FISIOLOGIA HUMANA
HOMEOSTASE: é o termo usado para descrever os vários ajustes que o
organismo efetua para manter as funções normais e reguladas das células e de
todo o organismo. Exemplo: manter normais a pressão arterial, temperatura,
funcionamento regular dos rins, ajustes respiratórios, etc.
VOLUMES CORPÓREOS: o corpo humano é formado em grande parte por
água. Essa água está distribuída nas porções intracelular e extracelular. A
porção intracelular compreende a água contida dentro das células e a substância
mais predominante é K+. A porção extracelular, onde predomina Na+,
compreende o plasma sangüíneo (que tem uma certa concentração salina) e o
líquido extracelular que está presente entre as células e da sustentação aos
tecidos além da troca de nutrientes .
FORÇAS
RESPONSÁVEIS
PELA
MOVIMENTAÇÃO
DE
SUBSTÂNCIAS ENTRE OS COMPARTIMENTOS: compreende:
Difusão: processo pelo qual uma substância em solução se expande, devido ao
movimento (choques) das partículas, a fim de ocupar todo o volume disponível.
O espalhamento das moléculas ocorrerá pela quantidade de choques dessas
moléculas. Se uma solução está muito concentrada, maior será o número de
choques entre as moléculas, e isso faz com que essas moléculas passem de um
lugar que tem constante choque para outro que está menos concentrado a fim de
estabelecer um equilíbrio. A tendência da difusão de uma área para outra é
proporcional à diferença de concentração de uma certa substância nas duas
áreas (gradiente químico ou de concentração). A difusão dos íons também é
afetada pelas cargas elétricas. Há sempre uma diferença de potencial entre as
duas áreas.
Os canais das membranas são específicos para o tipo de substância. São
as proteínas transmembrana que formam os canais de passagem de substâncias.
Canais sempre abertos: difusão simples e contínua. Canais fechados: abrem-se
quando o potencial de voltagem das células muda, esses são os canais
dependentes de voltagem (os canais abrem e fecham sob o mesmo estímulo);
canais dependentes de receptor ou ligante, ex: canais de sódio na placa mio
neural.
Difusão facilitada depende de uma proteína carregadora. Também possuem um
canal seletivo. As moléculas batem em toda a membrana da célula e algumas
batem no sitio de ligação das proteínas, fazendo com que essa molécula entre na
célula. Os fatores que contribuem para essa difusão são: concentração,
permeabilidade da membrana e tamanho da molécula (que influi na quantidade
de choques).
Osmose: difusão de água. É o movimento das moléculas do solvente através de
uma membrana, para uma área uma área onde existe uma concentração maior do
soluto em relação ao qual a membrana é impermeável. A pressão osmótica é a
força que a solução concentrada exerce para que haja a difusão do solvente
(água). A pressão osmótica varia de acordo com a concentração da solução, se a
concentração de um a solução for alta a pressão osmótica é maior e a osmose é
mais rápida.
A difusão e a osmose são processos passivos, no sentido de que não
requerem consumo de energia.
Transporte ativo: as substâncias são transportadas no sentido contrário ao
gradiente, com gasto de energia. O transporte ativo secundário só ocorre quando
há na célula um transporte ativo primário.
POTENCIAIS DE MEMBRANA: é a voltagem constante de uma célula em
situação de repouso. Essa voltagem é em torno de –90 mV. Todas as células têm
potencial de membrana. Na membrana celular existem sistemas de transporte
que processam o fluxo de substâncias e de informações. O sistema mais
importante é a bomba de sódio e potássio, que está presente em todas as células.
Essa bomba está em constante entrada e saída de potássio e sódio
respectivamente e mesmo assim existe uma diferença nas concentrações IC e
EC.
A bomba manda e íons Na+ para for a da célula contra somente 2 íons K+
bombeados para dentro. Assim, ela tende a gerar uma separação de cargas
elétricas através da membrana, tornando o citoplasma negativo em relação ao
meio extracelular.
A negatividade elétrica do citoplasma decorre de uma combinação de
alguns fatores:
1. Existe uma assimetria de concentrações de Na+ e K+ entre o citoplasma e o
meio EC, mantida pela bomba de Na+ e K+.
2. Existem vias seletivas para os íons Na+ e K+ na membrana celular, os canais
iônicos.
3. A assimetria de concentração de outros íons com canais específicos na
membrana, como o Ca2+ e o Cl- contribui, em menor grau, para o potencial
de membrana. Em algumas células excitáveis a distribuição de íons cálcio e
os canais de Na+ e Ca2+ contribuem de modo muito mais importante para o
potencial de membrana e suas alterações.
O potencial de membrana em repouso é essencialmente um potencial de
equilíbrio de K+. o citoplasma é tanto mais negativo quanto menos a
concentração externa de potássio, de tal forma que, numa faixa razoável de
[K+]externo, o potencial de membrana (Vm) depende linearmente do logaritmo da
concentração externa de potássio.
O estabelecimento da DP depende de diferença de concentração,
permeabilidade e mais de 1 íon.
Todas as células exibem potencial de membrana. As células excitáveis,
quando estimuladas, vão ter uma variação da voltagem. As células excitáveis
tem duas populações de canais, eles estão fechados e com o estímulo eles se
abrem e fecham de novo. A abertura e fechamento do canal de Na + é mais
rápida que a do K+. os canais se abrem só com um estímulo mínimo de variação
na voltagem, que se chama linear de voltagem.
Na hora que se abrem os canais de Na+, aumenta a permeabilidade ao Na+
e ele entra na célula anulando as cargas negativas. Ele faz isso até atingir o pico,
depois disso, abrem-se os canais de K+ que saem da célula por causa do
gradiente favorável, fazendo com que a carga da célula volte a ser negativa pela
saída de K+. nesse processo, a concentração não varia.
Um estímulo de intensidade forte ou fraca é medido pela freqüência. O
potencial de um estímulo é o mesmo, o que varia é a freqüência. Ex: água
morna (menor freqüência) e água fervendo (maior freqüência).
A lei do tudo ou nada existe em função do limiar de disparo, onde ou os
canais se abrem ou eles ficam fechados. O fato de ser tudo ou nada faz com que
a intensidade do estímulo seja contínua. Ex: corte do dedo do pé, para a
informação chegar no córtex cerebral a intensidade do estímulo das células do
pé até chegar no córtex manterão a mesma intensidade.
Potencial propagado é onde o estímulo é feito em um certo local da
membrana e se propaga por toda a membrana da célula. Ex: axônio.
SISTEMA CIRCULATÓRIO
SEGMENTOS E SUAS FUNÇÕES:
1) CORAÇÃO: bomba que gera pressão. A pressão varia de 120 para 80
mmHg. É uma bomba de 2 tempos porque a pressão no ventrículo varia de 0
a 120 mmHg. A pressão arterial cai por causa da resistência dos vasos.
2) AORTA: possui muita musculatura, isso faz com que ela seja elástica. O
coração bate e joga o sangue na aorta com muita força, isso faz com que a
parede da aorta deforme, armazenando energia. A pressão não cai a zero
porque a aorta devolve aos poucos a energia armazenada nela.
3) ARTÉRIAS: grandes vasos que levam sangue para as arteríolas.
4) ARTERÍOLAS: possuem músculo liso, isso garante uma contração
mantida, o tônus muscular. É o segmento que mais tem músculo por
diâmetro. Elas são o principal ponto de resistência ao fluxo sangüíneo
porque estão sempre contraídas. Essa resistência dissipa tanta energia que a
pressão cai de 100 mmHg para 35 mmHg. Essa pressão tem que ser 35
mmHg porque os capilares não tem músculo e não suportariam uma pressão
alta. As arteríolas funcionam como o redutor de um encanamento.
5) CAPILAR: delgado, facilita as trocas difusionais.
6) LEITO VENOSO: vênulas, pouco músculo, leito distensível. 65% do
sangue está no leito venoso pois é distensível. Volume de reserva.
O coração funciona como um sincício funcional.
