UNIVERSIDADE DE SÃO PAULO
INSTITUTO DE BIOCIÊNCIAS
1° CURSO DE NEUROCIÊNCIAS
E COMPORTAMENTO
27 de junho de 2008
www.ib.usp.br/labnec
Sumário Cognição ......................................................................................................................................... 1 Biologia da Cognição: Introdução .................................................................................................... 7 Construção de circuitos e sua modificação pela experiência ............................................................ 10 Integração entre circuitos: o modelo de redes .................................................................................. 11 Biologia da Cognição: Integração Neural ........................................................................................ 15 Percepção envolve ação ..................................................................................................................... 16 Organização e hierarquia no ciclo percepção‐ação ........................................................................... 17 Integrando percepção e ação: o sistema de neurônios espelho ....................................................... 18 Percepção ..................................................................................................................................... 21 Vias perceptuais ................................................................................................................................. 22 Visão ............................................................................................................................................... 22 Audição .......................................................................................................................................... 24 Memórias atentas ao contexto .......................................................................................................... 25 Ilusões e hemisférios cerebrais ...................................................................................................... 26 Sinestesia ............................................................................................................................................ 28 Concluir é um problema ..................................................................................................................... 30 Atenção ......................................................................................................................................... 32 Atenção e percepção .......................................................................................................................... 32 Falha na percepção ............................................................................................................................ 34 Teste de Posner .................................................................................................................................. 34 Efeitos das lesões do sistema nervoso na atenção ............................................................................ 36 Memória ....................................................................................................................................... 38 Aspectos comportamentais e evolutivos ........................................................................................... 38 O sistema nervoso como uma estrutura que suporta os sistemas de memória ............................... 39 Aspectos fisiológicos da memória ...................................................................................................... 40 Plasticidade Neural ............................................................................................................................. 42 i Aquisição e manutenção da memória ............................................................................................... 43 Redes neurais e memória .................................................................................................................. 44 Modularidade e os diferentes processos de memória ...................................................................... 46 Modelos de memória ........................................................................................................................ 47 Memória de longa duração ........................................................................................................... 48 Memória Operacional ................................................................................................................... 49 Sistemas de memórias e seus aspectos evolutivos ........................................................................... 50 Tomada de decisões ...................................................................................................................... 51 Dilemas e Estratégias ......................................................................................................................... 52 Origens ............................................................................................................................................... 53 Interação ............................................................................................................................................ 54 Percepção temporal .......................................................................................................................... 55 Processos inconscientes .................................................................................................................... 55 Atenção .............................................................................................................................................. 56 Memória ............................................................................................................................................ 56 Controle executivo ............................................................................................................................ 56 Estudos clínicos .................................................................................................................................. 57 Livre‐arbítrio e determinismo............................................................................................................ 58 Emoção ......................................................................................................................................... 60 Introdução ......................................................................................................................................... 60 Emoção, cognição e comportamento ................................................................................................ 61 Neurobiologia das emoções .............................................................................................................. 63 Modelos animais................................................................................................................................ 66 Modelos e Cognição ...................................................................................................................... 68 Modelos sobre processos cognitivos ................................................................................................. 68 Exemplo 1 ‐ Memória .................................................................................................................... 69 Exemplo 2 ‐ Atenção ...................................................................................................................... 70 Modelagem Computacional .............................................................................................................. 72 ii Teoria da detecção de sinais .............................................................................................................. 72 Dois fatores são fundamentais para a decisão: a aquisição de informação e o critério ............... 72 Exemplo 1 ‐ Memória e a Teoria de Detecção de Sinais ................................................................ 74 Exemplo 2 ‐ Atenção e a Teoria de Detecção de Sinais ................................................................. 75 Conclusão ........................................................................................................................................... 76 Bibliografia .................................................................................................................................... 77 iii 1º Curso de Neurociências e Comportamento
Cognição
Wataru Sumi
Laboratório de Neurociências e Comportamento
[email protected]
Os animais exibem diferentes tipos de comportamento, uns mais simples, outros
mais complexos. Os mais simples são as respostas reflexas, que são respostas
estereotipadas e fixas a estímulos específicos (Dethier, 1973). A resposta à dor é um
exemplo clássico de como um estímulo ambiental desencadeia uma resposta motora
automaticamente (Fig. 1.A). Existem também respostas bastante elaboradas, que podem
durar alguns minutos, desencadeadas por um único estímulo, como é o caso da resposta de
fuga apresentada por algumas espécies de anêmonas-do-mar. Quando ela é tocada por
uma estrela-do-mar, seus receptores são estimulados e assim, é iniciada uma sequência de
movimentos estereotipados (Fig. 1.B) que a faz se desprender do substrato e iniciar o nado.
Figura 1 - A. Reflexo a dor. B. Comportamento reflexo de fuga na anêmona-do-mar. Retirado de: 1. A
http://scienceblogs.com e 1. B Dethier, 1973.
Como exemplo de comportamento altamente complexo, podemos citar a habilidade
dos corvos da Nova Caledônia para construir ferramentas, que são hastes manufaturadas a
partir das folhas das plantas locais e utilizadas para retirar insetos de dentro das cascas das
árvores ou troncos apodrecidos. Essas ferramentas possuem ganchos, uma característica
observada apenas nesses animais e em humanos (Hunt ,1996). Além disso, essas
ferramentas são altamente uniformes, porém, variando de acordo com as diferentes regiões
onde vivem os corvos, sugerindo que o conhecimento para produzir essas ferramentas seja
transmitido de um indivíduo para outro (Hunt e Gray, 2004).
Um exemplo mais próximo de nós humanos é o uso de diferentes ferramentas por
chimpanzés: eles são capazes de utilizar gravetos para “pescar” cupins ou formigas (Fig. 2);
pedras fazendo papel de martelo e bigorna para quebrar nozes ou, ainda; galhos como
1 www.ib.usp.br/labnec
lanças para espetar presas entocadas em buracos além do alcance de seus braços. Essas
habilidades são aprendidas por observação e transmitidas de geração a geração (i.e.
culturalmente) (Wilson, 2000).
Figura 2 - Uso de ferramentas por chimpanzés. Um graveto é usado para “pescar” formigas. Retirado de Naish.
Dentre os exemplos de comportamento apresentados até agora, todos concordariam
que, no primeiro caso (resposta reflexa), o comportamento não envolveria processos
cognitivos e, no segundo caso (uso de ferramentas), se trataria do mais genuíno exemplo de
cognição observado na natureza. Porém, entre esses dois tipos bastante distintos de
comportamentos, o reflexo e a capacidade de produzir e utilizar ferramentas, existe um
grande repertório comportamental regido tanto pelo instinto como pelo aprendizado.
Os instintos são padrões de comportamento estereotipados que aparecem em sua
forma funcional desde a primeira vez em que são executados, mesmo que o animal não
tenha experiência prévia com o estímulo eliciador do comportamento. A rede neural
responsável pela detecção do estímulo e ativação do programa motor é denominada
mecanismo de liberação inato (Alcock, 2005).
Esses mecanismos inatos muitas vezes são modulados a partir das experiências
vividas pelos animais, ou seja, o aprendizado pode modificar o comportamento inato. Os
esquilos, por exemplo, que comem diferentes tipos de sementes e nozes, reconhecem-nas e
abrem-nas instintivamente mas, dada a variedade de formatos de sementes, é necessária
uma técnica específica para abrir cada uma delas. A habilidade de abrir um determinado tipo
de noz é adquirida por tentativa e erro até que chegam à perfeição (Tinbergen, 1971).
Vimos que existe um continuum de complexidade do comportamento. Como já
mencionado anteriormente, nem todos eles são tratados como cognição. A partir de que
grau de complexidade podemos dizer que um determinado comportamento é cognitivo?
Essa resposta varia enormemente entre diferentes autores. Uma definição mais abrangente
2 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
entende a cognição como sendo os mecanismos pelos quais os animais captam a
informação do ambiente, a retêm e a usam para ajustar o comportamento às condições
locais ou, simplesmente, como processamento da informação. Em uma definição mais
estrita, cognição é tratada como o conjunto de processos que produzem o comportamento
intencional (Heyes e Huber, 2000), ou manipulação do conhecimento declarativo (saber
que), não sendo considerada cognição o conhecimento de procedimento (saber como)
(McFarland, 1991).
A definição adotada pela neurociência cognitiva é a mais ampla, ou seja, considera a
cognição como o processamento da informação. Se pensarmos que, por exemplo, a
memória pode ser dividida em explicita e implícita (Fig. 3), sendo que a memória explícita
seria responsável pelo comportamento intencional, a adoção da definição mais restrita de
cognição implicaria em estudar apenas parte desses processos.
Figura 3 - Existem diferentes tipos de memória. A memória de longa duração pode ser dividida em: memória
declarativa e memória não-declarativa (retirado de Gazzaniga e col., 2006).
Como vimos até agora, a nossa definição de cognição não se restringe apenas a
processos mentais mais elevados, aqueles que nos permitem filosofar, calcular etc.. Durante
o dia, realizamos inúmeras atividades nas quais utilizamos a cognição. Conversamos com
um amigo, lemos um jornal, vamos até a padaria da esquina, preparamos uma refeição,
assistimos à televisão, andamos de bicicleta etc.. A maioria das nossas ações envolve
cognição, ou seja, processos como percepção, memória, atenção, tomada de decisão e
emoção.
Então, qual será o papel dos processos cognitivos em nossas atividades diárias?
Será que todos eles são utilizados? Veremos o “passo a passo” da recepção da informação
3 www.ib.usp.br/labnec
e subsequente processamento. Antes de qualquer coisa, para interagir com o ambiente,
precisamos de uma interface que faça a ligação do mundo exterior com o mundo interior,
representada pelos diferentes receptores sensoriais (foto-receptor, quimio-receptor etc.),
que transformam os estímulos do ambiente em potenciais elétricos transmitidos pelos
neurônios.
Após o recebimento das informações do ambiente, elas são processadas pelo
sistema perceptual. Diferentes regiões do cérebro são responsáveis por processar as
diferentes características dos objetos. Por exemplo, quando vemos um pintinho amarelo
andando, essa informação é processada por três subsistemas distintos, responsáveis por
forma, cor e movimento. Apesar dessas características dos objetos serem separadas
durante o processamento da informação, elas são percebidas como uma unidade e não
apenas como forma, cor e movimento separadamente (Gazzaniga e col.,2002).
A qualidade da informação detectada do ambiente não recebe modulação dos
receptores sensoriais, isso depende basicamente das características do estímulo. A
quantidade de informações recebidas por nossos sistemas sensoriais é enorme. Para
entender essa grandeza, imagine perceber todos os detalhes existentes de uma paisagem
em alguns poucos segundos; isso é uma tarefa impossível. Nosso sistema nervoso é
simplesmente incapaz de processar todas as informações ambientais simultaneamente. Isso
fica evidente também quando tentamos realizar simultaneamente duas atividades distintas,
por exemplo, conversar e ler um livro.
O sistema nervoso, por meio da atenção, seleciona certos estímulos para serem
adequadamente
processados.
Os
objetos
ou
eventos
escolhidos
para
posterior
processamento variam de acordo com a sua relevância. Por exemplo, se queremos ler um
livro, direcionamos voluntariamente a atenção visual para as letras e palavras. Há também,
certos estímulos que atraem a atenção automaticamente. Esses estímulos se caracterizam
por ser mais salientes do que outros, como por exemplo, a sirene e as luzes intermitentes
das ambulâncias.
Vamos supor que estamos engajados em uma conversa. A atenção seleciona as
informações que julgamos relevantes e essas informações são processadas pelo sistema
sensorial auditivo e posteriormente enviadas para áreas responsáveis pela linguagem. É
importante ressaltar que o processamento da linguagem não envolve apenas o sentido
auditivo, mas também o visual e o somático. Quando lemos um texto utilizando a visão
(mais comum) ou o tato (leitura em braile), as informações desses diferentes sentidos são
igualmente processadas nas áreas da linguagem.
Para manter uma conversa, direcionar a atenção ou perceber o mundo como nós
percebemos, é necessária, além dos processos já mencionados, também a memória. A
memória nos permite lembrar a tabuada, o caminho para a faculdade, o rosto de nossas
4 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
mães, nossos nomes, o significado das palavras etc. Para mantermos uma conversa
precisamos da memória, caso contrário não nos lembraríamos da última palavra ouvida ou
falada.
A atenção sustentada, que é o comportamento de manter a atenção focada em um
objeto ou situação por algum tempo, é possível graças à memória. Se mantemos a atenção
voluntariamente direcionada para algo, é porque provavelmente isso é relevante para nós.
Ou seja, as informações da memória influenciam o controle do direcionamento da atenção.
Direcionar a atenção voluntariamente ou realizar qualquer outra atividade, envolve a
tomada de decisão. A todo instante devemos decidir: continuamos a assistir TV ou
começamos a estudar para a prova? Comer mais uma fatia de pão no café da manhã? Usar
a camiseta vermelha ou a azul? Viajar para a praia ou para a montanha no feriado? A
maioria dos nossos comportamentos envolve algum tipo de decisão. A decisão não é
apenas uma simples escolha entre diferentes opções, mas uma escolha dependente de
diversos fatores. Um deles é a memória: quando sabemos, por experiências passadas, que
uma determinada opção pode nos trazer mais benefícios, é natural que essa escolha seja
preferida em detrimento das outras.
Outro fator importante na tomada de decisão é a emoção. Se tivermos medo de algo,
certamente nos comportaremos de modo a evitá-lo. Em um experimento clássico avaliou-se
o efeito da emoção no comportamento de risco. Eram apresentados a voluntários dois
montes de cartas. Em um deles (A), ganhava-se uma recompensa de $50, correndo-se o
risco de perder até $100. Por outro lado, no outro monte (B), podia-se ganhar $100, mas
podia-se perder até $1200, ou seja, o risco de perder era muito maior comparado ao ganho.
Sabendo dos riscos, os voluntários poderiam escolher livremente entre os dois montes.
Voluntários controles evitavam as cartas do monte B e a simples cogitação de escolher a
pilha mais arriscada desencadeava uma clara resposta emocional involuntária. Por outro
lado, pacientes com lesões específicas no córtex cerebral, relacionadas à emoção,
escolhiam sempre o monte mais arriscado e não apresentavam resposta emocional.
Esses são apenas alguns exemplos de como os diferentes processos cognitivos
atuam para produzir o nosso comportamento. Cada um desses processos pode ser mais ou
menos utilizado de acordo com a situação, a atividade realizada. Isso fica bastante claro
quando comparamos dois tipos de atividades como, por exemplo, fazer uma prova e assistir
à TV. Em ambos os casos utilizamos a memória, mas esse processo cognitivo é muito mais
ativo na primeira situação.
Estudando o funcionamento de cada um desses processos e como eles se interrelacionam, a neurociência cognitiva tenta entender como o sistema nervoso produz o
comportamento. Nos capítulos seguintes estudaremos como os diferentes processos
5 www.ib.usp.br/labnec
cognitivos atuam, além, é claro, do funcionamento do sistema nervoso propriamente dito,
suas unidades funcionais e os mecanismos de integração e processamento da informação.
6 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
Biologia da Cognição: Introdução
Renata Pereira Lima
Laboratório Neurociência e Comportamento
[email protected]
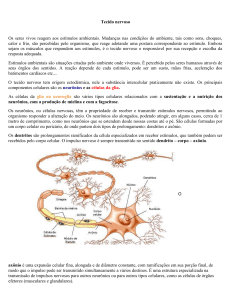
No sistema nervoso, neurônios nunca funcionam isolados; eles estão organizados
em circuitos que processam tipos específicos de informações. O sistema nervoso parece
organizado em grupos de circuitos, i.e., módulos, cujas funções servem a um propósito
comportamental específico. Desta maneira, sistemas sensoriais como a visão ou audição
adquirem e processam informações a partir do ambiente, o sistema motor permite que o
organismo responda a tais informações através da geração de ações. Há, entretanto, um
grande número de células e circuitos que estão entre estas mais ou menos bem definidas
aferências e eferências. Eles são coletivamente referidos como sistemas de associação e
são responsáveis pelas mais complexas funções.
Além destas amplas distinções, os neurocientistas têm convencionalmente dividido o
sistema nervoso dos vertebrados, sob o ponto de vista anatômico, em componentes centrais
e periféricos (Fig. 1). O sistema nervoso central (SNC) compreende o encéfalo e a medula
espinal. O sistema nervoso periférico (SNP) inclui fibras de neurônios que conectam os
receptores sensoriais na superfície do corpo ao SNC e a porção motora, que consiste em
axônios de nervos motores que conectam o encéfalo e a medula espinal aos músculos
esquelético, viscerais, cardíaco e glândulas.
Figura 1. Arranjo anatômico do sistema nervoso em
humanos. Em azul o sistema nervoso central (SNC) e em
amarelo, o sistema nervoso periférico (SNP). Retirado de
Bear, 1996.
7 www.ib.usp.br/labnec
Embora o arranjo dos circuitos que compõem estes sistemas varie grandemente de
acordo com suas funções, algumas características são comuns entre eles. As conexões
sinápticas que definem um circuito são tipicamente realizadas numa densa malha de
dendritos e terminais axonais. A direção do fluxo de informação em um circuito particular é
essencial para se entender sua função. Células nervosas que transmitem informações em
direção ao sistema nervoso central são chamadas de neurônios aferentes; já as que
transmitem informações para fora do encéfalo e da medula espinal (ou para fora do circuito
em questão), são chamadas de neurônios eferentes. Células nervosas que participam
somente no aspecto local do circuito são chamadas de interneurônios. Estas três classes –
neurônios aferentes, neurônios eferentes e os interneurônios – são os constituintes básicos
de todos os circuitos neurais.
De modo geral, podemos classificar os circuitos como:
Convergentes: aqueles nos quais um grupo de neurônios recebe uma
aferência (entrada) de um neurônio pré-sináptico e o circuito tende a se tornar concentrado.
Para demonstrar este tipo de circuito, imagine que tenhamos os neurônios A, B e C e que
cada um deles possua uma entrada diferente. Estes neurônios se projetam para um
neurônio D e este se projeta para outro neurônio E, realizando uma eferência (saída).
Circuitos convergentes são responsáveis, por exemplo, pela interpretação dos estímulos
sensoriais (Fig. 2, à esquerda).
Divergentes: são os circuitos que funcionam de maneira oposta aos circuitos
convergentes. Em vez de concentrar as aferências, estas se projetam separadamente para
diferentes neurônios. No caso do circuito divergente, o neurônio A possui uma aferência e
se projeta para os neurônios B, C e D. A característica básica de um circuito divergente é o
fato de que um único neurônio iniciará respostas de maneira crescente em outros neurônios.
Tais circuitos são encontrados nos sistema motores e sensoriais (Fig. 2, centro).
Reverberantes: o sinal de aferência é transmitido ao longo de uma série de
neurônios e cada um destes fará sinapses com neurônios de uma porção da via
previamente percorrida. O impulso reverbera sendo enviado ao longo do circuito
continuamente até que um neurônio seja inibido. Então, uma aferência no neurônio A se
projeta para o neurônio B, que se projeta para o neurônio C e então para o D e este se
projeta de volta para o neurônio A (ou para o B) e o ciclo se repete até que um neurônio
(que pode ser tanto A, quanto B, C ou D) seja inibido. Circuitos reverberantes estão
envolvidos no ciclo de sono-vigília, atividades motoras, memórias de longa duração, etc (Fig.
2, à direita).
8 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
Figura 2 - Esquema representativo dos modelos de circuitos. À esquerda, o modelo de circuitos convergentes,
no centro o modelo divergente e o reverberante à direita.
Além disto, circuitos podem funcionar paralela ou serialmente. No funcionamento
paralelo, sinais aferentes são processados em vias distintas e as informações são
analisadas de maneira analítica concomitantemente no tempo. Por exemplo, o sistema
visual funciona em vias paralelas que processam a informação neural de forma simultânea e
integrada. Sinais representando cores, movimento, forma e localização, por exemplo, são
processados simultaneamente em diferentes regiões do encéfalo. Atividades concomitantes
(e sincronizadas) nas vias visuais dorsal e ventral (que são anatomicamente distintas) são
responsáveis pela percepção unitária da imagem. No funcionamento serial, os resultados
dos processamentos de um circuito são necessários para que o próximo circuito possa
contribuir para o processamento total. Isto é, um neurônio estimula outro neurônio, que por
sua vez estimula outro neurônio e assim por diante. Um exemplo clássico de processamento
serial é o arco reflexo, em que há produz uma reação involuntária rápida, na maioria das
vezes inconsciente, que protege o organismo. Tal reação é originada a partir de um estímulo
externo que gera uma resposta antes mesmo do indivíduo tomar conhecimento da
existência do estímulo periférico e, conseqüentemente, antes deste poder comandá-la
voluntariamente. Muitos reflexos motores são controlados por neurônios localizados na
substância cinzenta da medula espinhal e do tronco encefálico (bulbo, ponte e
mesencéfalo), independentemente da vontade, como por exemplo:
• a retirada imediata da mão de uma panela muito quente;
• extensão da perna após a percussão e estiramento do tendão patelar;
• fechamento da pupila com o aumento da intensidade luminosa;
• aumento da secreção gástrica com a chegada do alimento no estômago.
Desta maneira, o ato reflexo é um mecanismo que gera uma reposta involuntária do
organismo a um determinado estímulo (dor, estiramento, aumento da intensidade luminosa,
variações da pressão arterial etc). Ocorrendo um estímulo, a fibra sensitiva de um nervo
aferente (ou sensitivo) transmite-o até a medula espinhal passando pela raiz posterior, ou
ao tronco encefálico, por meio de um nervo craniano. Na medula ou no tronco encefálico o
neurônio aferente comunica-se com o eferente diretamente ou por meio de interneurônios
associativos, gerando, no neurônio motor, a atividade que leva à ação. Os axônios
eferentes que levam essa ordem da medula (pela raiz anterior) ou do tronco encefálico (por
9 www.ib.usp.br/labnec
um nervo craniano) constituem as fibras eferentes motoras ou vegetativas que levam a
informação ao órgão efetor (músculo estriado esquelético, glândula, músculo liso ou
músculo cardíaco) que, por sua vez, executará a resposta ao estímulo inicial.
É importante ressaltar que o processamento serial é a maneira mais simples por
meio da qual um circuito pode funcionar. Este tipo de processamento está envolvido nas
respostas mais simples e estereotipadas. Durante o processamento de funções mais
complexas, de modo geral, os circuitos envolvidos, além de processar informações de modo
serial, funcionam concomitantemente em paralelo com outros circuitos de maneira
sincronizada.
Construção de circuitos e sua modificação pela experiência
A construção da circuitaria do sistema nervoso envolve processos ontogenéticos
associados à interação do sistema com o ambiente. Assim, fatores químicos liberados por
determinados neurônios em diferentes estágios do desenvolvimento ontogenético atraem
projeções de outros neurônios intrinsecamente; paralelamente, essas projeções e conexões
entre
neurônios
podem
originar-se
também
em associação com
a
estimulação
proporcionada pelo ambiente e/ou pela atividade de certos conjuntos de neurônios. Assim,
os padrões macroscópicos básicos das conexões no sistema nervoso estabelecidas
filogeneticamente podem ser microscopicamente alterados por padrões de atividade
neuronal (isto é, experiência), modificando a circuitaria sináptica do encéfalo. A atividade
neuronal gerada em decorrência de interações com o ambiente pré e pós-natal influencia a
estrutura e a função do sistema nervoso, além da construção de sua circuitaria.
A história de interação de um indivíduo com o ambiente, i.e., sua experiência
acumulada, molda os circuitos neurais, determinando seu comportamento. Em alguns
casos, as experiências funcionam primariamente como gatilhos que ativam alguns
comportamentos inatos. Mais freqüentemente, entretanto, experiências desenvolvidas em
períodos específicos no início da vida (referidos como períodos críticos) determinam um
repertório comportamental no indivíduo adulto. Estes períodos críticos influenciam
comportamentos diversos incluindo laços maternais, preferências sexuais e aquisição de
linguagem, entre outros.
Embora seja possível identificar conseqüências comportamentais de determinados
estímulos que foram apresentados em períodos críticos para determinadas funções, suas
bases biológicas ainda não estão completamente esclarecidas. Talvez o exemplo mais bem
investigado relacione-se ao período crítico no estabelecimento da visão. Alguns estudos
mostraram que a experiência é traduzida em padrões distintos de atividade neuronal que
influenciam a função e a conectividade dos neurônios relevantes. No sistema visual (e em
outros sistemas também) a competição entre aferências com diferentes padrões de
10 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
atividade é um determinante importante na consolidação dos padrões de conectividade. Em
um axônio aferente, padrões de atividade correlatos tendem a estabilizar as conexões.
