Biologia
Guto
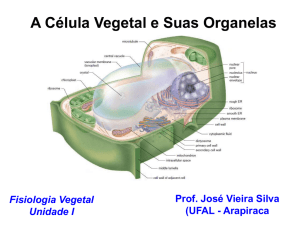
A ORIGEM DAS ORGANELAS
Texto adaptado pela Profa. Dra. Sônia Lopes, com base em trechos do capítulo “O mundo de
RNA e a origem da complexidade da vida”, escrito em 2001 pela Profa. Dra. Mariana Cabral de
Oliveira e pelo Prof. Dr. Carlos Frederico Martins Menck, ambos da Universidade de São Paulo, e
Publicado no livro Biologia Molecular e Evolução, Holos Editora, Editor Sérgio Russo Matioli.
A fotossíntese baseada em clorofila a é restrita a algumas eubactérias. Os eucariotos fotossintéticos
adquiriram essa capacidade por endossimbiose com eubactérias.
A clorofila a está presente em todos os organismos que fazem fotossíntese com desprendimento de
oxigênio. O surgimento da clorofila, uma molécula complexa, alterou completamente a atmosfera, causando
um enorme impacto na história do planeta e redirecionando a evolução dos seres vivos. Com o advento da
fotossíntese, o O2 começou a ser liberado para a atmosfera terrestre e foi se acumulando gradativamente. Por
volta de 2,8 a 2,4 bilhões de anos, o oxigênio já deveria estar presente em pequena quantidade (em torno de
0,5%), possibilitando a respiração aeróbia. Nessa época, devem ter-se originado os ancestrais da linhagem que
mais tarde iria originar as mitocôndrias. Nesse período, teriam surgido também as primeiras defesas celulares
contra os efeitos tóxicos do oxigênio, então um gás letal para a maioria das formas de vida existentes no
planeta. Apenas há cerca de 2 bilhões de anos é que o oxigênio deve ter-se acumulado na atmosfera em
quantidades suficientes para formar uma camada de ozônio (O3, a qual diminuiu a incidência de raios
ultravioleta sobre a superfície terrestre).
Os eucariotos, como conhecemos hoje, isto é, células nucleadas e com organelas, teriam surgido de
eventos de endossimbiose (simbiogênese) entre uma célula hospedeira e células procarióticas que deram
origem às mitocôndrias e aos cloroplastos. Simbiogênese é o surgimento de uma nova linhagem de
organismos como conseqüência de uma associação simbiótica estável.
A principal implicação da simbiogênese é que os eucariotos são, de fato, quimeras produzidas pela
combinação de diversos genomas.
As evidências que apóiam a simbiogênese das organelas como mitocôndrias e Cloroplastos são: (I) as
proteínas presentes nas organelas são mais semelhantes aos seus análogos procarióticos do que aos
eucarióticos; (2) existem procariotos de vida livre com forte semelhança estrutural, bioquímica e genética com
as respectivas organelas; (3) as organelas possuem genoma próprio, com organização semelhante ao genoma
procariótico; (4) os RNAs (ribossômico, transportador e mensageiro) das organelas também são mais
semelhantes aos de procariotos; (5) as organelas são semi-independentes, com capacidade de replicação; e (6)
as organelas e suas funções estão, alternativamente, presentes ou ausentes das células eucarióticas, não sendo
encontradas formas intermediárias.
Além das características mencionadas, a presença de uma membrana dupla ao redor dessas organelas
também é tomada como uma evidência de origem endossimbiótica. A membrana mais interna é de origem
procariótica (membrana plasmática do endossimbionte) e a mais externa é de origem eucariótica (membrana
do fagossomo da célula hospedeira). Pela criofratura, é possível verificar a orientação das membranas, pois as
duas camadas lipídicas têm aparência diferente quando estão voltadas para o interior ou exterior. A membrana
mais interna parece estar na orientação "correta", enquanto a membrana mais externa parece estar invertida,
indicando uma origem a partir do fagossomo.
Uma vez ocorrida a endossimbiose, genes do endossimbionte podem ser transferidos lateralmente para
o núcleo da célula hospedeira. Os produtos desses genes devem ser então direcionados às organelas.
Mitocôndrias e cloroplastos são, portanto, semi-independentes, já que necessitam dos produtos de alguns
genes que agora são codificados no núcleo. Em alguns complexos enzimáticos, uma parte das subunidades é
codificada pelo genoma nuclear e uma parte pelo genoma da organela. Transferências de genes entre
mitocôndrias e cloroplastos também podem ter ocorrido. A transferência específica de genes entre os
compartimentos celulares varia nos diferentes organismos. O mecanismo pelo qual se dá essa transferência
lateral no interior das células não está estabelecido. Esses eventos de transferência intensificam a dependência
entre organelas e núcleo, e provavelmente são essenciais à manutenção da associação endossimbiótica.
