
4.4 ASPECTOS BIOLÓGICOS
4.4.1 Prospecção Parcial do Ciclo Biológico
Com base em todas as técnicas empregadas bem como os resultados obtidos
das mesmas e, na observação da interação entre esses parasitos com o hospedeiro
Ageneiosus ucayalensis, postulamos três hipóteses referentes ao papel desse
hospedeiro no ciclo de vida e na biologia desses parasitos conforme o diagrama a
seguir.
Figura 44: Diagrama ilustrativa de possíveis formas de atuação de Ageneiosus
ucayalensis, no ciclo biológico de Metacestódeos Proteocephalídeos de Belém –PA.
114
Na primeira hipótese, Ageneiosus ucayalensis atua como hospedeiro
paratênico para as larvas Plerocercóides (Morfotipo I). Neste caso o hospedeiro se
infecta pela ingestão da forma larval Plerocercóide. Estas larvas são conduzidas
para o intestino onde atravessam a parede e se encistam na cavidade celomática de
A. ucayalensis, estabelecendo deste modo a infecção. Estas larvas permanecem em
quiescência até que este hospedeiro sofre predação por peixes maiores onde estas
larvas conseguem se desenvolver para formas adultas.
Na segunda hipótese, Ageneiosus ucayalensis possivelmente está atuando
como hospedeiro intermediário das formas larvares dos Morfotipos II e III. O
hospedeiro se infecta ingerindo as formas larvares Plerocercóides que sofrem
mudas no trajeto pelo trato gastrointestinal transformando em larvas Merocercóides
que atravessam a parede intestinal e se instalam na cavidade celomática de A.
ucayalensis. Neste local estas larvas então aguardam o momento em que o
hospedeiro intermediário será ingerido pelo hospedeiro definitivo onde poderão se
desenvolver em formas adultas.
Na terceira hipótese, Ageneiosus ucayalensis seria um hospedeiro paratênico
para as formas larvares dos Morfotipos II e III, onde este se infecta pela ingestão das
formas larvares Merocercóides. No intestino estas larvas atravessam a parede
intestinal e se instalam na cavidade celomática de A. ucayalensis, onde
permanecerão
aguardando
a
ingestão
pelo
hospedeiro
adequado
para,
posteriormente, se desenvolverem em formas adultas.
Das três hipóteses apenas a primeira parece estar completamente elucidada,
uma vez que as larvas não crescem neste local, permanecendo na primeira fase
larval do ciclo biológico desses parasitos e a invaginação do escólex está associado
provavelmente ao desconforto da larva frente às condições fisicoquímicas a ela
oferecidas, e que são diferentes da sua exigência para o desenvolvimento em fase
adulta. A segunda e a terceira hipótese só seriam completamente elucidadas se
soubéssemos a fase larvar da forma ingerida, o que seria possível através de
infecção experimental. Quaisquer das três hipóteses só serão confirmadas a partir
da observação e identificação das fases adultas que podem ser encontradas em
possíveis predadores de A.ucayalensis ou no estabelecimento de protocolos de
infecção experimental.
115
4.4.2 Ecologia do parasitismo
As diversas formas larvares deste estudo coabitam um mesmo nicho dentro
do hospedeiro Ageneiosus ucayalensis, onde provavelmente compartilham dos
mesmos nutrientes necessários á sua sobrevivência tendo em vista que exploram de
maneira semelhante a mesma classe de recursos nutritivos.
Do ponto de vista ecológico, duas espécies coexistem em um ambiente de
forma estável, se houver uma diferenciação de seus nichos, isto é, um espectro
limitado de condições e recursos que permitem a permanência de uma espécie,
mesmo na presença de competidores e predadores. Caso contrário, uma das
espécies eliminará ou excluirá a outra (Peroni et al., 2011). No entanto o que nós
observamos é que os três Morfotipos larvares (I, II e III) estão em um mesmo nicho;
a bolsa cística rica e complexamente vascularizada. A difusão de nutrientes
proporcionada pela arquitetura tecidual e vascular desta bolsa não permite a
compartimentalização de nicho e ou aporte de nutrientes, promovendo a ampla
distribuição de nutrientes.
Os diferentes morfotipos apresentam diferenças significativas de tamanho
corpóreo. Segundo Peroni et al. (2011), a competição entre espécies coexistentes
em um mesmo ambiente é resultado da diferenciação dos nichos e ambos são
evidenciados por padrões morfológicos em que, animais que competem fortemente
tendem a exibir diferenças regulares em tamanho do corpo. Como verificado
anteriormente, não há diferenciação de nicho e logo não há competição entre as
espécies. Deste modo, os diferentes tamanhos dos três Morfotipos larvares podem
estar relacionados a características genéticas implícitas à espécie de origem de
cada larva. Além disso, observamos que o Morfotipo I é mais abundante que os
Morfotipos II e III, sendo o Morfotipo III o menos frequente de todos. Isto pode estar
relacionado ao facto das espécies I, II e III ocorrerem com diferentes índices de
frequência na natureza.
Com base nestas observações, nós acreditamos que o que pode estar
acontecendo é a união das larvas I, II e III, promovendo a modulação conjunta do
hospedeiro e consequentemente remodelação tecidual em prol da nutrição e
manutenção de todos. Em um processo inicial, poucas larvas vão sendo adquiridas
pelo
peixe
e
vão
invadindo
a
cavidade,
localizando-se
dispersamente.
