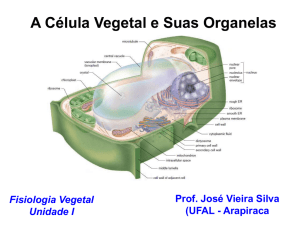
Membrana Plasmática
Funções
A membrana plasmática cumpre uma vasta gama de funções. A
primeira,
do
ponto
de
vista
da
própria
célula
é
que
ela
dá
individualidade a cada célula, definindo meios intra e extra celular.
Ela forma ambientes únicos e especializados, cuja composição e
concentração molecular são consequência de sua permeabilidade
seletiva e dos diversos meios de comunicação com o meio extracelular.
Além de delimitar o ambiente celular, compartimentalizando moléculas,
a membrana plasmática representa o primeiro elo de contato entre os
meios intra e extracelular,
transduzindo informações para o
interior da célula e permitindo que ela responda a estímulos externos
que podem, inclusive, influenciar no cumprimento de suas funções
biológicas.
Também
nas
interações
célula-célula
e
célula-matriz
extracelular a membrana plasmática participa de forma decisiva. É, por
exemplo,
através
de
componentes
da
membrana
que
células
semelhantes podem se reconhecer para, agrupando-se, formar tecidos.
A
manutenção
desempenho
da
das
individualidade
outras
funções
celular,
da
assim
membrana,
como
o
bom
requerem
uma
combinação particular de características estruturais da membrana
plasmática: ao mesmo tempo que a membrana precisa formar um
limite “estável”, ela precisa também ser dinâmica e flexível. A
combinação destas características é possível devido `a sua composição
química.
1
Composição química e estrutura
As membranas celulares consistem de uma dupla camada
contínua de lípides, com a qual proteínas e carboidratos das mais
diversas naturezas interagem das mais diversas maneiras…
Justamente a bicamada lipídica é que confere estabilidade e
flexibilidade, ao mesmo tempo, `a membrana. Pode-se dizer que os
lipídeos são os componentes que compõem a estrutura básica da
membrana. Existem 3 grandes classes de lipídeos que compõem a
membrana plasmática: fosfolipídeos, esteróis e glicolípides, sendo
que fosofolípides são os mais abundantes, via de regra.
A molécula de lípide possui uma característica bioquímica essencial
para formar uma bicamada estável, ainda que fluida. Ela possui uma
região hidrofílica e caudas hidrofóbicas. Enquanto que a região
hidrofílica interage bem com a água, altamente abundante nos
meios intra e extracelular, a região hidrofóbica busca “esconder-se”
da água. A intenção natural desta molécula anfipática, ou seja,
composta por regiões hidrofóbica e hidrofílica, de atingir um estado
que
seja
energeticamente
estável
e
termodinamicamente
favorável, faz com que elas arranjem-se na forma de uma bicamada.
A estabilidade é, então, dada pela necessidade termodinâmica do
próprio lípide em manter suas regiões hidrofílica e hidrofóbica em
posições adequadas em relação à água. Desta forma, se a bicamada
lipídica sofre um dano, onde algumas moléculas são removidas, sua
tendência
natural
é
a
de
se
regenerar.
Nos endereços abaixo você pode aprender mais sobre a estrutura dos
principais
lípides
de
membrana.
http://www-class.unl.edu/bios201/chapter4WEB/sld009.htm
http://www-class.unl.edu/bios201/chapter4WEB/sld010.htm
2
Os lipídeos distribuem-se assimetricamente nas duas monocamadas
lipídicas e estão em constante movimentação. Eles movem-se ao
longo do seu próprio eixo, num movimento chamado rotacional e
movem-se lateralmente ao longo da extensão da camada. Estes dois
movimentos não representam qualquer alteração `a termodinâmica
natural da membrana e, portanto, ocorrem constantemente. Um outro
movimento chamado flip-flop, que consiste em mudar de uma
monocamada `a outra, é menos frequente, pois envolve a passagem
da
cabeça
polar
(hidrofílica)
dentro
da
região
apolar
(hidrofóbica) da bicamada.