Quando padrões normais de atividade são rompidos (experimentalmente, em animais, ou
patologicamente, em humanos) durante um período critico na infância, a conectividade no
córtex visual é alterada, assim como a função visual. Se não é feita a manutenção destes
padrões até o final do período critico, estas alterações estruturais da circuitaria nervosa
dificilmente se restabelecem posteriormente.
A conectividade nervosa estabelecida ao longo do desenvolvimento normal
possibilita ao encéfalo armazenar vasta quantidade de informações que refletem a
experiência
específica
daquele
individuo.
Como
esperado,
a
construção
dessa
conectividade que tanto influencia o desenvolvimento do sistema nervoso gera alterações
maiores nos estágios iniciais de desenvolvimento. Assim, em um animal adulto, o sistema
nervoso se torna gradativamente mais refratário a lições da experiência e os mecanismos
celulares que medeiam as alterações da conectividade neuronal se tornam menos plásticos.
Integração entre circuitos: o modelo de redes
O conceito de que no córtex cerebral há domínios discretos dedicados mais ou
menos exclusivamente a algumas funções cognitivas, tais como discriminação visual,
linguagem, atenção espacial, reconhecimento de face, retenção de memória, memória
operacional, etc., tem sido questionado devido à falta de evidências conclusivas que o
apóiem. Em seu lugar, modelos de redes neurais têm sido apresentados como uma
alternativa mais coerente com as evidências disponíveis sobre seu funcionamento.
Em 1949, Donald Hebb hipotetizou uma forma de plasticidade sináptica
proporcionada por uma continuidade temporal das atividades pré e pós-sinápticas. Além de
acreditar que as conexões sinápticas eram as bases das associações mentais, ele foi além
do simples conexionismo dos behavioristas. Primeiro, ele argumentou que uma associação
não poderia ser localizada numa simples sinapse. Ao contrário, os neurônios estariam
agrupados em “assembléias de células” e esta associação era distribuída nas suas
conexões sinápticas. Segundo, Hebb rejeitou a noção de que a relação estímulo-reposta
poderia ser explicada somente por um simples arco reflexo conectando neurônios sensoriais
a neurônios motores. Assim, era necessário postular “um mecanismo central que explicasse
o atraso existente entre o estímulo e a resposta que é tão característico do pensamento”
(Hebb, 1949). Seguindo as idéias do neurofisiologista Lorente de Nó, Hebb acreditava que a
estimulação sensorial poderia iniciar padrões de atividade neural que eram mantidas
centralmente pela circulação em loops de feedbacks sinápticos. Tal “atividade reverberante”
torna estes padrões possíveis para as respostas que são subseqüentes aos estímulos
posteriores ao atraso. Em resumo, Hebb hipotetizou um “mecanismo com fundamentos
11 www.ib.usp.br/labnec
duplos” da memória. A atividade neural reverberante era o fundamento da memória de curta
duração, enquanto as conexões sinápticas eram o fundamento da memória de longa
duração. Desta maneira, Hebb propôs que:
“A persistência ou repetição de uma atividade reverberante tende a induzir mudanças
celulares permanentes que promovem estabilidade no sistema” (Hebb, 1949).
Esta proposição pode ser precisamente colocada da seguinte forma: quando um
axônio da célula A repetidamente ou persistentemente dispara, alguns processos de
crescimento ou mudanças metabólicas acontecem em uma ou em ambas as células (A ou
B) de tal modo que a eficiência de A, uma das células que estão agindo sob B, é
aumentada.
Além disto, Hebb hipotetiza uma função específica para esta “sinapse hebbiana”: a
conversão da memória de curta duração em memória de longa duração pela estabilização
de padrões de atividade reverberante. Uma vez que este padrão de atividade foi
armazenado nas conexões sinápticas, ele pode ser resgatado repetidamente a partir da
excitação de neurônios sensoriais ou a partir de outros padrões de atividade reverberante.
A hipótese de Hebb foi verificada décadas depois com a descoberta da potenciação
de longa duração, LTP (do inglês, long-term potentiation) (Fig. 3). A LTP é um estreitamento
da conexão entre dois neurônios que resulta de uma estimulação simultânea de ambos e
pode ser induzida experimentalmente aplicando-se uma seqüência de pequenos estímulos
de alta freqüência na célula nervosa. Este estreitamento pode durar de minutos a horas (in
vitro) ou de horas a dias ou meses (in vivo).
Pela eficiência aumentada da transmissão sináptica, a LTP aumenta a habilidade de
dois neurônios, um pré-sináptico e outro pós-sináptico, de comunicarem-se através da
sinapse. O mecanismo preciso para este aumento da transmissão ainda não é bem
estabelecido, em partes porque a LTP é controlada por múltiplos mecanismos que variam de
acordo com a região em que acontecem, a idade do animal em questão e espécie.
Entretanto, nas formas de LTP mais compreendidas, a melhora desta comunicação é
predominantemente feita através do aumento da sensibilidade das células pós-sinápticas
em receber sinais das células pré-sinápticas. Estes sinais, na forma de moléculas de
neurotransmissores, são recebidos por receptores presentes na superfície da célula póssináptica. Este aumento de sensibilidade é devido não somente ao aumento da atividade
dos receptores já existentes na superfície, mas também por um aumento do número destes
receptores.
Interessantemente, a LTP compartilha muitas características com a memória de
longa duração, o que faz dela uma candidata muito atrativa como um mecanismo celular do
aprendizado. Por exemplo, a LTP e a memória de longa duração dependem da síntese de
novas proteínas, possuem propriedades associativas e podem durar potencialmente vários
12 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
meses. A LTP também pode responder por vários tipos de aprendizado, desde o
relativamente simples condicionamento clássico presente em todos os animais, até
respostas mais complexas, como a cognição observada em humanos.
De acordo com essa concepção, a alteração estrutural leva ao armazenamento da
informação podendo explicar o fenômeno da memória. Este modelo postula que todas as
representações cognitivas consistem em redes de neurônios cuja atividade foi associada
pela experiência (estímulos repetidos). Nesse contexto, pode-se assumir que memórias
filogenéticas correspondem a redes que se consolidaram ao longo das gerações e não
necessitam de experiência individual para serem funcionais, embora possam ser
aprimoradas pela experiência individual.
Figura 3 - Modelo representativo do funcionamento da Potenciação de Longa-Duração (LTP). Os receptores
NMDA (vermelho) constituem a maquinaria molecular da aprendizagem. O neurotransmissor é libertado durante
atividade basal e durante a indução de LTP (topo, à esquerda). A expressão de LTP pode dever-se à presença
de mais receptores AMPA (receptores em amarelo, à esquerda, abaixo) ou à presença de receptores AMPA
mais eficientes (à direita, abaixo). Disponível em www.braincampaign.org - 09/06/2009.
Se considerarmos que um neurônio tipicamente recebe informações de cerca de 104
neurônios e, por sua vez, projeta-se para outros 104 neurônios e, que o encéfalo humano
13 www.ib.usp.br/labnec
contém pelo menos 1011 neurônios, isto significa dizer que pelo menos 1019 conexões
sinápticas são formadas no cérebro. Entretanto, a complexidade de seu funcionamento é
evidentemente maior, em particular quando se considera os arranjos seqüenciais pelos
quais uma informação pode viajar ao longo de seqüências de neurônios. Quanto mais
freqüentes as exposições a estímulos relevantes, mais fortes tornam-se essas conexões.
Como conseqüência, a informação tende a ser arquivada de maneira relacional. Isso
permite entender porque a recordação envolve, usualmente, categorias. Por exemplo, ao
pedirmos para uma pessoa listar todos os animais de que se recorda, não raro a lista
conterá animais agrupados por categorias de similaridade, ou seja, quadrúpedes, aves,
animais aquáticos, invertebrados etc. O mesmo ocorre em relação a alimentos; a
recordação também será categórica (frutas, verduras, legumes, carnes etc.). Isso ocorre
porque o aumento de atividade eletrofisiológica em determinados circuitos neurais (que
levam à recordação de uma dada informação) tende a estimular a atividade em circuitos
relacionados. Assim, quando aprendemos que determinado estímulo se refere a um
determinado conceito, estamos na verdade fazendo associações com conceitos que já
conhecemos (associando nós de uma rede com outros). Então, quando visualizamos a
imagem de uma maçã caindo, integramos todas as informações disponíveis (cor, forma,
contexto, movimento) com os circuitos já consolidados previamente e que em algum
momento foram associados ao conceito “maçã”. O mesmo vale para uma outra modalidade
de estímulo, ou seja, um som específico que atribuímos como característico de um
determinado animal, o cheiro de uma comida que está intimamente ligado com o seu sabor
etc.
14 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
Biologia da Cognição: Integração Neural
Renata Pereira Lima
Laboratório Neurociência e Comportamento
[email protected]
Todas as formas de comportamento adaptativo requerem o processamento de um
fluxo de informação sensorial e sua transdução em uma série de ações direcionadas a um
objetivo. Desde a mais primitiva espécie animal, todo o processo é regulado por feedbacks
externos (ambiente) e internos (Fig.1). Esse padrão de funcionamento torna o organismo
apto a forragear, fugir de predadores, lutar e reproduzir-se.
Figura 1 - Uma das finalidades da percepção é permitir uma interação com o ambiente. Interações podem incluir
andar de um lugar para outro, pegar um objeto, conversar com uma pessoa ou dirigir um carro. De modo circular,
tais ações afetam diretamente nossa percepção do mundo. Esta interdependência entre ação e percepção é
ilustrada pelo “Ciclo Percepção-Ação” da figura acima. A visão que temos na integração sensoriomotora é que
em vários aspectos do comportamento, ações motoras e processos sensoriais estão conectados
inseparavelmente e, desta forma, precisam ser estudados juntos. O sistema nervoso evoluiu, sobretudo nos mamíferos, de tal forma que uma grande
complexidade estrutural e funcional foi alcançada não tanto pelas vias aferentes,
responsáveis por canalizar as informações sensoriais, ou pelas vias eferentes, responsáveis
por emitir as respostas motoras, mas por circuitos neurais que intermedeiam essas vias de
entrada e saída. Os complexos circuitos neurais que se localizam entre as vias sensoriais e
motoras são os principais responsáveis pela riqueza, flexibilidade e plasticidade de
comportamentos observados. Isso se manifesta na enorme diversidade de estímulos que
podem ser reconhecidos pelos sistemas sensoriais, na multiplicidade de graus de liberdade
com que ações são organizadas pelos sistemas motores e, sobretudo, pela rica e plástica
relação que se estabelece entre esses dois conjuntos.
A progressiva elaboração dos circuitos neurais pode ser entendida como uma
conseqüência da seleção de ações mais vantajosas (organizadas por circuitos “pré15 www.ib.usp.br/labnec
motores”) em resposta à identificação seletiva de estímulos específicos (realizada por
circuitos “perceptivos”), provavelmente pressionada por fatores ambientais. Podemos supor
então que, ao tornar-se cada vez mais complexo, o funcionamento dos circuitos neurais que
organizam a integração sensório-motora expressa aquilo que chamamos de “percepção”,
“atenção”, “aprendizado”, “memória”, “ação” e, por fim, “consciência”. Esses rótulos estão
longe, em sua maioria, de uma definição completa e consensual. Eles são, mais
provavelmente, o resultado das limitações que ainda temos em compreender a essência do
funcionamento do sistema nervoso, não se constituindo em entidades separadas e
independentes da função neural.
Desta forma, se considerarmos que a percepção do mundo, onde “perceber” algo,
derivado do latim, significa “apoderar-se” dele, logo veremos que não há percepção sem que
alguma forma de atenção esteja em jogo. E é só por meio da percepção atenta que temos
de um estímulo que sentimos, de um evento que presenciamos ou de uma resposta que
emitimos, que poderemos mais tarde nos lembrar desse objeto, desse evento ou dessa
resposta, resgatando uma memória arquivada por meio de um processo de aprendizado. E,
de forma um tanto óbvia, todo trabalho investido em se “apoderar” do mundo, “arquivá-lo” e
“resgatá-lo”, seria inútil e sem sentido se não usássemos essa informação na organização e
emissão de uma ação sobre o mundo, com ele interagindo de forma contínua e coerente,
permitindo nossa permanência nesse mesmo mundo, apesar de seus constantes desafios.
Percepção envolve ação
Perceber algo geralmente requer alguma ação por parte de quem esta percebendo.
Freqüentemente temos que olhar (direcionar os olhos) para ver, fazendo uma varredura
visual do ambiente até que o objeto de desejo seja encontrado. Da mesma forma, para um
som ser audível, temos que direcionar nossos ouvidos em sua direção. Quando tocamos um
objeto, ele é mais facilmente identificado se for explorado pelos nossos dedos.
Todos estes exemplos demonstram que a percepção é um processo ativo que
funciona para direcionar e otimizar o comportamento através do seu refinamento. Além
disso, uma vez que um objeto tenha sido percebido, podemos decidir se iremos nos
aproximar ou nos afastar. Ao ouvir um barulho podemos responder a ele ou ficar quieto. Ao
identificar um objeto pelo toque podemos descartá-lo ou mantê-lo conosco. Em cada um
destes casos nosso comportamento depende do que é percebido.
A orientação da percepção por meio de uma ação induz uma distinção interessante
entre os vários sentidos que tem a ver com a proximidade do observador em relação ao
objeto percebido. Tocar e saborear algo requer um contato direto entre o observador e a
fonte de estimulação. Cheirar também é um certo contato com a fonte de estímulação;
substâncias químicas voláteis são diluídas conforme a distância da fonte aumenta; desta
16 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
forma, o cheirar funciona mais eficientemente para substâncias que estão próximas. Em
contraste, ver e ouvir,não dependem tanto deste contato. Os olhos e os ouvidos podem
capturar a informação originária de fontes remotas, neste sentido eles funcionam como um
radar. Eles permitem que o indivíduo faça contato perceptual com um objeto que não está
próximo, eles estendem a percepção para um mundo além dos limites dos dedos e do nariz.
Estes dois sentidos substituem o deslocamento até a fonte de estímulo, permitindo que o
indivíduo explore a vizinhança.
Organização e hierarquia no ciclo percepção-ação
Em todo o sistema nervoso central, o processamento de seqüências de ações
guiadas sensorialmente segue um fluxo a partir de estruturas geralmente posteriores
(sensórias), em direção a estruturas anteriores (motoras), com feedbacks em todos os
níveis. Assim, no nível cortical, a informação flui de maneira circular ao longo de uma série
de áreas hierarquicamente organizadas e entre conexões que constituem o ciclo percepçãoação (Fig. 2).
Ações automáticas e/ou muito freqüentes em resposta a estímulos sensoriais são
integradas em níveis mais inferiores do ciclo, nas áreas sensoriais da hierarquia (perceptiva)
e em áreas motoras da hierarquia (executiva). Comportamentos mais complexos, guiados
por estímulos também mais complexos e distantes no tempo, requerem uma integração em
níveis corticais mais superiores de ambas as hierarquias (perceptuais e executivas),
basicamente áreas superiores de associação sensorial e córtex frontal anterior.
Figura 2 - O substrato cortical do ciclo percepçãoação. Em azul está representado o lado da
percepção no ciclo e em vermelho o lado da ação.
Os retângulos vazios representam áreas
intermediárias ou subáreas do córtex. As setas
representam vias anatomicamente identificadas
em macacos e ressaltam a conectividade
recíproca entre os córtices posterior e anterior.
Retirado de Fuster, 2006. Para garantir as interações entre as duas hierarquias corticais, longas fibras corticocorticais conectam recíproca e topologicamente as áreas da hierarquia perceptual com as
áreas equivalentes executivas. Assim, áreas pré-motoras se conectam com áreas sensoriais
associativas relativamente inferiores (áreas inferiores de ambas as hierarquias), enquanto
17 www.ib.usp.br/labnec
áreas frontais anteriores se conectam com áreas associativas superiores do córtex posterior
(áreas superiores). Do mesmo modo, há evidências anatômicas de conexões ordenadas
descendentes do córtex frontal anterior ao córtex pré-motor e deste para o córtex motor. Em
cada estágio deste processo em cascata na hierarquia executiva, a próxima ação de uma
seqüência é determinada por dois tipos de influências: 1) o processamento dos aspectos
globais da seqüência nas áreas frontais superiores e 2) os sinais sensoriais que estão
ocorrendo naquele momento. A ativação progressiva de áreas frontais inferiores que
processam a ação é cumulativa. Da mesma forma, as entradas sensoriais associativas do
córtex posterior são progressivamente mais concretas e mais dependentes de um contexto
espacial e temporal imediato. Sinais que necessitam ser processados em um contexto
temporal mais amplo (episódico) requerem ações que dependem de uma integração
temporal em graus mais elevados. Estes sinais são processados no córtex posterior e
concomitantemente nas áreas superiores do córtex frontal anterior (rostral). Em ambos os
córtices, os sinais são integrados simultaneamente com as informações prévias (as regras
de uma determinada tarefa e as instruções eventualmente dadas) antes mesmo de serem
enviados para o processamento em estágios inferiores da hierarquia frontal. Sendo assim, o
córtex frontal anterior integra as mais elaboradas associações da informação sensorial que
estão armazenadas em redes dos córtices sensoriais e motores.
Se considerarmos que a execução de uma ação não se limita, em geral, a uma única
oportunidade, temos uma grande vantagem ao construirmos representações perceptivas do
mundo e guardá-las na memória, podendo usar essa informação em uma próxima
oportunidade em que ações semelhantes sejam requeridas. Esse aprendizado permite um
refinamento a longo prazo de nossas ações, fornecendo subsídios para ações mais
complexas, mais integrativas e de maior alcance adaptativo.
Integrando percepção e ação: o sistema de neurônios espelho
Quando temos que explicar uma ação humana, a neurociência tem duas abordagens
maiores: a sensoriomotora e a ideomotora. Na abordagem sensoriomotora, tudo começa
com uma estimulação, e as ações são consideradas uma conseqüência desta estimulação.
De modo inverso, na abordagem ideomotora, tudo começa com uma intenção, e as ações
são consideradas como o meio de realizar estas intenções, isto é, as ações são vistas como
o meio para determinados fins que seguem a intenção.
Assim como vimos acima, existe uma sobreposição e uma dependência entre as
percepções e as ações, tanto nos seus sistemas quanto nas respostas comportamentais.
Desde modo, fica difícil imaginar que nossas ações sejam meras escravas de nossas
percepções.
18 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
Em uma situação em que uma pessoa observa as ações de outra pessoa, a
abordagem ideomotora oferece uma predição muito consistente. Considerando o fato de
sermos seres sociais, nós humanos passamos boa parte do nosso tempo observando as
outras pessoas, tentando entender o que elas estão fazendo e por que. Esta “comunicação
primitiva” é essencial para estratégias de sobrevivência e sociabilidade do indivíduo.
Contudo, como reconhecemos e entendemos as intenções das outras pessoas? Quais as
bases neurofisiológicas desta habilidade? A recente descoberta de neurônios espelho tem
inspirado uma série de estudos em busca destas respostas.
O reconhecimento de uma ação foi inicialmente concebido como baseado apenas no
sistema visual (abordagem sensoriomotora); isto é, numa análise dos componentes visuais
da ação específica, do agente envolvido, do objeto ao qual a ação é direcionada e do
contexto no qual ela está inserida. Assim, a interação de todos estes elementos identificados
visualmente permitiria ao observador reconhecer e entender uma ação feita por outra
pessoa. Uma hipótese alternativa admite que a observação de uma ação estimularia uma
“representação motora interna” que envolveria as mesmas estruturas neurais envolvidas na
execução da ação observada; de acordo com esta concepção, embora nenhum movimento
efetivo seja executado, a representação motora evocada pela observação permitiria o
reconhecimento do significado do que é visto. Com a descoberta de que há ativação de
neurônios na região do córtex pré-motor durante a observação de ações, os assim
denominados “neurônios espelho”, e considerando que esta hipótese não exclui a
possibilidade de que outro processo cognitivo, baseado na descrição do objeto e do
movimento, possa participar desta função, esta hipótese motora vem ganhando cada vez
mais adeptos. Todavia, tem sido proposto que os neurônios espelho formam um sistema
que combina observação e execução – percepção e ação.
Neurônios espelho são um grupo particular de neurônios cuja atividade aumenta
durante a execução de uma ação motora particular ou da observação da mesma ação
desempenhada por outro indivíduo. Sua descoberta ocorreu durante experimentos com
macacos envolvendo o controle motor de ações desempenhadas com as mãos, como por
exemplo, pegar/manipular um objeto ou alimento. Os descobridores destes neurônios, entre
eles Giacomo Rizzolatti, implantaram eletrodos no córtex frontal inferior de macacos (área
F5) e registraram a atividade dos neurônios individualmente enquanto os animais
alcançavam pedaços de alimentos. Eles observaram que alguns destes neurônios (situados
no setor superior da área F5), disparavam não somente quando o macaco pegava o
alimento, como também quando ele observava outro indivíduo (macaco ou humano)
desempenhando esta ação, como se a mesma tivesse sido “refletida” no seu córtex motor
(Fig. 3). Estudos posteriores mostraram que pelo menos 10% dos neurônios envolvidos no
controle motor de ações desempenhadas com as mãos são “neurônios espelho”.
19 www.ib.usp.br/labnec
Figura 3 - Experimento feito com macacos em que
ele executa uma ação (pegar o amendoim) e também
observa esta mesma ação sendo feita pelo
experimentador. À direita está um esquema que
exemplifica a atividade dos neurônios espelho nas
duas situações. Retirado de Rizzolatti,1996.
Estes estudos mostram que além do reconhecimento da ação motora por meio de
informações visuais, o sistema de neurônios espelho lida com informações mais abstratas, a
fim de reconhecer o objetivo final da ação. Esta resposta, baseada também em outras
modalidades, isto é, auditiva, sugere que a atividade espelho depende da riqueza das
experiências próprias do observador e de ações presentes em seu repertório motor
(memória de planos motores). Entretanto, aparentemente, o reconhecimento do objetivo
final de uma ação baseado em exposição prévia do observador só parece possível se
houver dicas suficientes no ambiente acerca da intenção desse outro indivíduo. Isto é, uma
ação implica em um agente e um objetivo. Conseqüentemente, o reconhecimento de uma
ação implica no reconhecimento de um objetivo e, em outra perspectiva, o entendimento da
intenção do agente: “João vê Maria pegando uma maça”. Vendo sua mão movimentando-se
em direção à maça, ele reconhece o que Maria fará (pegará algo), e também reconhece que
Maria quer pegar uma maça, isto é, o estímulo é ligado à intenção do agente. Desta maneira, o sistema de neurônios espelho oferece um modelo de integração
entre percepção e ação bastante interessante. Através do reconhecimento de ações e, não
apenas pelo sistema sensorial, mas também no próprio sistema motor do observador, ocorre
uma integração online das informações recebidas do ambiente - a ação observada sendo
executada por outra pessoa - e também entre informações presentes no sistema nervoso do
observador - representação motora da ação observada.
20 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
Percepção
Felipe Viegas Rodrigues
Laboratório de Neurociência e Comportamento
[email protected]
Percepção é um produto do sistema nervoso central que depende do
entendimento dos sistemas sensoriais, mas vai além destes. Entender percepção é
entender não somente como percebemos alguma coisa (seja vendo, ouvindo ou sentindo
estímulos), mas também por que percebemos e quais as implicações para com outros
aspectos da cognição, como a memória ou a atenção. Falar em percepção é falar sobre os
córtices associativos. Esse campo de estudo lida com dois problemas: (1) como todos os
aspectos de um estímulo sensorial são entendidos e processados (cor, forma, movimento
para visão; intensidade, timbre, altura para audição, por exemplo) e (2) qual a relação com
outros produtos da cognição, especialmente atenção e memória.
Uma das principais diferenças entre a percepção e as sensações é a constância
perceptual. Tome por exemplo a Fig. 1. Não importa qual a posição do carro mostrado na
figura, sabemos que se trata do mesmo carro, apesar das quatro imagens serem distintas e
provocarem estimulações diferentes
nas
regiões
visual.