Provavelmente produtos de excreção/secreção (E/S) promoveriam inicialmente a
116
aglomeração das larvas I e II e com a proximidade e aumento do número, estas
podem entrar num segundo momento de síntese e sinalização de produtos de E/S
que induzem a neoformação de vasos e tecido conjuntivo, tornando-se perceptíveis
ao hospedeiro que os encerra em uma bolsa cística. Nas infecções sucessivas
proporcionadas pela alimentação do peixe, as larvas que chegarem posteriormente
ao processo de angiogênese migrariam por quimiotaxia ao sítio formado
anteriormente pelas larvas e somariam ao grupo, inclusive na sincronização da
síntese de produtos de E/S fazendo o aumento da bolsa cística e a manutenção da
neoformação tecidual e de vasos.
Considerando que as larvas dos Morfotipos I e II são mais numerosas que as
larvas do Morfotipo III, esta última pode ser oportunista e se associar à estrutura
induzida pelas larvas dos Morfotipos I e II e com isto garantir a sobrevivência ou
compartilhar da capacidade de produzir esses produtos de E/S, porém por ocorrer
em menor número, necessita desta associação das espécies I e II para ocorrer e
sobreviver.
117
5
CONCLUSÕES
a)
As formas larvares de Cestódeos observadas neste estudo parasitando
Ageneiosus ucayalensis pertencem à Família Proteocephalidae e foram
agrupadas em três morfotipos diferentes com grande probabilidade de
pertencerem a três gêneros e espécies distintas. Essas espécies podem estar
em fases diferentes do ciclo de vida, estando o primeiro Morfotipo na fase
Plerocercóide e o segundo e terceiro Morfotipos na fase Merocercóide.
b)
A presença dessas larvas na cavidade celomática de Ageneiosus
ucayalensis estimula a proliferação celular resultando em um tecido
conjuntivo, rico em fibras e células, altamente irrigado que envolve as larvas,
conferindo-lhes a sustentação, proteção e alimentação e, atuam no transporte
de oxigênio e nutrientes aos parasitos, e na condução da excreção de
produtos metabólicos dos parasitos através dos vasos sanguíneos.
c)
A membrana cística e a membrana do endotélio dos vasos sanguíneos
estabelecem contato íntimo através de junções evidenciando a troca de
metabólitos entre esses compartimentos.
d)
Os parasitos absorvem os metabólitos do compartimento cístico por
meio das microtríquias presentes na superfície corporal e através de
transporte vesicular do tipo pinocitose.
e)
Ageneiosus ucayalensis pode ter participação tanto como hospedeiro
paratênico quanto hospedeiro intermediário para o ciclo de vida desses
parasitos, em virtude das formas larvares se encontrarem em fases
diferentes.
f)
As diversas formas larvares deste estudo coabitam em um mesmo
nicho dentro do hospedeiro Ageneiosus ucayalensis, compartilhando
nutrientes necessários à sua sobrevivência, sem a necessidade de eliminação
de uma dessas formas.
118
PRESPECTIVAS DE ESTUDOS FUTUROS
1. Identificação de formas adultas dos três Morfotipos larvares por infecção
experimental.
2. Utilização da técnica de Reação em cadeia da Polimerase em Tempo
Real (RT-PCR) para identificação da expressão de genes de Fator de
Crescimento de Fibroblastos e Fator de Crescimento Endothelial
Vascular, nos parasitos e em tecidos vizinhos.
3. Estudos de Biologia Molecular com outros genes além do gene 18S para
separação mais específica entre as formas larvares.
4. Estudos de Proteômica das três formas larvares para caracterização de
substâncias angiogênicas e supressoras de sistema imume.
5. Estudos de influência de extratos somáticos das larvas em meio de cultivo
cellular para verificação de inibição de macrófagos, ativação de
fibroblastos e ativação de células endoteliais.
119
REFERÊNCIAS
AGUSTI, C., AZNAR, F.J., OLSON, P.D., LITTLEWOOD, D.T.J., KOSTADINOVA,
A., RAGA, J.A. Morphological and molecular characterization of tetraphyllidean
merocercoids (Platyhelminthes:Cestoda) of striped dolphins (Stenella
coeruleoalba) from the Western Mediterranean. Parasitology 130: 461–474,
2005.
ALICIA, A., DE PERTIERRA, G. Nomimoscolex semenasae n.sp. (Proteocephalidea:
Monticelliidae), acesto de parasite of Diplomystes viedmensis (Pisces:
Siluriformes) from the Patagonian region of Argentina. Systematic Parasitology
53: 183–190, 2002.
ALVAREZ, J.I., COLEGIAL, C.H., CASTANO, C.A., TRUJILLO, J., TEALE, J.M.,
RESTREPO, B.I. The human nervous tissue in proximity to granulomatous
lesions induced by Taenia solium metacestodes displays an active response.
Journal of Neuroimmunology 127: 139– 144, 2002.
ALVAREZ, J.I., LONDONÄO, D.P., ALVAREZ, A.L., TRUJILLO, J., JARAMILLO,
M.M., RESTREPO, B.I. Granuloma Formation and Parasite Disintegration in
Porcine Cysticercosis: Comparison with Human Neurocysticercosis. Journal of
Comparative Pathology 127: 186-193, 2002.
ALVAREZ, J.I., MISHRA, B.B., GUNDRA, U.M., MISHRA, P.K., TEALE,
J.M.Mesocestoides corti intracranial infection as a murine modelfor
neurocysticercosis. Parasitology 1-14, 2009.
ALVES, M.P. Avaliação imunohistoquímica para as galectinas 1, 3, 4 e 7 em
carcinoma epidermoide de língua.Tese, (Programa de Pós-Graduação em
Patologia Oral) - Universidade Federal de Rio Grande do Norte, 2009.132p.
ANDRADE, Z.A. Schistosomiasis and liver fibrosis. Parasite Immunology 31: 656–
663, 2009.