A fluidez da membrana é controlada por diversos fatores físicos e
químicos. A temperatura influencia na fluidez: quanto mais alta ou
baixa, mais ou menos fluida será a membrana, respectivamente. O
número de duplas ligações nas caudas hidrofóbicas dos lípides
também influencia a fluidez: quanto maior o número de insaturações,
mais fluida a membrana pois menor será a possibilidade de intração
entre moléculas vizinhas. Também a concentração de colesterol
influencia na fluidez: quanto mais colesterol, menos fluida. O colesterol,
por ser menor e mais rígido, interage mais fortemente com os lipídeos
adjacentes, diminuindo sua capacidade de movimentação.
Se os lípides são as moléculas mais expressivas em termos de
estrutura de membrana, as proteínas o são em termos de funções.
Considerando-se sua interação com a bicamada lipídica, as proteínas
podem
ser
classificadas
como:
ancoradas,
periféricas
ou
transmembrana (integrais). Naturalmente que as proteínas também
possuem características estruturais que as permitem interagir com a
bicamada lipídica: algumas delas possuem regiões polares e apolares,
sendo também anfipáticas.
3
Inúmeras
funções
são
desempenhadas
pelas
proteínas
de
membrana: elas comunicam célula e meio extracelular, servindo
como poros e canais, controlam o transporte iônico, servem como
transportadoras, realizam atividade enzimática e ainda podem ser
antigênicas, elicitando respostas imunes.
Os
carboidratos,
que
são
exclusivamente
encontrados
na
monocamada externa de membranas plasmáticas, interagem ora
com
proteínas
(glicoproteínas),
ora
com
lípides
(glicolípides),
formando uma estrutura denominada glicocálice. O glicocálice
desempenha inúmeras funções e elas refletem, na verdade, funções
desempenhadas por seus componentes. Por exemplo, a inibição do
crescimento celular por contato depende de glicoproteínas do
glicocálice. Se tais proteínas forem perdidas ou modificadas, como
acontece em alguns tumores malignos, mesmo o glicocálice ainda
existindo, esta função será comprometida. O glicocálice é importante na
adesão e reconhecimento celular, na determinação de grupos
sanguíneos, entre outras funções.
Glicocálice apresenta uma vasta gama de características interessantes…
Você
pode
aprender
mais
sobre
ele
no
endereço:
http://www.d.umn.edu/~sdowning/Membranes/glycocalyx.html
Transporte
Como a célula delimita o meio intra e extracelular, é necessário que
formas de transporte e de comunicação sejam constantemente
estabelecidas com o meio. Os transportes podem ou não envolver
gasto
de energia,
sendo
classificados
como
ativo
ou
passivo,
respectivamente. Exemplos de transporte passivo são as difusões
simples
e
facilitada.
As
bombas
de
íons
são
exemplos
de
4
transportes
ativos.
O endereço abaixo mostra de forma simples e elegante uma série de
figuras
compreensivas
sobre
transportes
celulares.
http://personal.tmlp.com/jimr57/textbook/chapter3/cmf1.htm
Também podem ser encontradas várias animações sobre transportes e
comunicações
com
o
meio
ambiente.
http://www.d.umn.edu/~sdowning/Membranes/imagesidx3.html
Especializações
As membranas celulares apresentam também diversos tipos de
especializações.
De
acordo
com
sua
localização,
podem
classificadas
ser
em:
Apicais:
Microvilosidades,
cílios
Laterais: zônula de adesão, de oclusão, interdigitações, desmossomas
Basais: pregas basais, hemidesmossomas
O complexo juncional é formado por junções oclusivas e adesivas. A
Zônula de Oclusão tem a função de vedar a passagem de
substâncias entre as células epiteliais, delimitando compartimentos
apicais, baso-laterais e intercelulares. É formada por ligações de
proteínas transmembranares de ocludina
entre células adjacentes,
formando um cinturão apical que une uma célula às outras que a
circundam. A Zônula de Oclusão associa-se aos microfilamentos de
actina através de moléculas importantes na sinalização intracelular.
