OS PROTOZOÁRIOS
Texto extraído do Livro Zoologia dos Invertebrados – Ruppert e Barnes – 6ª edição
Os protozoários são uma reunião diversa de cerca de 80.000 organismos de célula única que possuem
organelas celulares conjugadas a membranas típicas (eucarióticas). Como muitos são semelhantes a
animais, sendo móveis e apresentando uma nutrição heterotrófica, esse grupo foi tratado no passado
como um filo único dentro do Reino Animal - o filo Protozoa. Eles são hoje colocados dentro de um
número de diferentes filos unicelulares, que junto com a maioria dos filos de algas, constituem o reino
Protista, também chamados Protoctista. Protozoa é hoje utilizado como um nome comum conveniente
para protistas móveis unicelulares.
O nível de organização unicelular é a única característica pela qual os protozoários podem ser descritos
como um todo; em todos os outros aspectos, eles exibem uma diversidade extrema. Os Protozoa exibem
todos os tipos de simetria, uma grande variedade de complexidade estrutural e adaptações para todos os
tipos de condições ambientais. Embora a maioria dos protozoários exista como indivíduo solitário, há
numerosas formas coloniais. Algumas delas, tais como as espécies de Volvox, atingem tal grau de
interdependência celular que alcançam um nível de estrutura tecidual multicelular. Os protozoários
independentes individuais variam em tamanho de um micrômetro, no caso do planctônico Micromon, até
uns poucos milímetros em alguns dinoflagelados, amebas e ciliados.
Os protozoários de vida livre ocorrem onde quer que se encontre presente umidade - no mar, em todos os
tipos de água doce e no solo. Existem também muitas espécies parasitas, comensais e mutualistas.
ORGANELAS DOS PROTOZOÁRIOS E FISIOLOGIA GERAL
O corpo do protozoário é geralmente limitado somente pela membrana celular. A rigidez ou a flexibilidade
do corpo e a sua forma são enormemente dependentes da natureza do citoesqueleto. Os citoesqueletos
localizam-se tipicamente, mas nem sempre, imediatamente abaixo da membrana celular, onde, junto com
ela e outras organelas, formam a película, um tipo de "parede corporal" dos protozoários. O citoesqueleto
compõe-se frequentemente de proteínas filamentosas delgadas, microtúbulos ou vesículas, ou de
combinações variadas dos três. As proteínas filamentosas podem formar uma malha de suporte densa
denominada epiplasma, corno, por exemplo, no caso da Euglena e dos ciliados. Os microtúbulos
peliculares são as organelas citoesqueléticas mais evidentes que ocorrem em todos os flagelados, nos
protozoários formadores de esporos e nos ciliados. Eles podem arranjar-se como um colete microtubular,
ou, como em alguns flagelados, os microtúbulos originam-se nos corpos basais flagelares e irradiam-se
para trás a partir deles para a extremidade oposta da célula como um tipo de esqueleto axial (axóstilo).
Tais microtúbulos lembram os microtúbulos de um fuso mitótico, que se irradiam entre os centríolos e
formam o aparelho mitótico. Em outros protozoários, tais como os radiolários e os heliozoários, os feixes
de microtúbulos irradiam-se para fora a partir de um centrossomo ou de estruturas semelhantes, com
cada feixe estendendo- se para o interior e suportando uma projeção celular em forma de raio (axópodo).
O centrossomo e os microtúbulos lembram os ásteres em forma de estrela que se formam ao redor dos
centríolos nos pólos do fuso mitótico.
As vesículas ocorrem imediatamente abaixo da membrana celular em muitos protozoários, tais como os
dinoflagelados, os protozoários formadores de esporos e os ciliados. Quando essas vesículas ficam
achatadas e formam uma camada mais ou menos contínua por baixo da membrana celular, elas são
denominadas alvéolos. Os alvéolos "vazios" (como os que ocorrem nos ciliados) podem ficar túrgidos e
ajudar a sustentar a célula Membrana celular. Nos dinoflagelados, são secretadas placas de celulose no
interior das vesículas alveolares para formar um esqueleto rígido.
Os esqueletos dos protozoários, como os dos metazoários, também podem ser exoesqueletos. São
secretados sobre a superfície externa e são geralmente chamados testes.
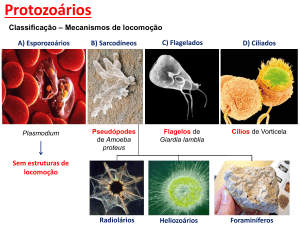
As organelas locomotoras dos protozoários podem ser os flagelos, os cílios ou extensões fluidas do corpo
chamadas pseudópodos. Essas organelas são importantes para definir os grupos de protozoários.
Ocorrem todos os tipos de nutrição nos protozoários. Alguns protozoários confiam no fotoautotrofismo,
outros absorvem nutrientes orgânicos solúveis de seu ambiente e podem ingerir partículas alimentares ou
predá-las e digeri-las intracelularmente no interior de vacúolos alimentares.
Como as outras organelas, as paredes dos vacúolos alimentares compõem-se de membranas de duas
camadas de lipídios, que podem ser sintetizadas ou removidas rapidamente e recicladas. O alimento
atinge o Vacúolo por meio de uma fagocitose, frequentemente através de uma boca celular ou citóstomo.
As partículas muito finas em suspensão podem entrar por meio de pinocitose, que pode ocorrer em toda
a superfície da célula. A digestão intracelular foi mais estudada nas amebas e nos ciliados.