ANDRADE, Z.A., SANTANA, T.S. Angiogenesis and schistosomiasis. Mem. Inst.
Oswaldo Cruz 105 (4): 436-439, 2010.
ANIKIEVA, L.V., MALAKHOVA, R.P. & IESHKO, E.P. Ecological analysis of
parasites of coregonid fish. Leningrad, Publ, 1983. 168 p.
120
ATTOUT, T., HOERAUF, A., LLE DENECE, G., DEBRAH, A.Y., MARFODEBREKYEI, Y., BOUSSINESQ, M., WANJI, S., MARTINEZ, V., MAND, S.,
ADJEI, O., BAIN, O., SPECHT, S., MARTIN, C. Lymphatic Vascularisation and
Involvement of Lyve-1+ Macrophages in the Human Onchocerca Nodule. Plos
One 4 (12): 1-9, 2009.
BAO, P. The role of Vascular Endothelial Growth Factor in Wound Healing. Journal
of Surgical Research 153 (1016): 347-358, 2008.
BAPTISTA, A.P., ANDRADE, Z.A. Angiogenesis and schistosomal granuloma
formation. Mem. Inst. Oswaldo Cruz 100 (2): 183-185, 2005.
BEFUS, A.D. Hymenolepis diminuta and Hymenolepis microstoma: Mouse
immunoglobulins binding to the tegumental surfasse. Experimental
Parasitology 41: 242-251, 1977.
BIAN, X.W., CHEN, J.H., JIANG, X.F., BAI, J.S., WANG, Q.L., ZHANG, X.
Angiogenesis as an immunopharmacologic target in inflammation and cancer
International Immunopharmacology 4: 1537-1547, 2004.
BRÄNNSTRÖM, K., SELLIN, M.E., HOLMFELDT, P., BRATTSAND, M.,
GULLBERG, M. The Schistosoma mansoni protein Sm16/SmSLP/SmSPO-1
assembles into a 9-subunit oligomer with potential to inhibit Tolllike receptor
signaling. Infect Immunology, 2009.
BREHM, K., SPILIOTIS, ZAVALA-GONGORA, R., KONRAD, C., FROSCH, M. The
molecular mechanisms of larval cestode development: First steps into an
unknown world. Parasitology International 55: 15-21, 2006.
CAO, Y. Positive and Negative Modulation of Angiogenesis by VEGFR1 Ligands.
Science Signaling 2 (59): 1-11, 2009.
CARMENA, D., BENITO, A., MARTÍNEZ, J., GUISANTES, J.A. Preliminary study of
the presence of antibodies against excretory secretory antigens from
protoscoleces of Echinococcus granulosus in dogs with intestinal echinococcosis.
Mem. Inst. Oswaldo Cruz 100 (3): 311-317, 2005.
CARMELIET, P., JAIN, R.K. Molecular mechanisms and clinical applications of
angiogenesis. Nature 473: 298-307, 2011.
121
CAVALCANTI, M.G.S., ARAUJO, H.R.C., PAIVA, M.H.S., SILVA, G.M., BARBOSA,
C.C.G.S., SILVA, L.F., BRAYNER, F.A., ALVES, L.C. Ultrastructural and
cytochemical aspects of Schistosoma mansoni cercaria. Micron 40: 394–400,
2009.
CHAMBRIER, A., SCHOLZ, T. Tapeworms (Cestoda: Proteocephalidea) of firewood
catfish Sorubimichthys planiceps (Siluriformes: Pimelodidae) from the Amazon
River. Folia Parasitologica 55: 17–28, 2008.
CHAPPELL, L. H. 1980. Physiology of Parasites. Blackie, Glasgow, London.
CHERVY, L. The terminology of larval cestodes or metacestodes. Systematic
Parasitology 52: 1–33, 2002.
CHOI, W.W.L., LEWIS, M.M., LAWSON, D.Y.G.Q., BIRDSONG, G.G., COTSONIS,
G.A. Angiogenic and lymphangiogenic microvessel density in breast carcinoma:
correlation with clinicopathologic parameters and VEGFfamily gene expression.
Modern Pathology 18: 143–52, 2005.
COGGINS, J. R. Apical end rgan structure and histochemistry in Plerocercoids of
Proteocephalus ambloplitis. International Journal for Parasitology 10: 97-101,
1979
COIL, W.H., Platyhelminthes : Cestoidea In : Microscopic Anatomy of
Invertebrates, Vol. 3 : Platyhelminthes and Nemertinea (Harrison, F. W. and
Bogitsch, B. J., eds.).Wiley-Liss, New York, 211-283. 1991
DAINICHI, T., MAEKAWA, Y., ISHII, K. Nippocystatin, a cysteine protease inhibitor
from Nippostrongylus brasiliensis, inhibits antigen processing and modulates
antigen-specific immune response. Infect Immunology 69: 7380–7386, 2001.
DEBRAH, A.Y., MAND, S., TOLIAT, M.R., MARFO-DEBREKYEI, Y., BATSA, L.,
LAWSON, P.N., ADJEI, B.O., HOERAUF, A., PFARR, K. Plasma Vascular
Endothelial Growth Factor-A (VEGF-A) and VEGF-A Gene Polymorphism are
Associated with Hydrocele Development in Lymphatic Filariasis. American
Society of Tropical Medicine and Hygiene 77 (4): 601–608, 2007.
DENNIS, R.D., SCHUBERT, U., BAUER, C. Angiogenesis and parasitic helminth
associated neovascularization. Parasitology 138: 426–439, 2011.
122
DEY, C., MISRA, K.K. Ultrastructural studies of internuncial processes in the
tegument of a cyclophyllid cestode Raillietina (Fuhrmannetta) echinobothrida, a
parasite of country fowl Gallus domesticus. Proc. zool. Soc. 62 (1): 29-37, 2009.