POTENCIAL DE AÇÃO CARDÍACO: existem canais de sódio que se
abrem, quando eles se fecham, abrem-se os canais de cálcio que são pouco
seletivos, deixando entrar também, um pouco de sódio. Quando os canais de
cálcio se fecham, abrem-se os canais de potássio para a repolarização.
Período refratário: a célula excitável não responde ao estímulo, é
impossível reexcitar aquela célula antes dela se recompor. Período difícil de
reestimular a célula nervosa, a não ser por um estímulo muito forte. Mas há uma
parte que vai do pico até a repolarização que não reestimula. No coração esse
período é importante porque quando estiver relaxado é o momento em que ele
se enche de sangue novamente.
O cálcio em grande quantidade estimula a contração. Para manter o platô
não é a quantidade e sim os íons que são os primeiros a entrar no canal.
Para a contração muscular é necessário muito cálcio.
Automaticidade: 3 conjuntos de células auto-excitáveis: SA, AV e
Purkinje. Esses 3 conjuntos trabalham com velocidades diferentes, mas geram
um potencial de ação. Age primeiro nos átrios, contraindo-os e depois age nos
ventrículos.
Ciclo cardíaco: sístole e diástole. Na diástole o sangue passa direto do
átrio para o ventrículo, pois a pressão do ventrículo é zero. Átrio: forma de
reserva para o enchimento ventricular, em uma situação de stress. Período
isovolúmico: pressão ventricular maior que a da aorta.
Automaticidade: batimento cardíaco autônomo. Com células marcapasso.
Os canais de sódio estão inativos, o próximo a ser ativado é o de cálcio,
entrando sódio.
FLUXO SANGÜÍNEO: a quantidade de sangue que passa por um determinado
ponto da circulação em dado período de tempo.
DÉBITO CARDÍACO: quantidade de sangue bombeada pelo coração no
período de uma unidade de tempo.
PRESSÃO SANGÜÍNEA: força exercida pelo sangue contra qualquer área
unitária da parede do vaso.
COMPLACÊNCIA: quantidade total de sangue que pode ser armazenada em
determinada parte da circulação por mmHg de elevação de pressão.
POTENCIAL DE AÇÃO NO MÚSCULO CARDÍACO: os canais rápidos de
Na+ ocasionam a despolarização; os canais de Ca 2+ e Na+ ocasionam o platô; os
canais lentos de K+ ocasionam a repolarização.
CICLO CARDÍACO: período compreendido entre o começo de um batimento
cardíaco e o começo do seguinte. Cada ciclo se inicia com a geração espontânea
de um potencial de ação no nodo SA. O ciclo cardíaco é composto por um
período de relaxamento, diástole, seguido de um período de contração, sístole.
Os átrios funcionam como bombas de reforço, pois somente 25-30% do
sangue é bombeado, os outros 70-75% de sangue flui naturalmente para os
ventrículos.
O período de diástole ventricular é caracterizado pela queda da pressão
ventricular e aumento da pressão atrial. As válvulas AV se abrem causando o
enchimento rápido dos ventrículos. Neste momento ocorre a sístole atrial
(contração dos átrios).
Durante um período extremamente curto ocorre a contração isométrica
dos ventrículos (início da sístole). As válvulas átrio ventriculares se fecham e as
válvulas S-L ainda não se abriram. Ocorre um aumento na pressão ventricular.
Esse aumento de pressão faz as válvulas SL se abrirem e acontece a ejeção do
sangue dos ventrículos. Ao final da sístole, mas antecedendo o início da diástole
do ventrículo ocorre o relaxamento isométrico. Diminui a pressão ventricular,
aumenta a pressão na aorta e nas artérias pulmonares, as válvulas SL se fecham
e as válvulas AV ainda não estão abertas.
REGULAÇÃO DA ATIVIDADE CARDÍACA:
1) RETORNO VENOSO: quantidade de sangue que chega ao coração.
DC = FC x VS
PA = CD x RT
DC é a quantidade de sangue que sai do coração por minuto. Na venoconstrição
Na venoconstrição chega mais sangue ao coração, aumentando o DC.
RETORNO VENOSO = DÉBITO CARDÍACO
2) MECANISMO DE FRANK-STARING: (regulação intrínseca) é a
capacidade do coração de aumentar sua força de contração, ao passo que o
volume de sangue que chegou até ele (retorno venoso) também aumentou.
3) REGULAÇÃO NERVOSA: no SNA.
Simpático: excita o músculo cardíaco (libera noradrenalina). É capaz de regular
a freqüência dos átrios e ventrículos. Aumenta a freqüência cardíaca e a força de
contração, aumentando o débito cardíaco, portanto, aumentando a PA.
A noradrenalina é vasoconstritora e liberada pelo simpático. Ela se
liga a 2 tipos de receptores: α e β. O α é vasoconstritos e o β é vasodilatador.
Nos músculos os receptores são β.
Parassimpático: inibe o músculo cardíaco (libera acetilcolina). Inerva só o átrio
e por isso diminui a freqüência, a força de contração, o débito cardíaco e a PA.
RITMICIDADE CARDÍACA: determinada pelas células marcapasso que
geram impulsos rítmicos para contração.
O coração possui um sistema especializado a excitação – condução do
impulso elétrico.
1) NODO SINO-ATRIAL: parte do coração localizada na região
superolateral do Ad, logo abaixo da abertura da veia cava superior. Possui
fibras musculares extremamente diferenciadas quase sem elementos de
contração. Especializadas em gerar impulsos rítmicos. Em suas membranas
plasmática há contínua vazão da Na+.
2) VIAS INTERNODAIS: anterior, mediana e superior que conduzem o
impulso cardíaco pelos átrios.
3) NODO ÁTRIO-VENTRICULAR: está situado na parte posterior da
parede septal do AD, logo atrás da válvula tricúspide. Causam retardo
fisiológico. Essas fibras oferecem um pouco de resistência ao potencial de
ação. Evita que os ventrículos contraiam ao mesmo tempo que os átrios.
4) FIBRAS ÁTRIO-VENTRICULARES: também ajudam a retardar o
impulso cardíaco.
5) FIBRAS DE PURKINJE: fazem a propagação imediata e com altíssima
velocidade do impulso cardíaco para todo o ventrículo
SISTEMA VENOSO:
1) PRESSÃO VENOSA: pressão venosa central = pressão atrial direita. Essa
pressão é regulada pela capacidade do coração em bombear o sangue para
fora do átrio.
Aumento da pressão venosa: no caso de transfusões, pois o volume
de sangue aumenta.
Diminuição da pressão venosa: em casos de hemorragia
2) BOMBA VENOSA: a contração da musculatura comprime a veia
bombeando o sangue. A presença de válvulas impedem o retorno do sangue.
Bombeamento aumenta a pressão venosa. Em uma pessoa parada (imóvel)
não ocorre bomba venosa. Veias varicosas prejudicam ainda mais a bomba
venosa.
3) VEIAS COMO RESERVATÓRIOS: 60% do sangue está nas veias.
Diminuição do volume de sangue e diminuição da PA estimula os
quimioreceptores, ocorrendo um reflexo nervoso simpático, com contração
das veias, aumentando a PA e o retorno venoso.
MICROCIRCULAÇÃO:
O leito capilar é extremamente delgado, com uma camada de células
endoteliais justapostas, formando fendas. Assim a difusão de substâncias é mais
fácil.
O capilar tem bastante resistência em sua parede, quando o sangue passa
com a pressão hidrostática (35 mmHg), o plasma extravaza formando o líquido
intersticial que tem forma de gel por causa das proteínas intersticiais.
As proteínas dos poros capilares são negativas e as proteínas plasmáticas
também são negativas, assim essas proteínas plasmáticas não passam para o
interstício. Essas proteínas plasmáticas também não passam para o interstício
porque elas são muito grandes. A força que elas fazem para tentar sair dos
capilares gera uma pressão osmótica, que no caso das proteínas chama-se
pressão oncótica, que só ocorre nos capilares com relação ao interstício.