A Zônula de Adesão forma também um cinturão contínuo ao redor
da célula que se une às adjacentes através de ligações entre moléculas
de
adesão
dependentes
de
Cálcio
(Caderinas).
Essas
proteínas
transmembranares estão ancoradas aos microfilamentos de actina
através de moléculas sinalizadoras.
5
A função principal da Zônula de Adesão é a de proporcinar a
coesão entre as células tornando a camada epitelial mais resistente
ao
atrito,
trações
e
pressões.
Além
disso,
participa
do
estabelecimento e manutenção da polaridade celular em associação com
a Zônula de Oclusão .
Os Desmossomas (“grampos”) formam regiões pontuais de
adesão entre as células e estão ancorados ao citoesqueleto de
filamentos intermediários através de moléculas com a desmoplaquina, a
desmina entre outras. Os desmossomas estão relacionados à forma e ao
padrão de distribuição das organelas na célula. Conferem resistência à
tração e à pressão. As Caderinas são as moléculas responsáveis pelas
ligações homeotípicas (entre moléculas da mesma família) entre as
células.
Os Hemi-desmossomas são regiões de união pontual entre células
e a lâmina ou membrana basal. São responsáveis pela adesão do tecido
epitelial ao tecido conjuntivo subjacente. A estrutura intracelular dos
hemidesmossomas é semelhante á dos desmossomas, mas as proteínas
transmembranares que ligam a célula às proteinas que formam a lâmina
basal são as integrinas e não as caderinas.
O endereço abaixo traz imagens excelentes de diversas especializações
de
membrana,
com
pequenos
textos
sobre
suas
funções.
http://www.med.uiuc.edu/histo/small/atlas/search.htm
6
Diferenciação
celular
e
interações
celulares
Para o estabelecimento dos metazoários, um dos eventos mais
importantes
foi
a
organização
de
folhas
epiteliais
que
separam
compartimentos externos e internos. A possibilidade de adesão e
comunicação entre as células através de junções e a polarização celular
em regiões basal e apical, parecem ser os principais requisitos para a
multicelularidade. Os epitélios são tecidos especializados que vedam
espaços e selecionam a passagem de substâncias entre diferentes
compartimentos.
Além
disso,
eles
participam
da
proteção
dos
organismos contra forças mecânicas, a desidratação, alterações da
microbiota, ação enzimática, extremos de pH e agentes patogênicos.
A formação de um espaço intercelular, a matriz extracelular,
possibilitou a manutenção
de
um
meio
mais constante
para
o
metabolismo e para trocas e interações entre as células. A criação desse
meio possibilitou também a diferenciação celular, sendo as atividades de
manutenção
do
organismo
assumidas
por
tipos
celulares
mais
especializados.
A diferenciação celular resulta da expressão diferencial de
genes e é um processo ontogenético que ocorre no
desenvolvimento dos seres multicelulares. Nesse sentido, é
uma sequência precisa de eventos que devem acontecer em tempos e
locais apropriados.
Gerando e mantendo estados de diferenciação estáveis.
Todas as células de organismos multicelulares se originam do ovo
fecundado. Elas proliferam-se, diferenciam-se, migram e interagem
7
umas com as outras e com a matriz extracelular, formando diferentes
tecidos e órgãos. A manutenção de células indiferenciadas (células
tronco ou precursoras) é de fundamental importância para a renovação
dos tecidos, cada qual com sua capacidade de regeneração e reposição.
Os epitélios e as células sangüíneas são os tecidos com maior taxa de
renovação e possuem células tronco bastante estudadas.
Morfologia
de
células
indiferenciadas
(blastos)
Existem células com potencial de gerar qualquer tipo celular e são
chamadas de células tronco totipotentes. São as células tronco
embrionárias, que formam o blastocisto. Elas representam um enorme
potencial terapêutico para a obtenção de tecidos para reconstituir
órgãos comprometidos. São as células utilizadas como receptoras na
clonagem.
