EMBRIOLOGIA
GASTRULAÇÃO
A gastrulação é o processo de migração de células e tecidos,
através do qual as células da blástula são totalmente rearranjadas.
A blástula é constituída por um grande no. de células; estas devido
à gastrulação ganham novas posições, sendo estabelecida a
estrutura multilamelar do organismo. Durante esta fase do
desenvolvimento, são formadas os 3 epitélios germinativos:
ectoderme (externa); mesoderme (mediana) e endoderme (interna).
A gastrulação envolve ao seguintes tipos de movimentos:
1. Epibola - camadas epiteliais envolvem as camadas mais
profundas do embrião.
2. Invaginação - ou Embolia, é o dobramento, (para dentro) de uma
região de células.
3. Involução - movimento de interiorização de uma camada externa
em expansão.
4. Delaminação - divisão de um epitélio em 2 camadas mais ou
menos
paralelas.
GASTRULAÇÃO DO OURIÇO DO MAR
As células do ouriço do mar são orinárias de diferentes regiões
do zigoto, têm tamanhos e propriedades diferentes. O destino de
cada camada celular, pode ser percebido por meio dos movimentos
das células, durante a gastrulação.
Após a ruptura da membrana de fecundação e eclosão da
blástula, o hemisfério vegetativo começa a se espessar e se
achatar. No centro dessa placa vegetativa, célula divididas dos
micrômeros, migram para o interior da blastocele. Essas células
correspondem ao mesênquima 1o. Duas proteína são importantes
para a migração dessas células (componente da lâmina basal da
blastocele de ouriço). Durante a gastrulação ocorre um aumento de
afinidade dos micrômeros por esta molécula; e as glicoproteínas
sulfatadas.
- Invaginação do arquêntero
1o. estágio :
As células que ficam na placa vegetativa (após a formação do
anel de células mesênquimais primárias) se deslocam para
preencher, os espaços deixados pelo ingresso de células
mesênquimais na blastocele. Portanto, a placa vegetativa se achata
ainda mais; dobrando-se, em seguida para dentro e se estendo
cerca de ¼ até metade de seu futuro comprimento na blastocele. A
região invaginada corresponde ao arquêntero (intestino primitivo), e
sua abertura, ao blastóporo. Essa invaginação da placa vegetativa,
para ser causada por forças internas da própria placa. O blastóporo
se encarrega da formação do ânus.
2o. estágio :
O arquêntero se estende muito. Neste processo de extensão, o
intestino primitivo é transformado em um tubo delgado. Para
realizar essa extensão, as células do arquêntero sofrem um
rearranjo, migrando umas sobre as outras e se achatando.
3o. estágio :
Células mesênquimais secundárias formam-se na extremidade
do arquêntero. Estas células entram em contato com superfície
interna da parede da blastocele, puxando o arquêntero.
Quando o topo do arquêntero encontra a parede da blastocele,
as células mesênquimais secundárias se dispersam na blastocele,
onde proliferam e vão formar os órgãos mesodérmicos. A boca é
formada no local, onde, o arquêntero entra em contato com a
parede da blastocele. A boca se funde com o arquêntero para
formar um tubo digestivo contínuo.
GASTRULAÇÃO NOS ANFÍBIOS :
- Movimentos celulares durante a gastrulação :
Nesses animais a gastrulação inicia-se no futuro lado dorsal do
embrião, na região do crescente cinzento. Nesse momento, há a
invaginação de células endodérmicas, formando um blastóporo em
forma de fenda. As “células
“e, vão revestir o arquêntero
inicial.
A gastrulação em anfíbios tem início na zona marginal (próxima
ao equador da blástula).