A pressão oncótica tende à entrada de líquido nos capilares (28mmHg). A
pressão hidrostática tende à saída e plasma dos capilares (35mmHg). A
resultante é a saída de fluido até que essas 2 pressões se igualem.
As trocas nos capilares são feitas por difusão e não por filtração.
A pressão cai porque as trocas são feitas por difusão, por causa da
resistência ao fluxo, dissipando energia.
Os 10% do sangue restante é reabsorvido pelo sistema linfático. Todo o
fluido coletado é devolvido no leito venoso na veia subclávia. O vaso linfático
tem células endoteliais valvuladas com capacidade contrátil.
REGULAÇÃO DE FLUXO SANGÜÍNEO:
Intrínseca – não depende de inervação. Não se sabe certamente quais os
mecanismos intrínsecos de regulação, existem somente teorias.
O aumento do metabolismo do corpo significa aumento da quebra de
ATP formando adenosina difosfato (ADP+P), CO2, hidrogênio e água que são
os metabólitos dessa quebra. Isso aumenta o consumo de O2.
Teoria metabólica: o CO2, ADP+P e H+ são vasodilatadores.
O fluxo capilar é intermitente por causa do esfíncter que determina a
entrada ou não de sangue nos capilares. O fluxo sangüíneo pode aumentar com
a abertura mais prolongadas dos esfíncteres.
Na teoria metabólica esses vasodilatadores irão abrir o esfíncter
aumentando o fluxo, até que a quantidade desses metabólitos se iguale
novamente. Quando a quantidade desses metabólitos aumenta, o fluxo aumenta
por causa da vasodilatação, ele vai aumentando até se igualar à quantidade
necessária de sangue para lavar o corpo desses metabólitos. Depois que o
sangue lava esses metabólitos o fluxo normaliza.
Teoria da demanda de O2: nela o O2 é vasoconstritor e quando aumenta o
metabolismo, aumentando o consumo de O2 e as células irão necessitar de muito
O2, como o O2 livre diminuiu, ocorreu vasodilatação e, portanto, aumento do
fluxo sangüíneo pois aumenta a quantidade de esfíncteres contraindo-se, assim
desloca o sangue para onde necessita.
Quando apertamos o pulso, a mão fica branca, quando soltamos, a mão
fica vermelha porque aumentou o fluxo para limpar a quantidade de metabólitos
presentes na mão. Essa vermelhidão se chama hiperemia reativa.
Hiperemia reativa é quando aumenta o fluxo de sangue depois de uma
interrupção. Hiperemia ativa é quando aumenta o fluxo de sangue por causa do
aumento do metabolismo.
AUTOREGULAÇÃO: quando varia a pressão, varia o fluxo.
Metabólica: aumento da pressão resulta em aumento do fluxo que limpa
o CO2, ocorrendo vasoconstrição, regulando o fluxo de novo. Quando aumenta
a pressão, aumenta o fluxo, aumentando a quantidade de O 2. A quantidade de
O2 é maior que as células necessitam, fazendo com que sobre muito O2
inutilizado, o O2 que é vasoconstritor vai fazer com que o fluxo diminua,
normalizando.
A autoregulação regula a pressão arterial aumentando ou diminuindo o
fluxo. Isso não quer dizer que a pressão vai aumentar ou diminuir, mas sim, o
fluxo vai ser regulado de acordo com a pressão.
A regulação da PA não é feita pelos capilares. Essa autoregulação vai
diminuir o número de alças de capilares, fazendo com que diminua o fluxo.
Miogênica: (não é válida) se aumenta a PA, aumenta a força na parede
dos vasos e ele vai constritar cada vez mais, de acordo com o aumento da PA,
até a morte. Nessa teoria quem regula é o movimento da parede dos vasos.
REGULAÇÃO DA PA A LONGO PRAZO: o organismo vai responder a
longo prazo. Quando abaixa a pressão, diminui o fluxo sangüíneo, diminuindo a
quantidade de O2, provocando a angiogênese (formação de novos vasos). Quem
tem pressão alta, o número de vasos diminui, porque eles fibrosam.
REGULAÇÃO DA PA A CURTO PRAZO: através de mecanismos do SNC.
Para a regulação da PA, é necessário ou regular o débito cardíaco, ou a
resistência ou os dois juntos. Todas essas regulações são para garantir o fluxo.
O SNA controla e regula a PA, e por conseqüência, o fluxo. Só o
simpático inerva vasos.
No SNC, entre o bulbo e a ponte, existe uma formação reticular, com
células que regulam várias funções (respiração, digestão, etc.). Dentro da
formação reticular está o centro vasomotor que se subdivide em regiões:
C1 – vasoconstritora + simpático
A1 – dilatadora , tem um efeito inibitório a C1
A2 – moduladora, porque recebe informações da periferia de quanto é a PA e
envia para C1 e A1.
NMDV (núcleo motor dorsal do vago): a principal eferência ao
parassimpático. É um estimulador do parassimpático e recebe informações da
periferia de quanto está a PA.
Existem regiões que controlam o centro mesencefálico. Essas regiões
(sistema límbico) controlam o centro vasomotor, aumentando ou diminuindo a
PA. Ex: se ouvirmos um alarme de incêndio, esse sistema límbico irá aumentar
a PA.
Nosso sistema de regulação funciona como um sistema de alça fechada.
Por exemplo: aumentou a PA, isso fez com que o receptor para o aumento da
PA seja ativado, enviando uma aferência ao SNC que responderá com uma
eferência estimulando o coração e os vasos que são as células efetoras da
resposta ao aumento da PA.
TIPOS DE RECEPTORES:
Baroceptor: função de estiramento, localizam-se no arco da aorta e no seio
carotídeo. Percebe o grau de estiramento da artéria e envia essa informação
pelos nervos vago e glossofaríngeo. Eles tem um sistema inibitório sobre o
sistema vasomotor (sistema simpático). Se a PA aumenta, os baroceptores
enviam a mensagem para o SNC, que inibe I simpático. Regulam agudamente a
PA, e atuam por no máximo 48 hrs. Mas eles trabalham constantemente para
que o organismo tenha o mínimo de variação a PA.
Quimioreceptores: o tecido que recebe mais sangue, e são muito sensíveis à
quantidade de O2 que chega nos vasos. Eles localizam-se no mesmo local que os
baroceptores. Atuam de forma contrária aos baroceptores. Só atuam quando a
PA cai a 60 mmHg. Quando a PA diminui, os quimioreceptores enviam a
mensagem para o centro vasomotor que ativa o sistema simpático, aumentando
a PA.
Só haverá variação da PA se variar o DC ou a R, ou os 2 juntos.
REGULAÇÃO DA PA A MÉDIO PRAZO: através de hormônios. É o mais
importante (SRAA).
Quando diminui a PA, ocorre uma pequena queda no fluxo e diminui a
filtração nos capilares glomerulares (que são mais porosos que os capilares
normais).
Na filtração, como esses capilares são muito porosos, passa muito sódio,
água e outras substâncias. Quando diminui a filtração, diminui a quantidade de
sódio. Essa diminuição é percebida pelo aparelho justaglomerular. As células
desse aparelho justaglomerular vão liberar renina, que vai atuar sobre a proteína
chamada angiotensinogênio (que existe no plasma).
A renina quebra o angiotensinogênio, formando a angiotensina I. Quando
a angiotensina I passar nos pulmões, outra enzima é liberada
( ECA–enzima conversora de angiotensina I), quebrando a angiotensina I,
fomando angiotensina II.
Essas 2 angiotensinas são vasoconstritoras, e por isso, quando está no
sangue, aumenta a PA.
Renina
vasoconstritora ( R)
Angiotensinogênio -- angiotensina I -- angiotensina II
Aldosterona ( DC)
Esses dois fatores juntos (vasoconstrição e a aldosterona) aumentam a
PA.
A angiotensina II, além de ser vasoconstritora, ela estimula a produção e
liberação de aldosterona.
A região glomerulosa do cortez cerebral libera aldosterona, que estimula
a reabsorção de H2O e Na+ pelos rins. Quando ocorre essa absorção, eles voltam
para a corrente sangüínea aumentando o leito venoso, aumentando assim o DC e
por conseqüência a PA.