Alguns protozoários (especialmente os que vivem no trato digestivo dos animais) são anaeróbicos
obrigatórios. Os que vivem em água onde houver uma decomposição ativa de matéria orgânica podem ser
anaeróbicos facultativos, utilizando o oxigênio quando estiver presente, mas também capazes de
respiração anaeróbica. A disponibilidade mutável de alimento e de oxigênio associada à degradação
resultam tipicamente em uma sucessão distinta de populações e espécies de protozoários. Devido ao seu
tempo de geração curto, os protozoários podem ser importantes na monitoração da poluição. A sua
resposta relativamente rápida às alterações no ambiente aquático podem ser indicativas de oxigênio
reduzido.
Muitos protozoários osmorregulam para remover o excesso de água (regulação do volume) e para ajustar
a concentração e a proporção de seus íons internos (regulação iônica). A água em excesso entra nos
protozoários através de osmose quando a concentração osmótica interna excede a da água circundante.
Pode entrar água adicional com o alimento nos vacúolos e na vesículas pinocitárias; por exemplo, uma
ameba experimentalmente alimentada em uma solução protéica absorve, através da endocitose, uma
quantidade de água equivalente a um terço de seu volume corporal. A osmorregulação é obtida através
de um transporte iônico ativo na membrana celular e através de um sistema de organelas de
bombeamento de água e de íons chamado de complexo vacuolar contrátil. O complexo compõe- se de
uma grande vesícula esférica - o Vacúolo contrátil propriamente dito - e de um sistema circundante de
pequenas vesículas ou túbulos denominado de espongioma. O espongioma proporciona a coleta de
fluido, que é entregue ao Vacúolo contrátil, que por sua vez expele o fluido para fora do organismo
através de um poro temporário ou permanente. Em alguns protozoários (algumas amebas e flagelados), o
Vacúolo desaparece completamente após a contração e refaz-se por meio de uma fusão de vesículas
pequenas. Em outros (muitos ciliados), o Vacúolo rompe-se na descarga e é repreenchido por fluido a
partir dostúbulos circundantes do espongioma. A taxa de descarga depende da pressão osmótica do meio
externo. O Paramecium caudatum (que vive na água doce) pode completar um ciclo de preenchimento e
descarga de Vacúolo rapidamente, em 6s, e expelir um volume equivalente ao seu corpo inteiro a cada
15min. A base para a contração pode diferir entre os grupos de protozoários. Nos dinoflagelados, uma
radícula flagelar ramifica-se para formar uma bainha contrátil ao redor do Vacúolo. No caso do
Paramecium, filamentos de actina circundam o Vacúolo. Nos protozoários de água doce, o conteúdo do
Vacúolo é mais diluído que o do citoplasma, porém mais concentrado que o do ambiente externo
circundante. Embora o mecanismo de função do Vacúolo contrátil não seja completamente
compreendido, parece atualmente provável que a água entra no espongioma à medida que as
membranas tubulares secretam íons provenientes do citoplasma no interior do túbulo. A água e os íons
passam ao longo do túbulo até uma região impermeável à água, na qual reabsorvem-se seletivamente
determinados íons e talvez outras substâncias. A água remanescente, os íons e talvez os metabólitos
residuais secretados são liberados da célula à medida que o Vacúolo se contrai. O sistema vacuolar
contrátil não tem nenhuma importância particular na remoção dos detritos metabólicos, tais como a
amônia e o CO2; esses difundem-se simplesmente para o lado externo do organismo.
Os protozoários com paredes celulares (significando que têm uma camada de celulose ou de outro
material exterior à membrana celular) não têm vacúolos contrateis. À medida que a água entra na célula
por osmose, a parede rígida resiste ao inchaço e estabelece-se uma pressão hidrostática. Quando a
elevação da pressão hidrostática iguala-se à pressão osmótica, não entra nenhuma água adicional na
célula. Como os animais, os protozoários respondem a estímulos químicos ou físicos de maneira a permitir
que evitem condições adversas, localizem alimento e achem parceiros. Nesse sentido, cada célula de
protozoário deve ser tanto receptora como efetora. Na sua receptividade aos estímulos ambientais, os
protozoários lembram as células nervosas sensoriais dos animais.
Como os receptores animais, os protozoários frequentemente recebem estímulos externos como
substâncias de sinalização que se conjugam com moléculas de membrana específicas. A conjugação pode
fazer com que se abra um canal iônico específico, permitindo que os íons (frequentemente Na+ e K+)
fluam para baixo de seu gradiente de concentração (Na+ dentro, K+ fora). Como a membrana celular em
repouso polariza-se em relação à distribuição desses íons, a abertura dos canais iônicos despolariza a
membrana. (As despolarizações podem ser medidas como alterações nos potenciais elétricos ou na
voltagem, utilizando eletrodos e um voltímetro.) Quando a membrana se despolariza, os canais de Ca++
abrem-se, e os íons de cálcio entram na célula. A entrada do cálcio no interior da célula dispara outras
alterações (tais como uma reversão do batimento ciliar), que fazem com que a célula se retire do
distúrbio. Por exemplo, o Paramecium tem pelo menos nove canais iônicos diferentes, alguns dos quais
localizados na frente e outros atrás da célula. Tais campos receptores localizados diferenciam a "cabeça"
da "cauda" e são conseqüentemente análogos à concentração de órgãos receptores de muitos animais. A
sinalização química intercelular (feromônios) nos protozoários, de fato, envolve frequentemente
moléculas de sinalização (tais como a serotonina, a B-endorfina, a acetilcolina e o AMP cíclico), que, nos
animais, funcionam como neurotransmissores e mensageiros internos.