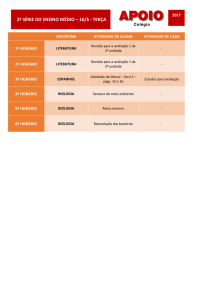
11/06/15
NG110 Métodos Computacionais em Bioinformá9ca Introdução a Biologia de Sistemas Prof. Renato Vicentini – [email protected]
•
•
•
•
NG264 Princípios da Biologia de Sistemas • Introdução • Conceitos básicos de Biologia de Sistemas Prof. Renato Vicentini – [email protected]
O método cienGfico A complexidade na Biologia Observação do sistema de interesse; Desenvolvimento de hipóteses sobre o sistema observado; Predições baseadas nas hipóteses propostas; Condução de experimentos que validem ou invalidem a predição (e conseqüentemente as hipóteses); • A biologia é uma disciplina experimental; • Mas bons experimentos são o resultado de elucubrações, planejamento e análises consideráveis; • Com as tecnologias experimentais se tornando mais complexas, estes três passos demandam cada vez mais o uso do intelecto; • Como resultado, a construção de modelos computacionais e a analises destes modelos se tornam parte integral da biologia; A complexidade na Biologia O ciclo de uma pesquisa em Biologia Molecular • Theorist vs. experimentalist jokes. Bancos de dados/literatura – Experimentalists observe things that cannot be explained – Theore9cians explain things that cannot be observed Analise de dados • The only true science is physics, everything else is stamp collec9ng Experimentos para testar as hipóteses Construção do modelo Analise do modelo Planejamento experimental Ernest Rutherford
Formulação de hipóteses 1
11/06/15
Biologia de Sistemas • A disciplina da Biologia de Sistemas examina: – como os componentes biológicos, em uma célula ou organismo, interagem e produzem redes de interação; – como as redes fornecem a função celular necessária para o fenó9po observado. A necessidade da integração das partes Biologia de Sistemas • Premissa: – A complexidade da biologia pode ser compreendida em termos da organização de seus componentes (“design principles”); • Generalizando: – Porque um sistema é construído da forma em que ele é ? Circuitos gené9cos • Funções celulares são resultantes de uma ação coordenada do produtos de múl9plos genes; O paradigma da Biologia de Sistemas As duas raízes da Biologia de Sistemas Os quatro passos principais na implementação da Biologia de Sistemas. Notem que o segundo passo é único e é a interface entre dados HT e analises in silico. 2
11/06/15
Compreendendo o sistema biológico como um sistema • Iden9ficação da estrutura do sistema: – Relacionamento entre os componentes (genes, proteínas, etc); – Microarrays, qRT-­‐PCR, PPI; • Analise do comportamento do sistema: Iden9ficação da estrutura do sistema • Bo/on-­‐up: – Compilação de dados experimentais independentes; • Top-­‐down: – Dados HT; – Resposta a es9mulo; • Controle do sistema: – Perturbação; • Design: – Biologia sinté9ca; A Biologia no tempo e no espaço • A descrição de en9dades biológicas e suas propriedades abrangem diferentes níveis de organização e diferentes escalas de tempo; O que é um modelo ? Modelos e Modelagem • Se observarmos os processos biológicos, seremos confrontados com vários processos complexos que não podem ser explicados pelos princípios básicos da biologia, e os resultados não podem ser previstos intui9vamente; • A modelagem matemá9ca e a simulação computacional podem nos ajudar a entender a natureza interna e a dinâmica destes processos; O que é um modelo ? • Alguns modelos podem ser aplicados a muitos elementos similares: – Ciné9ca de Michaelis-­‐Menten: várias enzimas; – TF-­‐promotor: vários genes; • Enquanto outros são específicos de um elemento em par9cular: – Modelo 3D de uma proteína; • Resolvendo as equações, são ob9das predições da concentração/tempo. Se estas predições não corroboram os dados experimentais, isto quer dizer que o modelo esta errado ou muito simplificado. 3
11/06/15
O que é um modelo ? Duas estratégias de modelagem matemá9ca • Os modelos são geralmente baseados em leis hsicas bem definidas: – ex: termodinâmica das reações químicas; Método exploratório Testa a hipótese “Assumindo estas interações, pode o sistema se comportar da forma como observamos?” Método predi9vo Extrapolação “O que acontece se nocautearmos estes genes?” Classes de redes biológicas Biologia de Sistemas Biologia de Sistema Biologia de Sistema • O sistema deve ser descrito igualmente tanto no nível dos genes, das vias bioquímicas, das organelas, das células, dos órgãos, do organismo, e da comunidade e ambiente (ecossistema). • Aumento de complexidade quando nos movemos da escala molecular para a escala do organismo como um todo. • Estudar processos em larga escala: – não como mecanismos individuais, mas a par9r de interações e regulações. • A unidade fundamental dos processos biológicos é a célula. • A melhor forma de construir uma ponte da biologia molecular para a fisiologia é reconhecer que as redes de interação formam um sistema dinâmico que evolui no espaço e no tempo de acordo com leis fundamentais. • Integrar informações complexas e altamente diversas em uma mesma estrutura conceitual: – sendo esta holís9ca, quan9ta9va e predi9va. 4