O
iniciais
mesmo
do
sistema
princípio
é
verdadeiro para a percepção de uma
mesma nota musical tocada por
instrumentos diferentes. Embora as
frequências
produzidas
por
eles
sejam diferentes, com alterações dos
harmônicos que compõem o som
Figura 1 – A imagem na retina é imensamente diferente para os
quatro desenhos. Ainda assim, perceptualmente logo nos damos
conta de que se trata do mesmo carro. Retirado de Gazzaniga,
Ivry e Mangun (2006).
resultante (dando a cada instrumento
seu timbre), a percepção de uma
determinada nota é mantida.
A constância perceptual só é possível pela integração da informação sensorial
com a informação de outras regiões encefálicas, inclusive (ou talvez principalmente) das
memórias adquiridas ao longo da vida. Esse mecanismo depende, portanto, de aprendizado
e ele é possivelmente uma particularidade da espécie humana. Experimente colocar um
capacete de ciclismo (que cobre apenas a parte superior da cabeça) e aparecer diante do
seu cachorro. Ele seguramente o estranhará. Por outro lado, o reconhecerá pelo cheiro e
voz, o que o fará parar de hesitar após algum tempo. Humanos são únicos em sua
capacidade de abstração, capazes de ver um tronco cortado em uma floresta e
imediatamente pensar: “Que bom! Um banco para descansar!”.
21 www.ib.usp.br/labnec
O interesse pelos mecanismos de percepção veio a partir de casos clínicos de
lesões cerebrais, em geral por acidentes vasculares cerebrais (AVC), em que os pacientes
tiveram comprometimento da percepção. Tais pessoas se tornaram incapazes de
reconhecer objetos ou pessoas que antes lhes eram muito familiares. Uma investigação
minuciosa evidencia que tais pessoas podem descrever em detalhes o que lhes é pedido, o
que descarta problemas de memória. Mais do que isso, a estimulação por outra modalidade
sensorial resulta em imediata identificação do objeto ou pessoa em questão, levando ao
entendimento de que o problema é perceptual e, em geral, associado a apenas uma
modalidade sensorial. Ao conjunto de sintomas de incapacidade de percepção é dado o
nome agnosia.
Vias perceptuais
As lesões cerebrais que levam a problemas de percepção frequentemente são
aquelas que ocorrem em áreas dos córtices parietal posterior, temporal inferior ou face
lateral do córtex occipital. Essas regiões encontram-se na confluência das áreas sensoriais
e, como já mencionado, são parte dos chamados córtices associativos, pois recebem
aferências corticais das regiões sensoriais e integram entradas múltiplas para desempenhar
funções cognitivas supramodais e comportamentais específicas. Algumas dessas regiões
são neoformações em primatas e elas constituem a maior parte do córtex cerebral,
particularmente no caso da espécie humana (Preuss, 2006).
Visão
O sistema visual é a modalidade mais estudada de todos os sistemas sensoriais
conhecidos. No capítulo sobre fisiologia sensorial foi possível entender como se dá o
processo de transdução do estímulo luminoso em sinal elétrico e como essa informação é
levada até o córtex. Vamos elucidar agora como essa informação é manipulada e integrada
com informações de outras regiões corticais para, de fato, entender como percebemos.
A informação que chega até o córtex visual não para em V1, muito pelo contrário,
essa informação continua avançando por diferentes regiões, cada vez mais próximas dos
córtices temporal inferior e parietal posterior, passando por populações de neurônios
especializadas no processamento de características específicas de um estímulo visual. Uma
particularidade desse sistema sequencial é que a cada sinapse que é realizada a partir de
V1, mais fibras vão convergindo para um mesmo neurônio. Com esse arranjo, quanto mais
adiante na sequência esteja um neurônio, mais específica é sua função no processamento
visual: enquanto aqueles no início da cadeia de processamento disparam para simples
estímulos em forma de barra (com populações específicas para as diversas angulações
possíveis dessa barra), há neurônios mais adiante nessa cadeia que só dispararão para
22 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
combinações dessas barras ou
se o estímulo em questão tiver
características de um móvel
(Fig. 2).
Apesar do arranjo sequencial,
as evidências atuais apontam
para um processamento em
paralelo
regiões.
dessas
Casos
diversas
clínicos
de
pacientes que tiveram um AVC
em regiões muito específicas
do
encéfalo
(nos
córtices
associativos) revelam a perda
de
percepção
de
algum
componente da visão, como
movimento ou cor, mas não de
outras características, mesmo
que estas sejam processadas
mais adiante na sequência de
Figura 2 – Estrutura sequencial na organização dos córtices
associativos do SNC. Quanto mais adiante na sequência, mais
complexo é o estímulo para qual a população de neurônios irá
responder. Modificado de Lent, 2006.
processamento visual. O maior
tempo
detecção
de
de
reação
um
para
estímulo
visual quando mais de uma característica precisa ser analisada em um teste perceptual (cor
e forma, por exemplo) também reforça a ideia de processamento em paralelo. Se apenas
uma das características for necessária para a detecção do estímulo, independente de qual
delas, o tempo de reação é menor.
Na Fig. 3 pode ser vista uma representação das diferentes regiões de
processamento visual e o papel de cada uma delas na construção de um percepto visual.
Vale ressaltar que o arranjo existente nos permite definir uma via dorsal e outra ventral de
processamento. Através da via dorsal, podemos entender “onde” vemos um objeto, já que
essa via nos trás informações sobre movimento e posição espacial de um objeto. Já a via
ventral nos traz informações de “o quê” vemos, permitindo identificar características como
cor e forma de um objeto.
23 www.ib.usp.br/labnec
Figura 3 – Vias paralelas de processamento do estímulo visual: via dorsal (córtex parietal posterior), para
processamento de informações sobre localização espacial e movimento, e uma via ventral (córtex temporal
inferior), para processamento de informações como cor e forma do objeto em questão. Retirado de Kandel e col.
(2000).
Evidências clínicas, mais uma vez, não deixam dúvidas de que essas vias
colaboram de forma independente para a percepção de um objeto qualquer. Um paciente
com lesão em regiões da via ventral poderá afirmar não existir uma caneta (objeto) sobre
uma mesa diante dele. Apesar disso, se ele for instruído a imaginar um objeto sobre a mesa
e demonstrar como seria o movimento para pegar esse objeto, esse indivíduo faria o
movimento correto e até mesmo poderia pegar a caneta. A ativação de todas as regiões
corticais é necessária para que possamos ter a “correta” percepção de um objeto à nossa
frente; o uso de aspas justifica-se porque, falando-se em percepção, simplesmente não há
“correto”, mas sim uma experiência pessoal que é fortemente influenciada pelas nossas
memórias, emoções e a atenção deslocada a um dado estímulo do ambiente. Falaremos
mais sobre isso nos tópicos seguintes.
Audição
O sistema auditório e seus córtices associativos adjacentes têm sido mais bem
estudados nos últimos anos. Novos experimentos têm trazido evidências de que o
processamento de diferentes características do som também ocorre em diferentes regiões
corticais. Semelhantemente ao sistema visual, existiriam duas vias de saída para os córtices
associativos: uma anteroventral, relacionada à percepção de características do som como
timbre e tonalidade; e outra posterodorsal para a percepção de características espaciais e
localização do estímulo.
De fato, Bendor e Wang (2005) encontraram no córtex auditivo de saguis-comuns
(na região anteroventral) neurônios capazes de perceber tons, isto é, que disparam para
24 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
uma determinada frequência e também para seus múltiplos. Essa relação entre frequências
é exatamente aquela encontrada entre duas oitavas musicais. Essa população de neurônios
provavelmente existe também em outras espécies de primatas, incluindo os humanos. É
possivelmente pelo disparo desses neurônios que identificamos as notas semelhantes entre
dois instrumentos musicais diferentes. Como no carro da Figura 1, é a constância perceptual
acontecendo para estímulos auditivos.
Por outro lado (ou, melhor dizendo, por outra via...), morcegos são um exemplo
brilhante da capacidade de localização por estímulos sonoros. Acredita-se que eles sejam
capazes de estabelecer um mapa do ambiente por onde se locomovem tão preciso quanto
aquele que estabelecemos pela estimulação visual. Tentar imaginar algo como isso é quase
impossível, mas, novamente, isto é apenas um reflexo da forma como percebemos o
mundo. Seria como tentar imaginar como um cego (de nascença) percebe o mundo. Embora
você provavelmente tenha pensado em fechar seus olhos e prestar atenção aos sons,
cheiros e pressões (táteis) ao seu redor, isto não é o que um cego percebe do mundo. Para
ele a estimulação visual nunca existiu, logo, perceber o mundo não é “ver” uma imagem
preta e atentar às outras sensações. Para ele, são apenas as outras sensações.
Há casos bem documentados de pessoas que conseguiram desenvolver a
capacidade de se ecolocalizar (como os morcegos) para se locomover. Essas pessoas
parecem criar mapas rudimentares do ambiente, precisos o suficiente para se locomoverem
sem maiores problemas.
Memórias atentas ao contexto
Em diversos mamíferos, após um estímulo percorrer todos os circuitos
necessários à sua percepção (ainda que de forma inconsciente), invariavelmente ele
chegará à região anterior do lobo frontal (ou estruturas homólogas). Essa região está
envolvida com memória operacional e atenção, especialmente no caso de primatas (e
possivelmente em outros mamíferos), e é onde o estímulo será integrado com memórias
passadas e, se o estímulo tiver maior relevância para o organismo (ou simplesmente se for
um estímulo muito forte – como um ruído muito alto), ganhará maior processamento neural
destes circuitos, resultando em um fenômeno que chamamos comumente de atenção.
É interessante notar que a definição de qual estímulo receberá atenção em um
dado momento também dependerá do contexto em que se encontra uma pessoa. Imaginese na sua rotina diária no colégio alguns anos atrás. Você consegue se lembrar com que
facilidade você percebia o sinal da sua escola soar perto do horário de ir embora? Ou
mesmo quantos “alarmes-falsos” você tinha durante essa espera? Da mesma forma,
círculos vermelhos não devem significar nada para você neste exato momento, mas eles
terão muita importância quando estiver dirigindo para algum lugar. Essas diferenças sutis
25 www.ib.usp.br/labnec
naquilo que percebemos são produto de ativação de circuitos de atenção e das memórias
que acumulamos ao longo da vida.
Ilusões e hemisférios cerebrais
Ter memórias significa aprender sobre o ambiente que nos rodeia. Quando essas
memórias são integradas com nossa percepção, não é raro que tenhamos uma visão
distorcida daquilo que está diante de nós. Tome por exemplo a Fig. 4A. Qual das duas
barras horizontais é maior? À primeira vista, todos dirão que a barra superior é maior.
Apenas alguns, após uma análise mais cuidadosa, dirão que ambas tem o mesmo tamanho.
Isso não significa que falhamos em enxergar. Apenas nos deixamos levar pelo aprendizado
que tivemos em toda nossa vida: ao longo dos anos, vemos que linhas de mesmo tamanho
parecem diferentes quanto mais distantes elas estão de nós. As barras convergentes na Fig.
4 criam a ilusão de algo que se distancia. Assim, percebemos as barras paralelas como
sendo de diferentes tamanhos. Olhe a Fig. 4B e isso ficará ainda mais claro.
Figura 4 – Ilusão de Ponzo. As
linhas paralelas em (A) parecem ter
diferentes tamanhos, apesar de
serem iguais. Em (B) uma possível
explicação biológica para esse
efeito. (A)
(B)
Nosso treino para perceber formas geométricas nos faz enxergá-las até mesmo
onde elas não existem. A Fig. 5 sugere o formato de um triângulo, mas sem todas as suas
bordas esperadas, de fato. A figura é conhecida como Triângulo de Kanisa. Algumas
pessoas chegam a dizer que ele é mais branco que as áreas em volta! A explicação direta é
que nos acostumamos a enxergar com mais luz algo que está em primeiro plano.
As ilusões de óptica não se resumem apenas a
fenômenos mnemônicos (que dizem respeito à memória).
Há também efeitos causados pelos próprios receptores
sensoriais. Você provavelmente já se deparou com
imagens como as que estão na Fig. 6. A estimulação de
um determinado receptor retiniano para cor por um
período
prolongado
leva
à
percepção
da
cor
complementar correspondente, o que faz com que, ao
Figura 5 – Triângulo de Kanisa.
olhar para um fundo neutro (branco, preto ou qualquer
tom de cinza), perceba-se cores trocadas na imagem.
26 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
Figura 6 – Efeito de pós-imagem. Uma ilusão criada pelos receptores sensoriais quando
superestimulados por uma determinada cor. Olhe fixamente por cerca de 30 segundos para
qualquer um dos pontos pretos nas imagens e, em seguida, para uma parede branca. O que você
vê?
Essa questão torna-se extremamente importante quando pensamos em contraste.
A percepção de uma cor em um determinado momento é influenciada não somente pela cor
em si, mas pelas cores em volta da mesma. Quão diferentes são as cores dos quadrados
“A” e “B” na Fig. 7? A resposta correta é: nada diferentes! Não há modificações! Isso
acontece porque as cores ao redor da cor atentada influenciam a percepção da mesma.
Figura 7 - Os quadrados
“A” e “B” da figura são
diferentes na cor? Não! Os
quadrados
não
são
diferentes! De forma mais ampla, somos influenciados por diferenças entre nossos
hemisférios cerebrais. Apesar de estes trabalharem sempre em conjunto, com ativações
bilaterais, diferenças sutis na ativação refletem certas dominâncias inter-hemisféricas que
podem também resultar em diferenças na percepção. Testes com pacientes que sofreram
um AVC e estudos com animais lesionados sugerem que o hemisfério esquerdo se
encarrega primordialmente da percepção de detalhes de uma imagem, enquanto que o
hemisfério direito se encarrega das características globais. Veja na Fig. 8 como estes
pacientes desempenham em um teste simples de cópia de uma figura. Essas diferenças
manifestam-se também na percepção de figuras com conteúdo ambíguo. O que você
percebe à primeira vista na Fig. 9?
27 www.ib.usp.br/labnec
Figura 8 – Desempenho de pacientes com hemisférios
cerebrais paralisados em um teste de cópia de figura.
Pacientes que tem apenas o hemisfério esquerdo
funcionante, percebem os detalhes das imagens
originais, mas perdem a forma global. Por outro lado,
pacientes com apenas o hemisfério direito funcionante
percebem a forma global, mas não se dão conta dos
detalhes. Retirado de Lent, 2006.
Sinestesia
A sinestesia é um caso muito
específico
de
percepção
em
que
uma
determinada modalidade sensorial gera a
percepção de outra modalidade. Um dos
eventos mais frequentes é a percepção
secundária de cores após a estimulação primária por um grafema, seja um número ou uma
letra (ou até mesmo palavras). A percepção induzida pelo estímulo primário é sempre muito
específica e unidirecional (a estimulação pelo percepto induzido não gera a percepção do
estímulo indutor pareado, isto é, se a palavra “casa” induz a percepção da cor amarela, o
contrário não acontecerá). Um sinesteta pode repetir mais de centenas de pares de
percepções com pouco ou nenhum erro.
Frequentemente a percepção induzida
é a de cores, seja por grafemas, como dito
acima, ou por sons (palavras em geral); mas há
relatos bem documentados de palavras gerando
percepção de gostos, gostos gerando formas,
cheiros para cores e, mais curiosamente, música
(ou intervalos tonais ou simplesmente tons) para
cores ou formas. As percepções secundárias de
gostos e também cheiros são menos comuns,
embora exista pelo menos um caso bem
documentado de percepção secundária de
gostos induzida por intervalos tonais (musicais).
A mesma pessoa reporta possuir o caso mais
comum de sinestesia entre tonalidades musicais
Figura 9 – O que você vê nesse quadro? e cores.
A investigação sobre o fenômeno é ainda muito recente e algumas perguntas
básicas sobre o assunto só agora começaram a ser respondidas. Em relação aos
mecanismos neurais que possibilitam a sinestesia, duas proposições foram feitas:
28 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
alterações estruturais e alterações funcionais. A Fig. 10 apresenta um resumo dos modelos
de mecanismos possíveis.
Figura 10 - Modelos de Sinestesia. Os modelos diferem na rota proposta de ativação cruzada (direta ou indireta)
entre as regiões indutora e concorrente e nas diferenças subjacentes ao sinesteta (estruturais ou funcionais).
Regiões em amarelo estão ativas (começando pela região indutora) e, em azul, inativas. Conexões excitatórias
são mostradas como flechas e inibitórias como pontas em traço. Linhas pontilhadas representam conexões
presentes estruturalmente, mas funcionalmente inativas. Modificado de Bargary e Mitchell (2008).
As evidências de casos clínicos e fenomenologia da sinestesia apontam mais
fortemente para alterações estruturais na conectividade cerebral, com ligações anormais
entre as regiões indutora e induzida no cérebro de sinestetas (Bargary e Mitchell, 2008).
Vale ressaltar que diferentes possuidores de uma mesma sinestesia (tons para cores, por
exemplo) podem reportar associações diferentes para a cor induzida. Se um deles disser
que um dó maior é azul, o outro poderá dizer: “Isto está errado!”. Não se sabe por que a
indução de cores é muito mais frequente que a indução de outras percepções.
Diferenças na manifestação da sinestesia ainda levaram à sugestão de uma
classificação em dois tipos de sinestetas: (1) de ordem baixa e (2) de ordem alta
(Ramachandran e Hubbard, 2003). Essa divisão leva em consideração o estágio de
processamento em que ocorre o fenômeno perceptual. Sinestetas de ordem baixa tendem a
ter o efeito de indução apenas com estímulos muito específicos, por exemplo: números
escritos na língua de origem. Já os sinestetas de ordem alta têm o efeito de indução toda
vez que o conceito que um determinado indutor sugere está presente. Tomando por base o
exemplo anterior, nos sinestetas de ordem alta mesmo algarismos escritos em números
romanos (que nada mais são do que letras) poderiam gerar a percepção induzida.
29 www.ib.usp.br/labnec
A incidência da sinestesia na população mundial é de algo entre 1% e 4% (Simner
e colaboradores, 2006), um valor bem diferente dos 0,05% anteriormente sugeridos.
Estudos em primatas dão indícios de que essas conexões “anormais” estão naturalmente
presentes no organismo durante a fase fetal e o período de lactância, mas após esse
período essa hiperconectividade de regiões sensoriais tende a ser removida do cérebro. Isto
ainda
não
fora
comprovado
em
recém-nascidos
humanos,
mas
observações
comportamentais levam à sugestão de que há uma “confusão sinestésica” nas primeiras
semanas de vida. A plena maturação perceptual e a segregação dos sentidos viriam apenas
após alguns poucos meses de vida, portanto. De qualquer forma, não ouse afirmar que um
sinesteta tem sentidos menos maduros ou perguntar a ele “como é viver assim?”. A resposta
sempre presente após essa pergunta é: “Como você vive assim?!”.
Concluir é um problema
Uma das maiores questões ainda não respondidas com respeito à percepção é
como geramos um percepto único das estimulações constantes à nossa frente se aspectos
diferentes de um estímulo são processados em regiões distintas do córtex cerebral (e.g. cor,
forma, movimento, etc., no caso da visão). É o chamado binding problem.
Uma das possíveis explicações para a forma como geramos um percepto é a de
que, pelo sequenciamento de neurônios no encéfalo, com cada vez mais neurônios se
juntando em um próximo neurônio (e, consequentemente, complexando o estímulo
processado), ao final do processamento, invariavelmente todas as informações sobre o
estímulo estariam ali reunidas. A quantidade de regiões envolvidas e a divisão do
processamento em duas vias (dorsal e ventral), porém, não favorece essa explicação.
Parece mais plausível aos pesquisadores que o encéfalo forme um percepto único
pela sincronização do disparo dos neurônios das diferentes regiões corticais, ainda que
cada uma delas esteja envolvida no processamento de distintos aspectos de um estímulo
apresentado. Essa explicação, porém, ainda carece de comprovações.
O estudo de casos de sinestesia tem trazido algumas colaborações para aquilo
que entendemos sobre percepção. Alguns sinestetas relatam a percepção de cores
estranhas, diferentes de qualquer cor que eles já tenham visto em algum objeto ou lugar.
Um deles chegou a chamar essas percepções sinestésicas de “cores marcianas”.
Ramachandran e Hubbard (2003) atribuem essas cores estranhas à ligação cruzada (ou
direta) de um córtex sensorial para outro, o que “desviaria” o processamento de estágios
iniciais da percepção de cores. Segundo os autores, isso sugere que a experiência subjetiva
da percepção de cores depende não só do processamento final, mas de todo o padrão de
atividade neural que leva à formação de um percepto, incluindo as fases iniciais do
processo.
30 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
Estando certa ou não a sugestão dada por Ramachandran e Hubbard (idem), fica
claro que ainda precisamos entender muito sobre os mecanismos pelos quais simplesmente
percebemos o mundo que está ao nosso redor. Ou talvez um dia tenhamos a certeza de
que, desde sempre, apenas representamos internamente o que é percebido externamente.
Pelo menos é isso que os estudos sobre atenção e memória sugerem cada vez mais
fortemente.
31 www.ib.usp.br/labnec
Atenção
Wataru Sumi
Laboratório de Neurociências e Comportamento
[email protected]
A todo instante somos expostos a uma grande quantidade de estímulos ambientais
que são captados por nossos órgãos sensoriais. A maioria desses estímulos não é
percebida pelo observador, apenas alguns selecionados para serem posteriormente
analisados. Acredita-se que o sistema nervoso é incapaz de processar todas essas
informações. Assim, para permitir um processamento eficiente, a atenção selecionaria
apenas algumas poucas informações que nós recebemos (Bear e col., 1996).
O conhecimento acumulado sobre atenção no último século tem trazido à luz, mais e
mais fenômenos relacionados com esse processo cognitivo. A partir da década de 1950,
muitos cientistas cognitivos propuseram diferentes teorias para abarcar o conjunto de dados
revelados pelas mais diversas técnicas de avaliação do comportamento e de medições das
atividades do cérebro. O presente texto será focado nos aspectos comportamentais da
atenção, portanto, não nos ateremos aos processos neurofisiológicos responsáveis pela
atenção nem nas diferentes teorias criadas ao longo das últimas décadas para explicar o
funcionamento geral desse que é um dos mais interessantes e enigmáticos processos
cognitivos.
Atenção e percepção
Como mencionado anteriormente, a atenção seleciona um conjunto de informações
do ambiente enquanto ignoram outros. Veremos ao longo do texto diferentes exemplos de
experimentos nos quais é evidenciado esse fenômeno, principalmente na atenção visual.
Antes, começaremos com os efeitos da atenção sobre a percepção auditiva.
Na década de 1950, Cherry realizou um experimento no qual era avaliada a
capacidade
de
selecionar
um
dentre
dois
estímulos
auditivos
simultaneamente
apresentados. O voluntário utilizava fones de ouvido e recebia diferentes estímulos, um para
cada ouvido. O voluntário era então instruído a prestar atenção apenas a um dos ouvidos.
Ele observou que os voluntários eram incapazes de relatar o que foi apresentado ao ouvido
não atendido (Gazzaniga e col., 2002). Esse efeito não aparece apenas quando ouvimos
estímulos diferentes em cada ouvido. Somos frequentemente expostos a situações nas
quais recebemos diferentes estímulos auditivos, selecionando os que nos interessa e
ignorando os demais. Para ilustrar melhor esse efeito, podemos nos imaginar em uma festa:
existem dezenas de pessoas, umas falando mais alto que outras, além da música no volume
32 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
máximo. Apesar disso, somos capazes de selecionar estímulos específicos que nos
interessam como a fala de um amigo ou eventualmente a música sendo tocada.
Apesar da avaliação da atenção auditiva ter contribuído bastante com entendimento
da atenção, os estudos nessa área concentram-se principalmente na atenção visual. Isso
fica claro quando tanto pelo numero de artigos publicados como pela diversidade de tarefas
desenvolvidas pelos cientistas. Consequentemente, o conhecimento acumulado sobre esse
sistema perceptual é muito maior. Vamos a seguir ver os diferentes efeitos comportamentais
da atenção sobre a percepção visual.