A aldosterona vai fazer com que saia H2O e Na+ dos túbulos renais para o
interstício, do interstício a H2O e Na+ serão reabsorvidos pelo capilar,
aumentando o leito venoso, aumentando o DC.
Se aumenta a PA, diminui a renina e vice-versa
O hormônio ADH (antidiurético) tem um efeito de vasoconstrição. Ele
aumenta a PA em doses fisiológicas.
O ADH aumenta a R, aumentando um pouco a PA. Os baroceptores
(inibem o simpático) vão perceber esse pequeno aumento, provocando uma
braquicardia reflexa (diminuem os batimentos cardíacos), fazendo com que
diminua o DC. Assim, como a resistência está alta e o DC está baixo, o PA é
constante.
Em experimentos o NO é produzido pelas células endoteliais dos vasos
(ele é produzido pela ligação da acetilcolina com seu receptor na membrana das
células endoteliais). O NO então, difunde-se para a musculatura lisa.
O NO pode ser produzido pelo atrito que a hemácia faz nas células
endoteliais dos capilares. Quando aumenta o fluxo, aumenta a velociadade do
sangue e as hemácias passam com maior atrito nas células endoteliais.
REGULAÇÃO DA PA A LONGO PRAZO: através dos rins.
Quando aumenta a PA, aumenta a excreção (perda) de H2O e sal pelo rim.
Isso faz com que diminua o volume de sangue, diminuindo o DC, e também a
PA. Esse processo se chama natriurese pressórica.
Nas capilars peritubulares (próximos ao túbulo do rim) a pressão oncótica
(puxa palsma para dentro dos capilares) é maior que a pressão hidrostática,
fazendo com que haja reabsorção. Mas quando aumenta a PA, a pressão
hidrostática também sobe, mas a pressão oncótica é constante, isso diminui a
capacidade de reabsorção de excretas desses capilares, e esses excretas vão para
os tubos renais, sendo escretados. Como diminui a quantidade de excretas
reabsorvidos, diminui a quantidade deles no leito venoso, diminuindo o DC e a
PA.
SISTEMA RESPIRATÓRIO
A principal função do sistema respiratório é fornecer O 2 e eliminar CO2
para manter o pH em níveis constantes. O O2 é necessário para síntese de ATP.
Composição do ar: O2: 21%, CO2: 0,3% e N2: 76%. A pressão
atmosférica do ar cai determinar a quantidade de O2 que entra no corpo.
Se a pressão no alvéolo é alta, os choques das moléculas são miores e as
moléculas de O2 vão para o sangue. Se a pressão no alvéolo é baixa, os choques
são menores e as moléculas não se difundem muito para o sangue.
O ar , no nariz, tem um fluxo turbilhonante e é aquecido. O ar aquecido
muda sua composição. A temperatura influencia na pressão do oxigênio.
Nós temos sempre a mesma composição do ar porque temos uma
temperatura constante.
Os alvéolos possuem uma grande superfície de contato, são delgados e
propícios às trocas gasosas. As trocas gasosas são feitas nos ductos e alvéolos.
O resto do trato respiratório é chamado trato morto porque não efetua trocas
gasosas.
Na inspiração, ocorre o aumento da caixa torácica, diminuindo a pressão,
fazendo com que o ar entre. Na expiração, o volume da caixa torácica volta ao
normal (os músculos relaxam), aumentando a pressão interna e o ar sai.
A renovação do ar alveolar não é imediata, acontece aos poucos. O ar que
entra se mistura aos poucos. Mas sempre fica um pouco de ar nos pulmões para
garantir as trocas constantes de gases.
Quando se inspira, a pressão do ar é 160 mm Hg e a pressão alveolar é
105 mm Hg. A pressão do ar na expiração é 120 mm hg. Isso ocorre porque o ar
inspirado fica no espaço morto e o ar que será expirado passa pelo espaço
morto, misturando os 2 ares e a pressão que estava 105 mm Hg foi para 120 mm
Hg.
O líquido surfactante mantém os alvéolos abertos.
A tensão da pressão do alvéolo é muito grande, se o alvéolo fechar as
suas paredes grudam. O ar não sai do alvéolo, o O 2 se difunde até o ar álveolar e
até o sangue.
Quando o sangue entra na circulação pulmonar a PO 2 é 40 mm Hg. A PO2
do alvéolo é 105 mm Hg. O sangue chega no capilar pulmonar a PO2 40 mm Hg
e rapidamente satura, chegando a PO2 105 mm Hg.
Mesmo quando aumenta o metabolismo, o sangue consegue ser
oxigenado.
Quando o sangue venoso sai do coração, parte dele vai para o espaço
morto (para oxigenar essa parte) e o resto vai ser oxigenado nos alvéolos.
Quando o sangue sai do pulmão para o coração, junta o sangue que havia no
espaço morto com o sangue oxigenado dos alvéolos. Essa mistura de sangue faz
com que haja uma diluição na concentração de O2 no sangue. A PO2 cai de 105
mm Hg para 97 mm Hg.
TRANSPORTE DE O2
Pigmentos respiratórios: metais, o que importa não é a cor e sim a
capacidade de transportar substâncias. Em mamíferos o pigmento respiratório é
a hemoglobina.
O CO2 se liga mais fortemente na hemoglobina que o O2. a hemoglobina
tem 4 sítios de ligação com o oxigênio.
Hiperventilação: aumento da freqüência respiratória mais que o
necessário.
O O2 dissolvido no plasma não é suficiente para a sobrevivência.
Quando a pessoa hiperventila e prende a respiração, como é o caso de
mergulhadores, na hiperventilação o O2 está 100%, isto é, está ligado nos 4
sítios de ligação da Hb e o CO2 está muito baixo (o CO2 que sinaliza a vontade
de respirar). Como o Co2 está baixo, não se sente vontade de respirar e enquanto
isso o O2 está sendo consumido. Como o O2 e o CO2 caem ocorre o desmaio.
Quando a Hb liga-se ao O2 e ao CO2 ela se modifica estruturalmente,
sendo mais ou menos associável ao O2 ou CO2
CURVA DE DISSOCIAÇÃO Hb/O2
Só é possível medir com certeza a afinidade da Hb com o O2 em 50%
(P50).
P50: PO2 na qual 50% da Hb está saturada no O2. É um artifício
matemático para indicar o valor de 50% de saturação, já que não dá pra medir
100%.
Fatorem que ativam a variação da curva: PCO2 e pH.
A afinidade da Hb com O2 pode estar aumentada ou diminuída, variando
a P50 e a curva para a direita ou para a esquerda.
Bohr:
Aumento do PCO2 + diminuição do pH = diminuição da afinidade
Aumento do pH + diminuição do PCO2 = aumento da afinidade.
Aumento da temperatura resulta na diminuição da afinidade, liberando O 2
para os tecidos.
Os tecidos doam CO2 para a Hb, aumentando a afinidade do CO2 na Hb,
diminuindo a afinidade do O2 na Hb, isso faz com que o O2 se disprenda da Hb,
indo para os tecidos.
Quando aumenta a temperatura, aumenta o metabolismo e o consumo de
O2. Nesse caso, a afinidade do O2 na Hb diminui para que o O2 seja liberado
para os tecidos.
Efeito Bohr: quando aumenta a PCO2 a curva varia para a direita
(tecidos), quando diminui a PCO2 a curva varia para a esquerda (pulmões). Isso
ocorre constantemente.
A hipoxia (falta de O2) estimula a hemácia a produzir 2,3 DPG (atua na
Hb), que desvia a curva para a direita. A liberação de O2 para os tecidos é mais
fácil.
DPG é uma substância fabricada pela hemácia que atua na Hb,
diminuindo a afinidade da Hb ao O2, liberando mais O2. O DPG é produzido em
casos onde a pressão atmosférica é baixa, diminuindo a quantidade de O 2 no ar
(por exemplo no Peru).
O CO2 é constantemente produzido pelas células, aumento da PCO2 ele é
transferido para os pulmões para ser liberado.