11/06/15
Redes de regulação ... Signal 1 Signal 2 Signal 3 Signal 4 Signal N Environment Transcrip9on factors Conceitos básicos de Biologia de Sistemas X2 gene 2 gene 3 X3 ... Xm genes gene 1 Prof. Renato Vicentini – [email protected]
Redes de regulação gene 4 gene 5 gene 6 ... gene k Redes de regulação promoter X DNA X1 X Ac3vator Y gene Y X binding site protein Y Y Y Sx X TRANSLATION mRNA gene Y RNA polymerase gene Y Y X* X* TRANSCRIPTION Y INCREASED TRANSCRIPTION Bound ac3vator Redes de regulação Rede de regulação em E. coli X Bound repressor Y • Representação de 20% das interações transcricionais conhecidas. Sx X X* NO TRANSCRIPTION X* Y Bound repressor Unbound repressor X Y Y Y 5
11/06/15
Modelagem de sistemas dinâmicos Modelagem de sistemas dinâmicos • Descreve a evolução temporal de um sistema usando um conjunto fixo de regras: – ODE derivação de uma ciné9ca enzimá9ca; Modelagem de sistemas dinâmicos Equação de Michaelis-­‐Menten • A obtenção de dados quan9ta9vos na biologia começou a cerca de um século atrás; • Quando Michaelis e Menten publicaram seu trabalho sobre os mecanismos da ciné9ca enzimá9ca (1913); • Eles mostraram que a taxa de uma reação enzimá9ca é hiperbólica e depende da concentração do substrato; • Base da maior parte da ciné9ca enzimá9ca mono-­‐substrato; Curva de saturação para uma enzima mostrando a relação entre
a concentração do substrato e a velocidade de reação.
Km (constante de Michaelis) é definida como a concentração
para a qual a velocidade da reação enzimática é metade de Vmax
O que é um sistema ? • Linearidade vs não-­‐linearidade Comportamento de sistemas dinâmicos Função linear de uma única entrada
Função não-linear de uma única entrada
• Analise do comportamento de sistemas dinâmicos: – Função linear de uma única entrada – Função não-­‐linear de uma única entrada 6
11/06/15
Linearidade vs não-­‐linearidade • O comportamento de sistemas não lineares é geralmente dihcil de predizer; • Sistemas não lineares são a norma, e não a exceção em biologia; • GRN e redes de interação bioquímicas apresentam geralmente um comportamento dinâmico não-­‐linear; Modelagem ciné9ca de reações enzimá9cas • O modelo de Michaelis-­‐Menten para a taxa de uma reação irreversível de um substrato é parte integral da bioquímica; • Mass ac9on rate law; Lei da ação das massas • A taxa de uma reação é proporcional a probabilidade de colisão dos reagentes; • Ou seja: Modelagem de sistemas dinâmicos • Função linear de duas entradas; • Considere um gene G regulado por dois fatores de transcrição A e B; – proporcional a concentração do substrato; A B G B* A* e B* A* Função linear de duas entradas; • Se A e B regulam G de forma independente, o efeito combinado dos dois em G será adi9vo; Função não linear de duas entradas • Se A e B interagem entre eles, o padrão de expressão de G não é a simples soma do padrão de expressão de A e B. • A regulação de G por A e B não é linear; A B G B* A* e B* Padrão de expressão de G A* B* A* e B* G apresenta uma função linear de A e B A* 7
11/06/15
Função não linear de duas entradas Sistemas não lineares • A e B sozinhos não são capazes de explicar a expressão de G; • Interações não lineares de elementos cis são comuns no desenvolvimento celular; • Variáveis de uma função não linear se afetam entre si e influenciam no resultado; B* A* e B* A* Sistemas não lineares Sistemas não lineares • Considere um cenário no qual células de interesse estão em suspensão (ex: sangue); • Imagine que A e B são iniciadores de duas vias de sinalização diferentes; • Exp1: A9vamos apenas a via de sinalização A e medimos G; • Exp2: A9vamos apenas a via de sinalização B e medimos G; A B • Nos dois casos, G não foi a9vado; • Erroneamente concluímos que as vias A e B não regulam G; • Esta conclusão estaria correta apenas para um sistema linear onde A e B atuam de forma