Algumas células têm potencial de diferenciação limitado, podendo dar
origem a poucos tipos celulares, sendo chamadas de células tronco
pluripotentes. Elas iniciam sua diferenciação durante a gastrulação.
Essas
células
passaram
por
algumas
etapas
de
diferenciação
e
apresentam modificações permanentes (imprinting). Células tronco
podem ser obtidas do cordão umbilical, após o nascimento do bebê e
congeladas para utilização posterior.
Exemplos:
Células
Mesenquimais
(Tecido
Conjuntivo,
Adiposo,
Cartilaginoso,
Ósseo)
Células Tronco Hematopoiéticas (Precursoras de eritrócitos, monócitos,
linfócitos,
basófilos,
Células
Tronco
neutrófilos,
Epiteliais
eosinófilos
e
(intestino
megacariócitos)
e
pele)
Células Tronco Germinativas (dos testículos no macho adulto e no ovário
embrionário)
8
Processo
de
diferenciação
Falta muito para que possamos compreender este processo. É consenso
que para que ocorra a diferenciação a célula deve parar de proliferar. A
diferenciação depende de sinais provenientes de hormônios, da matriz
extracelular, de contato entre células e de fatores de diferenciação
chamados genericamente de citocinas. A orquestração dos sinais
recebidos pela célula resulta então na repressão de certos genes e
ativação de outros. Esse fenômeno pode alterar a forma da célula, seus
produtos para exportação e para sua própria estrutura e as moléculas
de sua superfície. Essas alterações refletirão no modo com que essa
célula interage com outras células e com a matriz extracelular. Ela pode
permanecer no lugar, proliferar ou migrar para outros tecidos ou outras
regiões do tecido. Como exemplo, veja o processo de queratinização do
epitélio.
Dados recentes das experiências com clonagem, mostram que os
núcleos de células adultas apresentam modificações permanentes
(imprinting) que refletem, de alguma maneira, a história dessa célula.
Marcadores
de
diferenciação
Durante a diferenciação celular, algumas proteínas são expressas
transitoriamente
e
podem
ser
detectadas
através
de
anticorpos
específicos produzidos em animais imunizados com essas proteínas.
Esses anticorpos são ferramentas importantes para o estudo da
diferenciação celular e no diagnóstico de leucemias e outros tipos de
câncer. Em alguns tumores, proteínas expressas apenas no estágio
embrionário voltam a ser expressas no adulto. Ex: A alfa-fetoproteína é
expressa principalmente no período fetal em contraposição à expressão
da albumina que ocorre principalmente após o nascimento. No câncer
hepático,
a
quantidades,
alfa-fetoproteína
podendo
passa
ser
a
ser
expressa
detectada
em
no
grandes
soro.
9
No desenvolvimento de linfócitos, vários marcadores são conhecidos por
exemplo o CD4, importante na interação linfocitária e também na
infecção pelo HIV.
Mecanismos
conservados
de
sinalização
intracelular
A interação das células com hormônios, neuro-transmissores, com
outras células e com a matriz extracelular ocorre geralmente através de
receptores na membrana celular ou dentro da célula. Essa ligação
desencadeia uma série de reações dentro da célula, como a liberação de
Ca2+ do REL, mudanças na concentração de AMP cíclico, a fosforilação e
desfosforilação de proteínas e o metabolismo de lipídios importantes.
Mediadores secundários são produzidos, podendo ser translocados para
o núcleo ativando ou reprimindo a expressão de diferentes genes. Além
disso, alterações do citoesqueleto podem alterar a forma e a adesão
celular.
Os mecanismos de sinalização intracelular são bastante conservados,
sendo alguns dos mediadores presentes em bactérias e em metazoários.