O CO2 está 7% livre no plasma pois se dissolve com grande facilidade. O
CO2 se difunde rapidamente na hemácia, e daí pode ter 2 caminhos: um é se
difundir com a Hb (23%) e o outro é CO2 + H2O que é catalisada pela anidrase
carbônica, formando ácido carbônico.
Essa reação CO2 + H2O
H2CO3 só ocorre com a presença da emzima
catalisadora anidrase carbônica.
O H2CO3 libera H+, formando HCO3- e H+. Esse H+ se liga na Hb (no seu
respectivo sítio de ligação). O HCO3- se liga a uma proteína transportadora,
sendo levado para fora da célula e no seu lugar entra Cl- (70%)
O Cl- é osmoticamente ativo. Quando o Cl - entra na célula, aumenta o
número de íons nessa célula, aumentando o volume da célula.
Hematócrito é a quantidade proporcional de células e plasma na sangue.
O hematócrito do sangue venoso é maior que a do sangue arterial, pois o
volume das células venosas é maior.
Quando a hemácia chega no alvéolo, o CO2 livre se difunde para o
alvéolo, quebrando o equilíbrio (7%, 23%, 70%), isso daz com que o CO 2 saia
da hemácia. O pouco de HCO3- que está na célula se liga ao H+, formando
H2CO3, a bomba Cl-/HCO3 inverte, saindo o Cl- e entrando HCO3-, isso libera
mais CO2. esse H+ foi liberado pela Hb devido à formação da oxiemoglobina
que é um ácido forte e dissocia mais facilmente.
Quando a Hb + O2, a Hb libera mais facilmente o Co2 (diminui a
afinidade).
CO2 + H2O
H2CO3
HCO3- + H+
O pulmão auxilia a regulação do pH. Quando aumenta o pH, aumenta o
H livre e ele se liga ao HCO3- livre (que está em maior quantidade que o H+)
formando H2CO3, liberando CO2 pelos pulmòes, regulando portanto essa
variação do pH.
O HCO3- livre (que está em alta quantidade no corpo) é uma solução
tampão (o pH é estabelecido), ele atua para manter o equilíbrio do pH, quando o
H+ se liga ao HCO3- formando H2CO3 que libera CO2.
O pH 7,4 é mantido pela quantidade de H+ livre no plasma. O organismo
depende da pH 7,4 e quando ocorre a variação, ou seja, aumento de H+, o pH
tem que ser mantido a 7,4, para isso existe o HCO3- livre.
Quando o nível de H+ diminui, a freq”:uência respiratória diminui,
acumulando CO2 nas células e aumentando H+.
+
REGULAÇÃO DA RESPIRAÇÃO
Existe um padrão automa’tico para a respiração. Para que haja respiração
tem que haver potenciais de ação para que o músculo esquelático torácico seja
estimulado.
O núcleo respiratório dorsal exibe atividade durante a inspiração, e na
expiração exibe repouso.
O músculo (diafragma) é contraído até certo ponto.
Quando acaba o potencial de ação do dorsal, o centro pneumotoráxico
entra em atividade. É o centro pneumotoráxico que determina o fim da
inspiração. O núcleo dorsal e o centro pneumotoráxico trabalham juntos para a
respiração.
O reflexo de Heing-Brewer limita a alta inflação do pulmão. Ele corrige
uma super inflação do pulmão. Atua quando ele já atuou muito e o reflexo
interrompe a continuação da inflação do pulmão acima da sua capacidade.
Quando a respiração fica mais rápida (em exercício)entra em atuação o
núcleo ventral, que emite potenciais de ação.
Corpo em repouso – núcleo dorsal
Corpo em exercício – núcleo ventral
A regulação da respiração em seres terrestres é feita pelo CO 2, porque o
CO2 em excesso muda o pH do organismo.
Controle químico: através dos quimiorreceptores centrais e ventrais.
Esses quimiorreceptores vão perceber acúmulos de CO2 no corpo e estimular
uma resposta. É preciso manter constate o nível de CO2.
Quimiorreceptores centrais são sensíveis ao pH e ao CO 2. se aumenta a
PCO2 e/ou diminui o pH, aumenta a freqüência respiratória. O único detalhe é
que esses quimiorreceptores estão na barreira hemato-cefálica (os capilares do
cérebro são bem fechados, formando essa barreira). O sangue passa nesses
capilares com alta PCO2, esse CO2 se difunde para o líquido que banha o SNC,
revertendo a reação CO2 + H2O H2CO3
HCO3- + H+, esse H+ estimula os
quimiorreceptores para aumentar a freqüência respiratória e limpar o CO2. Mas
o H+ não passa dos capilares para atingir os quimiorreceptores por causa da
barreira, só o CO2 passa porque ele se difunde facilmente.
Quimiorreceptores periféricos são sensíveis à quantidade de O 2. são
ativados quando PO2 cai abaixo de 60 mm Hg. Atuam em uma situação
extrema.
Regulação superior: é um controle voluntário da respiração. Ele acontece
até um certo limite, em casos de prender a respiração.
SISTEMA DIGESTÓRIO
A função da digestão é quebrar substratos em partículas menores para
facilitar a absorção.
As vitaminas não são fontes de energia. As fibras são importantes para
aumentar o bolo fecal e facilitar a eliminação desses resíduos.
BOCA: com mucosa e amilase salivar (pH 6,7) que é responsável pela digestão
do amido. A digestão do amido começa na boca e termina no estômago.
ESTÔMAGO: possui secreções ácidas em sua parede e o movimento
peristáltico do estômago faz com que o bolo alimentar se movimente em
círculos dentro do estômago e se misture com essa secreção. O alimento
deglutido demora para se misturar completamente com o ácido do estômago e
enquanto ele não se mistura, a amilase ainda está digerindo o amido (enquanto o
pH ainda for + ou – 6,0).
Enzimas do estômago: pepsina, formada pela ligação do pepsinogênio
com HCl. O pepsinogênio cai na luz do estômago, se liga ao HCl e se
transforma em pepsina. Ela não é produzida na parede do estômago pois senão
ela destruiria a parede interna do estômago. A pepsina quebra o tecido
conectivo da carne (principalmente fibras colágenas), ou seja, a superfície, para
depois a carne ser mais quebrada.
Quanto maior for a relação superfície/volume, mais rápido o alimento
será digerido. Ex: quanto menor for o tamanho do alimento, mais rápido ele será
digerido.
A tributase age na gordura da manteiga.
INTESTINO DELGADO:
Duodeno: presença se suco pancreático da parte exócrina do pâncreas. Esse
suco é composto por amilase pancreática (quebra amido), lípase pancreática
(quebra lipídeos), lipsina, quimiotripsina e carboxipeptidase (quebram
proteínas) e HCO3-.
A amilase quebra até a forma de dissacarídeo. E a lipsina, a
quimiotripsina e a carboxipeptidase quebram até dipeptídeos.
O nosso corpo não absorve dissacarídeos nem dipeptídeos, eles precisam
ser quebrados. No intestino existem microvilosidades e as células de sua parede
produzem enzimas dissacaridases e dipeptidases que quebra em
monossacarídeos e monopeptídeos. Depois dessa quebra ocorre a absorção.
Essas últimas enzimas estão misturadas no muco das microvilosidades.
ABSORÇÃO DE MONOSSACARÍDEOS: glicose e galactose – transporte
ativo secundário. Frutose – difusão facilitada.
Glicose e
galactose
Na+
K+
Na+
Na+
As secreções biliares são produzidas pelo fígado. Os sais biliares são
formados pela bilirrubina. A secreção biliar é importante excretora de sais
biliares. Como a bilirrubina é tóxica em altas quantidades ela precisa ser
eliminada, é por isso que as fezes apresentam cor marrom.
A bilirrubina (age como um detergente) é importante porque liga a
gordura na água, formando uma emulsão, isso é para a eliminação. Micelas:
fragmentos de gorduras + bilirrubina.