Posteriormente, as células do polo animal sofrem epibolia e
convergem para o blastóporo. Chegando ao lábio do blastóporo, as
células marginais se deslocam ao longo da superfície interna do
epitélio superficial. As 1o.s células que constituem o lábio dorsal,
são células endodérmicas ( que formarão a extremidade anterior do
arquêntero; posteriormente, tais células se tornam as células
faríngeas do intestino anterior). Ao penetrarem no interior do
embrião, estas células estarão representando os precursores da
mesoderme da cabeça. Em seguida, estas células sofrerão
involução, formando as células da cordomesaderme. Estas
formarão a notocorda ( a “espinha dorsal” mesodérmica transitória
essencial) para iniciar a diferênciação do sistema nervoso .
À medida que novas células migram para dentro do embrião, a
blastocele é deslocada para o lado oposto ao lábio dorsal do
blastóporo. Este, desloca-se em direção ao pólo vegetativo e se
expande (devido ao acréscimo de células do pólo animal). O
blastóporo em expansão “crescente”, desenvolve os lábios laterais
e o lábio ventral, pelo qual passam células precursoras da
mesoderme e endoderme. Com a formação do lábio ventral, o
blastóporo ganha de um anel, ao redor das células endodérmicas,
que permanecem expostas na superfície. Esta porção de
endoderme é chamada de “rolha de vitelo”(com o fechamento do
blastóporo, esta rolha de vitelo é levada para dentro do embrião).
- Migração da mesoderme :
Um aspécto importante da gastrulação de anfíbios, é a
convergência e a extensão da zona marginal em involução, que
corresponde à região de células, que fica acima do lábio do
blastóporo. A zona marginal em involução, contém a endoderme do
teto do arquêntero (na sua camada superficial) e, as células da
mesoderme (em sua região mais profunda).
Durante a gastrulação, o hemisfério animal e as células da zona
marginal, se expandem por epibola a fim de cobrirem todo o
embrião.
As células da zona marginal superficial em involução, sofrem
extensão convergente para formar a endoderme do teto do
arquêntero. A endoderme sub-blastoporal é coberta pelas células
da zona marginal (que não sofrem involução). Esta endoderme
forma o assoalho do arquêntero.
As células precursoras da mesoderme (que estão involuindo)
migram em direção ao pólo animal sobre uma rede de fibronectina,
secretada pelas células do teto da blastocele. A mesoderme parece
deslizar sobre estas fibras de fibronectina.
A matriz extracelular contendo fibronectina, parece transmitir
sinais para o direcionamento da migração celular.
- Epibolia da ectoderme :
O principal mecanismo da epibolia, corresponde a um aumento
no no. de células (através de divisão), simultaneamente a um
integração de várias camadas profundas, formando um única
camada.
O epitélio superficial se expande, em consequência da divisão e
de o achatamento celulares.
Todos os movimentos celulares vistos, resultam no
posicionamento dos 3 epitélios germinativos, que se preparam para
a diferênciação nos órgãos do corpo.
- GASTRULAÇÃO NAS AVES :
Nos embriões de aves, durante a clivagem, forma-se um
blastodisco sobre uma enorme quantidade de vitelo. As células da
região central do blastodisco, são separados do vitelo por uma
cavidade subgerminativa. As células da região centram do
blastodisco são claras, sendo esta região chamada de área
pelúcida. As células à margem da área pelúcida são opacas.
Enquanto a maioria das células permanece na superfície,
formando o epiblasto, algumas delas migram para a cavidade
subgerminativa, a fim de formarem o hipoblasto 1o. Posteriormente,
uma camada de células da blastoderme, migra, juntando-se às
células do hipoblasto 1o., formando o hipoblasto 2o. Na blastoderme
diblástica (epiblasto e hipoblasto), o espaço entre os 2 epitélios,
corresponde à blastocele.
As células do hipoblasto atuam na formação das membranas
externas, que alimentam e protegem o embrião. Os 3 folhetos
germinativos do embrião, são formados a partir de células do
epiblasto.
A estrutura mais evidente da gastrulação de aves, répteis e
mamíferos é a linha primitiva. Esta, surge como um espessamento
da camada celular, no centro da região posterior da área pelúcida.