Moléculas
de
Adesão
Celular
As moléculas de adesão conferem à célula um tipo de endereço. Elas
podem ancorar células em tecidos específicos ou "endereçar" células
migrantes
a
determinadas
regiões
do
corpo.
Principais moléculas de adesão:
As selectinas interagem com açúcares nas glicoproteínas e na
Selectinas matriz. Promovem uma adesão fraca inical que pode dar
início às etapas posteriores
Moléculas de adesão dependentes da concentração de Ca+2.
Caderinas São importantes para adesão das células de um mesmo
tecido.
10
Ex. E-caderina nos epitélios. Associam-se ao citoesqueleto.
São moléculas importantes na adesão das células à lâmina
basal e demais componentes da matriz extracelular, assim
Integrinas como na interação linfocitária. Diferentes integrinas são
expressas em tecidos diferentes, que possuem matriz e tipos
celulares diferentes.Ex: a4b1 nas mucosas
InterCelular Adhesion Molecules. Pertencem à superfamília
ICAMs
das imunoglobulinas. Ex: V-CAM vascular, E-CAM endotélio,
N-CAM no tecido nervoso
A perda de certas moléculas de adesão em células tumorais pode levar
à migração das células para outros tecidos: Metástase.
Citoesqueleto
O Citoesqueleto das células eucariontes é composto de complexos
protéicos
fibrilares,
formados
pela
polimerização
de
proteínas
globulares. Sua pricincipal função é coordenar a distribuição de
organelas
na
célula
e
orientar
sua
forma
geral.
Ele é responsável pelas alterações de forma e da distribuição de
organelas desencadeadas por interações entre a célula e seu meio e
entre células diferentes. É também responsável pela sustentação e
resistência da célula.
O Citoesqueleto é composto por Microfilamentos, Filamentos Espessos,
Filamentos
Intermediários
e
Microtúbulos.
Esses
componentes
se
associam entre si, formando uma complexa rede citoplasmática.
Através de proteínas associadas, eles se ligam à membrana plasmática
e às membranas de outras organelas. É uma estrutura dinâmica, que se
altera
através
de
variações
entre
taxas
de
polimerização
e
11
despolimerização.
Componentes
Diâmetro Estrutura
Principal proteína
Participam de:
Contração muscular,
Microfilamentos
~5nm
ooooo
Actina
endocitose,
pinocitose, migração
celular
Filamentos
Espessos
~15nm ______oO
Miosina
Contração muscular
Citoqueratina
(células epiteliais)
Filamentos
Intermediários
~10nm
--o--o--o
----o---o
Vimentina(células
Sustentação:
mesenquimais)
desmossomas e
Neurofilamentos, etc. hemidesmossomas
Lamina (lâmina
nuclear)
Tubulina-a e
Microtúbulos
~25nm ::::::::::::::::
Tubulina-b
Cilindro oco de
diâmetro
Estruturas com microtúbulos
organizados
Formação do fuso
mitótico
Transporte de
vesículas e outras
organelas
Estrutura Localização
Flagelos
9+2
Espermatozóide e em protozoários
Cílios
9+2
Epitélio das viás aéreas
Centríolos
3x9
Corpúsculos Basais
3x9
Todas as células animais; região
organizadora de microtúbulos
Ancoragem e origem dos cílios
12
Associação do Citoesqueleto com a membrana plasmática:
As junções entre células podem apresentar diferentes funções de acordo
com
sua
estrutura.
A Junções de Oclusão são contínuas em torno da célula nos pontos de
contato com as células vizinhas e vedam compartimentos. São formadas
por
proteínas
transmembranares,
associadas
internamente
ao
citesqueleto de microfilamentos de actina. Ex: Lume Intestinal, bexiga,
vias
respiratórias.
As junções de adesão podem ser contínuas como a Zona de Adesão, ou
pontuais como os desmossomas e hemidesmossomas. A Zona de
Adesão associa-se aos microfilamentos de actina da malha próxima à
membrana
plasmática.