A lípase atua junto com as colipases, quebrando triglicerídeso em ácidos
graxos e um pouco de monoglicerídeos que batem na parede do intestino e se
difundem para dentro das células. Quando ele entra nas células, essas
substâncias sofrem reesterificação pelo REL para formar triglicerídeos. Esses
triglicerídeos se juntam com o colesterol e com fosfolipídeos formando o
quilimícron.
O quilimícron sai das células e cai no sistema linfático e depois para o
circulatório.
A vantagem do armazenamento de gordura é que ela é mais energética
que o glicogênio.
CAMADAS DO TUBO DIGESTIVO:
Luz do tubo – região mucosa – região sub-mucosa com glândulas exócrinas –
músculo circular – feixes longitudinais – plexos mioentérico e submucoso.
Mesmo quando o tubo digestivo está vazio ele exibe contrações que
mantém o fluxo das secreções (porque a parede descama), mantendo o sistema
limpo.
Quando a glicemia abaixa, as contrações aumentam (são as contrações de
fome). A taxa de glicemia determina também as contrações do sistema.
Nas células do músculo liso do tubo não há potencial de ação, e sim uma
variação da voltagem.
Quando a onda chega no pico, quando o tubo está vazio, não há potencial
de ação porque está a –50 e não há limiar de disparo. Quando o tubo está cheio,
neurormônios são liberados, aumentando essas ondas para –40 e aí, no pico da
onda há o potencial de ação.
SNE – sistema nervoso entérico. Contém dois plexos: o mioentérico e o
submucoso.
Na mucosa das paredes existem receptores para estímulos mecânicos e
químicos. O estímulo é sentido por esses receptores sobre a chegada do
alimento e ativa os dois plexos que determinam a contração do músculo. O SNE
é completamente independente do SNC.
O alimento chega no tubo, ativa os receptores que ativam o SNE (plexos)
e estes emitem a resposta que é a contração muscular.
Esses receptores também ativam o SNC que modula o SNE via SNA
(simpático e parassimpático).
O SNA funciona assim: o simpático inibe a atividade do sistema, o
parassimpático estimula a atividade do sistema. O parassimpático libera
acetilcolina que estimula os potenciais de ação. A acetilcolina despolariza as
células. O simpático libera noradrenalina que hiperpolariza a célula.
O SNE, quando estimula o submucoso, ativa a liberação da secreção e
aumenta a motilidade. Como o mioentérico está entre 2 camadas de células, ele
é responsável muito mais para a motilidade que para secreção. O submucoso
ativa mais a secreção que a motilidade.
FLUXO SANGÜÍNEO: artéria mesentérica se ramifica em vasos menores que
banham o mesentério. Conforme o sangue passa pelo vilo vai liberando O 2 e o
lado esquerdo tem mais O2 que o direito, para corrigir isso, ocorre difusão do
capilar arteriolar para o capilar venular.
MOVIMENTOS:
Mastigação: é voluntário, mas existe um estímulo reflexo. O alimento na boca
encosta no palato, ativando os receptores que levam a ativação para o tronco
cefálico (centro de deglutição) e a maxila cai, nisso o estímulo pára e a maxila
volta, que é a mastigação.
Deglutição: o início é voluntário, mas quando o alimento pressiona o palato
mole ele ativa o centro de deglutição e o movimento se torna involuntário.
Quando começa a engolir, as dobras palatofaringial diminuem o tamanho
da garganta para garantir que o alimento não fique intalado e para garantira eu
só passe alimento de um tamanho pequeno. A epiglote fecha par impedir que o
alimento não entre no sistema respiratório, por isso, não há entrada nem saída
de ar na respiração.
Esôfago - 1˚ esfíncter esofagiano que abre e fecha levando o alimento em
direção ao estômago. Se essa primeira onda não leva o alimento diretamente
para o estômago, existe a segunda onda que é muito forte.
O estômago tem um pH de trabalho em torno de 3, por isso que ele tem mucosa
para proteger esse ácido. Esse segundo esfíncter é bem fechado para evitar a
passagem de HCl no esôfago. O HCl do estômago corrói a parede do esôfago,
por isso que o esfíncter é bem fechado.
Quando o alimento chega no estômago ele vai primeiro para o fundo que
é a região de armazenamento. Começam as contrações do estômago em direção
ao esfíncter pilórico, mas o alimento não entra de uma vez no duodeno mas sim
aos poucos. O alimento que não passa para o duodeno volta, se misturando com
o HCl.
Depois de bolo alimentar, a partir do estômago, o alimento passa a se
chamar quimo.
Intestino delgado: movimentos peristálticos e segmentação que é uma mistura.
Controle neuro-endócrino: pelo SNA. Simpático inibe e parassimpático
estimula.
Regulação endócrina: o estômago produz o hormônio gastrina que
aumenta a motilidade estomacal e o quimo vai para o duodeno.
O duodeno produz CCK que inibe a motilidade gástrica. O alimento vai
passando pelo duodeno e quanto mais ele sai do duodeno, menos CCK produz.
Por isso que o sistema é paulatino, ou seja, o duodeno que controla o fluxo do
alimento aos poucos.
SECREÇÕES DIGESTIVAS:
Saliva: componente mucoso – parte líquida, componente enzimático – amilase
salivar.
Glândulas parótidas, submandibulares e sublinguais, várias e pequenas
glândulas espalhadas.
A saliva inicial é muito parecida com o plasma, é fornecida nos ácinos e
avança nos ductos. A saliva é hipotônica em relação ao plasma. Intensa
reabsorção de Na+ e reabsorção de bicarbonato (HCO3-) e K+.
Regulação da secreção salivar: quando se observa alguma coisa, pelo cheiro,
fome; acaba excitando o núcleo salivatório (regulação direta com a glândula
salivar) entre o bulbo e a ponte = regulação neural, não há regulação endócrina
para a secreção salivar.
Liberação de bradicinina – vasodilatador local, quanto maior o fluxo
sangüíneo maior a quantidade de plasma. A bradicinina atua sobre os capilares
periféricos à glândula salivar.
Esôfago: não produz muco.
Estômago (secreção estomacal): o estômago é composto por dois tipos de
glândulas que são compridas e liberam sua secreção na luz do estômago.
Glândula oxíntica com células parietais que produzem HCl, células
mucosas que produzem muco e células pépticas que produzem pepsinogênio.
Glândula pilórica com células parietais (HCl), células mucosas (muco),
células G que produzem gastrina que estimula a secreção de HCl.
Regulação da secreção gástrica: o simpático inibe e o parassimpático
estimula, ele estimula o SNE que estimula a glândula.
A secreção gástrica tem três fases:
Fase cefálica: quando você vê o alimento, aumentando a secreção de HCl no
estômago. Essa estimulação provém do nervo vago que é a principal eferência
do parassimpático e estimula o SNE que aumenta a secreção de HCl.
Fase gástrica: quando o alimento chega no estômago (distende). Estimulação
mecânica e química, chagando a informação no SNE e SNA. Regulação
hormonal do parassimpático a regulação de HCl pelo estômago dá-se por:
Parassimpático, gastrina, CCK, que estimulam a secreção enzimática e secretina
que estimula a liberação de bicarbonato. Todos esses 4 fatores vão atuar juntos.
Fase intestinal: o alimento chega no intestino que estimula a secreção de
gastrina, CCK e secretina. A gastrina tem efeito estimulatório no estômago
(estimula a secreção gástrica de HCl) e no pâncreas. O duodeno produz, além do
CCK a gastrina que estimula o estômago, porém toda a resultante ainda vai ser a
inibição da gastrina pelo CCK, já que sua produção é bem maior que a da
gastrina; tem maior efeito inibitório do que estimulatório da gastrina.
A histamina assim como a gastrina, estimula a secreção gástrica (HCl) e
tem um efeito sinérgico, ou seja, age com a gastrina aumentando e muito a
produção de HCl (efeito multiplicativo) mais do que os dois separadamente. A
histamina atua em diferentes receptores; ex: no estômago (H2) e no capilar (H1).
A secreção intestinal propriamente dita é apenas eletrólitos e íons.
METABOLISMO: reações de síntese e degradações de produtos, reações
necessárias para manter nosso corpo vivo.