Para compreendermos melhor a atenção, podemos utilizar a metáfora do holofote:
enxergamos os objetos iluminados pela luz, mas não somos capazes de enxergar os objetos
que permanecem nas sombras. De forma semelhante, para percebermos os estímulos do
ambiente, eles devem estar sob o foco da atenção. Um experimento realizado no final do
século XIX por Herman von Helmholtz demonstra isso claramente. Nesse experimento, os
voluntários eram colocados em frente a um painel e eram instruídos a direcionar a atenção a
um ponto específico da tela. O painel era mal iluminado e o voluntário era incapaz de
observar qualquer letra impressa nele. Quando um flash de luz era acionado, era então
possível ver as letras. Porém, os voluntários eram capazes de discriminar apenas as letras
localizadas na região onde a atenção estava previamente focada (Gazzaniga e col., 2002).
Como citado anteriormente, não somos capazes de processar eficientemente todas
as informações que recebemos do ambiente. Quando procuramos algo específico no
ambiente, podemos ter mais ou menos facilidade de acordo com a característica do alvo.
Quando um estímulo se destaca muito no meio de outros estímulos, a atenção pode ser
atraída automaticamente, como ocorre, por exemplo, com as luzes intermitentes dos
automóveis. Por outro lado, quando a diferença entre o alvo e os outros elementos da cena
(distratores) é pequena, é necessário procurá-la, ou seja, direcionar voluntariamente a
atenção. Ambos os processos podem ser avaliados na tarefa de busca visual (Fig. 1).
Quando o alvo (1-A: barra vertical e; 1-B: barra vermelha) é muito diferente dos distratores,
a sua detecção é quase imediata, independentemente do número de elementos, ou seja,
podem ser cinco ou cinquenta distratores sem que o tempo para a detecção do alvo seja
afetado. Isso porque a atenção é atraída automaticamente. Porém, quando a diferença entre
alvo (1-C: barra azul horizontal) e os distratores é pequena, o tempo de detecção aumenta
de acordo com o aumento do número de elementos distratores, isso porque é necessário
analisar cada um dos itens isoladamente de forma serial; assim, quanto mais elementos
precisarem ser analisados, maior será o tempo necessário para detectar o alvo (Treisman e
Gelade, 1980).
33 www.ib.usp.br/labnec
Figura 1- Tarefa de busca visual. 1a e 1b: o alvo difere em apenas uma característica em relação aos
distratores: forma e cor, respectivamente. 1c: o alvo possui duas características que o torna diferente dos
distratores (horizontal azul).
Falha na percepção
Quando não prestamos atenção ao ambiente por estarmos distraídos ou por
estarmos prestando atenção fixamente em algo, deixamos de perceber diferentes estímulos.
Existem experimentos nos quais são evidenciados esses efeitos. Um deles é denominado
cegueira inatencional. Isso se caracteriza pela incapacidade do voluntário de reportar a
presença de um objeto no centro de seu campo visual, perfeitamente visível, mas
inesperado, porque a atenção estava engajada em outra tarefa (Neisser e Becklen, 1975).
Outra evidência do papel da atenção na percepção é a cegueira para mudança, um
fenômeno relacionado com a cegueira inatencional, mas com sutis diferenças. Esse efeito
se refere à incapacidade em identificar diferenças entre duas imagens apresentadas em
seqüência (Simons e Rensink, 2005). Essas imagens podem ser fotografias diferentes, algo
como os jogos de sete erros, ou mesmo objetos do ambiente. Em uma “pegadinha”
realizada por uma emissora de TV, o atendente de uma loja abaixa-se atrás do balcão para,
supostamente, pegar uma mercadoria, ele então troca de lugar com outra pessoa que, após
se levantar, continua a interagir com os clientes-vítimas como se nada tivesse acontecido.
Poucos clientes percebiam a troca.
Teste de Posner
Uma das maiores contribuições para os estudos da atenção foi feita por Posner, que
desenvolveu uma tarefa que pode ser utilizada para testar diferentes aspectos da atenção.
Essa tarefa (Fig. 2) consiste em manter o olhar fixo no centro de um monitor de vídeo.
Apresenta-se, então, uma pista indicando o provável (por exemplo, 80%) local de
aparecimento de um alvo, que pode ser para a direita ou esquerda do monitor. Nesse caso,
a pista é chamada de válida. No restante das tentativas (20%), a pista indica o local oposto
de aparecimento do alvo, sendo chamada então de pista inválida. Após o aparecimento da
pista, o voluntário deve direcionar a atenção, mas não o olhar, para o local indicado pela
34 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
pista. Finalmente, após o aparecimento do alvo, o voluntário deve responder pressionando
um único botão, independente do lado e da validade da pista (Bear e col., 1996).
Quando o voluntário direciona a atenção para o local de aparecimento do alvo (i.e.
tentativa com pista válida) ele responde mais rápido do que na situação em que ele
direciona a atenção para o lado oposto do monitor de vídeo. Essa tarefa nos permite avaliar
diferenças entre esses tempos de resposta na ordem de dezenas de milissegundos.
Se alterarmos a intensidade do estímulo alvo, será possível observar claramente os
efeitos atencionais sobre a percepção. Nesse caso, o estímulo deve estar um pouco acima
do limiar de percepção do voluntário. Então, se o alvo for precedido da pista válida, quando
o sujeito direciona a atenção para o local de aparecimento do alvo, ele responde
normalmente, mas, quando o voluntário direciona a atenção para o lado errado em
decorrência da pista inválida, ele simplesmente não vê o alvo.
Figura 2- Tarefa de orientação espacial da atenção. O voluntário deve manter o olhar fixo no centro do monitor.
Ele será, então, instruido a direcionar a atenção para um dos lados de acordo com uma pista e, finalmente,
responder ao aparecimento do alvo. Retirado de Lent, 2002.
Uma alteração dessa tarefa pode nos mostrar o efeito do direcionamento atencional
na percepção da coincidência temporal da apresentação de estímulos. Nessa tarefa, os
voluntários eram orientados a direcionar a atenção para um dos lados do monitor, de acordo
com uma pista sinalizadora, mas ao invés de aparecer apenas um alvo, à direita ou à
esquerda, apareciam dois alvos, um de cada lado, mas com um intervalo de algumas
35 www.ib.usp.br/labnec
dezenas de milissegundos. Observou-se que os voluntários percebiam um estímulo mais
rapidamente quando prestavam atenção ao local de aparecimento; por exemplo, se ele
direcionasse a atenção para o lado direito e aparecesse um estímulo à esquerda e 30ms
depois na direita, o voluntário relataria que os alvos apareceram simultaneamente, apesar
da defasagem temporal entre os estímulos (Stelmach e Herdman, 1991).
Efeitos das lesões do sistema nervoso na atenção
Até agora, vimos os efeitos produzidos pela alocação ou não da atenção nas
atividades do cotidiano ou em condições experimentais que nos auxiliam a entender como
ela funciona. Outra fonte muito importante de informação sobre as relações desse processo
cognitivo que tem contribuído com os avanços nessa área é a observação de indivíduos
com graves deficiências atencionais, decorrentes de lesões provocadas por AVC, tumores,
traumas, etc.
Lesões no córtex parietal, na junção com o córtex temporal (principalmente no
hemisfério direito), produzem um efeito conhecido como síndrome de heminegligência (Fig.
3), que consiste em ignorar objetos ou eventos presentes no lado oposto à lesão (Robertson
e Rafal, 2000). Apesar do prejuízo em relatar eventos no campo contralateral, os pacientes
dessa síndrome ainda são capazes de identificá-los precariamente, porém, quando
estímulos são apresentados simultaneamente em ambos os lados do campo visual, eles
identificam apenas os estímulos apresentados no lado ipsolateral (Gazzaniga e col., 2002).
Esse efeito é chamado de extinção.
Figura 3- Desenho feito por um paciente com a
sindrome de heminegligência. À esquerda,
desenho modelo e, à direita, cópia feita pelo
paciente. Metade da figura é ignorada pelo
paciente. Modificado de Gawryszewski e col.,
2007.
Em um experimento clássico é possível observar que a heminegligência não afeta
apenas a percepção. Um paciente que sofre dessa síndrome foi orientado a descrever uma
paisagem com a qual ele estava bastante familiarizado (a praça central da cidade em que
vivia). Imaginando-se posicionado em um dos lados da praça, ele descrevia apenas a
metade da paisagem. Mas, quando ele era orientado a se imaginar do lado oposto da praça,
36 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
ele descrevia os objetos anteriormente ignorados e mantendo a tendência de ignorar
metade do campo visual (Gazzaniga e col., 2002).
A lesão no córtex parietal dos dois hemisférios cerebrais produz a síndrome de
Balint. O portador dessa síndrome percebe apenas um objeto de cada vez (agnosia
simultânea), mesmo quando dois objetos estão próximos ou sobrepostos (Gazzaniga e col.,
2002). Para esses pacientes, os objetos aparecem de repente e as mudanças do campo
visual para outros objetos são aleatórias. A capacidade de reconhecer rostos, formas, cores
e palavras são mantidas, mas a incapacidade de ver dois objetos simultaneamente faz com
que esses pacientes percam a noção de espaço. Eles são incapazes de dizer se um objeto
está à direita, à esquerda, acima ou abaixo em relação a eles ou outros objetos (Robertson
e Rafal, 2000).
As propriedades atencionais descritas aqui mostram explicitamente a relação entre
atenção e percepção, mas não apenas isso. Sabendo, por exemplo, que a nossa memória
se constrói principalmente a partir do que percebemos do mundo, fica claro também que a
atenção tem grande importância na formação de memória. É importante ressaltar também
que, se por um lado a atenção afeta o funcionamento das outras funções cognitivas, a
recíproca também é verdadeira, pois todas as funções cognitivas são inter-relacionadas,
modulando-se mutuamente, como será visto nas demais aulas desse curso.
37 www.ib.usp.br/labnec
Memória
Leopoldo F. Barletta Marchelli
Laboratório de Neurociência e comportamento
[email protected]
Das propriedades que emergem da organização e funcionamento do sistema
nervoso, a memória é tida como um dos resultados mais fascinantes. O que demonstra a
presença de memória em um organismo é a capacidade que ele tem de alterar seu
comportamento em função de informações adquiridas e armazenadas. Uma vez que ocorra
interação entre ambiente e indivíduo, os sistemas de memórias, aptos a aprenderem sobre
informações
e
regras
ambientais
relevantes
(altamente
informativas),
guiam
adaptativamente o comportamento desses indivíduos. Com o acúmulo de informações, o
sistema nervoso passa a detectar regularidades e antecipar eventos em função de
experiências anteriores. Desta forma, organismos portadores de memórias podem relacionar
grandes quantidades de informações passadas e presentes e selecionar quais receberão
um processamento preferencial por meio do direcionamento da atenção. Isso quer dizer que
em função de experiências prévias, tais organismos podem flexibilizar o controle de seus
comportamentos. Isso lhes garante um repertório de soluções para os mais diversos
problemas que a sobrevivência impõe.
Aspectos comportamentais e evolutivos
Aos olhos da teoria proposta por Charles Darwin em 1859, sistemas biológicos são
tidos como produtos da evolução por seleção natural, que pode favorecer o
desenvolvimento de um sistema mais adaptado. Além de a seleção atuar sobre estruturas e
mecanismos, ela age também selecionando comportamentos. Por exemplo, se um ambiente
é relativamente simples e possui certa regularidade, a seleção natural pode favorecer
indivíduos que sejam capazes de gerar “previsões” de tal ambiente e responder de maneira
antecipatória. Neste caso esses indivíduos estariam então mais aptos para tal ambiente. Se,
no entanto, a complexidade de tal ambiente aumentar, a imprevisibilidade pode tornar-se um
problema. Indivíduos que tiverem um sistema mais flexível, capaz de obter e armazenar o
máximo de informações relevantes sobre o ambiente, estarão mais aptos a reagirem
prontamente a estímulos ambientais.
Assim sendo, serão capazes de solucionarem
problemas de maneira antecipatória quando um padrão regular puder ser identificado. Ainda
sim, mesmo quando um padrão não puder ser identificado e os problemas forem
inesperados, com informações prévias, os indivíduos portadores de sistemas flexíveis de
armazenamento de informações poderão resolver problemas de forma não-antecipatória.
38 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
O processo de evolução do sistema nervoso, sobretudo da memória, parece estar
relacionado com a ideia do desenvolvimento de sistemas seletivos capazes de lidar com
novidades ao longo da vida do individuo. À medida que um individuo consegue identificar
estímulos, prever o ambiente e gerar as “inferências” e respostas mais adequadas, ele se
beneficiará; estará, portanto, mais apto para determinado ambiente. Deste modo, a emissão
de comportamento antecipatório e a resolução de problemas, ambos baseados em
experiências antecedentes, conferem ao repertório comportamental do sujeito alto valor
adaptativo. Uma vez que a emissão de determinados comportamentos diante de algumas
situações também traz ganhos adaptativos, parece razoável considerar que a memória seja
um dos resultados de maior sucesso ao longo da evolução biológica.
O conceito de memória pode estar relacionado com uma ideia de “representação
interna” do ambiente em organismos mais desenvolvidos. Do ponto de vista evolutivo, isso
pode ser altamente vantajoso, pois tais representações permitem avaliar consequências
futuras de ações correntes, sem comprometer de algum modo a integridade do sistema no
desempenho da ação.
O sistema nervoso como uma estrutura que suporta os sistemas de memória
O funcionamento dos sistemas de memória implica no armazenamento de uma
quantidade substancial de informações sobre o ambiente, sobre suas regularidades e sobre
os efeitos de ações anteriores. Essas informações ficam inteiramente armazenadas no
sistema nervoso do indivíduo. Assim, os sistemas de memória são claramente dependentes
da estrutura e do funcionamento do sistema nervoso.
O sistema nervoso de um humano adulto possui bilhões de neurônios, células
nervosas capazes de processar e conduzir impulsos elétricos. O processamento neuronal
visa receber a informação, avaliá-la e passar o sinal a outros neurônios. Cada neurônio
envia projeções para milhares de outros neurônios e, por sua vez, recebe projeções de
outros milhares de neurônios. Uma mensagem passa de um neurônio para outro através da
sinapse. É nesse momento que ocorre modulação do processamento de informações.
Conectadas aos neurônios, há células especializadas para a recepção de informações
ambientais (receptores sensoriais), que transformam diferentes formas de energia (e.g. luz,
som, odores etc.) em potenciais elétricos, influenciando assim tanto a atividade elétrica
quanto a química dos neurônios. Essas informações sensoriais, sob a forma de impulsos
elétricos, são transmitidas por circuitos definidos do sistema nervoso, havendo circuitos
neurais dedicados ao processamento preferencial de informações de cada uma das
modalidades sensoriais, e outros circuitos responsáveis pela integração de informações de
diferentes modalidades sensoriais.
39 www.ib.usp.br/labnec
Aspectos fisiológicos da memória
Pressupõe-se que a atividade eletrofisiológica, gerada por atividade espontânea,
estímulos ambientais e respostas a esses estímulos, desencadeie processos que levam à
alteração da conectividade entre células nervosas, alterando a transmissão de impulsos
elétricos por esses circuitos neurais. Todas essas modificações provocadas em elementos
constituintes do sistema nervoso caracterizam (representam) assim o armazenamento de
informações, as memórias. Uma decorrência lógica dessa suposição é que seja possível
detectar a ocorrência de alterações bioquímicas associadas à alteração da conectividade
nervosa relacionada ao processo de formação de memórias.
O conhecimento atual sobre memória é resultado do trabalho de inúmeros
personagens. Gold e colaboradores (1970) expuseram ratos a uma câmara clara conectada,
por uma porta tipo guilhotina, a uma câmara escura cujo assoalho é constituído de barras
metálicas eletrificáveis. Os ratos rapidamente entram na câmara escura; após entrarem
nessa câmara, levam um choque nas patas. Em uma etapa de teste, realizada 24 horas
depois, os animais inseridos na câmara clara não entram na câmara escura (ver a barra
vermelha da Fig. 1). Animais de um grupo controle, que não receberam choque nas patas
no dia anterior, entram rapidamente na câmara escura (ver barra verde da Fig. 1). Em
experimentos adicionais, depois do treinamento com choque nas patas, foram aplicadas
correntes elétricas no sistema nervoso dos animais com diferentes intervalos de tempo entre
o choque na pata e o choque eletroconvulsivo (ver Fig. 1 - esquerda). Observa-se que
quanto menor o intervalo de tempo entre o choque nas patas e o choque no sistema
nervoso, maior é o prejuízo de memória aversiva sobre o ambiente escuro. À medida que
esse intervalo de tempo aumenta, menor é o efeito, como se o choque eletroconvulsivo
perdesse sua efetividade para evitar sua consolidação. (ver Fig. 1 – direita: barras de cor
laranja).
Intervalo de tempo entre choque nas
patas e choque eletroconvulsivo (S).
Figura 1 – Experimento de Gold e colaboradores (1970). A organização temporal dos eventos (esquerda) e os
resultados (direita): o tempo que os ratos submetidos aos diferentes tratamentos demoraram para entrar na
câmara escura – quanto menor o intervalo de tempo entre o choque nas patas e o choque eletroconvulsivo
menor é a lembrança do evento aversivo. Modificado de Pavão (2009) e Gold (1970).
40 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
Outro experimento que trata de questões fisiológicas sobre a memória foi feito por
Shashoua (síntese publicada em 1985). O experimentador prendeu um flutuador nas
nadadeiras peitorais de peixinhos dourados para fazer com que os animais ficassem em
posição desconfortável. Após longo esforço de cerca de 3 horas, alguns peixes voltaram à
posição normal, apesar do flutuador (Fig. 2, treino inicial representado pela curva verde). Se
o flutuador for removido e recolocado três dias depois, os animais realizam a tarefa mais
rapidamente; i.e., os peixes retornam à posição normal em apenas 15 minutos, o que indica
que eles aprenderam e retiveram a solução desse desafio (Fig. 2, curva azul) (para detalhes
sobre esses experimentos, ver Helene e Xavier, 2007). Em outro teste, Shashoua (1985)
injetou valina marcada com hidrogênio radioativo (valina-H*) no ventrículo encefálico de
animais que ficaram por 4h com o flutuador, e valina marcada com carbono radioativo
(valina-C*) no ventrículo de animais que não foram treinados. Os encéfalos dos animais dos
dois grupos foram homogeneizados conjuntamente e as proteínas foram separadas por
peso molecular. A maioria das proteínas presentes estava marcada tanto com valina-H*
quando com valina-C*; porém, algumas delas estavam mais marcadas com valina-H*,
indicando que elas foram incorporadas no cérebro dos animais que aprenderam a tarefa;
essas proteínas foram denominadas ependiminas. Num terceiro teste, as ependiminas
foram isoladas e injetadas em coelhos para produção de anticorpos específicos contra as
ependiminas. Então, os anticorpos foram injetados no ventrículo encefálico de peixes que
tinham acabado de aprender a tarefa de nadar com o flutuador; no teste de memória
realizado 3 dias depois, esses peixes demoraram cerca de 3h para voltar à posição normal
(Fig. 2, curva vermelha). Ou seja, esses animais comportaram-se como se nunca tivessem
sido submetidos ao treinamento. Atualmente, as ependiminas são denominadas “moléculas
de adesão celular” e estão diretamente relacionadas com o fortalecimento e formação de
sinapses.
Figura 2 – Experimentos de Shashoua (1985) envolvendo aprendizagem em peixes dourados. Flutuadores
foram presos aos animais, que em ficavam em posição desconfortável (esquerda, acima). com treino de cerca de
180 minutos, ficavam em posição confortável (esquerda, abaixo) – curva verde. Em segundo momento, 3 dias
depois, os flutuadores foram recolocados, e os animais demoraram cerca de 15 minutos para ficar na posição
confortável, indicando que aprenderam essa habilidade – curva azul. Animais treinados tratados com anticorpos
para proteínas envolvidas com a alteração de circuitos neurais apresentam desempenho similar a animais não
tratados – os traços de memória foram apagados pelo tratamento. Modificado de Pavão (2009) e Shashoua
(1985).
41 www.ib.usp.br/labnec
Em conjunto, os resultados obtidos a partir de experimentos envolvendo choques
eletroconvulsivos e síntese de proteínas sugerem que há dois processos envolvidos na
manutenção da memória. Um deles, mais instável, é prejudicado pelo choque eletroconvulsivo, estando relacionado ao padrão de atividade eletrofisiológica dos neurônios
(frequência de disparos, por exemplo). O outro, associado com produção de proteínas,
parece envolver alterações estruturais nas sinapses, gerando circuitos alterados no sistema
nervoso.
Posteriormente ao experimento de Shashoua, muitos trabalhos com proteínas
associadas aos processos de arquivamento de informação ao nível celular vêm sendo
desenvolvidos. Muitas moléculas subjacentes à formação de memória já foram descobertas.
Isso tem trazido importantes informações acerca das diferentes etapas e modalidades do
processo de formação de memórias em nível celular, inclusive o envolvimento dessas
proteínas na alteração plástica do sistema nervoso.
Plasticidade neural
O sistema nervoso possui a capacidade de se modificar estruturalmente e
funcionalmente em decorrência de estímulos que de algum modo incidem sobre ele. Tal
fenômeno denomina-se neuroplasticidade ou, simplesmente, plasticidade. Inerente ao
funcionamento do sistema nervoso, a neuroplasticidade é uma característica marcante e
constante da função neural. Muito dos processos cognitivos depende de tal propriedade.
Parece haver dois tipos básicos de plasticidade sináptica, uma de curta duração e a
outra de longa duração. A plasticidade sináptica de curta duração pode ser induzida
rapidamente; parece não requerer síntese proteica e mantém-se por, no máximo, algumas
horas. Esse tipo de plasticidade reflete alterações na força de sinapses pré-existentes, pela
modificação de proteínas pré e pós-sinápticas. Diferentemente, a plasticidade sináptica de
longa duração (que parece ter sido a modalidade principal investigada nos estudos de
Shashoua) dura dias, meses ou anos, envolve processos de transcrição gênica e síntese de
novas proteínas; esse tipo de plasticidade sináptica parece envolver a remodelação de
sinapses existentes ou a formação de novas sinapses.
Com base nessas e em outras características do sistema nervoso apresentadas até
aqui, percebe-se que além de aumentar a capacidade de comunicação entre as diversas
populações de neurônios, sua estrutura e funcionamento possibilitam a formação de
memórias em decorrência de experiências vividas. As diferentes modalidades de
arquivamento parecem envolver alguns tipos de alterações no sistema: (1) alterações
transitórias na atividade eletrofisiológica (taxa de disparos) de populações de neurônios, que
estariam ligadas ao arquivamento por curtos períodos de tempo; (2) alteração na facilidade
42 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
com que a atividade eletrofisiológica é transmitida entre neurônios, relacionada com o
arquivamento por períodos intermediários de tempo (que pode durar de minutos até meses);
(3) alterações estruturais permanentes na conectividade neuronal que levam à formação de
circuitos neurais, ou redes nervosas, cuja atividade representaria informações mantidas por
um longo período de tempo, anos ou até mesmo uma vida inteira.
Aquisição e manutenção da memória
Donald Hebb (1949) baseou-se na plasticidade sináptica para afirmar que a
transmissão de informações entre dois neurônios deveria ser facilitada e tornar-se estável
quando ocorresse sincronia entre os disparos do primeiro e do segundo neurônio. Sendo
assim, a transmissão de mensagens entre os neurônios poderia ser regulada: não seria um
fenômeno rígido e imutável, mas sim algo modulável de acordo com as circunstâncias.
Um importante elemento descrito inicialmente no hipocampo que atua na alteração
de sinapses (portanto, na formação de memórias) é o fenômeno denominado potenciação
de longa duração (LTP). Aparentemente, o hipocampo (e outras estruturas do lobo temporal
medial) está envolvido em um processo de ativação repetitiva de circuitos envolvidos na
representação da informação que determina alteração estrutural desses circuitos. Trata-se
de uma plasticidade sináptica específica que ocorre entre um neurônio pré e um neurônio
pós-sináptico, assim como Hebb havia proposto. Acredita-se que a LTP seja um importante
mecanismo envolvido no armazenamento de informações cuja natureza é essencialmente
associativa. Tal mecanismo pode envolver a interação entre diferentes sinapses de um
mesmo neurônio, permitindo que uma sinapse fraca se fortaleça pelo disparo concomitante
com uma sinapse forte, tornando-as associadas. Sendo assim, a LTP permite uma
facilitação na comunicação sináptica.