Os
desmossomas
e
hemidesmossomas
se
associam a filamentos intermediários. Ver: Junções
Doenças
Ação
associadas
de
drogas
a
alterações
sobre
do
o
citoesqueleto:
citoesqueleto:
Microtúbulos:
A Colchicina é um alcalóide que se liga aos dímeros da tubulina e
impede a polimerização dos microtúbulos mais sensíveis como os do
fuso mitótico. Ela é utilizada para se estudar células em mitose.
O Taxol é um alcalóide utilizado no tratamento de tumores pois
também impede a mitose. Ele acelera a polimerização e estabiliza os
microtúbulos, impedindo a despolimerização. Outras drogas utilizadas
no tratamento de tumores, que também interferem com a dinâmica de
microtúbulos, são a Vincristina e a vimblastina.
Microfilamentos:
Produtos de fungos como as citocalasinas e as faloidinas interferem com
a dinâmica de polimerização e despolimerização da actina, interferindo
com o movimento celular. As citocalasinas se ligam à actina impedindo a
13
polimerização, enquanto
microfilamentos
as faloidinas se
estabilizando-os.
ligam lateralmente
Filamentos
intermediários
aos
-
A
disposição dos filamentos intermediários depende da presença de
microtúbulos.
Organelas e Inclusões Citoplasmáticas
Mitocôndria: a produção da energia aeróbia
Estrutura Morfológica
Qualquer mitocôndria é formada por duas unidades de membrana
separadas por um espaço intermembranoso, cada qual com funções e
proteínas diferentes associadas à sua bicamada lipídica. A membrana
interna
emite
numerosas
cristas
para
o
interior
da
organela,
aumentando substancialmente a sua superfície. Nessas cristas pode-se
visualizar ao microscópio eletrônico em grande aumento, partículas em
forma de raquete, denominadas corpúsculos elementares. São
encontrados a intervalos de 10 nm, podendo haver 10 4 a 106
corpúsculos por mitocôndria.
Na intimidade da mitocôndria, delimitada pela membrana interna, está a
matriz mitocondrial. A matriz contém material protéico granular de
alta densidade, capaz de se ligar à fosfatos de cálcio e magnésio,
precipitando-se na forma de grânulos elétron-densos. É por isso que
as mitocôndrias são o segundo destino preferencial de acúmulo do cálcio
intracelular (o primeiro sendo o retículo endoplasmático liso). A matriz
possui todas as enzimas necessárias ao ciclo de Krebs.
14
A
membrana
externa
mitocondrial
é
semelhante
às
demais
membranas da célula eucarionte. A membrana interna, possui os
elementos da cadeia respiratória necessários para a respiração aeróbica.
Eletromicrografia de Intestino delgado de rato, mostrando
mitocôndrias.
Foto elaborada pelo aluno de mestrado Ramon Lamar de
Oliveira Junior. ICB-UFMG.
Clique na foto para maior
aumento
Cadeia Respiratória
A cadeia respiratória ocorre na membrana mitocondrial interna e
compreeende três complexos enzimáticos principais através dos quais
elétrons fluem do NADH para o O2, utilizando a energia daí gerada para
bombear H+ da matriz para o espaço intermebranoso. Na membrana
nativa, os carreadores de elétrons móveis ubiquinona e citocromo c
completam
a
cadeia
transportadora
de
elétrons
ao
mediar
a
transferência de elétrons entre os complexos enzimáticos. Por fim, os
elétrons são transferidos para o oxigênio molecular (O2 ), essencial ao
processo aeróbico, unindo-se a átomos de hidrogênio para formar água.
O gradiente eletroquímico de prótons resultante é adaptado para
sintetizar ATP por outro complexo protéico transmembrana, ATP
sintetase, através do qual H+ flui de volta à matriz. Esse complexo está
localizado nos corpúsculos elementares.
Quando a mitocôndria fica exposta a um meio citoplasmático com altas
concentrações de ADP, ela assume uma forma condensada. Isso se deve
15
ao fato de estar em síntese máxima de ATP e, junto com ele, de H2O,
que
se
acumula
no
espaço
intermembranoso.