Metabolismo basal: reações químicas que acontecem quando o indivíduo não
está com frio ou calor, em repouso sem estar realizando atividades físicas e sem
sofrer digestão.
À medida que você gasta ATP, mais ATP é sintetizado. A glicose entra
na célula mais facilmente quando o corpo está em exercício.
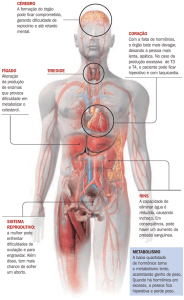
FISIOLOGIA ENDÓCRINA
Funções do sistema endócrino: reprodução (hormônios reprodutivos),
desenvolvimento e crescimento, manutenção do meio interno e produção,
armazenamento e utilização dos substratos ou metabólitos energéticos.
Glândula Tiróide: seus hormônios atuam sobre o metabolismo e são
responsáveis por quase todos os tecidos. Quando acabamos de comer,
hormônios são liberados para aumentar o metabolismo e quando passa um
tempo, o metabolismo volta ao normal por causa dos hormônios.
A ação do Sistema Nervoso é rápida e a ação do Sistema Endócrino é
mais lenta e mais duradoura, embora haja exceções (adrenalina e
noradrenalina).
O ciclo menstrual é alterado pelo ciclo claro-escuro. Conclui-se que o SE
está sob controle do SN. A luz é captada pelo olho e é transmitida para o SE,
através de alguns nervos ópticos ligados ou SE.
Como o SN tem uma linguagem elétrica, ele consegue controlar o SE
através de sinais que o SN emite. Quem converge esse sinal do SN para o SE é
o hipotálamo.
Hipotálamo:
O hipotálamo está no tronco encefálico, abaixo do 3˚ ventrículo, ligado à
glândula hipófise. Como o hipotálamo é muito irrigado, ele controla carias
funções.
No hipotálamo, os neurônios vão liberar hormônios. Existem núcleos
neuronais para hormônios específicos. O hormônio é liberado através de
potenciais de ação.
O hipotálamo vai liberar suas secreções sobre a hipófise. A hipófise é
quem recebe os hormônios do hipotálamo e os libera para o corpo. O
hipotálamo não é considerado glândula endócrina.
Sistema porta-hipotálamo-hipofisário: liga o hipotálamo a adenohipófise. Sistema de vasos responsável pelo transporte de hormônios do
hipotálamo para a adeno-hipófise.
Vem um estímulo nervoso para a produção de um hormônio no núcleo
hipotalâmico, é nesse local que estão os corpos celulares que produzem o
hormônio. Esse hormônio caminha pelo axônio e devido à abertura dos canais
de Ca2+, ocorre exocitose do hormônio que vai para a adeno-hipófise e se
difunde na rede capilar.
Tipos de hormônios: protéicos (hidrossolúvel), colesterol (lipossolúvel) e
aminoácidos (hidrossolúvel).
Adeno-hipófise:
A hipófise fica bem atrás do nariz e o acesso a ela é pela boca. A hipófise
possui duas glândulas distintas, mas associadas: a adeno-hipófise e a neurohipófise.
Adeno-hipófise: atua sobre a glândula adrenal, tireóide, gônadas, produz
GH e prolactina.
-Hormônio do crescimento (GH): o GH têm um efeito mitogênico sobre todas as
células, ele favorece as sínteses protéicas. O GH:
Aumenta o transporte de aa intracelular
Aumenta a transcrição do DNA, gerando RNAm
Ativa os processos traducionais (transformação de RNAm em
proteína)
O GH tem um efeito anabólico (armazenador) sobre a síntese de proteínas
e também tem um efeito hiperglicemiante (o GH isolado), mas não manifesta
hiperglicemia pois a insulina que é hipoglicemiante age equilibrando essa
diferença.
GH: anabólito de proteínas e catabólito de glicose e gorduras.
O GH vai atuar no fígado que produz um fator de crescimento (IGF ou
somatomedina) que cai na corrente sangüínea e ocasiona o crescimento.
Regulação da secreção de GH: feedback (pode ser tanto + quanto-). O
próprio hormônio vai inibir ou estimular a sua secreção para garantir uma
regulação com uma variação bem pequena. Lembrando que nunca “liga” ou
“desliga” e sim aumenta ou diminui. Ex: quanto mais T3 e T4 é produzido, maior
é o efeito biológico.
Sabe-se que o TGF tem um efeito inibidor ou estimulador sobre o GH.
Em casos de hipoglicemia, produz-se GH que tem efeito hiperglicêmico.
Prolactina: produção de leite em seres humanos. (peixes também têm prolactina,
mas para outras funções).
A prolactina existe no corpo em níveis basais tanto em mulheres quanto
em homens. Ao longo da gravidez percebe-se um alto nível de prolactina.
Quando os níveis de estrógeno e progesterona (produzidos pela placenta) estão
altos, não se produz leite. Depois do parto, que é quando esse hormônio
diminui, o leite é produzido.
PRF + Prolactina
PIF
-
Prolactina
Neuro-hipófise:
A neuro-hipófise é uma glândula de origem diferente da adeno-hipófise.
Ela tem origem neural e não secreta hormônios.
Existem dois núcleos hipotalâmicos: o supraótico e o paraventricular.
Eles contém os corpos celulares dos neurônios e esses respectivos axônios estão
direcionados para a neuro-hipófise.
Esses neurônios secretam ADH e oxitocina separadamente (ou o neurônio
é produtor de ADH ou ele é produtor de oxitocina). Em ambos os núcleos
existem neurônios que produzem ADH e outros que produzem oxitocina.
No potencial de ação, os canais de Ca2+ do terminal do axônio se abrem e
o Ca2+ entra e fosforila proteínas que ancoram as vesículas que contém
hormônios, liberando essas vesículas por exocitose. A neuro-hipófise vai liberar
os hormônios produzidos nos núcleos supraótico e paraventricular.
Efeitos biológicos e regulação do ADH: em doses fisiológicas não ocorre
aumento de pressão, ocorre sim a conservação de água. Em doses de emergência
(hemorragia) o ADH vai controlar a PA.
-Antidiurese: ocorre no rim. O sangue é filtrado no glomérulo. Ele filtra
aproximadamente 120 ml/min, mas 90% é reabsorvido no tubo distal e 10% é
ou não reabsorvido no tubo coletor.
Quando bebemos muita água, ela vai para o sangue e dilui a concentração
do plasma. O mecanismo de ação que percebe a concentração do plasma é no
OVLT (órgão vasculoso da lâmina terminal), que é um órgão que possui
osmorreceptores. Esses osmorreceptores percebem a diferença da concentração
de plasma.
Quando o plasma está diluído, entra água nessas células
(osmorreceptores) que mandam uma mensagem para os núcleos supraótico e
paraventricular, diminuindo a produção de ADH. O ADH aumenta a reabsorção
de H2O no tubo coletor do rim.
A PKA ativa proteínas que são “túbulos”, e esses se inserem na
membrana plasmática. Esses “túbulos” são específicos para H2O.
H
R
Fosforila
proteínas
Pr G
Adenil
ciclase
ATP
AMPC
PKA
Fosforila outras
proteínas
LUZ
PKA
AMPC
R
H
O álcool inibe a produção de ADH. Quando se bebe muito álcool, o ADH
não consegue agir. Então diminuem o número de microtúbulos de membrana e
perde mais água que se bebe.
O que regula o ADH é a osmolaridade plasmática e em casos de
emergência a PA.
Oxitocina:
Começa a ter efeito na hora do parto. O estrógeno deixa a célula
despolarizada e a progesterona deixa hiperpolarizada. Esse equilíbrio impede a
contração do útero.
A primeira contração do útero ocorre pois o feto produz corticóides que
promovem a transformação da progesterona em 17-OH progesterona. Isso faz
com que o nível de progesterona diminua ocorrendo desequilíbrio da relação
estrógeno/progesterona e por conseqüência ocorre a contração. Essa primeira
contração é a que ativa a produção da oxitocina. Essa primeira contração emite
uma mensagem para os núcleos supraótico e paraventricular que produzem a
oxitocina. Quanto mais oxitocina é produzida, mais contrações o útero vai
sofrer, até o bebê ser expulso do útero.