independente; G Sistemas não lineares • Na realidade, A e B podem interagir e regular G de forma não linear; • Esta dificuldade em predizer o comportamento de um sistema, faz com que seja necessário um planejamento experimental cuidadoso; • Se nós sabemos que A e B são a9vadores, um conjunto de experimentos mais informa9vo seria começar com células onde as duas vias estão a9vas e então desa9var uma via de cada vez. • Mas se um for a9vador e o outro repressor ? Sistemas não lineares • O conjunto de componentes determina o estado do sistema; • Sistemas podem ser: • complicados (número de componentes); • complexos (comportamentos dihceis de imaginar); • Algumas sistemas podem ser tão complexos que chegam a ser contra-­‐intui9vos: • comportamento emergente; 8
11/06/15
Comportamento emergente Comportamento emergente • Pontos randomicamente distribuídos se auto-­‐organizam em um circulo ao longo do tempo; • Apenas duas regras simples: – Ficar próximo a seus dois visinhos mais próximos; – Distanciar-­‐se de todos os outros pontos; • Não existe uma definição explicita para forma o circulo; Comportamento emergente • A formação do circulo é uma propriedade emergente da interação dos agentes autônomos; Comportamento emergente • Não existe nada de mágico nestas propriedades; • Neste exemplo as regras: – Maximizam a área total; – Minimizam a distancia entre os vizinhos mais próximos; • Uma propriedade bem conhecida dos círculos; Por que desenvolver modelos ? • Imagine que a transcrição de um gene de interesse seja a9vada por um complexo formado por três proteínas; • Uma destas proteínas atua como scaffold para as outras duas; • Apenas o ul9mo caso é capaz de a9var a transcrição; • Vamos considerar que a concentração das três proteínas é estacionaria (steady state); • U9lizando mass ac9on kine9cs nós podemos calcular a concentração estacionaria para o complexo; Por que desenvolver modelos ? • A não linearidade leva a comportamento surpreendentes; Como esperado a concentração segue uma simples ciné9ca de saturação 9
11/06/15
Por que desenvolver modelos ? • Variando a proporção da concentração final das três proteínas: – Afeta o estado estacionário do complexo; Efeito bifásico • É o mecanismo para a regulação da sinalização na via das MAP kinases; Por que desenvolver modelos ? • No experimento acima não será possível verificar o comportamento bifásico; • A concentração do scaffold nunca será maior que a das outras proteínas; Comportamento surpreendente • Dependência da concentração do complexo em relação concentração total da proteína scaffold. • Surpreendentemente a concentração do complexo cai a medida que a concentração do scaffold aumenta em relação a concentração das proteínas vermelha e verde. Por que desenvolver modelos ? • Vamos supor que as três proteínas são necessárias para a9var a transcrição; • Mas nos não sabemos que uma é um scaffold; • Precisaríamos começar um experimento com quan9dade iguais das três proteínas; • Então ir diminuindo a concentração de cada uma das proteínas e medir o decréscimo na concentração do complexo; Somente modelos mecanis9camente corretos realizam boas predições Modelo simulado
Interpolação – predições dentro das amostras
Extrapolação– predições fora dos dados
10
11/06/15
Por que desenvolver modelos ? • Todos os modelos são aproximações; • Novos experimentos devem ser montados para testar o modelo; • Falhas no modelo podem auxiliar no desenho de novos experimentos; O que é um modelo ? • É tudo aquilo que é u9lizado como um subs9tuto para o sistema que desejamos entender; • U9lizado para visualizar sistemas complexo; • Explorar seus comportamentos em condições de interesse; O que é um modelo ? Editorial
All systems go!
November 2006
Reviews
Linking publication, gene and protein data
Data-driven modelling of signal-transduction networks
Paul Kersey and Rolf Apweiler
Nature Cell Biology advance online publication, 22 October 2006
Kevin A. Janes and Michael B. Yaffe
Nature Reviews Molecular Cell Biology 7, 820-828 (2006)
Modelling data across labs, genomes, space and time
Modelling microtubule patterns
Jason R. Swedlow, Suzanna E. Lewis and Ilya G. Goldberg
Nature Cell Biology advance online publication, 22 October 2006
Eric Karsenti, François Nédélec and Thomas Surrey
Nature Cell Biology advance online publication, 22 October 2006
Collecting and organizing systematic sets of protein data
John G. Albeck, Gavin MacBeath, Forest M. White, Peter K. Sorger, Douglas A.