Com o aumento na frequência de disparos das sinapses produzidas em decorrência
de estímulos ambientais, ocorrem alterações na eficiência sináptica dos neurônios
recrutados, de maneira a intensificar a comunicação dessas células. Uma vez que a
comunicação sináptica seja facilitada, qualquer referência ao estímulo inicial já causa um
disparo das células envolvidas. Com estímulos muito pequenos pode-se desencadear um
processo efetivo de ativação neural. A LTP é fundamental para o arquivamento de
informações sobre eventos experienciados. Esse processo parece essencial para a retenção
de informações sobre “o que” ocorreu, mas não sobre “como” desempenhar uma tarefa
perceptomotora.
Estímulos ambientais e experiências geram atividade eletrofisiológica em conjuntos
de neurônios. Como vimos, essa atividade pode levar à formação de novas sinapses ou à
alteração das sinapses já existentes, o que permite estabelecer circuitos neurais envolvendo
43 www.ib.usp.br/labnec
populações de neurônios cuja atividade, correspondente àquela gerada durante a
experiência original, representa a experiência adquirida.
A recordação da informação representada em circuitos se dá pela ativação
eletrofisiológica de sua população de neurônios; isso pode ocorrer tanto em decorrência de
estímulos que de alguma forma estão relacionados à experiência original, como por um ato
de vontade para recordar aquela experiência. É curioso notar que os sistemas de memória
permitem identificar estímulos muito específicos e responder a eles, mesmo quando estes
não são apresentados em sua totalidade. Uma vez ativos, esses circuitos podem
estabelecer novas conexões com outros circuitos ativos, ou contar com a adição de novos
elementos em decorrência de novas experiências.
Quanto mais frequentes as exposições a estímulos relevantes, mais fortes tornam-se
as conexões. Como consequência, a informação tende a ser arquivada de maneira
relacional. Isso permite entender porque a recordação envolve, usualmente, categorias. Tal
fato ocorre porque o aumento de atividade eletrofisiológica em determinados circuitos
neurais (que levam à recordação de uma dada informação) tende a estimular a atividade em
circuitos relacionados.
É importante ressaltar que os mesmos circuitos neurais associados à atenção,
percepção, ação e outros processos cognitivos, são os que se alteram para a formação de
memórias de diferentes tipos. Isso significa que, quando esses circuitos forem
posteriormente mobilizados, o processamento das informações será diferente em relação às
experiências anteriores, dado que o circuito vem sendo alterado a cada uma delas. Assim
sendo, a percepção e as habilidades se alteram ao longo da história de vida. Além disso,
estão profundamente associadas com os processos de memória.
Redes neurais e memória
Praticamente todas as regiões do sistema nervoso estão envolvidas de alguma forma
no arquivamento de memórias de um tipo ou de outro. Em primatas, costuma-se atribuir
uma grande importância ao neocórtex (a porção filogeneticamente mais recente do córtex)
no arquivamento de informações. Esse tecido envolve sistemas de processamento
modalmente específicos e sistemas de integração de informações de diferentes
modalidades (denominados polimodais e supramodais).
A maioria das experiências humanas inclui diferentes modalidades sensoriais,
organizadas no tempo e espaço. Por exemplo: a partir de uma estimulação perceptual
específica, o sistema nervoso mobilizaria um grupo de neurônios para representar o evento,
por meio de sua atividade e conexões, produzindo uma espécie de "rede" de interconexões
que se mantém em contínua reconstrução ao longo da vida. A formação de uma memória
sobre esse evento envolveria o fortalecimento das conexões entre as células dedicadas a
44 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
essa percepção, resultando num grupamento celular cujas conexões seriam mais eficientes.
Depois do desaparecimento do estímulo gerador da atividade, "nós" da rede, quando
ativados, excitariam ou inibiriam outros nós numa rica e complexa rede de conexões, de
forma que representações seriam mantidas enquanto houvesse reverberação da atividade
nervosa correspondente ao estímulo inicial.
Nessa rede, uma dada população de nós disparando, provavelmente com níveis de
atividade diferentes em várias regiões nervosas, representa uma determinada informação,
enquanto a malha representa as ligações associativas das relações entre os nós; essas
ligações podem variar em intensidade. Nesse sentido, um mesmo nó pode estar envolvido
em representações distintas, já que a informação é representada pelo conjunto de disparos
dos nós a ela relacionados e não por um nó individual. Isso nos sugere que processos de
memória estariam baseados em um funcionamento sistêmico de determinadas populações
de neurônios.
Hebb (1949) propõe algumas previsões sobre o funcionamento da memória. Por
exemplo, parece plausível pensar que estimulações parciais correspondentes à experiência
original sejam capazes de regenerar a atividade em toda a rede, contribuindo para a
lembrança completa da experiência original. Além disso, se dois eventos forem pareados no
tempo supõe-se que haja a formação de redes tais que a estimulação da atividade do
primeiro evento gera o padrão de atividade eletrofisiológica associada ao segundo evento,
levando à sua previsão.
O autor sugere que haveria apenas três aspectos centrais que determinariam o
funcionamento de um sistema neuronal: (1) a conexão entre neurônios é mais eficaz quanto
maior for o grau de relação entre as porções pré e pós-sináptica; (2) grupos de neurônios
que tendem a disparar conjuntamente irão formar agrupamentos celulares cuja atividade se
mantém expressa mesmo após o fim do estímulo que gerou a atividade e; (3) cognição
deriva da atividade sequencial destes agrupamentos celulares facilitados.
Figura 3 - Esquema representativo de redes neurais de Hebb. Os pontos pretos são os neurônios e as linhas
são as conexões. A rede tem uma organização inicial como representado em (A); ao receber um estímulo, é
ativada (B); esse estímulo pode ser apresentado repetidas vezes, ou pode ter reverberado nessa rede, de modo
que as conexões entre os neurônios são fortalecidas (C e D); então, um estímulo mais fraco ou mesmo
incompleto, mas que mantenha algumas das características do inicial (D) é capaz de ativar a rede fortalecida (E).
Modificado de Bear, 2002 e de Helene e Xavier, 2007.
45 www.ib.usp.br/labnec
Modularidade e os diferentes processos de memória
A noção de que a memória compõe um conjunto de habilidades mediadas por
diferentes módulos do sistema nervoso, que funcionam de forma independente, porém
cooperativa, parece atualmente bem difundida. Este conceito de modularidade de funções
tem embasado investigações acerca dos processos de memória. Segundo essa ideia, o
processamento de informações nesses módulos acontece de forma paralela e distribuída,
possibilitando que um grande número de unidades de processamento influencie outras em
qualquer momento no tempo, e que grande quantidade de informações seja processada
concomitantemente.
O refinamento nas técnicas de neuroimagem permite investigar unidades funcionais
em indivíduos normais durante o desempenho de tarefas que envolvem o engajamento dos
diferentes módulos de memória, trazendo informações mais precisas sobre as regiões e
processos cerebrais envolvidos nessas funções. No entanto, muitas das evidências
relevantes para o desenvolvimento de modelos de memória - correlações entre funções e
módulos do sistema nervoso – derivaram de correlatos anatomofuncionais, isto é, estudos
envolvendo dificuldades de memória em pacientes com danos cerebrais identificáveis. A
partir de então foi possível chegar a definições de memória e modelos baseados na dupla
dissociação entre memória de curta e longa duração. Inclusive os conceitos de dissociações
entre os sistemas particulares da memória de longa duração foram também amplamente
desenvolvidos.
Um estudo que muito contribuiu para o desenvolvimento e formalização dos modelos
de memória foi o caso do paciente H.M., descrito por Scoville e Milner (1957). Na ocasião, o
paciente sofria de epilepsia intratável. O foco epiléptico, que se situava no lobo temporal
medial (bilateralmente), foi removido cirurgicamente; isso resultou na remoção dos 2/3
anteriores do hipocampo e da amígdala, além de outras porções corticais. Após a remoção
das estruturas, H.M. apresentou um quadro de amnésia anterógrada (era incapaz de formar
novas memórias) e também retrógrada (eventos ocorridos pouco antes da cirurgia); porém,
neste último caso a amnésia era temporalmente graduada. O prejuízo cognitivo de H.M.
estava restrito à aquisição de memórias de longa duração; suas capacidades perceptuais se
mantiveram, assim como seu QI, sua personalidade e a memória de curta duração.
Mesmo apresentando alguns prejuízos de memória, H.M. ainda conseguia adquirir e
reter diversas informações. Por exemplo, aprendeu a ler palavras invertidas, como se
apresentadas por meio de um espelho e também novas habilidades motoras e cognitivas
(ver Helene e Xavier, 2007). O paciente apresentava um bom desempenho nessas tarefas.
Curiosamente, quando consultado sobre seu treinamento prévio, ele alegava nunca ter feito
isso.
46 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
Como dito anteriormente, o hipocampo atua em um processo de ativação repetitiva
de circuitos envolvidos na representação da informação, uma espécie de reverberação da
atividade neural que resulta no arquivamento de informação. Essa reverberação seria
essencial para o arquivamento das informações sobre “o que” ocorreu, mas não sobre
“como” desempenhar uma tarefa perceptomotora. Trazendo esse conceito para o caso do
paciente H.M, pode-se dizer que, embora o paciente seja capaz de adquirir uma habilidade
motora, ele não é capaz de se recordar “que” já a praticou. Em suma, a natureza da
informação “saber que” é diferente da natureza da informação sobre “saber como” (ver
Helene e Xavier, 2007).
Curiosamente, pacientes com doença de Parkinson (caracterizada por disfunções em
estruturas nervosas denominadas gânglios da base) possuem um quadro oposto ao dos
amnésicos (que, como visto, têm lesão no lobo temporal medial). Os pacientes com
disfunções nos gânglios da base exibem dificuldades na aquisição de habilidades motoras e
cognitivas, ao mesmo tempo em que são perfeitamente capazes de descrever verbalmente
as experiências vivenciadas nessas situações de teste. Neste contexto, pacientes
parkinsonianos exibem, por exemplo, prejuízo na aprendizagem da habilidade de leitura de
palavras invertidas.
Duplas dissociações, caracterizadas pelo prejuízo de desempenho em algumas
tarefas concomitantemente ao desempenho normal em outras tarefas, são apontadas como
evidência da existência de sistemas de memória distintos no sistema nervoso. Resultados
de estudos como do paciente H.M. e pacientes parkinsonianos sugerem a existência de
módulos de memória cujo funcionamento seria relativamente independente, embora possam
cooperar entre si.
Modelos de memória
Baseados em estudos envolvendo duplas dissociações, Cohen (1984) e Squire e
Zola-Morgan (1991) propuseram uma distinção para os sistemas de memória de longa
duração segundo a qual haveria uma memória declarativa (ou explícita), usualmente
prejudicada em pacientes amnésicos e preservada em pacientes cerebelares ou com
disfunções nos gânglios da base, e uma memória de procedimentos (ou implícita),
usualmente preservada nos pacientes amnésicos, mas prejudicada nos pacientes
cerebelares ou com danos nos gânglios da base (Fig. 4). Em outras palavras, memórias que
atualmente são denominadas memórias implícitas correspondem ao “saber como” (o que faz
bastante sentido, pois é muito difícil declarar como se anda de bicicleta) e “saber que” são
denominadas memórias explícitas.
47 www.ib.usp.br/labnec
Paciente
H.M.
Aquisição de
novos fatos e
eventos (saber
QUE)
Prejuízo
Aquisição de
novas
habilidades
(saber COMO)
Preservada
Pacientes com
doença de
Parkinson
Preservada
Prejuízo
Figura 4 – Esquema da dupla-dissociação entre funções e áreas envolvendo os
sistemas de memória de longa duração. Modificado de Helene e Xavier, 2007.
Memória de longa duração
A memória de longa duração se refere à retenção de informações por prolongados
períodos de tempo. Sendo assim, ela pode ser dividida em dois tipos (ou módulos): memória
explícita e memória implícita (Fig. 5). Tanto no caso das memórias explícitas como no caso
das implícitas, o arquivamento de informações envolveria alterações sinápticas, como já
descritas; porém, em cada caso, elas ocorreriam em diferentes regiões do sistema nervoso
com diferentes regras de funcionamento.
Figura 5 - Taxonomia dos sistemas de memória de longa duração. Modificado de Helene e Xavier, 2007. A memória explícita (ou declarativa) caracteriza a retenção de experiências sobre
fatos e eventos passados e é passível de relato verbal, ou seja, possui um acesso
48 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
consciente. Além disso, o arquivamento de informações pode se dar por associações
arbitrárias que podem formar-se mesmo após uma única experiência.
A memória implícita (ou de procedimentos) se expressa pelo desempenho habilidoso
das atividades previamente treinadas. Sua aquisição é gradual e dependente de treino,
ocorre de forma cumulativa. O conhecimento contido neste tipo de memória manifesta-se
pela ativação das estruturas nervosas envolvidas no processo de aquisição.
Memória operacional
Baddeley e Hitch (1974) conceberam um modelo de memória denominado "memória
operacional". Tal modelo refere-se a um arquivamento temporário e gerenciamento de
informações para o desempenho de uma diversidade de tarefas cognitivas. Segundo os
autores, memória operacional compreende um sistema de controle de atenção, a central
executiva, auxiliado por dois sistemas de suporte responsáveis pelo arquivamento
temporário e manipulação de informações, um de natureza vísuo-espacial e outro de
natureza fonológica.
Posteriormente, para lidar com a associação entre as informações mantidas nesses
sistemas de apoio e promover sua integração com informações da memória de longa
duração, Baddeley inseriu um quarto componente no modelo, denominado de retentor
episódico, que corresponderia a um sistema de capacidade limitada no qual a informação
evocada da memória declarativa tornar-se-ia consciente. A central executiva proporcionaria
a conexão entre os sistemas de suporte e a memória de longa duração e seria o
responsável pela seleção de estratégias e planos; sua atividade estaria relacionada ao
funcionamento do lobo frontal, que teria a função de supervisionar informações a serem
codificadas, armazenadas e evocadas concomitantemente ao seu ingresso no sistema (Fig.
6).
Figura 6 - Modelo de
memória operacional: três
componentes
propostos
inicialmente por Baddeley e
Hitch (1974). A área central
executiva se refere ao
componente de gerenciamento atencional (a central
executiva), enquanto as
áreas laterais da figura
representam as alças de
manutenção de informações
por curto período de tempo
(adaptado de Baddeley,
1982). 49 www.ib.usp.br/labnec
Ainda sim, a memória operacional estaria ligada ao desempenho de uma grande
variedade de funções cognitivas, incluindo raciocínio lógico, resolução de problemas,
imagética (relacionado ao treinamento imaginativo) e compreensão de linguagem.
Sistemas de memórias e seus aspectos evolutivos
Em conclusão, a evolução filogenética teria atuado na seleção de sistemas neurais
capazes de modificar-se gradualmente pelo desempenho de ações repetitivas (o exemplo
mais típico seria o caso de habilidades motoras e perceptuais) de sistemas capazes de
arquivar informações depois de uma única experiência, e de sistemas capazes de reter
informações temporariamente, enquanto úteis. É provável que a seleção desses sistemas,
com propriedades distintas, esteja relacionada ao fato de que memórias são especializações
adaptativas que proporcionam vantagens seletivas para a solução de determinados tipos de
problema; as propriedades que tornam um sistema eficiente para a solução de determinados
tipos de problema (e.g., aquisição após uma única experiência de treino) o tornam
incompatíveis com a solução de um problema de natureza diversa (e.g., aquisição de
conhecimento pela mudança cumulativa e gradual de experiências). Assim, do ponto de
vista evolutivo, a organização do sistema nervoso, inclusive dos diferentes módulos de
memória, teria derivado da interação do organismo com demandas ambientais específicas,
resultando em especializações adaptativas que permitem ao organismo lidar com problemas
específicos.
50 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
Tomada de Decisões
Camile Maria Costa Corrêa
Laboratório de Neurociências e Comportamento
[email protected]
A pesquisa sobre tomada de decisões é a área da neurociência pela qual se
desenvolvem métodos voltados à compreensão dos processos neurais responsáveis pelas
escolhas. Isso aguça a curiosidade na busca por respostas aos processos subjacentes aos
nossos julgamentos e ações.
As decisões estão constantemente presentes em nossas vidas, de forma mais ou
menos explícita. Decidimo-nos, a todo instante, entre alternativas banais, cotidianas,
chegando a juízos mais complexos. Embora percebamos o quanto esses atos são
registradamente humanos, eles também se fazem presentes no repertório comportamental
de outros animais.
Decisões são consideradas escolhas baseadas em propósitos; ações orientadas
para alcançar determinado objetivo. Para isso, uma entre muitas alternativas de ações
possíveis é escolhida quando do confronto com um problema, a fim de resolvê-lo.
Muitas situações podem ser problematizadas para apreendermos o fenômeno da
tomada de decisão. Que roupa vestir antes de sair de casa? Descer pela escada ou pelo
elevador? Onde almoçar hoje? Comprar ou não uma bicicleta agora? Onde prestar
mestrado? Como julgar inocentes e condenáveis? Que opinião formar sobre questões
polêmicas? Algumas escolhas são mais simples que outras, ou porque têm sua resolução
mais rápida, ou por envolver menos alternativas a serem processadas. Na problematização
dessas questões, podem-se levar em conta diferentes critérios.
Entretanto, por mais que se tente discriminar quais são as estratégias utilizadas,
sabemos que nossas decisões provavelmente não sofrem o mesmo tipo de processamento
a cada instante e a todo contexto. Muitas das nossas escolhas são automáticas,
emocionais, inconscientes. Muitas vezes somos impulsivos ou completamente indecisos
frente a dilemas. Tanto que não é difícil encontrar pessoas que reportam dificuldades para
decidir, e há mesmo casos de pacientes incapazes de tomar decisões. Percebemos, então,
que se podem propor vários métodos para compreender esse fenômeno.Com efeito, o tema
da decisão acompanha as produções da humanidade tanto nas artes como na filosofia e
ciência, chegando às abordagens clínicas e experimentais do tema. A psicologia cognitiva, a
partir de 1950, vem integrando modelos tradicionais de tomada de decisões em humanos,
aproximando-se de modelos de processamento de informações (Sternberg, 2000). A
pesquisa básica da chamada “decision making” vem tomando corpo com a realização de
experimentos associados a estudos da neurobiologia (vias de neurotransmissores,
51 www.ib.usp.br/labnec
correlatos anátomo-funcionais), principalmente em ratos e primatas não humanos, lançando
mão de modelos matemáticos e probabilísticos para a análise dos dados (e.g., Körding e
Wolpert, 2006; Kepecs, 2008). Recentemente, pesquisas clínicas têm se debruçado sobre o
tema e suas correlações com o desempenho em tarefas que envolvem decisão
(distratibilidade, perseveração) em populações com alterações funcionais em áreas
específicas do sistema nervoso e em grupos de pacientes psiquiátricos (e.g., Bechara, 2001;
2004; Cavedini, 2002; Schurman, 2005).
A neurociência, por sua vez, vem desenvolvendo métodos para avaliar a contribuição
da cognição, emoção, atenção e memória, além de outras variáveis, partindo do
pressuposto de que a decisão não é uma simples escolha entre alternativas, mas um
processo dependente da experiência do indivíduo que decide e de sua capacidade de
identificar os principais fatores da situação na qual se deve decidir. Esses estudos têm
permitido concluir que o processo, tanto em humanos quanto em outros animais, é
modulado não só pelo contexto ambiental, mas também pelo estado do organismo.
Entender como decidimos é perguntar como processamos as informações, de que
forma atribuímos diferentes valores a elas e como optamos entre alternativas, direcionando
as ações pelas quais somos, em princípio, responsáveis. Para que se escolha, então, é
necessário não só um funcionamento íntegro e orquestrado do sistema nervoso, como
também a capacidade de selecionar informações e estímulos aos quais o organismo é
exposto e aos quais deve reagir - optando. Nesse processo contínuo, sugere-se que tanto
variáveis extrínsecas quanto intrínsecas, conscientes ou não, concorrem para a tomada de
decisão.
Dilemas e Estratégias
A teoria dos jogos, desenvolvida por Nash, estuda situações estratégicas onde
jogadores escolhem diferentes ações na tentativa de melhorar seu retorno. Um exemplo
envolvendo estratégias mútuas é ilustrado pelo dilema do prisioneiro, em que dois suspeitos,
A e B, são presos pela polícia. A polícia tem provas insuficientes para condená-los, mas,
separando os prisioneiros, oferece a ambos o mesmo acordo: se um dos prisioneiros,
confessando, testemunhar contra o outro e esse outro permanecer em silêncio, o que
confessou sai livre enquanto o cúmplice silencioso cumpre 10 anos de sentença. Se ambos
ficarem em silêncio, a polícia só pode condená-los a 6 meses de cadeia cada um. Se ambos
traírem o comparsa, cada um leva 5 anos de cadeia. Cada prisioneiro faz a sua decisão sem
saber que decisão o outro vai tomar, e nenhum tem certeza da decisão do outro. A questão
que o dilema propõe é: o que vai acontecer? Como o prisioneiro vai reagir? Abaixo, uma
matriz de ganhos do dilema do prisioneiro:
52 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
Tabela 1 – Dilema do prisioneiro
Prisioneiro B fica em silêncio
Prisioneiro B confessa
Prisioneiro A fica em silêncio
6 meses cada
Prisioneiro A: 10 anos
Prisioneiro B: liberdade
Prisioneiro A confessa
Prisioneiro A: liberdade
Prisioneiro B: 10 anos
5 anos cada
Em linhas gerais, não importa os valores das penas em si, mas o cálculo das
vantagens de uma decisão cujas consequências estão atreladas às decisões de outros
agentes, onde a confiança e a traição fazem parte da estratégia em jogo. Dilemas que
envolvem estratégias mútuas em grupos sociais também podem ser simulados: Robert
Axelrod estudou uma extensão do dilema do prisioneiro, denominada dilema do prisioneiro
iterado (DPI), em que esse problema é jogado repetidas vezes. Num torneio de
programação, os participantes deveriam escolher uma e outra vez a sua estratégia mútua, e
tinham memória dos seus encontros prévios (encontros com outros programas em que
deveriam optar por ser altruístas ou egoístas). Os programas que participaram variavam
amplamente a complexidade do algoritmo: hostilidade inicial, capacidade de perdão e
similares.
Axelrod descobriu que, durante a repetição dos encontros com muitos jogadores,
cada um com estratégias distintas, as estratégias "egoístas" tendiam a ser piores a longo
prazo, enquanto que as estratégias "altruístas" eram melhores, julgando-as unicamente com
respeito ao interesse próprio. Nesse torneio, a melhor estratégia determinista foi a de “olho
por olho” ("tit for tat"), desenvolvida e apresentada no torneio por Anatol Rapoport: o mais
simples de todos os programas apresentados, contendo apenas quatro linhas de BASIC, foi
o que ganhou o concurso. A estratégia consistiu em cooperar na primeira interação do jogo,
e, depois disso, escolher aquilo que o oponente escolhera na rodada anterior. Esse padrão
de decisão pôde demonstrar, por exemplo, um possível mecanismo que explicasse de que
forma é possível evoluir um comportamento altruísta a partir de mecanismos puramente
egoístas na seleção natural.
Origens
Campos, Santos e Xavier (1997) defendem que regularidades ambientais presentes
ao longo da evolução das espécies possibilitaram a seleção de sistemas que assim
tornaram-se adaptados e otimizados para esses ambientes, possibilitando a emissão de
respostas comportamentais antecipatórias. Porém, a complexidade ambiental teria
contribuído também para a seleção de mecanismos mais flexíveis, envolvendo o acúmulo de
informações sobre o ambiente, que permitiram a extrapolação, com base nessas
informações arquivadas, para a solução de novas demandas ambientais. Esses
53 www.ib.usp.br/labnec
mecanismos seriam adaptativos, pois permitem lidar com circunstâncias novas e
inesperadas. Mesmo bactérias, como a E. coli, apresentam sistemas sensórios voltados à
detecção de nutrientes, fontes de energia, toxinas e capacidade para armazenar e avaliar as
informações vindas desses receptores. O resultado final da integração sensorial no
comportamento de busca por nutrientes é a decisão, por exemplo, de continuar nadando
numa mesma direção ou mudar de rumo. Assim, características fundamentais de integração
cognitiva, tais como integração sensorial, memória, tomada de decisão e controle
comportamental podem ser encontradas em organismos muito simples. (Allman, 1999).