DIAS, P.F., RIBEIRO-DO-VALLE, R.M., MARASCHIN, R.P., VENDRUSCOLO, L.F.,
MARASCHIN, M. Na formação dos vasos sangüíneos. Fronteiras entre saúde e
doença. Ciência Hoje 32 (190): 22-27, 2003.
DIAS, P.F., RIBEIRO-DO-VALLE, R.M., MARASCHIN, R.P., MARASCHIN, M. Novos
moduladores da formação de vasos sanguíneos. Biotecnologia Ciência &
Desenvolvimento 25: 28-32, 2002.
DORSEY, C.H., COUSIN, C.E., LEWIS, F.A., STIREWALT, M.A. Ultrastructure of the
Schistosoma mansoni Cercaria. Micron 33: 279-323, 2002.
DVIR, E., CLIFT, S.J. Evaluation of selected growth factor expression in canine
spirocercosis (Spirocerca lupi)-associated non-neoplastic nodules and sarcomas.
Veterinary Parasitology 174: 257–266, 2010.
ESTERRE P., PLICHART C., HUIN-BLONDEY M.O., NGUYEN L.N. Soluble cellular
adhesion molecules, selectins, VEGF and endothelin- 1 in patients with
Wuchereria bancrofti infection and association with clinical status. Parasite
Immunology 27: 9–16, 2005.
FALAVIGNA, D.L.M., VELHO, L.F.M., PAVANELLI, G.C. Proteocephalidean Larvae
(Cestoda) in Naturally Infected Cyclopid Copepods of the Upper Paraná River
Floodplain, Brazil. Mem. Inst. Oswaldo Cruz 98 (1): 69-72, 2003.
FECHINE-JAMACARU, F.V., JÚNIOR, J.U.F., FILHO, M.O.M. Model of inflammatory
angiogenesis in rabbit cornea induced by punctual alkaline cauterization. Acta
Cirúrgica Brasileira 20 (1) 64-73, 2005.
FERRARA, N. Vascular endothelial growth factor: basic science and clinical
progress. Endocrine Reviews 25 (4): 581-611, 2004.
FERRARA, N., CARVER-MOORE, K., CHEN, H., DOWN, M., LI, L., OSHEA, K.S.,
POWELL-BRAXTON, L., HILLAN, K.J., MOORE, M.W. Heterozygous embryonic
lethality induced by targeted inactivation of the VEGF gene. Nature 380: 439442, 1996.
123
FERREIRA, L.A., ANDRADE, Z.A. Capillaria hepatica: a cause of septal fibrosis of
the liver. Mem. Inst. Oswaldo Cruz 88: 441-447, 1993.
Fishwise - Ageneiosus ucayalensis - Castelnau 1855. Disponível em
www.fishwise.co.za › acesso em 12 de agosto de 2011.
FLOHR, C., QUINNELL, C.R.J., BRITTON, J. Do helminth parasites protect against
atopy and allergic disease? Clinical and Experimental Allergy 39: 20–32,
2008.
FOLKMAN, J. Role of angiogenesis in tumor growth and metastasis. Seminars in
Oncology 29 (6): 15-18, 2002.
FONSECA, A.A.H., PEREIRA, M.J.S. Classificação e morfologia de plathelmintos em
medicina veterinária: Cestoides. Seropédica, 2002.
FOUQUET, B., WEINSTEIN, B.M., SERLUCA, F.C., FISHMAN, M.C. Vessel
Patterning in the Embryo of the Zebrafish: Guidance by Notochord.
Developmental Biology 183: 37-48, 1997.
FREEMAN, R.S. On the biology of Proteocephalus parallacticus MacLulich (Cestoda)
in Algonquin Park, Canada. Canadian Journal of Zoology 42, 387-408, 1964.
FROESE, R., PAULY, D. FishBase. World Wide Web Electronic Publication,
Disponível em www.fishbase.org, version, 2010.
JENSEN, K., BULLARD, S. A. Characterization of a diversity of tetraphyllidean and
rhinebothriidean cestode larval types, with comments on host associations and
life-cycles. International Journal for Parasitology 40: 889-910, 2010.
GABAN, L., RAMOS, C.D.L., JUNIOR, A.A.B., SOUZA, M.M., ANDRADE, Z.A.
Dynamics of Capillaria-hepatica-induced hepatic septal fibrosis in rats.
Dinâmica da fibrose septal hepática induzida por Capillaria hepatica em ratos.
Revista da Sociedade Brasileira de Medicina Tropical 43 (6): 643-646, 2010.
GIESE, E.G., FURTADO, A.P., LANFREDI, R.M., SANTOS, J.N. A New Cucullanid
Species (nematoda) from Ageneiosus ucayalensis castelnau, 1855 (pisces:
auchenipteridae) from Pará Brazil. Journal of Parasitology 96 (2) 389-394,
2010.
124
GIESE, E.G. Biodiversidade de helmintos de Ageneiosus ucayalensis
Castelnau 1855 (pisces siluriformes) da Foz do rio Guamá e baía do
Guajará, Belém, Pará. Tese (Doutorado em Biologia de Agentes Infecciosos e
Parasitários) – Belém, Universidade Federal do Pará, 2010.157F.
GIESE, E.G., SANTOS, J.N., LANFREDI, R.M.A New Species of Camallanidae from
Ageneiosus ucayalensis (pisces: siluriformes) from Para State, Brazil.Journal of
Parasitology 95 (2): 407-412, 2009.
GOTTSTEIN, B., HEMPHILL, A. Echinococcus multilocularis: The parasite–host
interplay. Experimental Parasitology 119: 447-452, 2008.