Genoma Mitocondrial
A mitocôndria possui DNA próprio, o que talvez reflita o curso do
processo evolutivo. O genoma mitocondrial se restringe a uma fita de
DNA circular na célula animal, sendo o sistema genético mais simples
conhecido, onde todos os nucleotídeos fazem parte de sequências
codificantes. Nos vegetais, o DNA mitocondrial é 10 a 150 vezes maior,
mas a quantidade de proteínas sintetizadas é quase a mesma, pois
muitas sequências adicionadas não constituem genes nesses tipos
mitocondriais, chegando, até mesmo, a existir íntrons.
O núcleo deve fornecer cerca de 90 genes para a realização de funções
mitocondriais. Em animais, o genoma mitocondrial está na ordem de 10 5
do nuclear, o que corresponde a 16.500 pares de bases. Isso é
suficiente para que sejam capazes de sintetizar suas próprias proteínas
e se autoduplicarem. Em números, esse material genético é capaz de
codificar: 2 RNAs ribossomais, 22 RNAs transportadores e 13 cadeias
polipeptídicas. Como nas bactérias, o DNA mitocondrial não é envolto
em histonas e seu empacotamento não é bem explicado.
As mitocôndrias só possuem 22 RNA transportadores, enquanto na
célula tem-se 30. Isso faz o sistema de pareamento códon-anticódon
relaxado; muitos dos RNAts reconhecem qualquer nucleotídeo na
terceira posição dos códons, podendo se ligar a até quatro sequências
diferentes. Da mesma forma, o significado de 4 dos 64 códons possíveis
encontra-se
alterado
nas
mitocôndrias.
16
Síntese Protéica
Muitas das proteínas mitocondriais são produzidas a partir do próprio
DNA nuclear e sintetizadas no citoplasma. Uma boa parte da síntese
protéica mitocondrial consiste em polipeptídeos que precisam associarse
à
subunidades
produzidas
pela
célula.
O
transporte
destas
subunidades ocorre através de sítios de adesão entre as membranas
mitocondriais interna e externa, denominados sítios de contato.
Para que uma proteína celular chegue até a mitocôndria e se estabeleça
na sua matriz são necessários eventos de sinalização celular. As
proteínas de origem nuclear normalmente possuem peptídios de
sinalização que endereçam a cadeia protéica para a matriz da
mitocôndria. As proteínas atravessam as membranas impulsionadas por
um gradiente eletroquímico existente entre as faces da membrana
mitocondrial interna e pela energia do ATP, num estado desdobrado que
facilita o transporte. Já na matriz, a proteína pode associar-se à outras
subunidades mitocondriais e atinge a sua conformação efetiva.
A maior parte dos lipídios mitocondriais são importados de outros
compartimentos
celulares.
Os
endoplasmático
liso,
prontamente
são
lipídios,
sintetizados
aderidos
no
à
retículo
membrana
mitocondrial interna. A mitocôndria pode realizar simples modificações
nessas moléculas, podendo também atuar na conversão de lipídios
importados em cardiolipina, que constitui 20% dos lípides de sua
membrana
interna.
17
Hipótese Endossimbiótica, Multiplicação e Herança
O caráter procariótico do sistema genético das mitocôndrias, bem como
dos cloroplastos, sugere que essas organelas originaram-se de bactérias
endocitadas há mais de um bilhão de anos, quando o oxigênio
atmosférico terrestre atingiu níveis elevados. De fato, evidencia-se uma
grande
semelhança
entre
o
funcionamento
e
constituição
das
mitocôndrias e bactérias.
Segundo essa hipótese, as células eucarióticas iniciaram sua existência
estabelecendo
uma
relação
endossimbiótica
com
uma
bactéria,
responsável pelo sistema de fosforilação oxidativa. Ela seria uma
bactéria púrpura fotossintetizante que teria perdido a capacidade
fotossintética, se especializando na cadeia respiratória.