Para além disso, a flexibilidade comportamental de diferentes grupos de animais
parece estar relacionada com a quantidade relativa de tecido nervoso (proporcionalmente ao
tamanho corpóreo). Em vertebrados, as porções anteriores do sistema nervoso,
relacionadas à manipulação e integração de informações, memória, antecipação, atenção e
produção de respostas, variam enormemente, sendo maiores nos primatas, particularmente
em seres humanos (Campos, Santos e Xavier, 1997). Apesar de estar presente em todas as
espécies de mamíferos, o córtex frontal sofreu grande expansão ao longo da evolução dos
primatas, especialmente nas regiões mais anteriores, contribuindo para o grande
desenvolvimento de capacidades cognitivas (Gazzaniga e coll., 2006). Não surpreende,
portanto, seu envolvimento em processos de tomada de decisões. A função dessa classe de
comportamentos se estabelece na relação direta de nossas ações, pois são elas as que são
selecionadas ao agirmos sobre o meio de forma adaptativa.
Interação
Observemos um quadro em que o organismo se relaciona com eventos exteriores a
ele: as informações vindas do meio devem ser processadas de forma a serem traduzidas
em códigos reconhecíveis pelo sistema nervoso. Essa conversão de diferentes formas de
energia, incluindo a química, a térmica, a mecânica, a sonora, entre outras, em energia
elétrica, passível de ser traduzida e conduzida na forma de potenciais de ação, é
denominada transdução. Ao longo desse processamento, em que estruturas nervosas são
funcional e hierarquicamente mobilizadas, identificam-se interações entre percepções –
tanto internas quanto externas; o acesso e mesmo alterações em registros de memória; a
confluência de motivações e a construção de um programa que habilite o sujeito a fazer uma
nova transdução, convertendo agora a informação processada em planos de ação
direcionados ao meio. Na modulação desse processo, influenciariam estados imunitários,
emocionais e atencionais, diferenciando nossas escolhas e imprimindo nossa personalidade
a elas.
Percepção, emoção, atenção e memória, entre outras funções cognitivas, interferem
nesse processo, dependente da experiência prévia do indivíduo que decide, de sua
54 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
capacidade de identificar os principais fatores da situação na qual se deve decidir, de quais
desses fatores são ressaltados e valorizados, além da afetividade relacionada à decisão.
Revisamos, na sequência, a contribuição relativa de processos cognitivos envolvidos nas
nossas escolhas.
Percepção temporal
Demandas por escolhas frequentemente envolvem a ponderação entre vantagens e
riscos assumidos em curto prazo frente a expectativas de longo prazo. Diariamente, e isto
se intensifica nas manhãs de inverno, ao termos o sono interrompido pelo despertador,
iniciamos uma luta travada entre o prazer de continuar no estado de sonolência e a
obrigação de aumentarmos a vigília, iniciando os afazeres pelos quais assumimos
responsabilidade. De forma semelhante, pessoas que querem entrar em dieta encontram
dificuldades para iniciá-la e mantê-la; as tentativas de parar de fumar podem ser frustrantes;
o impulso por comprar imediatamente pode conflitar com planejamentos de economia a
médio e longo prazo. Interessantemente, a percepção temporal que os sujeitos têm na hora
de avaliar alternativas pode fornecer pistas interessantes na pesquisa sobre como
decidimos, apontando diretamente para comportamentos, práticas de consumo, economia e
política.
Processos inconscientes
Ao estabelecermos que uma decisão se inicia com uma percepção, seja de
estímulos, seja de variáveis ou de probabilidades, muitas vezes não se tem acesso a tais
percepções em nível consciente. Num artigo de 1987, Kihlstrom apontou o impacto de
estruturas e processos inconscientes na experiência consciente do indivíduo, reportando-se
a von Helmholtz, para quem a percepção consciente era produto de inferências
inconscientes baseadas no conhecimento do mundo e de experiências prévias. Para
Kihlstrom, processos perceptivos são capazes de ativar nós da rede neuronal, os quais
codificariam representações mentais de estímulos externos, ou mesmo por mecanismos
internos de pensamento.
Se, no sistema nervoso, subsistemas podem operar independentemente e sob
regras diversas, apenas alguns módulos seriam acessíveis à consciência; portanto,
potencialmente sob domínio de controle voluntário. Há situações em que tanto o número de
processos ativos simultâneos quanto a velocidade com que a informação é trocada podem
exceder a capacidade de atenção consciente. Os processos inconscientes seriam rápidos e
paralelos, enquanto que o processamento consciente seria lento e serial.
Do ponto de vista experimental, portanto, tem-se conhecimento sobre os objetivos e
condições dos procedimentos, bem como dos produtos de suas execuções, mas não se tem
55 www.ib.usp.br/labnec
acesso às operações por elas mesmas. De forma análoga, o conhecimento sobre as
operações que levam às decisões não seriam acessíveis à consciência. E aí o papel da
experimentação é fundamental, na tentativa de desvelar esses processos.
Atenção
A orientação da atenção pode ser considerada um processo decisório. Desde o que
se considerem os níveis mais elementares de tomada de decisão até as escolhas mais
complexas, o ato de decidir envolve o engajamento, consciente ou não, de um foco
atencional. Decidir, em outras palavras, envolve seleção e processamento preferencial de
alguns estímulos em detrimento de outros. Os processos que levam a esse processamento
dependem não apenas da história prévia do sistema selecionador, isto é, suas memórias,
como também de expectativas geradas com base em memórias sobre regularidades
passadas e planos de ação.
Memória
Com o acúmulo de registros sobre ocorrências anteriores, memórias no sentido
amplo da palavra, e a identificação de regularidades na ocorrência desses eventos, o
sistema nervoso passa a gerar previsões (probabilísticas) sobre o ambiente. Xavier, Saito e
Stein (1991) sugeriram que a antecipação, com base na identificação de regularidades
ambientais passadas, permite reagir mais prontamente à estimulação esperada, pois o
organismo direciona a atenção para os setores do ambiente que são relevantes.
A memória operacional, um tipo de memória temporária, contém representações
ativas do organismo em seu ambiente atual, com seus objetivos em curso e com estruturas
de conhecimentos explícitos (declarativos) já existentes, ativadas por entradas perceptivas
ou por outros processos dos quais não se tem consciência.
Helene e Xavier (2007) postularam que, como produto final de seu funcionamento, a
memória pode ser vista como base fundadora dos processos de formação, não somente de
comportamentos estereotipados, mas também de um vasto conjunto de comportamentos
adaptativos, dentre eles, podemos dizer, a decisão.
Controle executivo
Ao planejar, ao agir antecipatoriamente, o sistema nervoso pode tanto gerar ações
que levem aos resultados desejados como atuar no sentido de selecionar determinados
tipos de informação para processamento adicional (direcionamento da atenção). Decidir
envolve não só a percepção de regularidades passadas, como a lembrança dos planos de
ação, a prever os efeitos prováveis da escolha, considerando todos os reflexos possíveis
56 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
que ela pode causar no curso do tempo. O conceito de controle executivo ilustra, então, a
capacidade que temos de planejar, gerenciar ações, modular o comportamento e criticar
processos. Ao decidirmos, ponderamos de forma mais ou menos flexível, transitando entre
alternativas que se nos apresentam, fazendo com que nossas escolhas situem-se entre a
impulsividade e a perseveração.
Estudos clínicos
O desempenho de pacientes neurológicos ajuda a direcionar as investigações sobre
o processo de tomada de decisão em humanos. Bechara e col. (1997) questionaram a
premissa segundo a qual decidir de forma vantajosa numa situação complexa requer
racionalização de conhecimento declarativo. Esse grupo investigou a possibilidade de que a
racionalização manifesta fosse precedida por uma etapa não consciente, cujos sistemas
neurais seriam diferentes dos que suportam o conhecimento declarativo. Para isso,
participantes normais e pacientes com lesão prefrontal (e deficits na tomada de decisão)
realizaram o Yowa Gambling Task, um simulador de tomada de decisões, que envolve
escolhas monetárias, permitindo classificar o comportamento de decisão do indivíduo em
termos de aversão ou busca pelo risco. A tarefa envolve a escolha de uma carta de um
dentre quatro baralhos (cinco blocos de vinte jogadas cada). Cada um desses trabalhos
inclui uma longa série de ganhos e perdas. A partir de um processo de aprendizagem, os
participantes criam padrões de probabilidade e inferem quais baralhos são vantajosos e
quais não são. Eles devem desenvolver o conhecimento de quais baralhos são arriscados e
quais são lucrativos em longo prazo (Schneider e Parente, 2006).
No estudo do grupo de Bechara, pessoas normais começaram a escolher de forma
vantajosa antes que percebessem qual era a melhor estratégia, enquanto pacientes com
disfunções prefrontais continuaram a escolher de forma desvantajosa, mesmo depois de
terem conhecimento de qual era a estratégia correta. Além disso, os sujeitos normais
começaram a gerar respostas antecipadas de aumento da condutância de pele frente a uma
escolha arriscada, sendo que os pacientes nunca chegaram a desenvolver essas respostas
antecipatórias, embora alguns tenham eventualmente percebido quais escolhas eram
arriscadas.
Os resultados sugerem que, em indivíduos normais, vieses não conscientes são
capazes de guiar comportamentos antes mesmo que o conhecimento consciente o faça.
Sem a influência de tais tendenciosidades o conhecimento manifesto pode ser insuficiente
para assegurar comportamentos vantajosos. Assim, uma vez que comportamentos de maior
risco foram encontrados na amostra clínica (lesão frontal) e não no grupo controle, tornou-se
possível identificar níveis progressivos tanto de desempenho na tarefa como de acesso
57 www.ib.usp.br/labnec
explícito ao conteúdo da informação. Os autores sugerem, a partir disso, uma relativa
dissociação entre desempenho e consciência.
Ainda no âmbito das pesquisas clínicas, apontam-se correlações entre o
desempenho de tarefas que envolvem tomada de decisões em humanos e um aumento da
atividade em regiões definidas do sistema nervoso, como revelado por estudos de
neuroimageamento funcional. Essas regiões parecem estar afetadas em pessoas com
disfunções patológicas nos processos de tomada de decisões, tais como pacientes com
lesões frontais, pacientes esquizofrênicos e jogadores compulsivos. (Bechara, 2001; 2004;
Cavedini, 2002; Schurman, 2005).
Livre arbítrio e determinismo
O estudo da tomada de decisões envolve, em última análise, não só a compreensão
de diversas funções cognitivas, como permite pensarmos questões éticas sobre como os
seres humanos assumem responsabilidade sobre a própria vida.
O livre arbítrio é a crença filosófica que defende que as escolhas e julgamentos
morais possam ser realizados de forma autônoma, livre e isenta de influências. Esse
conceito, ainda que aqui delineado de maneira geral, leva a importantes implicações
religiosas, morais, psicológicas e científicas. A doutrina oposta a essa é a do determinismo
psíquico, que afirma que todos os eventos, incluindo as vontades e escolhas humanas, são
causados diretamente por acontecimentos anteriores, o que abala a noção de liberdade de
escolha.
Voltados a esse tema, pesquisadores ousaram debater o tema da liberdade de
escolha de forma experimental. Por exemplo, Soon e col., num artigo amplamente divulgado
(2008), questionam em que extensão decisões podem ser subjetivamente consideradas
como “livres” se são determinadas por atividade cerebral detectada anteriormente no tempo;
os autores relatam que o resultado de uma decisão pode ser codificada na atividade
cerebral dos córtices prefrontal e parietal até 10 ms antes do acesso consciente à decisão.
Esse atraso, presumivelmente, refletiria a operação de uma rede nervosa de controle que
inicia o preparo da decisão em curso mesmo antes de qualquer acesso explícito ao seu
conteúdo.
Independente de nossas posições pessoais sobre o assunto, os resultados
experimentais influenciam no desenvolvimento dos métodos de investigação posteriores,
ajudando a determinar os passos seguintes da investigação sobre o tema.
Mecanismos decisórios fazem com que sejamos capazes de elaborar juízos ao
tomarmos contato com problemas a fim de resolvê-los. Essa capacidade aproxima o estudo
do processo da tomada de decisão a outras funções cognitivas, na medida em que são
58 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
necessários planejamento, gerenciamento de ações, modulação do comportamento, crítica
e flexibilidade.
Investigar a tomada de decisões é presenciar o momento em que as contingências
passam a ser atualizadas pelas ações do sujeito, que é capaz de fazer interfaces entre
eventos externos e internos e, assim, aprende a prever as consequências de seus
comportamentos.
59 www.ib.usp.br/labnec
Emoção
Diego de Carvalho
Laboratório de Neurociências e Comportamento
[email protected]
Introdução
A capacidade de racionalizar, sentir e se emocionar torna os seres humanos únicos
em relação às outras espécies e dentro da sociedade. A emoção está presente
permanentemente no nosso dia-a-dia e, apesar dessa existência corriqueira, é muito difícil
definir através de palavras o que sentimos. É possível que essa dificuldade resida na
aceitação do fato de que o controle das emoções, algo tão imaterial, esteja ligado a algo tão
físico como o cérebro e não a um fator externo ao corpo, como propôs Descartes em sua
teoria da mente e do corpo. Além disso, talvez até pela dificuldade de ligar o cérebro às
emoções, a neurociência negligenciou os estudos dos aspectos emocionais do
comportamento por um longo tempo.
Em 1848 um incidente trágico tornou claro que as emoções, a personalidade e a
vivência em sociedade são regidas por funções neurais. Phineas Gage, um operário de uma
estrada de ferro, estava dinamitando algumas rochas e ao pressionar a pólvora em um
buraco com uma barra de ferro iniciou o processo de detonação. A barra trespassou a face
de Gage e saiu pela testa. Surpreendentemente, ele continuou lúcido e foi declarado curado
em poucos dias, porém sua capacidade de se emocionar e tomar decisões foi comprometida
após o incidente, sendo o primeiro relato de que uma lesão nos lobos frontais (confirmada
posteriormente) pode alterar a personalidade de uma pessoa.
Após o caso Gage, visto que as funções comportamentais relacionadas à emoção
eram exercidas e/ou controladas por algumas regiões do encéfalo, a neurociência teve que
vencer alguns obstáculos para incorporar a emoção ao estudo científico: Como dimensionar
a emoção em pessoas com vivências diferentes, ou mesmo, como dimensionar o que está
se sentindo? Como gerar sentimentos que sejam próximos aos espontâneos? Como criar
um modelo palpável de manipulação experimental em humanos e animais?
Estas
perguntas aparentemente geraram, além de dúvida, um desconforto nos neurocientistas que
culminou em uma demora na inserção das emoções ao plano científico.
Embora todas estas perguntas tenham alta relevância científica, a maior dificuldade
ainda está na real definição de emoção. Uma noção generalista é de que a emoção é
composta por três fatores principais: um componente sentimental, uma resposta
comportamental e as adequações fisiológicas pertinentes. Esta noção, além de generalista,
descarta os componentes psicológicos que, no caso de humanos, altera significativamente o
modo que a emoção se processará. Contudo, é possível concluir que um dado sentimento
60 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
gerado por fatores exógenos ou endógenos, levará a alguma resposta motora, seja esta
estereotipada, isto é, simples e de natureza reflexa ou complexa de natureza volitiva; e
ainda terá os ajustes fisiológicos pertinentes, como a liberação de hormônios, dada a
intensidade do sentimento e diferindo quanto às situações e personalidade dos indivíduos.
Em casos normais as respostas emocionais devem ser de caráter imediato e transitório,
porém, em casos de desordens afetivas ocorrem respostas prolongadas.
Esta definição de emoção, ainda que generalista, permite entender o significado da
emoção, mas não define cada experiência emocional isoladamente. Por exemplo, estar feliz,
triste, furioso, entediado, desapontado, excitado, chateado, com medo, apaixonado,
exaltado ou satisfeito definem vários estados emocionais, sejam positivos ou negativos,
fortes ou fracos. É possível, então, notar que alguns destes exemplos são apenas variações
quanto à intensidade do sentimento: alegre, satisfeito, excitado são apenas termos que
exprimem diferentes amplitudes de felicidade. Então, nota-se que há dois novos
componentes das emoções: (1) o fator intensidade ou amplitude e; (2) há várias
denominações dependentes da intensidade de uma emoção básica.
Desde a expressão das emoções nos homens e nos animais de Charles Darwin
(1872), os cientistas vêm tentando definir um conjunto finito de emoções primárias. Em
estudos com expressões faciais foi descoberto que independente do local, cultura e etnia
algumas emoções tem respostas comportamentais muito semelhantes (Ekman e Frieser,
1971). Baseado nestes estudos foi proposto que existem 6 tipos de expressões faciais
básicas humanas que denotam emoções. São elas: raiva, medo, aborrecimento, felicidade,
tristeza e surpresa. A definição de emoções básicas permite diferentes manipulações
experimentais para investigação dos sistemas neurais envolvidos nas emoções e diferentes
propostas para quantificá-las. Chegando novamente à pergunta: como dimensionar e
quantificar a emoção em pessoas com vivências diferentes?
Emoção, Cognição e Comportamento
Aparentemente a correlação entre cérebro e emoções está, hoje, bem estabelecida.
Entretanto, a inserção dos processos emocionais dentro de estudos cognitivos soa um tanto
quanto abstrata. Algumas dualidades parecem perdurar na história humana, como era, por
exemplo, na teoria de alma e corpo de Descartes, a alma regeria os aspectos emocionais e
o corpo regeria a razão. Ainda hoje razão e emoção parecem palavras antônimas e da
mesma forma cognição e emoção podem parecer contrastantes. Mas novos estudos têm
demonstrado que a emoção é um fator que influencia, modula e pode ser até mesmo
preponderante em sistemas classicamente cognitivos como na memória e aprendizado
(Immordino-Yang e Damásio, 2007). Ainda neste mérito, Antônio Damásio, em o “Erro de
61 www.ib.usp.br/labnec
Descartes” (1994), propõe que a razão é consequência da avaliação emocional de um ato,
portanto tratar emoção e razão como domínios totalmente isolados seria um erro.
Mesmo estabelecendo que a emoção seja adequada aos estudos cognitivos, a
grande variedade de personalidades existentes na sociedade, mesmo em um grupo restrito,
torna o estudo em laboratório altamente desafiador; agora imagine um estudo global da
espécie humana, no qual além de personalidades diferentes, diferem também as culturas e
os valores. Portanto, a tarefa de manipular e estudar a emoção em situações controladas
exige algumas técnicas, que vem se desenvolvendo ao longo dos anos.
Os conceitos de que uma emoção pode ser positiva ou negativa parecem estar bem
claros. Por exemplo, é de comum acordo que felicidade é uma experiência positiva e tristeza
negativa. Também parece claro que uma emoção pode ter diferentes intensidades: A
felicidade de achar uma moeda na rua não é de mesma amplitude que ganhar na loteria.
Então, a avaliação da emoção em laboratório visa, basicamente, manipular a amplitude e o
contexto emocional a fim de elucidar como são deflagradas as ações subsequentes ao
estímulo, como é o processamento neural por detrás dos sentimentos e como isso pode
influenciar em outros processos cognitivos. Em termos gerais, em experimentos com
emoção, há uma tentativa de indução de um estado emocional no sujeito, seja por indução,
em que se pede que o indivíduo tente evocar um estado emocional em particular; por
métodos de recompensa e punição, nos quais um estímulo motivacional é a chave do teste,
seja ele reforçador ou aversivo; ou ainda pela apresentação de estímulos que evocam
emoções, como a apresentação de algumas cenas carregadas de sentidos emocionais. Em
1995, Lang e colaboradores reuniram uma coleção de imagens que evocam uma série de
respostas emocionais. Os autores pediram que várias pessoas de diversas etnias
classificassem estas figuras por valência (intensidade da emoção gerada) e por alerta
(quanto a imagem os deixou em alerta). Este trabalho acabou virando um sistema
internacional de figuras afetivas (IAPS – International Affective Pictures System) que é
usado como padrão em testes comportamentais (Figura 1).
Figura 1 – À esquerda exemplo de imagem de contexto emocional positivo, e à direita, negativo. Retirado de:
IAPS, 1995.
Os métodos de quantificação de impacto emocional que gerado por diferente
técnicas podem ser avaliados através de questionários ou através de testes subsequentes,
62 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
como por exemplo, a escolha entre dois objetos, em que a preferência será a medida
observada. Em modelos animais, algumas respostas comportamentais podem ser
quantificadas (ver adiante) ou ainda é possível avaliar algumas respostas autonômicas,
como pressão arterial, resposta de condutância da pele e frequência cardíaca.
A resposta de condutância da pele pode ser utilizada tanto em humanos como em
animais. As propriedades elétricas da pele são medidas através de eletrodos durante a
realização de uma tarefa ou apresentação de um estímulo. Em condições de estresse a
condutividade aumenta significativamente. Esta avaliação foi por muito tempo a base do
detector de mentiras.
Os processos emocionais podem ainda interferir na realização de algumas tarefas
que exigem outros processos cognitivos, como tomada de decisão, memória, aprendizado e
atenção. Em uma determinada tarefa o fator emocional pode influenciar de forma a inibir ou
facilitar o desempenho. Por exemplo, utilizando o IAPS os indivíduos tendem a lembrar mais
de imagens de caráter negativo quando solicitados a lembrar de algumas imagens que lhes
foram apresentadas (Pratto e Jonh, 1991).
Neurobiologia das Emoções
Ao longo dos anos diversas teorias foram construídas a fim de explicar como as
emoções se processavam no sistema nervoso. Uma das primeiras teorias é a de James e
Lange datada do início do século XIX, quando pouco era sabido das interações dos eventos
neurais e comportamento. Os autores postularam que a emoção decorria após alguns
eventos fisiológicos, ou seja, o comportamento emocional se dava após a percepção de
alterações fisiológicas ocorridas no organismo. Então poderia se supor com esta teoria que
a tristeza decorre por causa do choro e não o contrário.
Em seguida Cannon e Bard (1928) refutaram a teoria de James-Lange e propuseram
que a resposta emocional estava ligada a eventos no sistema nervoso central, mais
precisamente no tálamo e hipotálamo. Entretanto, o conhecimento atual permite avaliar que
faltam evidências de participações talâmicas nas emoções.
Enfim, Papez, em 1937, teorizou que haveria todo um sistema relacionado com o
processamento da emoção. Este sistema ficou conhecido anos mais tarde como sistema
límbico; após algumas alterações da teoria inicial, foram atribuídas as participações do
hipotálamo, hipocampo, tálamo anterior, giro do cíngulo, amígdala e o córtex orbitofrontal. A
teoria inicial de Papez foi atualizada algumas vezes baseada em novos experimentos,
atualmente sabe-se que muitas estruturas límbicas participam de fato do processamento
emocional. Entretanto nem todas as descritas como participantes iniciais do sistema tem
participação comprovada. Apesar de existir toda uma circuitaria neural que compreende
63 www.ib.usp.br/labnec
diversas estruturas e mediadores, duas estruturas têm se destacado particularmente: A
amígdala e o córtex orbitofrontal.
Voltando ao Phineas Gage, anos após sua morte, o crânio foi exumado e estudado
por técnicas de formação e recomposição computadorizada do acidente (Fig. 2). Ficou claro
que era uma lesão frontal, a qual abrangia o córtex orbitofrontal. Esta estrutura não fazia,
então, parte do sistema límbico descrito por Papez, mas desempenha papel fundamental
nas emoções e tomadas de decisão. O real papel do córtex orbitofrontal nos processos
emocionais ainda não está bem estabelecido, porém sabe-se que tem grande importância
nas tomadas de decisões sejam elas de caráter emocional ou não.
As lesões frontais têm sido alvo de estudos ao longo dos anos. Um indivíduo com tal
lesão perde o senso de responsabilidade, capacidade de concentração, perda do poder
discriminativo ao tomar decisões e ainda tem prejuízo na expressão de estados afetivos. Um
exemplo disso era quando se praticava a lobotomia pré-frontal como tratamento para
algumas psicopatologias, o indivíduo perdia a capacidade de expressar e reconhecer
respostas emocionais e afetivas.
Figura 2 - Reconstituição computadorizada do crânio de Phineas Gage após o acidente.