HELLEMOND, J.J.V., RETRA, K., BROUWERS ,J.F.M., BALKOM, B.W.M.,
YAZDANBAKHSH, M., SHOEMAKER, C.B., TIELENS, A.G.M. Functions of the
tegument of schistosomes: Clues from the proteome and lipidome. International
Journal of Parasitology 36: 691-699, 2006.
HEWITSON, J.P., GRAINGER, J.R., MAIZELS, R.M. Helminth immunoregulation:
The role of parasite secreted proteins in modulating host immunity. Molecular
and Biochemical Parasitology 167 (1-9): 1-11, 2009.
HIGAZI, T.B., PEARLMAN, E., WHIKEHART, D.R., UNNASCH, T.R. Angiogenic
activity of an Onchocerca volvulus Ancylostoma secreted protein homologue.
Molecular & Biochemical Parasitology 129: 61-68, 2003.
HOPKINS, C.A., LAW, L.M. Schistocephalus solidus: Pinocytosis by the plerocercoid
tegument. Experimental Parasitology 44: 161-172, 1978.
HYPSA, V., SKERIKOVA, A., SCHOLZ, T. Phylogeny, evolution and host–parasite
relationships of the order Proteocephalidea (Eucestoda) as revealed by
combined analysis and secondary structure characters. Parasitology 130,
p.359–371. 2005.
JACKSON, J.A., FRIBERG, I.M., LITTLE, S., BRADLEY, E.J. Review series on
helminths, immune modulation and the hygiene hypothesis: Immunity against
helminths and immunological phenomena in modern human populations:
coevolutionary legacies? Immunology 126: 18-27, 2008.
JO, J.O., KIM, S.R., BAE, M.K., KANG, Y.J., OCK, M.S., KLEINMAN, H.K., CHA,
H.J. Thymosin β4 induces the expression of vascular endothelial growth factor
(VEGF) in a hypoxia-inducible factor (HIF)-1α-dependent manner. Biochimica et
Biophysica Acta 1803:1244-1251, 2010.
125
JOHNSTON, M.J.G., MACDONALD, J.A., MCKAY M. Parasitic helminths: a
pharmacopeia of anti-inflammatory molecules. Parasitology 136: 125-147, 2009.
JONES, M.K. Structure and diversity of cestode epithelia. International Journal of
Parasitology 28: 903-923, 1998.
KANG, Y.J., JO, J.O., Cho, M.K., Yu, H.S., OCK, M.S., CHA, H.J. Trichinella spiralis
infection induces angiogenic factor thymosin b4 expression. Veterinary
Parasitology 181: 222- 228, 2011.
KANSE, S.M., LIANG, O., SCHUBERT, U., HAAS, H., PREISSNER, K.T.,
DOENHOFF, M.J., DENNIS, R.D. Characterisation and partial purification of
Schistosoma mansoni egg-derived pro-angiogenic factor. Molecular &
Biochemical Parasitology 144: 76-85, 2005.
KLAUS J.E. Helminths, allergic disorders and IgE-mediated immune responses:
Where do we stand? European Journal of Immunology 37: 1170-1173, 2007.
KNAPP, J., NAKAO, M., YANAGIDA, T., OKAMOTO, M., SAARMA, U.,
LAVIKAINEN, A., ITO, A. Phylogenetic relationships within Echinococcus and
Taenia tapeworms (Cestoda: Taeniidae): An inference from nuclear proteincoding genes. Molecular Phylogenetics and Evolution 61: 628-638, 2011.
KODEDOVA, I., DOLEZÏEL, D., BROUCÏKOVA, M., JIRKU, M., HYPSÏA, V.,
LUKESÏ, J., SCHOLZ, T. On the phylogenetic positions of the Caryophyllidea,
Pseudophyllidea and Proteocephalidea (Eucestoda) inferred from 18S RNA.
International Journal for Parasitology 30:1109-1113, 2000.
KREIDER, T., ANTHONY, R.M., URBAN JR, J.F., GAUSE, W.C. Alternatively
activated macrophages in helminth infections. Current Opinion in Immunology
19: 448-453, 2007.
LEMOS, Q.T., ANDRADE, Z.A. Angiogenesis and experimental
fibrosis.Mem. Inst. Oswaldo Cruz 105 (5): 611-614, 2010.
hepatic
LENZI, J.A., MOTA.E.M., MACHADO, M.P., VALE, L.S., VALE, B.S., ANDRADE,
Z.A., LENZI.H.L. Intestinal Fibrovascular Nodules Caused by Schistosoma
mansoni Infection in Calomys callosus Rengger, 1830 (Rodentia: Cricetidae): a
Model of Concomitant Fibrosis and Angiogenesis. Mem. Inst. Oswaldo Cruz 97:
117-127, 2002.
126
LEVRON, C., PODDUBNAYA, L.G., KUCHTA, R., FREEMAN, M., WANG, Y.H.,
SCHOLZ, T. Ultrastructure of the tegument of the cestode Paraechinophallus
japonicus (Bothriocephalidea: Echinophallidae), a parasite of the bathypelagic
fish Psenopsis anômala Invertebrate. Biology 127 (2): 153-161, 2008.
LEVY, A.P., LEVY, N.S., WEGNER, S. Transcriptional regulation of the rat vascular
endothelial growth factor gene by hypoxia. Journal Biology Chem 270: 13333–
13340,1995.
LOEFFLER, D.A., LUNDY, S.K., SINGH, K.P., GERARD, H.C., HUDSON, A.P.,
BOROS,
D.L. Soluble Egg Antigens from Schistosoma mansoni Induce
Angiogenesis-Related Processes by Up-Regulating Vascular Endothelial Growth
Factor in Human Endothelial Cells. Journal of Infectious Diseases 185: 16501656, 2002.