À medida que a área espessada se estreita, ela se estende em
direção à região anterior e, se contrai para formar a linha primitiva
definitiva. Esta, corresponde ao eixo ântro-posterior do embrião
(simetria bilateral). Entre as bordas da linha primitiva se estabelece
uma depressão. Esta, é conhecida como fenda primitiva, e
corresponde a um blastóporo, por onde migram as células que
passam para a blastocele.
Na extremidade anterior da linha primitiva, há uma região de
células (mais espessa),chamada de nó de Hensen. O centro deste
nó possui uma depressão em forma de funil, através do qual as
células passam para a blastocele; estas células se dirigem para a
região anterior, formando a mesoderme da cabeça e a notocorda.
As células que migram através da fenda primitiva, formam a maior
parte da endoderme e da mesoderme.
- Mecanismos de gastrulação nas aves :
- Papel do hipoblasto :
O hipoblasto não contribui com células para o adulto, mas é
essencial para o desenvolvimento, pois, ele (hipoblasto) induz a
formação e o direcionamento da linha primitiva.
- Movimento celular no interior da blástula :
Como na gastrulação de anfíbios, as células do embrião de ave,
que passam pelo blastóporo, adquirem a forma de garrafa. Logo,
que estas células entram na blastocele, elas se achatam e passam
a fazer parte de um fluxo celular, onde as células migram de forma
independente. Polissacarídeos, como o ácido hialurônico, facilitam
a migração de células isoladas, mas eles não conseguem orientar o
movimento dessas células.
Estes movimentos celulares, nas aves, também estão
relacionados com a presença de uma rede de fibronectina, na
lâmina basal das células do epiblasto.
- Epibolia da ectoderme :
Durante a gastrulação das aves, as células precursoras da
ectoderme se expandem, a fim de revestirem o vitelo. Estas células
estão associadas e, se deslocam como uma unidade, não como
células isoladas.
Há indicações, da resistência de um reconhecimento específico
entre as células marginais da área opaca (do blastodisco) e a
superfície interna da membrana vitelina. Este reconhecimento é
essencial para a migração epibólica das células da ectoderme, que
revestem o vitelo.
GASTRULAÇÃO NOS MAMÍFEROS :
O desenvolvimento dos mamíferos é semelhante ao das aves.
Porém, os mamíferos não se desenvolvem isoladamente no interior
de um ovo. Eles criaram estratégias de desenvolvimento, dentro da
própria mãe. O embrião de mamífero obtém nutrientes diretamente
da mãe, não havendo a necessidade de acumular vitelo.
Esta evolução exigiu uma reestruturação da anatomia materna,
como o desenvolvimento de um órgão fetal capaz de absorver
nutrientes, fornecidos pela mãe. Este órgão, a placenta, é derivado
das células trofoblásticas do embrião, às quais se juntam células
mesodérmicas, derivadas da massa celular interna. A 1o.
segregação de células, que ocorre na massa celular interna, da
origem ao hipoblasto.
As células do hipoblasto separam-se da massa celular interna a
fim de revestirem a cavi// da blastocele, onde elas originarão a
endoderme do saco vitelino.
As céls restantes da massa cel interna (acima do hipoblasto) são
denominadas epiblasto. Essas céls são separadas por pequenas
fendas, com o intuito de afastar o epiblasto, as quais formam o
revestimento do âmnio. Uma vez, formado, o âmnio é preenchido
pelo fluido amniótico, q possui a fç de amortecer choques
mecânicos e prevenir a dessecação do embrião em
desenvolvimento.
Acredita-se q o epiblasto embrionário contenha todas as céls q
formarão embrião. Na margem posterior do epiblasto, ocorre um
espessamento, q produz uma linha primitiva. Através desta, migram
os precursores da endoderme e da mesoderme.