Damásio (1994) demonstrou uma série de experimentos que comprovaram a
participação dos córtices frontais nos processos de tomada de decisão. O autor ainda
teorizou, a partir destes estudos com pacientes com lesões frontais, que o conhecimento de
características emocionais guia nossas escolhas racionais; por exemplo, um grupo era
convidado a escolher uma carta de duas pilhas distintas. A partir de certo ponto o indivíduo
aprendia que uma pilha continha cartas com premiações maiores, mas o risco de perda era
igualmente maior. Os indivíduos controles tendiam a evitar esta pilha, e quando decidiam
pegar uma carta desta, exibiam respostas emocionais autônomas claras, ao passo que os
pacientes com lesão frontal não evitavam a pilha arriscada e nem exibiam tais respostas
emocionais. Portanto, neste caso o papel das regiões frontais, sobretudo do córtex
64 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
orbitofrontal, parece ser o de basear as ações conforme as informações emocionais de cada
estímulo. Esta associação também parece ser altamente dependente de conexões das
áreas frontais e outras estruturas cerebrais, como a amígdala.
A amígdala desempenha papel fundamental na formação das emoções. Esta
estrutura em forma de amêndoa e seus variados núcleos têm ligações com outras
importantes estruturas cerebrais, como hipocampo, porção dorsal do tálamo, áreas préfrontais e núcleos do septo. Cada uma destas vias, e a própria amígdala, são importantes
mediadoras de comportamentos em resposta a um estímulo emocional. Ela é responsável
pela associação do estímulo emocional e a resposta comportamental, mediação da memória
associativa, alguns autores ainda a citam como responsável pelo comportamento agressivo
e mediador da consolidação de algumas memórias emocionais, podendo, por ação de
alguns hormônios liberados em situações de grande estresse, facilitar a retenção da
informação. Talvez por isso seja, na maioria das vezes, tão fácil lembrar episódios
carregados de contexto emocional. Um bom exemplo disso é que em testes que utilizam o
sistema de imagens afetivas a maioria das pessoas lembra mais fortemente de imagens de
contexto emocional negativo, com alta valência, em comparação às imagens neutras e
positivas (Hansen e Hansen, 1988; Pratto e Jonh , 1991; Huang e Luo, 2006)
As primeiras observações de que a amígdala tem participação importante nas
respostas emocionais datam de 1939 (Klüver e Bucy), em que macacos com lesões nesta
estrutura não reconheciam mais objetos antes carregados de grande contexto emocional. A
partir deste achado, muitos estudos têm sido realizados enfocando tal estrutura. Sabe-se
que a amígdala está envolvida em aprendizados que exigem associações, como por
exemplo, no condicionamento aversivo. Um rato que é colocado experimentalmente em uma
caixa onde um choque elétrico nas patas é pareado com um estímulo neutro, como uma luz,
exibirá após o pareamento uma resposta de medo, por exemplo, um sobressalto. Um animal
cujas funções amigdalares não estejam íntegras não responderá desta forma na
apresentação somente do estímulo neutro como ocorre normalmente em ratos sem dano
algum (Davis, 1992).
Mesmo nos aprendizados mais complexos, observados nos seres humanos, como
no medo instruído; no qual o evento aversivo não é vivido e sim contado por outra pessoa, a
amígdala é requerida. Isto porque ela tem conexões importantes com outras estruturas
neurais envolvidos fortemente com a memória, como por exemplo, o hipocampo. Então,
atuando em conjunto com outras estruturas, a amígdala é importante mediadora da
consolidação de eventos emocionais, modulando a intensidade e o impacto destas
memórias (McGaugh, 2004).
Lesões experimentais em determinados núcleos da amígdala ou desconexões de
algumas vias que ela faz parte têm sido praticadas para estudo da função da estrutura
65 www.ib.usp.br/labnec
neural e suas vias, bem como nas respostas emocionais. Diferentes vias e estruturas podem
participar da expressão das emoções, e grande parte dos achados até agora se devem, em
grande parte, a modelos animais.
Modelos animais
Os modelos animais permitem práticas de lesões precisas e desconexões funcionais
igualmente corretas para estudo da função neural, o que não é observado em casos
envolvendo humanos, nos quais a lesão acidental geralmente envolve múltiplas estruturas e
em geral é uma lesão difusa. Então, modelos animais podem ajudar a elucidar como a
emoção se processa no cérebro. Contudo, a linguagem ainda é uma barreira; como estudar
emoção se não conseguimos saber de fato se a emoção é pertinente ao animal? Para
responder esta pergunta e ainda ter ferramentas para responder outros questionamentos,
foram criados modelos animais para estudo do comportamento de respostas ao medo,
ansiedade e respostas à estímulos positivos e negativos. Testes como o labirinto em cruz
elevado (LCE), testes de esquiva, medo condicionado, e diversas tarefas que utilizam
reforços tem se mostrado muito eficazes em avaliar respostas emocionais, que são até de
certa forma subjetivas, principalmente para animais, como é o caso da ansiedade.
A ansiedade é um estado subjetivo de apreensão que se difere do medo por não
haver causa direta ou pelo menos aparente (File, 1992) provocando reações fisiológicas e
comportamentais. Tarefas comportamentais, como o LCE e o Campo Aberto (Fig. 3) permite
que façamos inferências sobre o estado e o nível de ansiedade que um animal, em geral
ratos ou camundongos, apresenta. No LCE existem quatro braços, sendo que dois possuem
paredes altas e dois são abertos. Um animal ansioso evitará a permanência prolongada nos
braços abertos, uma vez que estes animais tendem a evitar ambientes abertos e se
demonstram desconfortáveis com a altura. No campo aberto a premissa é semelhante, dado
o desconforto pela presença no novo ambiente o animal exibirá uma série de
comportamentos que nos permitem fazer uma análise do nível de estresse que ele está
submetido.
Figura 3 – À esquerda o LCE como dois braços abertos e dois fechados; à direita o teste do Campo Aberto.
66 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
Como anteriormente citado, o medo é um estado no qual um agente externo
conhecido provoca reações de tensão, como por exemplo, algumas respostas de defesa
como luta ou fuga. O condicionamento é a base das tarefas de medo, no qual um estímulo
neutro (luz), pareado com um estímulo incondicionado por algumas vezes gerará uma
resposta comportamental a apresentação apenas do estímulo neutro inicial (luz).
Manipulações usando esta premissa são freqüentes, por exemplo, avaliações de tarefas
potencializadas ou inibidas pelo medo. A associação de um estímulo incondicionado, como
um som alto, ao choque e uma luz potencializará uma reação de medo no animal, como por
exemplo, um sobressalto (Fig. 4). Portanto, diferentes pareamentos e manipulações podem
elucidar alguns pontos chaves do comportamento emocional e suas bases neurais, como a
função amigdalar.
Figura 4 – Exemplo de medo condicionado, no caso um teste de sobressalto
potencializado pelo medo, no qual primeiramente o animal recebe um
pareamento de luz e choque, em seguida um som que provoca uma reação de
sobressalto e depois o pareamento de luz e som que levará a uma resposta
exagerada de sobressalto. Retirado de Davis, 1992. Os modelos animais têm sido ferramentas de grande valia
no
estudo,
tanto
comportamentos
dos
substratos
emocionais,
bem
neurais
como
envolvidos
em
nos
experimentos
farmacológicos para o teste de drogas antidepressivas e
ansiolíticas, que são amplamente utilizados em casos de
desordens psicológicas humanas.
A inclusão do estudo dos comportamentos emocionais nas neurociências permitiu a
elucidação de alguns substratos neurais envolvidos nestes comportamentos. Entretanto,
novos avanços devem ser feitos com mais estudos. Algumas respostas, como os reais
papéis de determinadas estruturas, podem ser alcançadas. E ainda a inclusão da emoção
como um domínio cognitivo permite que a levemos em conta no estudo dos mais variados
comportamentos, influenciando diretamente em cada um deles.
67 www.ib.usp.br/labnec
Modelos e Cognição
Rodrigo Pavão
Laboratório de Neurociências e Comportamento
[email protected]
A construção de conhecimento científico envolve observação de fenômenos
associada à reflexão sobre eles, e também a formulação de hipóteses e criação de modelos.
Hipóteses e modelos científicos são criações humanas que tentam representar a realidade
de modo sintético e aproximado. Hipóteses e modelos têm o propósito de facilitar a
compreensão da realidade e permitir a previsão de fenômenos.
Existem diferentes tipos de modelos, dependendo de seus objetivos e organização.
Por exemplo, há modelos que remetem aos principais achados empíricos, ou que
representam relações entre partes do processo, ou ainda modelos classificatórios que
ressaltam semelhanças e diferenças entre processos, e até mesmo representações físicas,
como o modelo em estrutura metálica da molécula de DNA, desenvolvido por Watson e
Crick. Os modelos podem ser alterados para abordar aspectos não notados em sua criação.
Além disso, um mesmo fenômeno natural pode ser abordado por diferentes modelos.
Ademais, modelos podem exibir diferentes graus de generalidade e poder
explanatório. A teoria da evolução proposta por Darwin é um exemplo de modelo de ampla
generalidade e poder explanatório, pela sua aplicabilidade a uma ampla gama de
fenômenos biológicos. Em neurociências existem modelos de um único processo (p.ex., um
modelo de memória para tarefa de lembrança de listas de palavras) ou modelos mais gerais
(p.ex., o modelo hebbiano de plasticidade sináptica, que descreve que a eficácia sináptica
aumenta com a estimulação repetitiva). Um modelo geral é aquele que se aplica a uma
variedade de circunstâncias distintas, e pode ser definido como um paradigma (Sayão,
2001).
Modelos sobre processos cognitivos
Assim como em outras áreas da ciência, processos cognitivos também são
compreendidos e estudados por meio de modelos. Modelos de processos cognitivos são
representações dos processos mentais; aqueles propostos pela assim chamada área de
neurociência cognitiva representam sistemas e interações destes sistemas. Há modelos que
representam também o modo de funcionamento, relacionando funções cognitivas a
estruturas neuroanatômicas e a mecanismos neurofisiológicos, e que têm o objetivo de
facilitar a compreensão dos mecanismos. Adicionalmente, os modelos cognitivos podem ter
poder preditivo em relação ao comportamento. Serão apresentados dois exemplos de
funções cognitivas e alguns modelos relevantes para sua compreensão.
68 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
Exemplo 1 - Memória
Diversos modelos tentaram identificar a existência de múltiplas formas de memória
associadas a sistemas neurais distintos, cada qual com diferentes características. Por
exemplo, o modelo proposto por Atkinson e Shiffrin (1971), que ficou conhecido como
“modelo modal”, definia três tipos de estocagem mnemônica, incluindo (1) registros
sensoriais, o primeiro estágio da percepção, cujo conteúdo seria transferido para (2)
registros de curta duração, um armazenamento temporário das informações, que seriam
transferidas para (3) um registro de longa duração (Fig. 1 – esquerda). Assim, a informação
fluiria através de estágios sucessivos de processamento, podendo ser estocada em uma
memória de longa duração, o último estágio da cascata. No entanto, contrariamente a essa
proposta, havia evidências de que a informação poderia fluir para memória de longa duração
independentemente de sua permanência na memória de curta duração.
Como uma alternativa aos registros sensoriais e de curta duração do modelo modal
de memória, Baddeley e Hitch (1974) propuseram o modelo de memória operacional para
descrever a retenção temporária e manipulação de informações. A memória operacional
compreenderia um sistema de controle de atenção, o executivo central, auxiliado por dois
sistemas de suporte responsáveis pelo arquivamento temporário e pela manipulação de
informações, um de natureza visuo-espacial e outro de natureza fonológica (Fig. 1 – centro).
Adicionalmente, a memória de longa duração também foi dividida em sistemas
diferentes a partir de estudos de dupla dissociação envolvendo pacientes com lesões ou
disfunções no lobo temporal medial e nos gânglios basais. Assim, a memória de longa
duração pode ser dividida em conhecimento explícito - relacionado a fatos e eventos,
expresso pela lembrança da informação sob forma passível de relato verbal - e
conhecimento implícito, que poderia ser subdivididos em subsistemas (Squire e Knowton,
1995) (Fig. 1 – direita).
Figura 1 – Modelos modal de memória (à esquerda) (modificado de Atkinson e Shiffrin, 1971), memória
operacional (ao centro) (modificado de Baddeley e Hitch, 1974) e memória de longa duração (à direita)
(modificado de Squire e Knowton, 1995).
É possível exemplificar a atuação dos sistemas de memória operacional e de longa
duração através de atividades. A memória operacional é fundamental para a atividade de
lembrar uma lista de itens por um curto período de tempo, como ao memorizar um número
de telefone temporariamente até discá-lo. A memória explícita, por sua vez, é fundamental
69 www.ib.usp.br/labnec
para lembrar uma lista de itens por período de tempo prolongado, ou lembrar de evento
ocorrido em momento remoto. A memória implícita não está envolvida na lembrança
declarativa de itens; é fundamental, no entanto, para aprender e desempenhar relações
percepto-motoras como andar de bicicleta ou tocar um instrumento musical (como em
situação em que, ao se tocar uma série de notas no violão ou bateria, outras notas podem
ser tocadas sem fazer um planejamento explícito), cujo relato declarativo é freqüentemente
inviável.
Uma estratégia bem controlada de avaliar a memória implícita é o uso da tarefa de
aprendizagem de seqüências. Nessa tarefa o voluntário deve apertar, o mais rápido
possível, botões correspondentes a estímulos apresentados numa tela de computador. Os
estímulos podem ser apresentados aleatoriamente ou em uma seqüência. Pode-se controlar
quais os estímulos apresentados, sua duração, taxa de apresentação etc., além de acessar
com exatidão a velocidade das respostas e a precisão do voluntário, permitindo comparar o
efeito de diferentes tratamentos, incluindo o uso de seqüências. Em apresentação de
estímulos aleatórios os tempos de resposta são maiores do que para apresentação de
estímulos em seqüência; isso indica que a seqüência é aprendida. É interessante notar que
essa redução dos tempos de reação ocorre mesmo sem que a organização da seqüência
seja percebida conscientemente; essa é a razão de classificar essa tarefa como de memória
implícita. Exemplo 2 - Atenção
Modelos dos processos atencionais são menos consensuais que os modelos de
memória. Há classificações controversas como a atenção sustentada (prontidão do sistema
nervoso), atenção dividida (direcionamento da atenção concomitantemente a mais de uma
fonte) e atenção seletiva (processamento de informações oriundas de uma fonte, ignorando
as demais) (Muir, 1996), que não parecem de grande utilidade pela ampla sobreposição dos
processos e pelo fato de que em qualquer das situações existiria um processamento
seletivo. Há também o debate sobre como se daria essa seleção: como filtro (permitindo
processamento adicional de apenas uma parte da informação transmitida pelo sistema
sensorial), filtro atenuador (manutenção do sinal a ser processado, associado à redução dos
demais sinais não atendidos), ou intensificador (amplificação do sinal a ser processado,
associado à manutenção dos demais sinais não atendidos) (Fig. 1 – esquerda). A seleção
do que seria processado preferencialmente poderia se dar em diferentes níveis do sistema
nervoso – desde o sistema sensorial até as áreas integrativas. Há também o debate sobre
como se dá o direcionamento da atenção (Fig. 2 – centro e direita); há modelos que o
desmembram em direcionamento automático e direcionamento voluntário, envolvendo
diversas estruturas com diferentes funções (Posner, 1987; Laberge, 1989). Há ainda a
70 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
interpretação de que a atenção seja um processo de seleção modulado pelo registro do
passado, expectativa e funções superiores (Fig. 2 – direita).
Figura 2 – Seleção por filtros simples, atenuador ou amplificador (à esquerda) (modificado de Helene e Xavier,
2003). Modelo de etapas do direcionamento da atenção visual no espaço (ao centro) (modificado de Posner,
1987). Interação do filtro atencional com outros processos cognitivos (à direita). Modificado de Laberge, 1989.
Um tipo de avaliação amplamente utilizado em estudos sobre o direcionamento
espacial da atenção visual foi proposto por Posner (1980). Os participantes devem
pressionar um botão assim que detectam um estimulo luminoso (“alvo”) que aparece ou na
mesma região do espaço que um estímulo prévio (“pista válida”) ou na região oposta (“pista
inválida”) (Fig. 2 – centro, parte superior – representação da apresentação da pista, à direita
do ponto de fixação do olhar). O tempo de resposta quando a pista é válida é menor do que
quando a pista é inválida; essa diferença de tempo é uma medida do benefício da
orientação atencional gerado pela pista válida, que facilita o processamento do estímulo
visual, associado ao prejuízo gerado pela pista inválida em decorrência do direcionamento
da atenção para o local incorreto, dificultando assim a detecção do alvo que é apresentado
num outro local. Esse protocolo é comumente usado para avaliação da atenção automática
quando associado a 50% de tentativas válidas e 50% de tentativas inválidas (pistas não
preditivas da posição do alvo). Uma modificação desse protocolo, com o uso de pistas
simbólicas, centrais (apresentadas próximas do ponto de fixação) e outra proporção entre
pistas válidas e inválidas (tornando a pista preditiva da posição do alvo), é usada para
avaliação do direcionamento voluntário da atenção espacial. Além da orientação espacial
da atenção, estuda-se também o direcionamento atencional para processamento de formas,
contrastes etc.
Esses modelos esquemáticos são geralmente consistentes com achados empíricos e
clínicos, além de serem intuitivamente plausíveis. Há, no entanto, outras estratégias de
modelagem que tratam das computações envolvidas nos processos cognitivos.
71 www.ib.usp.br/labnec
Modelagem Computacional
A neurociência cognitiva tem usado a modelagem computacional como ferramenta
para explicação e entendimento dos mecanismos neurais subjacentes aos processos
cognitivos, por meio da implementação de programas de computador que traduzem modelos
abstratos em simulações concretas de processos cognitivos. Uma ampla gama de
processos pode ser modelada computacionalmente, desde a neurofisiologia neuronal até as
computações envolvidas em funções cognitivas complexas. A modelagem computacional
tem, portanto, um grande potencial na simulação de processos de integração incluindo os
níveis da neurofisiologia, neuroanatomia e neuropsicologia, podendo oferecer “insights”
sobre os processos computacionais complexos envolvidos no funcionamento integrado de
redes neuronais e na determinação do comportamento.
Um modelo computacional que vem sendo aplicado cada vez mais frequentemente
nas neurociências é a teoria de detecção de sinais, que apresentaremos a seguir.
Teoria de Detecção de Sinais
A teoria de detecção de sinais é uma adaptação da teoria de decisão estatística para
o campo da percepção (Swets e col., 1961). Uma estratégia interessante de explicação
dessa teoria é o uso do exemplo do diagnóstico de tumor por um médico observando
imagens de tomografia computadorizada (adaptado de Heeger, 2007).
A interpretação de imagens de tomografia é difícil e demanda bastante treino. Em
razão dessa dificuldade, há sempre incerteza sobre o julgamento. Pode existir um tumor
(sinal presente) ou não (sinal ausente). O médico pode ver o tumor (resposta “sim”) ou não
(resposta “não”). Existem quatro possibilidades, duas boas (identificação e rejeição corretas)
e duas más (omissão e alarme falsos).
sinal
presente
sinal
ausente
resposta
resposta
“sim”
“não”
acerto
omissão
alarme falso
rejeição correta
Figura 3 – Combinações possíveis entre presença/ausência de sinal e resposta sim/não da teoria de detecção
de sinais. Acertos (sinal presente, resposta sim) e rejeições corretas (sinal ausente e resposta não) são positivos;
alarmes falsos e omissões são negativos.
Dois fatores são fundamentais para a decisão: a aquisição de informação e o critério
A aquisição de informação, no nosso exemplo, se dá pela observação das imagens
da tomografia: formato, cor, textura etc. do tecido observado. Com bastante treino, o médico
consegue obter informação suficiente dessas imagens. Além disso, outros métodos
72 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
poderiam ser usados, como ressonância magnética, que poderiam fornecer informação
adicional. A aquisição de informação define a resposta interna (ver adiante).
O critério, por outro lado, é mais subjetivo ao próprio médico. Dois médicos
diferentes com mesma capacidade de análise, observando o mesmo exame, podem ter
diferentes opiniões sobre o que fazer. Um deles pode assumir que estará perdendo a
oportunidade de fazer um diagnóstico precoce que pode significar a diferença entre a vida e
a morte, e que um alarme falso poderia resultar em uma operação de rotina para biópsia; e,
nesse contexto, opta pela resposta “sim”. Outro médico pode assumir que cirurgias
desnecessárias, mesmo de rotina, são ruins, caras, estressantes etc.; e, nesse contexto,
pode adotar uma postura mais conservadora e optar pela resposta “não”. Este último médico
deixará de diagnosticar pacientes com tumor, principalmente em estágios iniciais, mas
estará reduzindo o número de cirurgias desnecessárias. Assim, o critério não se refere à
informação, mas sim à decisão que será tomada com essa informação.
Adicionalmente, existem ruídos que são processados juntamente com o sinal.
Ruídos, no nosso exemplo, correspondem às limitações da técnica, ou algo no tecido sadio
que é similar ao tumor. Além disso, o médico também exibe variações na maneira pela qual
analisa o exame. A soma do sinal com os ruídos determina a resposta interna.
A resposta interna poderia ser colocada de forma mais concreta, supondo que o
médico possua “neurônios-tumor” que têm a freqüência de disparo (em spikes/s) aumentada
ao ver exame com evidência de tumor. Note que apesar de este ser um exemplo bastante
didático, é bem pouco provável que o processamento realmente se dê desse modo. No
entanto, é bastante certo que o reconhecimento de tumores em exames de tomografia
envolva atividade diferenciada em alguns circuitos neurais de médicos neurologistas. A
atividade diferenciada nos circuitos neurais referentes ao reconhecimento de sinais será
referido como resposta interna.
O processo pode ser formalizado como representado na Fig. 4. A curva à esquerda
expressa apenas ruído (tecido sadio), e a curva à direita expressa sinal (tumor presente)
mais ruído. A abscissa representa a resposta interna, e a ordenada a probabilidade de
ocorrência. Numa situação envolvendo apenas ruído haverá, usualmente, 10 unidades de
resposta interna; porém, algumas vezes pode haver bem mais do que isso, i.e., até 18 ou 19
unidades de resposta interna. De maneira similar, numa situação envolvendo ruído mais
sinal pode haver menos do que 20 unidades de resposta interna, podendo gerar uma
sobreposição entre as curvas das duas condições.
73 www.ib.usp.br/labnec
Figura 4 – Resposta interna do observador (no exemplo, o médico que analisa os exames) para as condições
apenas ruído (tecido sadio) e sinal (tumor) mais ruído (à esquerda). Dois médicos com a mesma habilidade
podem adotar critérios distintos, levando a mais acertos e mais alarmes falsos (à direita, acima) ou menos
alarmes falsos e menos acertos, i.e., omissões (à direita, abaixo).
Exemplo 1 - Memória e a Teoria de Detecção de Sinais
A teoria de detecção de sinais tem sido usada nos modelos formais de
aprendizagem e memória. O modelo apresentado por Berry e col. (2008), por exemplo,
expressa valores de familiaridade amostrados em uma distribuição normal (análogo à
resposta interna) a cada item. A familiaridade exprime a força da memória que, na prática,
pode ser entendida como a facilidade de lembrar este item. O treinamento de um item
específico gera o aumento do valor da familiaridade daquele item (Fig. 5 – direita); assim,
assume-se que a média da familiaridade é maior para itens treinados do que para itens não
treinados, já que a familiaridade aumenta face a exposições repetidas do item em questão.
Tal valor de familiaridade é usado para fazer julgamentos de reconhecimento (“já vi” se valor
de “f” (familiaridade) for maior que um dado critério, e “não vi” se “f” for menor que o critério).
A familiaridade é usada também para obter medidas de pré-ativação, por exemplo, o tempo
de resposta para o item (Fig. 5 – esquerda).
74 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
Figura 5 – Esquerda: Familiaridade para um item em função da quantidade de treino e seu reflexo sobre a
lembrança e o tempo de reação. Esse modelo pode ser aplicado a diversas situações em que tradicionalmente
se julgam como necessários os sistemas de memória explícita (lembrança de lista de palavras, ou diferenciar
palavras apresentadas de não-apresentadas – inserindo critério de distinção dessas categorias) e implícita
(como tocar uma seqüência completa de notas, p.ex., 1-2-3-4, ou modificada, p.ex., 1-2-3-9). Direita: O efeito do
treino altera os valores de familiaridade fazendo com que itens inicialmente indiferenciados (parte superior)
tornem-se paulatinamente distintos (parte inferior). Esse modelo se aplica ao treinamento da capacidade de
médicos de diferenciar exames com tecido sadio e/ou tumor, treino para reconhecimento de palavras e
aprendizagem de seqüências, entre muitos outros. Exemplo 2 - Atenção e a Teoria de Detecção de Sinais
A aplicação dos conceitos da teoria de detecção de sinais ao estudo da atenção leva
à sugestão de que a atenção atua aumentando a resposta interna aos estímulos
selecionados (Fig. 6 – esquerda). Esse tipo de abordagem é bastante utilizada em
experimentos de detecção de contrastes, havendo relatos de alterações neurofisiológicas
associadas a estímulos aos quais a atenção foi direcionada. Por exemplo, quando a atenção
é direcionada para um dado estímulo, a taxa de disparos de neurônios isolados aumenta em
relação à apresentação de um estímulo de mesmo contraste, porém, sem o direcionamento
da atenção ao mesmo (Kim e col, 2007) (Fig. 6 – direita).