MAITY, A., BHATTACHARYA, D., BATABYAL, S., CHATTOPADHYAY, S., BERA,
A.K., KARMAKAR, P.K. Echinococcus granulosus of Buffalo in India: Partial
Characterization of Excretory-Secretory and Germinal Membrane Antigens.
Veterinary Research Communications 31: 457-460, 2007.
MELO, M.F.C., GIESE, E.G., SANTOS, J.N., SANTOS, C.P. First record of larval
Pseudoproleptus sp.(Nematoda: Cystidicolidae) in fish host. Acta Tropica 117:
212-215, 2011.
MIURA, K., FUKUMOTO, S., DIRGAHAYU, P., HIRAI, K. Excretory/secretory
products from plerocercoids of Spirometra erinaceieuropaei suppress gene
expressions and production of tumor necrosis factor-a in murine macrophages
stimulated with lipopolysaccharide or lipoteichoic acid. International Journal for
Parasitology 31: 39-47, 2000.
MALUCELLI, B.E., SAFATLE, A.M.V., BARROS, P.S.M., GUERRA, J.L. Implantation
of two biological membranes in a cornealmicropocket as an experimental model
for angiogenesis. Brazilian Journal of Veterinary Research and Animal
Science 39 (4): 189-195, 2002.
MARR, J.J., NILSEN, T.W., KOMUNIECKI, R.W. Molecular Medical Parasitology.
Academic press, 2003.488p.
MARSHALL, F.A., GRIERSON, A.M., GARSIDE, P., HARNETT, W., HARNETT,
M.M. ES-62, an immunomodulator secreted by filarial nematodes suppresses
127
clonal expansion and modifies effector function of heterologous antigen-specific
T cells in vivo. Journal of Immunology 175: 5817-5826, 2005.
MORAES, M.S. A Função do óxido nítrico no processo da angiogênese através
do controle da atividade do receptor de EGF. Tese (Doutorado em
Bioquímica), Universidade de São Paulo, 2008.
NAKAO, M., LAVIKAINEN, A., IWAKI, T., HAUKISALMI, V., KONYAEV, S., OKU, Y.,
OKAMOTO, M., ITO, A. Molecular phylogeny of the genus Taenia (Cestoda:
Taeniidae): Proposals for the resurrection of Hydatigera Lamarck, 1816 and the
creation of a new genus Versteria. International Journal for Parasitology 43:
427-437, 2013.
NICOLI, S., DE SENA, G., PRESTA, M. Fibroblast Growth Factor 2-induced
angiogenesis in zebrafish: the zebrafish yolk membrane (ZFYM) angiogenesis
assay. Journal of Cellular and Molecular Medicine 13: 2061-2068, 2008.
OLIVEIRA, S.A.L. Pesquisa de helmintos em musculatura e serosa abdominalde
peixes de importância comercial capturados no litoral norte do Brasil.
Dissertação (Mestrado em Ciência Animal) – Belém, Universidade Federal do
Pará, 2005.66p
OLSON, P.D., LITTLEWOOD, D.T.J., BRAY, R.A., MARIAUX, J. Interrelationships
and Evolution of the Tapeworms (Platyhelminthes: Cestoda). Molecular
Phylogenetics and Evolution 19 (3): 443-467, 2001.
OLSSON, A.K., DIMBERG, A., KREUGER, J., CLAESSON-WELSH, L. VEGF
receptor signalling — in control of vascular function. Molecular cell biology 7:
359-371, 2006.
PALM, H.W., POYNTON, S.L., RUTLEDGE, P. Surface ultrastructure of
plerocercoids of Bombycirhynchus sphyraenaicum (Pintner, 1930) (Cestoda:
Trypanorhyncha). Parasitology Research 84: 195-204, 1998.
PINHO M.S.L. Angiogênese: o gatilho proliferativo. Revista Brasileira de
Coloproctologia 25 (4): 396-402, 2005.
PERONI, N., HERNÁNDEZ, M.I.M. Ecologia de Populações e Comunidades.
Universidade Federal de Santa Catarina, 2011.453p.
128
QUINTAVALLE, C., GAROFALO, M., CROCE, C.M., CONDORELLI, G. ApoptomiRs‖
in vascular cells: Their role in physiological and pathological angiogenesis.
Vascular Pharmacology, 2011.
RASO, P., RASO, L.A.M., MELO, F.A., TAFURI, W.L. Schistosoma mansoni
granuloma in late evolutive phase, in a case of tumoral form in man. Revista da
Sociedade Brasileira de Medicina Tropical 45 (5): 627-632, 2012.
REGO, A.A. Problems of classification of South American Proteocephalids
(Cestoda).On a new classification for the group.Acta Scientiarum: Biological
Sciences 25 (1): 15-22, 2003.
___________ Scolex Morphology of Proteocephalid Cestodes Parasites of
Neotropical Freshwater Fishes. Mem.Inst.Oswaldo Cruz 94 (1): 37-52, 1999.
____________ Proteocephalid Cestodes parasites of Pseudoplatystoma (pisces,
pimelodidae) from south america. Revista Brasileira de Zoociências 4 (2): 269282, 2002.
____________ Estudos de Cestóides de peixes do Brasil.2.ª Nota: Revisão do
Gênero Monticellia La Rue, 1991 (Cestoda Proteocephalidae). Revista
Brasileira de Biologia 35 (3): 567 – 586, 1975.
REGO, A.A., CHUBB, F.C., PAVANELLI, G.C. Cestodes in South American
Freshwater Teleost Fishes: Keys to gen era and brief description os species.