As céls extra-embrionárias contribuem p/ a formação de tecidos,
que possibilitarão a sobrevivência do feto, no interior do útero.
As céls do trofoblasto originam uma população de céls, q
constituirão ( por meio de : nuclear o chamado citotrofoblasto; a
massa cel multinucleada forma o sinciciotrofoblasto. O
citotrofoblasto fixa-se à parede uterina (endométrio) por meio de
moléculas de adesão. As céls do citotrofoblasto humano possuem
enzimas proteicas, q permitem a sua penetração na parede uterina
e a remodelação dos vasos sanguíneos uterinos, de modo que o
sangue materno chegue até os vasos sanguíneos do feto.
O sinciciotrofoblasto invade a parede do útero, encaixando o
embrião no endométrio. O útero, supre esta área com vasos
sanguíneos que entram em contato com o sinciciotrofoblasto.
A mesoderme extra-embrionária associa-se às extensões do
trofoblasto, originando os vasos sanguíneos, q conduzem os
nutrientes da mãe para o feto.
O pedúnculo de conexão, formado por mesoderme extraembrionária, q liga o embrião ao trofoblasto, forma os vasos
sanguíneos do cordão umbilical.
O cório é o componente fetal da placenta. Ele é constituído por
trofoblasto (sincicio e citotrofo ) associado à mesoderme extraembrionária e contém tb vasos sanguíneos. O cório se funde à
parede uterina p/ formar a placenta. Portanto, esta é formada pelo
endométrio do útero (porção materna) e pelo cório ( porção fetal).
O embrião envolto pelo âmnio e protegido pelocório. Este,
possui vilosidades q se projetam da sua superfície externa. Estas
vilosi// contém vasos sanguíneos e, permitem ao cório ampliar a
área de exposição ao sangue materno. Embora o sist. Circulatório
fetal e materno nunca entrem em contato direto, ocorre difusão de
substâncias solúveis, através das vilosi// do cório. Assim, por meio
da circulação materna, o feto absorve nutrientes e oxigênio e,
elimina produtos residuais (como o dióxido de carbono e a uréia).
Os vasos sanguíneos das vilosi// coriônicas, formam-se a partir
da mesoderme extra-embrionária, q penetra nos montículos de
tecido citotrofoblástico; estes montículos são denominados vilosi//
1o. As vilosi//2o. formam-se durante a 2o. semana de gestação. No
final da 3o. semana surgem as vilosi// 3o. ( com vasos sanguíneos).
Resumindo : O trofoblasto é necessário p/ a adesão e
penetração do embrião no endométrio uterino. O cório atua na troca
de gases e nutrientes entre mãe e feto. O cório , ainda, atua como
um órgão endócrino, sua porção de sinciciotrofoblasto produz 3
hormônios (essenciais p/ o desenvolvimento de mamíferos). São
estes : gonadotrofina coriônica (um peptídeo que induz céls da
placenta e do ovário a produzirem progesterona); este, é um
hormônio esteróide q mantém a parede uterina espessa e cheia de
vasos sanguíneos. Somatomamotrofina coriônica ( ou lactogênio da
placenta ), durante a gestação, este hormônio é responsável pelo
desenvolvimento dos seios maternos, capacitando-os à produção
de leite. O cório pode, ainda, ter a função de proteger o feto da
resposta imune da mãe, por meio da secreção de proteínas
solúveis, q bloqueiam a produção de anticorpos; e pela produção
de certos tipos de linfócitos, q suprimem a resposta imune normal
interior do útero.
As fç da placenta incluem : suporte físico, troca de nutrientes e
regulação do relacionamento endócrino e imunológico entre mãe e
feto.
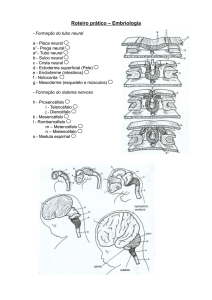
NEURULAÇÃO
Todos os vertebrados em desenvolvimento são muito
semelhante após a gastrulação. É só mais tarde, q as
características especiais da espécie, aparecem. Todos os
embriões de vertebrados têm arcos brânquiais, notocorda,
medula espinhal e rins pronéfricos.