Figura 6 – Curvas de probabilidades da resposta interna, conforme a teoria de detecção de sinais aplicada à
atenção (esquerda). Resposta neural a estímulos aos quais a atenção foi ou não direcionada (direita). Modificado
de Kim e col, 2007.
75 www.ib.usp.br/labnec
Assim, também a atenção também pode ser modelada pela teoria de detecção de
sinais. Processos atencionais envolvendo a facilitação do processamento (possibilitando a
emissão de respostas mais rápidas ou melhor detecção de estímulos) poderiam, inclusive,
ser interpretados como fundamentados na mesma base que os processos de memória. De
fato, o experimento de Kim e col. (2007) consiste em apresentar uma pista indicando o lado
provável de apresentação do estímulo (com diferentes contrastes) que se assemelha ao
experimento de aprendizagem de seqüências (estímulos anteriores indicam o próximo
estímulo); assim, parece bastante plausível o uso do mesmo modelo.
Conclusão
Esse capítulo apresenta a possibilidade de investigar memória e atenção sob um
mesmo prisma, isto é, adotando um mesmo modelo básico. O modelo apresentado aqui é a
teoria de detecção de sinais, cuja aplicação parece vantajosa, na medida que facilita a
compreensão de processos cognitivos como atenção e memória e é um modelo elaborado
de forma a permitir previsões.
Nesse modelo, o processamento de estímulos seria facilitado de acordo com
respostas internas; ou seja, os tempos de resposta, lembrança, detecção etc. seriam
definidos pelo grau de preparação prévio do sistema nervoso. Esse grau de preparação é
dado pela estrutura e atividade dos circuitos neurais. Assim, a força das sinapses, a
quantidade ou a sincronização da atividade elétrica, entre outros, definiriam a facilidade de
resposta aos eventos.
A estratégia aplicada na Neurociência Cognitiva de assumir que existem módulos
para cada uma das funções cognitivas tem seu ganho na organização do estudo da
cognição. Essa e outras abordagens similares criaram modelos para cada um desses
processos cognitivos. E, realmente, o uso de modelos específicos para cada um dos casos
tem sua função de facilitar a compreensão daquele fenômeno; porém, é clara a interação (e
até mesmo similaridade) entre os diversos processos cognitivos. De fato, a estreita relação
entre atenção e memória já foi apresentada previamente por Helene e Xavier (2003). A
visão defendida aqui, entretanto, é que a computação desses dois processos é de tal modo
similar que haveria ganho na compreensão e na previsão de fenômenos através da adoção
de um mesmo modelo geral que fizesse a tradução da neurofisiologia para o
comportamento. A teoria de detecção de sinais é um modelo que tem se mostrado capaz de
atuar desse modo; de fato, a generalidade dessa teoria é tal que outros processos
cognitivos poderiam vir a ser modelados vantajosamente. 76 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
Bibliografia
Akirav I. e col. (2004) A facilitative role for corticosterone in the acquisition of a spatial task under
moderate stress. Learning and Memory 11:188–195.
Alcock, J. (2005) Animal Behavior. An evolutionay approach. 8ª ed., Sunderland: Sinauer Associates,
Inc.
Allen E. (1912) The cessation of mitosis in the central nervous system of the albino rat. Journal of
Comparative Neurology. 19:547-68.
Allman, J.M. (1999) Evolving brains. Scientific american library, New York, NY. 224 pp.
Altman, J. (1969) Autoradiographic and histological studies of postnatal neurogenesis IV. Cell
proliferation and migration in the anterior forebrain, with special reference to persisting
neurogenesis in the olfactory bulb. Journal of Comparative Neurology. 137, 433–458.
Altman, J. (1966) Autoradiographic and histological studies of postnatal neurogenesis II. A longitudinal
investigation of the kinetics, migration and transformation of cells incorporating tritiated
thymidine in infant rats, with special reference to postnatal neurogenesis in some brain regions.
Journal of Comparative Neurology. 128, 431–474.
Altman, J. (1963) Autoradiographic investigation of cell proliferation in the brains of rats and cats.
Postnatal growth and differentiation of the mammalian brain, with implications for a
morphological theory of memory. Anatomical Record. 145, 573–591.
Altman, J. (1962) Are new neurons formed in the brains of adult mammals? Science 135, 1127–1128.
Altman, J. & Das, G. D. (1965) Autoradiographic and histological evidence of postnatal hippocampal
neurogenesis in rats. Journal of Comparative Neurology. 124, 319–335.
Altman, J. & Das, G. D. (1966) Autoradiographic and histological studies of postnatal neurogenesis. I.
A longitudinal investigation of the kinetics, migration and transformation of cells incorporating
tritiated thymidine in neonate rats, with special reference to postnatal neurogenesis in some
brain regions. Journal of Comparative Neurology. 126, 337–390.
Atkinson, R.C. e Shiffrin, R. M. (1971) The control of short-term memory. Scientific American 225, 8290.
Baddeley A.D. (2000). The episodic buffer: a new component of working memory? Trends in Cognitive
Sciences. vol.4 pp. 417-423.
Baddeley, A. D. e Hitch, G. (1974). Working memory. In: Bower, G. A. (Ed) The Psychology of
Learning and Motivation. New York: Academic Press. 8, 47-89.
Baldo, M. V. C. . Bases Neurais da Percepção e Ação (no prelo) (2009). In: Araújo Filho, J. P.; Rui
Curi. (Org.). Fisiologia. 1 ed. Guanabara-Koogan, Rio de Janeiro.
Bargary, G. e Mitchell, K. J. (2008) Synaesthesia and cortical connectivity. Trends in Neuroscience.
Vol. 31, nº 07, pp. 335-342.
Bear MF, Connors BW & Paradiso MA (2002). Neurociências – Desvendando o Sistema Nervoso.
Porto Alegre 2ed, Artmed Editora.
Bear, M.F., Connors, B.W. e Paradiso, M.A. (1996) Language and attention. In: Satterfield, T.S. (Ed).
Neuroscience: Exploring the Brain. Baltimore, Maryland: Williams & Wilkins. pp. 576-615.
Bechara, A. e col. (1997) Deciding advantageously before knowing the advantageous strategy.
Science, v. 275, pp.1293-95.
Bechara, A. e col. (2001). Decision-making deficits, linked to a dysfunctional ventromedial prefrontal
cortex, revealed in alcohol and stimulant abusers. Neuropsychologia, 39(4):pp. 376-89.
Bechara. A. (2004). The role of emotion in decision-making: evidence from neurological patients with
orbitofrontal damage. Brain and Cognition, volume 55, issue 1, pp. 30-40.
Bendor, D. e Wang, X. (2005) The neuronal representation of pitch in primate auditory cortex. Nature.
Vol. 436, pp. 1161-1165.
Brandão, ML. (2004) Comportamento emocional. In: Brandão, M.L. As bases biológicas do
comportamento. Introdução à neurociência. Editora Pedagógica e Universitária. pp. 117-157.
Cameron, H.A. & Christie, B.R. (2007) Do new neurons have a functional role in the adult
hippocampus? Debates in Neuroscience 1:26–32.
Campos, A; Santos, A. M. G., Xavier, G. F. (1997). A consciência como fruto da evolução e do
funcionamento do sistema nervoso. Psicologia USP, São Paulo, v. 8, n.2, pp. 181-226.
Carlson, N. R. (2005) Foundations of physiological psychology. Ed. Allyn & Bacon, Boston, MA. 592
pp.
Cavedini, P. e col. (2002). Frontal lobe dysfunction in pathological gambling patients. Biological
Psychiatry, vol. 51, nº 4, pp. 334-341.
Cohen NJ. (1984). Preserved learning capacity in amnesia: evidence for multiple memory systems. In:
Squire LR, Butters N. (Ed) The Neuropsychology of Memory. New York: Guilford Press, pp.83103.
77 www.ib.usp.br/labnec
Damásio, AR. (1994) Erro de Descartes. Companhia das letras. São Paulo.
Darwin, C. (1872) Expressão da emoção nos homens e nos animais. Companhia das Letras. São
Paulo.
Davis, M. (1992) The role of the amygdala in conditioned fear. In: Aggleton, J.P (Ed.)The amygdala:
Neurobiological aspects of emotion, memory, and mental dysfunctions. pp. 255-305
Dethier, V. G. e Stellar, E. (1973). Comportamento animal. Trad. Diva Diniz Corrêa. São Paulo:
Edgard Blücher.
Dilema
do
prisioneiro.
Disponível
em:
<http://pt.wikipedia.org/w/index.php?title=Dilema_do_prisioneiro&oldid=15230186>
Acesso
em:26/05/09.
Eisch, A. J., Barrot, M., Schad, C. A., Self, D. W. & Nestler, E. J. (2000). Opiates inhibit neurogenesis
in the adult rat hippocampus. Proceedings of the National Academy of sciences of the USA. 97,
7579–7584.
Ekman, P. Frieser, W. (1976) .Pictures of Facial Affects. Consulting Psychologist Press.
Eriksson, P. S. e col.. (1998) Neurogenesis in the adult human hippocampus. Nature Medicine. 4,
1313–1317.
File S.E. (1992). Behavioural detection of anxiolytic action. In: Elliott JM, Heal DJ, Marsden CA (eds).
Experimental Approaches to Anxiety and Depression. pp 25–44.
Fuster, J. (2004). Upper processing stages of the perception–action cycle. Trends in Cognitive
Sciences, Vol.8 No.4, pp 143-145.
Fuster, J. (2006) The cognit: A network model of cortical representation. International Journal of
Psychophysiology 60 (2), pp. 125-132.
Gawryszewski, L.G., Ferreira, F.M., Guimarães-Silva, S (2007) ATENÇÃO!. In: Landeira-Fernandez,
J; Araújo-Silva, MT. (Ed.). Intersecções entre Psicologia e Neurociências. São Paulo: Lovise
Ltda. pp. 83-101
Gazzaniga, M. S., Irvy R B, e Mangun, G. R. (2002) Cognitive neuroscience, 2nd Ed. New York:
Norton.
Gazzaniga, M.S., Ivry, R. B., Mangun, G. R. (2006) Neurociência cognitiva – a biologia da mente – 2ª.
Ed. – Porto Alegre: Artmed. pp. 767.
Gold PE., W Farrell e RA King (1971). Retrograde amnesia after localized brain shock in passiveavoidance learning. Physiology & Behavior, v.7, n.5, p.709-712.
Goldman, S. A. & Nottebohm, F. (1983) Neuronal production, migration, and differentiation in a vocal
control nucleus of the adult female canary brain. Proceedings of the National Academy of
Sciences of the USA. 80, 2390–2394.
Gould, E. e col.. (2000) Regulation of hippocampal neurogenesis in adulthood. Biological psychiatry
48(8):715-20.
Haeger,
D.
(2007).
Signal
Detection
Theory.
Disponível
em:
<http://www.cns.nyu.edu/~david/handouts/sdt/sdt.html> Acesso em: 19/06/09
Hamilton A. (1901) The division of differentiated cells in the central nervous system of the white rat.
Journal of Comparative Neurology. 11:297-320.
Hansen, RD. Hansen, C.H. (1988). Finding the face in the crowd: an anger superiority effect, J. Pers.
Soc. Psychol. 54. pp 917–924.
Heale VR, Vanderwolf CH, Kavaliers M (1994): Components of weasel and fox odors elicit fast wave
bursts in the dentate gyrus of rats. Behavior Brain Research. 63:159 –165.
Hebb, D.O. (1949) The organization of behavior, New York: Wiley. 378 pp.
Helene, A. F. e Xavier, G. F. (2003). A construção da atenção a partir da memória. Revista Brasileira
de Psiquiatria, 25(Supl.II), 12-20.
Helene, A. F. e Xavier, G. F. (2007) Memória e (a Elaboração da) Percepção, Imaginação,
Inconsciente e Consciência. In: Landeira-Fernadez, J. e Silva, M. T. A. (Ed). Intersecções entre
psicologia e neurociências, 1ª edição. MedBook, Rio de Janeiro, pp. 103-148.
Heyes,C., Huber, L (2000) The Evolution of Cognition. London: MIT Press.
Huang, YX; Luo, (2006) Temporal course of emotional negativity bias: an ERP study. Neuroscience
Letters, 398 (1-2)..pp 91-96.
Hunt, G.R. (1996) Manufacture and use of hook-tools by New Caledonian crows. Nature 379: 249251.
Hunt, G.R. e Gray, R.D. (2004). Direct observations of pandanus-tool manufacture and use by a New
Caledonian crow (Corvus moneduloides). Animal Cognition 7: 114-120.
Immordino-Yang, MH; Damásio, A. (2007). We feel, therefore we learn: The relevance of affective and
social neuroscience to education. Mind, brain and education. pp. 3-10.
78 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
Isaacs, K. R. e col.. (1992) Exercise and the brain: angiogenesis in the adult rat cerebellum after
vigorous physical activity and motor skill learning. Journal of Cerebral Blood Flow and
Metabolism 12, 110–119.
Kandel, E. R.; Schwartz, J. H. e Jessell, T. M. (2000) Principles of neural science. 4º ed. Ed. McGrawHill Companies. New York, NY. 1414 pp.
Kaplan, M. S. & Hinds, J. W. (1977) Neurogenesis in the adult rat: electron microscopic analysis of
light radioautographs. Science 197, 1092–1094.
Kaplan, M. S. (1981) Neurogenesis in the 3-month-old rat visual cortex. Journal of Comparative
Neurology. 195, 323–338.
Kaplan, M. S. (1984) Mitotitic neuroblasts in the 9-day-old and 11-month-old rodent hippocampus.
Journal of Neuroscience. 4,1429–1441.
Kaplan, M. S. (1985) Formation and turnover of neurons in young and senescent animals: an electron
microscopic and morphometric analysis. Annals of the New York Academy of Sciences. 457,
173–192.
Kempermann G, Gage FH. (2002) Genetic determinants of adult hippocampal neurogenesis correlate
with acquisition, but not probe trial performance, in the water maze task. European Journal of
Neuroscience 16:129–136.
Kempermann G, Kuhn HG, Gage FH. (1997) More hippocampal neurons in adult mice living in an
enriched environment. Nature 3;386(6624):493-5.
Kempermann, G. (2002) Why new neurons? Possible functions for adult hippocampal neurogenesis.
The Journal of Neuroscience. 22(3):635-8.
Kepecs, A. e col. (2008). Neural correlates, computation and behavioural impact of decision
confidence. Nature, vol 455/11 september pp.227-231.
Kihlstrom, J. F. (1987). The cognitive unconscious. Science, vol. 237 pp. 1445-1452.
Kim, Y.J., Grabowecky, M., Paller, K, Muthu K, Satoru Suzuki S (2007). Attention induces
synchronization-based response gain in steady-state visual evoked potentials. Nature
Neuroscience 10, 117 – 125.
Kolb, B. (2006) Do all mammals have a prefrontal cortex? In: Kaas, J. H. (editor) (2006) Evolution of
nervous systems – a comprehensive reference. Ed. Academic Press, New York, NY. 2.000 pp.
Körding, K. P. Wolpert, D. M. Bayesian decision theory in sensorimotor control. Trends in cognitive
sciences, vol. 10 no.7 july 2006. pp. 319-326.
Kornack, D. R. & Rakic, P. (1999) Continuation of neurogenesis in the hippocampus of the adult
macaque monkey. Proceedings of the National Academy of Sciences of the USA. 96 5768–
5773.
LaBerge, D. e Brown, V. (1989). Theory of attentional operations in shape identification. Psychological
Review, 96, 101-124.
Lang, P.J., Bradley, M.M., & Cuthbert, B.N. (2008). International affective picture system (IAPS):
Affective ratings of pictures and instruction manual. Technical Report A-8. University of Florida,
Gainesville, FL. McGaugh, 2004
Leggio, M.G. (2005) Environmental enrichment promotes improved spatial abilities and enhanced
dendritic growth in the rat. Behavioural Brain Research 163(1):78-90.
Lent, R. (2006) Cem bilhões de neurônios. Ed. Atheneu, Rio de Janeiro, RJ. 698 pp.
Malva, J. O. (2005) IBRO Brain Campaign. Neuroscience: Science of the Brain. Disponível em
http://www.braincampaign.org, Acesso em: 09/06/2009.
McFarland, D. (1991) Defining motivation and cognition in animals. International studies in the
philosophy of science, 5, 153 - 170.
Moghaddam B, Bolinao ML, Stein-Behrens B, Sapolsky R (1994): Glucocorticoids mediate the stressinduced extracellular accumulation of glutamate. Brain Research. 655:251–254.
Morris, R.G.M. (1981). Spatial localization does not require the presence of local cues. Learning and
Motivation 12: 239-260.
Muir, J. L. (1996) Attention and stimulus processing in the rat. Cognitive Brain Research. 3, 215–222.
Naish,
D.
Chimpanzees
make
and
use
spears
Disponível
em:
http://scienceblogs.com/tetrapodzoology/2007/02/chimpanzees_make_and_use_spear.php.
Acesso em: 02/06/09.
Neisser, U. e Becklen, R. (1975) Selective looking: Attending to visually specified events. Cognitive
Psychology, 7(4), 480-494.
Nowakowski RS, Lewin SB, Miller MW. (1989) Bromodeoxyuridine immunohistochemical
determination of the lengths of the cell cycle and the DNA-synthetic phase for an anatomically
defined population. Journal of Neurocytology. 18(3):311-8.
O’Keefe J, Nadel L. (1978). The hippocampus as a cognitive map. Oxford: Clarendon Press. 570 pp.
79 www.ib.usp.br/labnec
Papez, JW. (1937) A proposed mechanism of emotion. J Neuropsychiatry Clin Neurosci. 1995
Winter;7(1) pp .103-12.
Pavão R. (2009). Aprendizagem e Memória. Revista da Biologia, v.1 n.1 p. 16-20.
Posner, M. I., Inhoff, A. W., Friedrich, F. J. e Cohen, A. (1987) Isolating attentional systems: A
cognitive-anatomical analysis. Psychobiology, 15, 107-121.
Posner, M.I. (1980). Orienting of attention, Quatery Journal of Experimental Psychology, 32:2-25.
Pratto, F. John, O.P. (1991) Automatic vigilance: the attention-grabbing power of negative social
information, J. Pers. Soc. Psychol. Vol. 61. pp. 380–391.
Preuss, T. M. (2006) Primate brain evolution in phylogenetic context. In: Kaas, J. H. (ed.) (2006)
Evolution of nervous systems – a comprehensive reference. Ed. Academic Press, New York,
NY. 2.000 pp.
Prinz, W. (2005) An ideomotor approach of imitation. In: Hurley, S. e Chater, N. (Eds.), Perspectives
on Imitation: From Neuroscience to Social Science, The MIT Press, Vol 1 pp 141-156.
Ramachandran, V.S. e Hubbard, E. M. (2003) The phenomenology of synaesthesia. Journal of
Consciousness Studies. Vol. 10, nº 08, pp. 49-57.
Rizzolatti G. e Craighero L. (2004). The mirror neuron system. Annual Review of Neuroscience,
27:169-92.
Rizzolatti G., Fogassi L., Gallese V, Fogassi L. (1996). Premotor cortex and the recognition of motor
actions. Cognitive Brain Research, 3:131-41.
Robertson, L. C. e Rafal, R. (2000) Disorders of visual attention. In M. Gazzaniga (Ed.), The new
cognitive neurosciences, 2ª ed., Cambridge: MIT Press, pp. 633-649.
Sayão, L. F. (2001). Modelos teóricos em ciência da informação: abstração e método científico.
Ciência e Informação. 30, 1, 82-91.
Schneider, D. D. G. & Parente, M. A. M. P. (2006). O desempenho de adultos jovens e idosos no Iowa
Gambling Task (IGT): um estudo sobre a tomada de decisão. Psicol. Reflex. Crit. vol.19, n.3,
pp. 442-450.
Scoville WB., B Milner (1957). Loss of recent memory after bilateral hippocampal lesion. Journal of
Neurolog,. Neurosurgery and Psychiatry. v.20, p. 11-21.
Sekuler, R. e Blake, R. (1994) Perception. McGraw-Hill Companies, 3ª ed. 420 pp.
Seung, H. S. (2000) Half a century of Hebb. Nature Neuroscience Supplement. (3), pp. 1166.
Shashoua V. (1985). The role of extracellular proteins in learning and memory. American Scientist, v.
73, p. 364-370.
Shurman, B. Horan, W. P., Nuechterlein K. H. (2005). Schizophrenia patients demonstrate a
distinctive pattern of decision-making impairment on the Iowa Gambling Task. Schizophrenia
Research, vol. 72, nº 2, pp. 215-224.
Simons, D. J. e Rensink, R. A. (2005). Change blindness: past, present, and future. Trends in
Cognitive Sciences, 9(1), 16-20.
Soon C.S. e col (2008): Unconscious determinants of free decisions in the human brain. Nature
neuroscience vol 11 n5 may . pp. 543-545.
Sotelo, C. (2003) Viewing the brain through the master hand of Ramón y Cajal. Nature Reviews
Neuroscience. 4(1):71-7.
Squire LR, Zola-Morgan S. (1991). The medial temporal lobe memory system. Science v. 253 p.13801386.
Squire, L.R. e Knowlton, B.J. (1995). Memory, hippocampus, and brain systems. In Gazzaniga, M S
(Ed.). The cognitive neurosciences. Cambridge, A Bradford Book, 825-837.
Stelmach, L.B. e Herdman, C.M. (1991) Directed attention and perception of temporal order. Journal
of experimental Psychology; Human perception and performance. 17 (2), 539-550.
Sternberg, R. J. (2000). Psicologia Cognitiva, Ed. Artmed, Porto Alegre, RS. pp. 494.
Sugita N. (1918) Comparative studies on the growth of the cerebral cortex. Journal of Comparative
Neurology. 29:61-117.
Swets. J. A., Tanner, W. P. Jr e Birdsall, T. G. (1961). Decision processes in perception. Psychology
Review. 68, 301-40.
Tanapat P, Hastings N, Reeves AJ, Gould E (1999): Estrogen stimulates a transient increase in the
number of new neurons in the dentate gyrus of the adult female rat. Journal of Neuroscience.
19:5792–5801.
Teoria
dos
jogos.
Disponível
em:
<http://pt.wikipedia.org/w/index.php?title=Teoria_dos_jogos&oldid=15346385.> Acesso em:
26/05/2009
Tinbergen, N. (1971) Comportamento Animal. Trad. Pinheiro de Lemos. Rio de Janeiro: José Olimpio.
80 www.ib.usp.br/labnec
1º Curso de Neurociências e Comportamento
Tit for tat. Disponível em: <http://en.wikipedia.org/w/index.php?title=Tit_for_tat&oldid=292379412>
Acesso em: 26/05/2009.
Treisman, A. M. e Gelade, G. (1980) A feature integration theory of attention. Cognitive Psychology,
12, 97-136.
van Praag e col.. (2002) Functional neurogenesis in the adult hippocampus. Nature 415(6875):1030-4.
van Praag H, Kempermann G, Gage FH. (1999) Running increases cell proliferation and neurogenesis
in the adult mouse dentate gyrus. Nature Neuroscience, 2(3):266-70.
Wilson, E.O. (2000) Sociobiology: The New Synthesis, Twenty-fifth Anniversary Edition. Cambridge:
Belknap Press. 697 pp.
Xavier G.F. (1993). A modularidade da memória e o sistema nervoso. Psicologia USP v.4, p.61-115.
Xavier, G. F., Saito, M.I.P. (1991) Stein C. Habituation of Exploratory Activity to New Stimuli, to the
Absence of a Previously Presented Stimulus and to New Contexts, in Rats. The Quarterly
Journal of Experimental Psychology Section B, 438 (2) pp. 157-175.
81 www.ib.usp.br/labnec