Lauffenburger and Suzanne Gaudet
Perspective
Nature Reviews Molecular Cell Biology 7, 803-812 (2006)
Essay
Back to the future: education for systems-level biologists
Linking data to models: data regression
Ned Wingreen and David Botstein
Nature Reviews Molecular Cell Biology 7, 829-832 (2006)
Khuloud Jaqaman and Gaudenz Danuser
Nature Reviews Molecular Cell Biology 7, 813-819 (2006)
Tutorial
Physicochemical modelling of cell signalling pathways
Yasushi Sako
Molecular Systems Biology 2, published online: 17 October 2006
Imaging single molecules in living cells for systems biology
Bree B. Aldridge, John M. Burke, Douglas A. Lauffenburger and Peter K. Sorger
Nature Cell Biology advance online publication, 22 October 2006
Princípios da Biologia de Sistemas Redes de regulação gênica Elementos de regulação em redes de transcrição Prof. Renato Vicentini – [email protected]
11
11/06/15
Redes regulatórias da transcrição • A expressão de genes em um genoma é um processo cuidadosamente regulado; • Existem complexas redes regulatórias que controlam quais genes serão expressos em respostas a diversos esGmulos ambientais e a sinais de desenvolvimento; Propriedades básicas • Especificidade: – Especificidade da ligação e associação com macromoléculas; • Co-­‐localização dos componentes; • Passo cri9co, governado pela freqüência de colisão, ou pela ciné9ca de ação das massas; Propriedades básicas • Associação e interação de macromoléculas: – Proteína-­‐proteína; – DNA-­‐proteína; • Não se conhece bem a redes metabólicas envolvidas no processo de GRN; O operon lac em Escherichia coli • Três genes estruturais: – lacA, lacZ, e lacY; – Envolvidos na u9lização de lactose; • Regulado por sinais de lactose e glicose; • Envolvendo duas proteínas regulatórias de ligação ao DNA: – o repressor lac (lactose); – CAP (glicose); • O repressor lac se liga ao DNA somente na ausência de lactose; • A proteína CAP se liga ao DNA somente na ausência de glicose; O operon lac em Escherichia coli • Dependendo da concentração de glicose e lactose no meio, três estados diferentes do sistema podem ser observados: Regulon GAL em levedura • Genes GAL são necessários para a quebra da galactose; • Transcrição induzida pela galactose; – A9vador de ligação ao DNA, Gal4; • Reprimida pela glicose; – Repressor de ligação ao DNA, Mig1; 12
11/06/15
Regulon GAL em levedura • Três estados diferentes do sistema podem ser observados, sendo que Gal80 atua como um inibidor; Redes regulatórias da transcrição Redes regulatórias da transcrição • Notem que apesar de evidencias experimentais serem a base para estes dois úl9mos exemplos, eles são descritos essencialmente em termos de presença e ausência; • Podem muitas vezes ser representados matema9camente; Proteínas que se ligam ao DNA • Mas na realidade estes diferentes estados de ligação ao DNA representam eventos químicos, que são determinados por: – Concentração; – Afinidade de ligação; • Se as equações químicas que descrevem a ligação ao DNA de proteína regulatórias forem conhecidas, estes circuitos regulatórios podem ser descritos estequiometricamente; Proteínas que se ligam ao DNA • Quando todas as proteínas que se ligam ao DNA, assim como seus alvos, são considerados juntos, uma complexa rede regulatória emerge; • Com cascatas de transcrição e loops de auto-­‐regularão; Blocos fundamentas • Bloco fundamental de uma rede regulatória: – Região promotora; • Rede regulatória da transcrição: – Qual TF se liga a qual promotor, e qual é o efeito integrado de todos os TFs na expressão do gene; • Existe um pequeno conjunto de mo9vos comumente encontrados em bactérias; 13
11/06/15
Mo9vos comumente encontrados Hierarquia das redes de transcrição • Existem diversos níveis de abstração para as GRN Reconstruindo redes regulatórias Top-­‐down approach • Dados em larga escala (HT) que sejam capazes de medir simultaneamente um grande numero de variáveis/estados; • Medindo experimentalmente a expressão de um genoma: – WT/knockout de um TF; – Perturbando o experimento (es9mulo, etc); • Iden9ficação de todos os sí9os em promotores u9lizando bioinformá9ca; • Localização experimental da ligação DNA-­‐proteína: – ChIP-­‐chip; ChIP-­‐chip Boyom-­‐up approach • Uso de dados derivados da bioquímica/gené9ca tradicionais; • Focam em poucas variáveis, ou mesmo apenas uma; • Exemplo do operon lac e do regulon GAL; 14