Durante a evolução eucariótica, ocorreu uma grande transferência de
genes das mitocôndrias para o núcleo celular, com o objetivo de
favorecer a mitocôndria na execução de uma única função principal: o
fornecimento energético. Isso explica a importação de proteínas
citoplasmáticas e a existência de algumas sequências não codificantes
no DNA nuclear, correspondendo ao DNA importado recentemente e
sem função.
A teoria ainda abre espaço para explicar a presença de duas
membranas lipídicas na organela. A membrana mitocondrial interna
seria originária da membrana da bactéria endocitada, enquanto a
membrana mitocondrial externa seria derivada da própria membrana
celular.
18
AUTODUPLICAÇÃO
As mitocôndrias sempre se originam de outras pré-existentes por fissão,
podendo também fundirem-se umas com as outras. Os processos de
duplicação e fusão são controlados e capazes de manter um número
sempre estável de mitocôndrias por célula.
O
número
de
mitocôndrias
pode
ser
ainda
regulado
de
forma
adaptativa; o músculo esquelético submetido a esforço prolongado, por
exemplo, possui 5 a 10 vezes mais mitocôndrias.
O DNA mitocondrial se duplica durante a intérfase, mas não só neste
período, e em tempos diferentes se comparadas as mitocôndrias de uma
mesma célula. A duplicação se assemelha àquela observada nas
bactérias.
HERANÇA GENÉTICA
Em seres inferiores, como as leveduras, a herança do DNA mitocondrial
é biparental, ou seja, proveniente dos dois organismos formadores.
Já nos seres onde ocorre maior especialização do sistema reprodutor, a
herança mitocondrial encontra-se sempre no óvulo, e nunca no
espermatozóide; isso implica que a herança é uniparental, ou mais
especificamente materna.
Tipos de Mitocôndria e Patologia
Especificidade Tecidual
As mitocôndrias variam entre os tecidos, adaptando-se à cada célula em
particular. Essa variação pode ocorrer no número, forma, localização,
constituição protéica e funções principais exercidas pelas mitocôndrias.
19
As mitocôndrias estão associadas ao citoesqueleto, sendo posicionadas
na célula pela sua movimentação em associação a microtúbulos e
microfilamentos. Normalmente se encontram próximas dos locais que
mais necessitam de energia em cada célula em particular.
Para evidenciar a especificidade tecidual das mitocôndrias veja alguns
exemplos :
Espermatozóide
O dispêndio de energia nos espermatozóides é, basicamente,
com o seu movimento flagelar. O movimento flagelar é
alimentado por proteínas motoras chamadas dineínas que
utilizam a energia proveniente do ATP para o deslizamento
dos microtúbulos. As mitocôndrias, neste caso, concentramse na porção anterior da cauda, onde o ATP é necessário,
fomando um ``espiral´´ em torno do segmento de flagelo.
Músculo Estriado
Esquelético
No músculo estriado esquelético, as mitocôndrias são
alongadas e ramificadas para maximizar a fosforilação
oxidativa, minimizando a distância que o ATP percorre por
difusão. Podem ser vistas nesta formação em corte
transversal; em corte longitudinal, as mitocôndrias são
visualizadas aos pares na altura das bandas I do espaço
interfibrilar.
As
mitocôndrias
do
músculo
estriado
esquelético são ricas em fosfocreatina. A fosfocreatina pode
reagir com o ADP, formando ATP e creatina, ou seja, produz
energia na ausência de oxigênio. Se a necessidade de
Clique na foto para maior aumento
oxigênio não é suprida rapidamente, o estoque de
Mitocôndrias de célula
fosfocreatina é consumido e a célula muscular não consegue
muscular
realizar as etapas intramitocondriais do ciclo de degradação
da glicose, acumulando ácido láctico. O excesso de ácido
láctico intracelular pode baixar o pH celular para até 6,34 o
que origina dor e fadiga, sintomas da cãimbra.
20
21