Revista Brasileira de Zoologia 16 (2): 299-367, 1999.
REY, L.PARASITOLOGIA MÉDICA.Guanabara koogan S.A 2005.856p
RIBEIRO, O.M., MIRANDA, J.P., QUENTAL, F., DE LA TORRE, A., PORTELA,
R.C.Q.Relações tróficas entre peixes em uma área de várzea no Rio Solimões,
Amazônia Central. Curso de Campo Ecologia da Floresta Amazônica, 2003.
RISSAU, W.Mechanisms of angiogenesis. Nature 386: 671-674, 1997.
ROBERTS, L.S., JANOVY, JR. J. Foundation of Parasitology. Sixth edition. Mc
Graw Hill International Edition. 2000.
129
ROBINSON, M.W., MENON, R., DONNELLY, S.M., DALTON, J.P.,
RANGANATHAN, S. An integrated transcriptomic and proteomic analysis of the
secretome of the helminth pathogen, Fasciola hepatica: proteins associated with
invasion and infection of the mammalian host. Molecular & Cellular
Proteomics, 2009.
ROSS, M.H., PAULINA, W. Histologia, Texto e Atlas em Correlação com Biologia
Celular e Molecular, 5a Edição. Guanabara Koogan, 2008 908p.
RZEPECKA, J., LUCIUS, R., DOLIGALSKA, M., BECK, S., RAUSCH, S.,
HARTMANN, S. Screening for immunomodulatory proteins of the intestinal
parasitic nematode Heligmosomoides polygyrus. Parasite Immunology 28: 463472, 2006.
SAFATLE, A.M.V., BARROS, P.S.M., MALUCELLI.B.E., GUERRA, J.L. Implante de
duas membranas biológicas em microbolsa corneana como modelo experimental
de angiogênese. Journal of Veterinary Research and Animal Science 39 (4):
189-195, 2002.
SAMBROOK, J., FRITSCH, E. F., MANIATIS, T. Molecular Cloning. A Laboratory
Manual. New York: Cold Spring Harbor Laboratory, 1989. 417p.
SANTOS, G.M., JEGU, M., MERONA, B. Catalago de peixes comerciais do baixo
rio Tocantins; projeto Tucuruí. Manaus - AM, 1984.
______________, FERREIRA, E.J.G,; ZUANON, J.A.S. Peixes comerciais de
Manaus. Pro-Várzea p.144, 2006.
SCHMIDT, G.D., LARRY S.R. Foundations of Parasitology. Higher Education 8th
edition, 2009.701p.
SCHRAMLOVÁ, J., BLAZER, K. Cysticercus Bovis: Pinocytosis in the Cysticercus
Tegument. Folia Parasitológica 35: 223-226, 1988.
SCHOLZ, T. Life cycles of species of species of Proteocephalus Weinland 1858
(Cestoda: Proteocephalidae), parasitesof fishes in the Palearctic Region: a
review. Journal of Helminthology 73: 1-20, 1999.
SCHOLZ, T. Studies on the Development of the Cestode Proteocephalus Neglectus
La Rue, 1911 (Cestoda Proteocephalidae) under experimental conditions. Folha
Parasitologica 38: 39-55, 1991.
130
SCHOLZ, T., HANZELOVA, V. Tapeworms of the genus Proteocephalus Weinland,
1858 (Cestoda: Proteocephalidae), parasites of fishes in Europe. Studies of the
Academy of Sciences (2) 120 p, 1998.
SENGER, D.R., PERRUZZI, C.A., FEDER, J., DVORAK, H.F.A. Highly conserved
vascular permeabillity factor secreted by a variety of human and rodent tumor cell
lines. Cancer research 46 (11): 5629-5632, 1986.
SHARIATI, F., PÉREZ-ARELLANO, J.L., CARRANZA, C., LÓPEZ-ABÁN, J.,
VICENTE, B., AREFI, M., MURO, A. Evaluation of the role of angiogenic factors
in the pathogenesis of schistosomiasis. Experimental Parasitology 128: 44-49,
2011.
SHARIATI, F., PEREZ-ARELLANO, J.L., PEZ-AB, J.L.N., EL BEHAIRY, A.M.,
MURO, A. Role of angiogenic factors in acute experimental Strongyloides
venezuelensis infection. Parasite Immunology 32: 430-439, 2010.
SIKASUNGE, C.S., JOHANSEN, M.V., PHIRI, I.K., WILLINGHAM, A.L., LEIFSSON,
P.S. The immune response in Taenia solium neurocysticercosis in pigs is
associated with astrogliosis, axonal degeneration and altered blood–brain barrier
permeability. Veterinary Parasitology 160: 242-250, 2009.
SIRACUSANO, A., MARGUTTI, P., DELUNARDO, F. Molecular cross-talk in host–
parasite relationships: the intriguing immunomodulatory role of Echinococcus
antigen B in cystic echinococcosis. International Journal of Parasitology 38:
1371-1376, 2008.
Sistema
cardiovascular-Vasos
sanguíneos.
Disponível
em
http://www.auladeanatomia.com/cardiovascular/vasos.htm, acesso em 05 de
dezembro de 2012.
SMYTH, J.D. Intoduction to animal parasitology 3a Edição.Cambridge University
Press, 1994. 549p
SOBKY, E.EL., GOMHA, M., EL-BAZ, H., ABOL-ENEIN., SHAABAN, A.A. Prognostic
significance of tumor angiogenesis in Schistosoma-associated adenocarcinoma
of the urinary blader. BJU International 89: 126-132, 2002.