11/06/15
Reconstruindo redes regulatórias Modelagem implícita • Modelos computacionais de GRN não necessariamente precisam ser formulados matema9camente; • Dados de HT de medições de abundancia ou modificações de estados : – RNAs, proteínas, lipídios e metabolitos; • Culpados por associação; Modelagem implícita • Data-­‐driven (top-­‐down) • Associação por função; • Cytoscape; Modelagem implícita • Note a similaridade entre as condições na coluna da direita; • O mesmo para as colunas da esquerda; Modelagem implícita • Exemplos do uso de mapas de interação como modelos predi9vos: –
–
–
–
–
–
Novos membros em grupos da redes; Evidencias que suportam fracas predições; Padrões de interação em larga escala; Conservação evolu9va da topologia da rede; Rede em mul9escalas; Dados mul9dimensionais; Modelagem implícita • As duas colunas parecem ser complementares em termos da expressão gênica; • Padrões como este podem auxiliar a interpretar grandes volumes de dados; • Podem ser testados esta9s9camente; • Servem como ponto de inicio para novos estudos; 15
11/06/15
Modelagem implícita Rede de transcrição em E. coli • U9lizando a analise dos grafos podemos: – Avaliar a evolução no espaço e no tempo de um sistema; • Ajudando assim a encontrar potenciais relações causa-­‐efeito; • Valer-­‐se de ferramentas da teoria de grafos: – Uma área de matemá9ca já bem estabelecida; N=420 Nodes
E=520 Edges
Es=40 self-edges
Rede de transcrição em E. coli • Esta é uma rede complexa; • Vamos tentar definir padrões que sirvam como blocos básicos da construção desta rede (network mo3fs); Blue nodes have self-edges
Network mo9fs • Para definir um padrão esta9s9camente significante, nos comparamos a rede ob9da com um conjunto de redes randomizadas; • Devem possuir as mesmas caracterís9cas da rede real (ex: numero de nos e ligações); • Mas as conexões entre os nos são feitas randomicamente; Network mo9fs Network mo9fs • Padrões que ocorrem na rede real significa9vamente mais vezes do que nas redes randomizadas são chamados network mo9fs; • Ou seja, estão sendo evolu9vamente conservado: • Se um mo9vo não oferece vantagem sele9va, ele é removido e ocorrera em quan9dade similar em uma rede randomizada; – mutações que modificam as interações; • A perda de uma ligação é algo simples (uma mutação em apenas uma base); 16
11/06/15
Network mo9fs Auto-­‐regularão Randomized network
(Erdos – Renyi model)
‘Real’ Network
• Vamos comparar redes reais com redes randômicas; • Vamos começar pela auto regulação (40 casos no exemplo a ser visto para E. coli); • Acelera o tempo de resposta da rede; • Estabiliza a rede; N=10 nodes
E= 14 edges
Es=4 self-edges
N=10 nodes
E= 14 edges
Es=1 self-edge
Regulação simples Auto-­‐regularão • A maioria são auto-­‐reguladores nega9vos A
X
A
Auto-­‐regularão X
Auto-­‐regularão • 40 casos vistos na rede real, contra apenas 1 visto nas redes simuladas; Randomized network
‘Real’ Network
(Erdos – Renyi model)
N=10 nodes
E= 14 edges
Es=4 self-edges
K
• Porque a auto-­‐regularão é um network mo9f ? • Ela apresenta alguma função ú9l ? N=10 nodes
E= 14 edges
Es=1 self-edge
17
11/06/15
Auto-­‐regularão Auto-­‐regularão • Auto-­‐regularão nega9va pode u9lizar um promotor forte para uma produção inicial rápida; • E então u9lizar a auto-­‐repressão para parar a produção no estado estacionário desejado; • Em um gene simplesmente regulado, o estagio estacionário é um balanço entre a produção e a degradação; • Sumarizando: A auto-­‐regularão nega9va nos fornece o melhor de dois mundos: Dinâmica de um gene auto-­‐regulado nega9vamente (linha con9nua) Dinâmica do produto de um gene auto-­‐
regulado nega9vamente (linha con9nua) – Um promotor forte pode gerar uma rápida produção; – A auto-­‐repressão é finamente controlada; • O mesmo promotor em um circuito de regulação simples iria levar a uma super-­‐expressão indesejada; 2
X(t) / K
1.5
1.8
1.6
1.4
X/Xst
1
1.2
1
0.8
0.6
0.5
0.4
0.2
0
0
0
0.2
T1/2 (nar)
0.4
0.6
0.8
T1/2
1
(simple)
1.2
1.4
1.6
1.8
time α t
2
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
Time (α t)
18