Nos vertebrados, a gastrulação produz um embrião com uma
camada endodérmica interna, uma mesoderme intermediária e a
ectoderme externa. A interação com a mesoderme dorsal e a
ectoderme, q a recobre, inicia a organogênese (criação de tecidos e
órgãos específicos). Nessa interação, a cordomesoderme induz a
ectoderme, acima dela, a formar o tubo neural (oco)que,
posteriormente, irá se diferênciar em cérebro e medula espinhal.
Essa formação do tubo neural, é chamada de neurulação. Nesta
fase, a ectoderme original está dividida em 3 grupos de céls: o tubo
neural (posicionado interiormente), a epiderme da pele e as células
da crista neural (q migram da região que liga o tubo neural à
epiderme).Um embrião nesta fase é chamado de nêurula.
A 1o. indicação de que uma região da ectoderme está destinada
a tornar-se tecido neural, é uma mudança na forma das céls. As
céls ectodérmicas (na região mediana) torna-se alongadas,
enquanto que, as células destinadas a formar a epiderme, tornamse mais achatadas. O alongamento das céls ectodérmicas dorsais,
faz com que, a região neural cresça mais que a ectoderme
adjacente, criando assim, a placa neural. Em seguida, as bordas da
placa neural se espessam e deslocam-se p/ cima, p/ formar as
pegas neurais; enquanto um sulco neural (em forma de U) aparece
no centro da placa, dividindo os futuros lados direito e esquerdo do
embrião. As pregas neurais migram em direção à linha média do
embrião, unindo-se p/ formar o tubo neural (abaixo da ectoderme
superficial). As céls da porção mais dorsal do tubo neural,
transformam-se nas células da crista neural.
- O tubo neural e a origem do sistema nervoso central :
A formação do tubo neural, não ocorre simultaneamente ao
longo de toda a ectoderme.
Na extremidade cefálica (onde se formará e cérebro), a parede
do tubo é larga e espessa. O restante do tubo neural, permanece
como um tubo simples (q se afila em direção à cauda do embrião).
As duas extremidades abertas do tubo neural, são chamadas de
neuróporo anterior e neuróporo posterior. Em mamíferos, essas
aberturas permitem que o líquido amniótico circule, por certo tempo,
no interior do tubo neural. O não fechamento do neuróporo
posterior na espécie humana, resulta na chamada Spina Bífida. O
não fechamento do neuróporo anterior resulta na Anencefalia.
- Mecanismos de formação do tubo neural :
A formação do tubo neural está relacionada à mudanças na
forma das células. Nessas mudanças, estão envolvidos os
microtúbulos e os microfilamentos celulares.
As céls ectodérmicas, se alongam ao mesmo tempo em que os
microtúbulos se alinham, paralelamente, ao seu eixo de
alongamento. Uma 2o. mudança na forma das céls, envolve a
constrição apical das células cilíndricas. Nas laterais do tubo neural,
essa mudança é orientada por um anel de microfilamentos
contráteis, que circunda a margem apical de cada célula.
Outro mecanismo de formação do tubo neural, é a tração cortical
do epitélio da placa neural. Esse mecanismo ocorre pela
movimentação da córtex das céls da placa neural; essa
movimentação, tem inicio na extremi// basal e termina no ápice. As
céls ectodérmicas estão unidas entre si por moléculas, q são
carregadas por esse fluxo cortical. Se a veloci// média desse fluxo é
a mesma em todas as céls, estas permanecem ligadas umas às
outras, não ocorrendo nenhuma deformação da placa neural.
Porém se a veloci// dessa tração difere entre 2 céls adjacentes, a
cél mais veloz move-se em direção basal (p/ fora da placa neural).