131
SOUZA, F.P.C., VITORINO, R.R., COSTA, A.P., JÚNIOR, F.C.F., SANTANA, L.A.,
GOMES, A.P. Schistosomiasis mansoni: general aspects, immunology,
pathogenesis and natural history. Esquistossomose mansônica: aspectos
gerais, imunologia, patogênese e história natural. Revista Brasileira de Clinica
Medica 9 (4): 300-307, 2011.
SPILIOTIS, M., KRONER, A., BREHM, K. Identification, molecular characterization
and expression of the gene encoding the epidermal growth factor receptor
orthologue from the fox-tapeworm Echinococcus multilocularis. Gene 323: 5765, 2003.
STADELMANN, B., SPILIOTIS, M., MULLER, J., SCHOLL, S., MULLER, N.,
GOTTSTEIN, B., HEMPHILL, A. Echinococcus multilocularis phosphoglucose
isomerase (EmPGI): A glycolytic enzyme involved in metacestode growth and
parasite-host cell interactions. International Journal for Parasitology 40: 15631574.2010.
TAKAHASHI, H., SHIBUYA, M. The vascular endothelial growth factor (VEGF)/VEGF
receptor system and its role under physiological and pathological conditions.
Clinical Science 109: 227-241, 2005.
TARSITANO, M., DE FALCO, S., COLONNA, V., MCGHEE, J. D., PERSICO, M. G.
The C. elegans pvf-1 gene encodes a PDGF/VEGF-like factor able to bind
mammalian VEGF receptors and to induce angiogenesis. FASEB Journal 20,
227–233, 2006.
TCHAICHA, J.H., REYES, S.B., SHIN, J. Are Differentially Regulated by b8 Integrin
Glioblastoma Angiogenesis and Tumor Cell Invasiveness. Cancer Research 71:
6371-6381, 2011.
THOMPSON, R.C.A., HAYTON, A.R.,; JUE SUE, L.P.Z. An Ultrastructural Study of
the Microtriches of Adult Proteocephalus tidsweili (Cestoda: Proteocephalidea).
Parasitenkd 64: 95-111, 1980.
THREADGOLD, L.T., DUNN, J., Taenia crassiceps: Regional variations in
ultrastructure and evidence of endocytosis in cysticercus tegument.
Experimental Parasitology 55: 121-131, 1983.
THREADGOLD, L., HOPKINS, C.A. Schistocephalus solidus and Ligula intestinalis:
Piynocytosis by tegument. Experimental Parasitology 51: 444-456, 1981.
132
TOBIA, C., SENA, G., PRESTA, M. Zebrafish embryo, a tool to study tumor
angiogenesis. International Journal of Developmental Biology 55: 505-509,
2011.
VAN DER GEER, P., HUNTER, T., LINDBERGR, A. Receptor protein tyrosine
kinases and their signal transduction pathways. Annual Review of Cell Biology
10: 251-337, 1994.
Vasos
Sanguíneos.
Disponível
em
http://www.cabuloso.xpg.com.br/AnatomiaHumana/SistemaCardiovascular/Vaso
s-Sanguineos.htm, acesso em 05 de dezembro de 2012.
WAESCHENBACH, A.B.L., WEBSTER, D.T.J., WOOD, L. Adding resolution to
ordinal level relationships of tapeworms (Platyhelminthes: Cestoda) with large
fragments of mtDNA. Molecular Phylogenetics and Evolution 63: 834-847,
2012.
WARREN, K.S. The pathogeneses of ―clay-pipe atem cirrhosis‖ in mice with chronic
schistosomiasis mansoni, with a note on the lengevity of the schistosomiase.
American Journal of Pathology 49: 477-89, 1966.
WEINBERG, D.F., CORREA, E.B., LENZI, H.L., BOROJEVIC, R. Schistosoma
mansoni: Peritoneal Plasmacytogenesis and Polypoid Transformation of
Mesenteric Milky Spots in Infected Mice. Experimental parasitology 74: 408416, 1992.
WOHLLEBEN, G., TRUJILLO, C., MULLER, J., RITZE, Y., GRUNEWALD, S.,
TATSCH, U., ERB, K.J. Helminth infection modulates the development of
allergeninduced airway inflammation. Internacional Immunology 16: 585-596,
2004.
WOOD D. E. Nature of the end organ in Ophiotaenia filaroides (La Rue). Journal of
Parasitology 51: 541-544, 1965.
YAMAGUTI, S. Systema Helminthum, Volume II. The cestode of Vertebrates.
Interscience Publishers, 1959. 860p.
YANG, R.Y., RABINOVICH, G.A., LIU, F.T. Galectins: Structure, Function and
Therapeutic potential. Molecular Medicine 10: 1-24, 2008.
133
YOSHIDA, W.B. Angiogênese, arteriogênese e vasculogênese: tratamento do futuro
para isquemia crítica de membros? Jornal Vascular Brasileiro 4 (4): 316, 2005.
ZACCONE, P., COOKE, A. Helminth mediated modulation of Type 1 diabetes (T1D)
International Journal for Parasitology 4: 311–318, 2013.
ZALEŚNY, G., HILDEBRAND, J. Molecular identification of Mesocestoides spp. from
intermediate hosts (rodents) in central Europe (Poland). Parasitology Reserche
110: 1055-1061, 2012.
ZAVALA-GONGORA, R., KRONER, A., WITTEK, B., KNAUS, P., BREHM K.
Identification and characterisation of two distinct Smad proteins from the fox-tapeworm
Echinococcus multilocularis. International Journal of Parasitology 33: 1665-1677,
2003.
ZEHNDER, M.P., MARIAUX, J. Molecular systematic analysis of the order
Roteocephalidea (Eucestoda) based on mitochondrial and nuclear rDNA
sequences. International Journal for Parasitology 29: 1841-1852, 1999.