Apesar de eventos de transferência e perda de genes ocorrerem provavelmente ao acaso, aparentemente existe
uma direcionalidade, já que o núcleo apresenta uma tendência de adquirir genes, enquanto as organelas
parecem perder genes redundantes.
Uma explicação para essa direcionalidade seria que o núcleo é um ambiente geneticamente mais estável
que as organelas.
www.inforum.insite.com.br/22342
Biologia
Guto
A maioria das linhagens eucarióticas tem mitocôndrias que foram adquiridas por meio de um evento de
endossimbiose entre uma célula eucariótica e uma bactéria púrpura, provavelmente há cerca de 2,5 bilhões de
anos. O parente atual mais próximo das mitocôndrias é a bactéria Rickettsia, um parasita intracelular causador
do tifo, cujo genoma foi sequenciado em 1998.
As mitocôndrias apresentam duas membranas, a mais externa normalmente lisa e a mais interna com
dobramentos, e podem assumir diversas conformações (tubular vesicular ou lamelar). Apesar da variação de
forma, número e tamanho das mitocôndrias, seqüências moleculares têm mostrado uma origem monofilética,
indicando um único evento de aquisição de mitocôndrias por endossimbiose, seguido de algumas raras perdas
secundárias.
Existem linhagens eucarióticas que não possuem mitocôndrias e muitas dessas linhagens parecem ter
surgido antes da maioria das linhagens atuais de eucariotos. Esses protistas sem mitocôndria foram reunidos
no grupo arqueozoa (ameboflagelados, diplomonas, microsporídios e tricomonas), que se acreditava ter
divergido antes da aquisição de mitocôndrias. Entretanto, nos últimos anos, têm sido achados genes
tipicamente mitocôndrias no núcleo de todas essas linhagens, indicando que ocorreu uma perda secundária da
mitocôndria. Isso não significa que esses grupos não são basais na evolução dos eucariotos, ou seja, não
invalidam as árvores filogenéticas propostas, mas apenas altera a interpretação feita sobre a condição do
ancestral comum das linhagens atuais de eucariotos.
Atualmente não existe um forte candidato a um eucarioto que nunca tenha possuído uma mitocôndria.
Muitos dos organismos sem mitocôndria são parasitas internos (por exemplo,Giardia). Isso sugere uma perda
da mitocôndria, já que esses ambientes são pobres em oxigênio.
Autores argumentam que, se não forem encontrados eucariotos atuais que nunca tiveram mitocôndrias,
a ordem dos eventos que levaram à origem da célula eucariótica deveria ser revista.
A hipótese tradicional diz que a célula hospedeira, adquiriu sua mitocôndria, já possuía um núcleo.
Alguns autores propõem que uma ordem de origem das estruturas eucarióticas mais lógica seria: primeiro o
citoesqueleto, possibilitando a fagocitose, depois a origem da mitocôndria, seguido do sistema de
endomembranas e, por fim, o compartimento nuclear.
Todos os organismos que possuem cloroplastos também possuem mitocôndrias, o que sugere que as
mitocôndrias precederam os plastos. Entretanto, isso também pode indicar que as mitocôndrias são
obrigatórias para a manutenção de plastos.
A época de origem dos eucariotos tem sido estimada usando o tamanho das células no registro fóssil. A
descoberta de um fóssil denominado Grypania spiralis, com idade estimada em 2,1 bilhões de anos,
interpretado como uma alga eucariótica fotossintetizante indica que a origem do cloroplasto por
endossimbiose teria ocorrido antes dessa data.
A associação simbiótica entre organismos autótrofos e heterótrofos é extremamente comum na natureza.
Diversos animais e protozoários apresentam associações com microalgas (como corais, esponjas, ascídias e
foraminíferos). Os fungos associam-se às algas, formando os liquens. Em ambientes iluminados e ricos em
nutrientes, a fotossíntese tende a ser superprodutiva, a ponto de haver excesso de produção para o organismo
hospedeiro.