Como as céls estão ligadas pelos seus ápices, ocorre deformação
da placa neural.
Os modelos anteriormente referidos da formação do tubo neural,
têm como base 3 princípios :
1. todas as forças necessárias p/ criar o tubo neural, são intrínsecas
ao próprio epitélio;
2. a neurulação é dirigida por mudanças na forma das céls
neuroepiteliais;
3. as forças que causam as mudanças na forma das células, são
geradas pelo citoesqueleto.
Porém, as diferenças observadas nas formas das céls, nas
diferentes regiões do tubo neural, indicam q forças extrínsecas ao
neuroepitelio atuam na moldagem do tubo neural. As céls
adjacentes à notocorda e as céls q se encontram nos limites do
sulco neural, tomam a forma de garrafa, porém, as outras céls do
sulco neural não. Portanto, enquanto muitos aspéctos da
neurulação podem ser explicados, apenas por fatores intrínsecos
às próprias céls da placa neural, algumas diferenças entre as
células do tubo neural, podem resultar do meio ambiente em q
ocorre a neurulação.
ALGUMA COISA SOBRE ORGANOGÊNESE
Em mamíferos, a organogênese inicia-se entre a 3o. e 8o.
semanas do período embrionário.
O desdobramento do disco trilaminar, transforma o embrião em
um cilindro. Esse dobramento é provocado pelo crescimento do
tubo neural. Neste estágio o embrião é envolvido pelo saco
amniótico.
- Destino dos 3 folhetos germinativos:
Ectoderme - origina SNC, SNP, epitélios sensitivos, epiderme e
anexos, gl. Mamárias, subcutâneas, hipófise e esmalte dos dentes.
Mesoderme - origina os ossos, músculos, cartilagens tec. Conj.,
Céls e vasos sanguíneos e linfáticos, aparelho urogenital, serosas e
baço.
Endoderme - origina revestimentos gastrointestinal, respiratório,
da bexiga, uretra, ap. auditivo, fígado e pâncreas.
O controle do desenvolvimento embrionário pode ser genético
(contido nos cromossomas) ou, por indução de tecidos, geralmente
por mediação química.
Alguns fatores q influenciam o crescimento fetal são : a
desnutrição materna, fumo, drogas e álcool, gravidez múltipla,
defeitos placentários diabetis materna, fatores genéticos.
DIFERÊNCIAÇÃO CELULAR
O desenvolvimento dos tipos especializados de céls, a partir de
um único ovo fertilizado, é chamado diferênciação celular. Esta
mudança na bioquímica e função celulares, é precedida por um
processo chamado determinação, no qual o destino da célula é
decidido. A determinação ocorre por 2 maneiras: 1o. é a segregação
citoplasmática de moléculas determinantes, durante a clivagem
embrionária. Nesse momento, os planos de clivagem separam
regiões do citoplasma do zigoto, entre diferentes céls filhas. 2o. é a
indução embrionária, q envolve a interação de céls ou tecidos.
PRÉ-FORMAÇÃO E EPIGÊNESE
Qualquer explicação sobre a diferênciação celular, tem que levar
em conta a constância da morfologia de cada espécie, a
diversidade das partes do corpo de cada organismo. O
desenvolvimento envolve a expressão das proprie// hereditárias das
espécies.
No séc XVII, a relação entre desenvolvimento...
A teoria da pré formação explicava melhor a continui// das
gerações, enquanto a epigênese explicava melhor a variação e as
observações diretas da formação dos órgãos.
Os teratologistas franceses - Embriologia moderna
ESPECIFICAÇÃO CITOPLASMÁTICA EM EMBRIÕES DE
TUNICADOS
À medida que o embrião se divide, diferentes céls incorporam
diferentes regiões do citoplasma. Acredita-se que estas diferentes
regiões citoplasmáticas, contém determinantes morfogênicos, que
controlam o destino da célula.