Existem vários tipos de cloroplastos que diferem em sua forma, ultraestrutura e pigmentação. Entretanto,
seqüências de RNAr e de vários outros genes indicam uma origem monofilética para todos os cloroplastos
que teriam surgido a partir de um evento endossimbiótico entre uma célula eucariótica (célula hospedeira) e
uma cianobactéria. Esse resultado foi uma surpresa para alguns pesquisadores que consideravam a
diversidade pigmentar e estrutural encontrada nos organismos fotossintetizantes atuais como indicação de
uma origem polifilética dos cloroplastos. Atualmente, a origem endossimbiótica dos cloroplastos já está
plenamente estabelecida, embora ainda exista controvérsia quanto ao número de eventos de endossimbiose
que levou à formação dos diferentes tipos de cloroplastos. As árvores filogenéticas indicam um único evento
primário de endossimbiose. Entretanto, esses dados também poderiam ser interpretados como eventos
múltiplos com um grupo de cianobactérias filogeneticamente próximas. Além do evento de endossimbiose
primário, múltiplos eventos de endossimbiose secundários ocorreram nas linhagens eucarióticas
fotossintéticas. Um evento de endossimbiose secundário é aquele onde uma célula eucariótica engloba outra
célula eucariótica que já contenha um cloroplasto (produto do evento primário). Eventos de endossimbiose
secundários geraram cloroplastos complexos com mais de duas membranas (três ou quatro). Em alguns casos,
é ainda possível verificar a presença de um núcleo vestigial (chamado nucleomorfo) entre a segunda e a
terceira membranas do cloroplasto. Na endossimbiose secundária, o núcleo da célula englobada sofre o
mesmo processo de redução, com perda e transferência lateral de genes para o núcleo da célula hospedeira.
O número de membranas ao redor dos cloroplastos é interpretado como indicativo de uma
www.inforum.insite.com.br/22342
Biologia
Guto
origem primária ou secundária. Os cloroplastos com duas membranas são considerados produto de
endossimbiose primária, sendo que a membrana mais interna é de origem procariótica (membrana plasmática)
e a mais externa de origem eucariótica (membrana do fagossomo). O cloroplasto secundário, além das duas
membranas, possui ainda uma terceira (membrana do eucarioto que foi englobado) e uma quarta (do
fagossomo) membranas eucarióticas. Em alguns casos, parece ter ocorrido a perda de uma das membranas,
como nas euglenófitas e nos dinoflagelados.
Entre os eucariotos fotossintetizantes atuais, reconhecemos três grupos cujo cloroplasto seria produto de
um evento primário de endossimbiose: (1) as algas verdes (Chlorophyta) e as plantas terrestres; (2) as algas
vermelhas (Rhodophyta); e (3) um pequeno grupo de algas unicelulares flageladas (Glaucocystophyta). Esses
grupos, por sua vez, teriam originado os plastos de outras linhagens de eucariotos por meio de eventos de
endossimbiose secundária.
Essa hipótese baseia-se em evidências de ultra-estrutura, bioquímicas e em comparações de seqüências
moleculares. Por exemplo, as algas vermelhas deram origem aos cloroplastos das algas com clorofila c
(heterocontes, haptófitas e dinoflagelados). As algas verdes são apontadas como o grupo que deu origem aos
cloroplastos das euglenófitas e, possivelmente, do plasto nãofotossintetizante dos apicomplexa, grupo que
inclui o Plasmodium (causador da malária) e Toxoplasma (causador da toxoplasmose).
A origem endossimbiótica dos plastos e mitocôndrias já estão firmemente estabelecidas.
Entretanto, endossimbiose também tem sido proposta para explicar a origem de praticamente todas as
demais organelas das células eucarióticas. Para essas outras organelas, como o sistema relacionado à
motilidade (sistema microtubular e flagelo), retículo endoplasmático, peroxissomos, entre outros, não existe
evidência molecular ou bioquímica conclusiva de origem endossimbiótica. Ao contrário, os dados existentes
favorecem a hipótese de origem autógena, onde essas estruturas teriam se originado e se organizado
gradativamente na célula eucariótica. Uma possível presença de DNA e RNA nos centros organizadores de
microtúbulos não foi confirmada.
Os organismos eucarióticos diversificaram-se em várias linhagens filogenéticas, das quais as principais
(chamadas, em inglês, de crown lineages) são os animais (Metazoa, incluindo os animais invertebrados e os
vertebrados), os fungos verdadeiros, as plantas (com clorofila a e b, que incluem as algas verdes), as algas
vermelhas, os estramenopilas (inclui os oomicetos, labirintulomicetos e as algas heterocontes-pardas,
diatomáceas e outras algas com clorofilas a e c e os alveolados incluindo os dinoflagelados, ciliados e os
apicomplexa). A diversidade molecular e fenotípica encontrada nos estramenopilas e alveolados são
equivalentes à encontrada nos outros reinos e ambos possuem organismos fotossintetizantes e heterotróficos.
A rápida diversificação das principais linhagens dos eucariotos ocorreu em torno de 1 a 1,5 bilhão de
anos atrás e pode ter sido ocasionada por diversos fatores, como alterações ambientais, por exemplo, o
aumento de oxigênio na atmosfera que pode ter atingido patamares que possibilitaram a ocupação de novos
nichos.
www.inforum.insite.com.br/22342