A primeira contração do parto é independente da oxitocina e não é
determinada pela mãe, e sim pelo feto.
A oxitocina também é liberada durante a amamentação. E auxilia o
espermatozóide a chegar no óvulo.
GLÂNDULA TIRÓIDE
Localiza-se atrás da traquéia. É uma glândula com dois lobos que estão
ligados por um ístimo que passa pela frente da traquéia. A tiróide é formada por
folículos preenchidos por colóide e células C (que produzem calcitonina). Sobre
a tiróide existem 2 pares de glândulas, as paratiróides que produzem PTH
(hormônio paratiroideano).
Os hormônios tiroideanos são constituídos 90% de iodo, é por isso que é
necessário ter uma boa alimentação de iodo. O iodo é essencial para a síntese de
hormônio tiroideano.
Absorção do iodo: na membrana basal existe um transporte secundário de iodo.
As células possuem esse trocador com transporte ativo secundário de I para
dentro da célula. É necessário também a presença da tireoglobulina, que é uma
proteína enorme com vários aa tirosina. A tirosina têm 2 sítios de ligação com o
iodeto e a ligação do iodeto no sítio de ligação da tirosina é catalisada pela
peroxidase. Quase todos os aa da tireoglobulina têm os 2 sítios ocupados.
2 sítios ocupados: Di Iodo tirosina (DIT)
1 sítio ocupado: Mono Iodo tirosina (MIT)
2 DIT = T4
1DIT + 1 MIT = T3
T3 e T4 existem dentro de uma mesma proteína. A chance de formar T4 é
muito maior que a de formar T3, de formar T2 é ainda mais insignificante. O T3 e
T4 estão ligados na proteína.
Vem o estímulo para liberação de T3 e T4, ocorre pinocitose e entra na
célula uma vesícula com colóide, essa vesícula se liga a um lisossomo (que
contém enzimas proteolíticas) e ocorre a quebra da proteína, liberando T3 e T4.
A reabsorção do I livre é constante.
O T3 e T4 caem na circulação e se ligam a proteínas: pré-albumina
(TBPA) e globulina (TBG). Os receptores de hormônios têm mais afinidade ao
T3 que ao T4. Por isso, o T4 é considerado um pré-hormônio e tem que se
desiodar o T4 em T3 para ele poder atuar. Esse processo ocorre no fígado,
através da desiodase.
Desiodase 1: perto da membrana plasmática. No fígado. Isso faz com que tenha
muito T3 na circulação.
Desiodase 2: perto da membrana nuclear. Ocorre em outras células,
principalmente na adeno-hipófise.
O fígado é uma fonte extraordinária de T3, mas não o acumula.
O receptor do T3 está associado ao DNA (DNA codifica proteínas), na
região reguladora. O T3 ligado ao receptor no DNA vai inibir ou estimular a
transcrição de proteínas. Ex: o T3 ligado no receptor estimula a produção de GH
e inibe a do TSH.
Mesmo quando o receptor de T3 estiver vazio ele tem um efeito. Ex:
quando o receptor está vazio inibe GH e quando está cheio estimula GH.
T3 : ativa o metabolismo. Quando a temperatura do corpo cai, o T 3 é liberado
para ativar o metabolismo e produzir calor. o T3 ativa a calcioATPase que retira
Ca2+ da célula, mas mesmo assim o nível de Ca2+ não se altera porque a
quantidade de Ca2+ que sai, entra. O T3 também ativa quebra de gordura, mas
não altera o nível de gordura do corpo porque essa gordura quebrada sintetiza
depois. O T3 também atua sobre o crescimento e ossificação.
Uma pessoa que não tem T3 possui cretinismo, pois não tem crescimento
do corpo (raquítico) e deficiência mental, pois não ocorreu amadurecimento das
sinapses no cérebro.
Quem inibe a produção de T3 e T4 é o T4, mas em sua forma desiodada,
que é T3.
Bócio: não tem T3 e a tiróide incha porque aumenta a produção de colóide para
receber o iodo e como não tem iodo, acumula colóide.
PÂNCREAS ENDÓCRINO
O pâncreas endócrino é constituído pelas ilhotas de Langerhans que
possuem células alfa, beta e delta. As células alfa produzem glucagon, as beta
insulina e delta somatostatina.
Insulina: hormônio protéico, hidrossolúvel. É constituída por duas cadeias e a
insulina só é ativa quando essas duas cadeias estão presentes.
O receptor da insulina é composto de subunidades 2 alfa (extracelular) e
2 beta (intracelular). Quando o hormônio (insulina) se liga em alfa, ativa beta
que têm propriedade quinase. A quinase tem como propriedade fosforilar outras
proteínas.
EFEITOS: (aumenta a permeabilidade da glicose)
-No músculo: a insulina estimula o transporte de glicose para dentro da célula
(já que a membrana das células musculares é muito impermeável), aumentando
a síntese e a capacidade dos transportadores dessa hexose. Uma pequena fração
dessa glicose é oxidada, mas a maior parte é armazenada com glicogênio,
devido a uma ação específica da insulina ativando a glicogênio sintetase
muscular.
A insulina exerce importante ação anabólica sobre o metabolismo de
proteínas no músculo através dos seguintes mecanismos:
*Estimulando o transporte de aa para o interior da célula muscular,
independentemente do seu efeito sobre o transporte de glicose
*Aumentando a eficiência do processo de tradução, atuando na
etapa de iniciação da síntese protéica
*Inibindo a degradação protéica e a liberação de aa essenciais e
ramificados no músculo.
A insulina ativa a enzima hexoquinase, tornando possível o
processamento e utilização da glicose.
-No fígado: a insulina aumenta a captação de glicose, estimulando seu
armazenamento na forma de glicogênio e sua utilização como substrato para a
lipogênese; aumenta o fluxo na via glicolítica e na via das pentoses; reduz a
produção hepática de glicose e inibe a formação de corpos cetônicos.
A membrana do hepatócito é permeável a glicose, portanto não necessita
de insulina para entrar na célula. O fígado estoca glicogênio, e quando
necessário o glicogênio se transforma em glicose para mandar para o sangue. É
por isso que o fígado é um tampão da glicemia.
Quando se come um doce, a glicose é armazenada no hepatócito sob a
forma de glicogênio. Quando essa quantidade de glicogênio satura na célula, ele
é transformado em gordura.
-No tecido adiposo: a insulina estimula as atividades lipogenéticas e inibe a
motilização de ácidos graxos do tecido adiposo.
-Nas proteínas: a proteína é o último recurso que o corpo pode usar para gastar.
A insulina aumenta a transcrição do DNA em RNAm e a sua transformação
(tradução) em proteínas. A insulina aumenta a permeabilidade dos aa na célula.
-No cérebro: é altamente permeável à glicose. A hipoglicemia torna os
neurônios mais excitáveis.
Efeito cetogênico de falta de insulina: a insulina é quem transforma e estoca a
glicose em glicogênio, quando há falta de insulina, o processo de formação de
glicogênio inverte, liberando glicose para a produção de ATP.
Quando pára a produção de insulina, toda a glicose é utilizada e o corpo
começa a usar gordura (quebra de triglicerídeos). Existe pouco ácido
oxalacético e muita coenzima A, por causa da quebra do triglicerídeo para a
produção do ATP.
Duas moléculas de Aco-A formam ácidos que são transportados para
outras células e transformados em Aco-A de novo para se ligar ao ácido
oxalacético e entrar no ciclo de Krebs para produzir ATP.
Regulação da insulina: a glicemia e hormônios grastro-intestinais. Regulação
metabólica. A liberação dos hormônios gastro-intestinais estimulam a produção
de glicose.
Hipoglicemia: liberação de glucagon. O glucagon se opõe ao efeito da insulina,
ou seja, libera glicose.
GLÂNDULA ADRENAL
A glândula adrenal está localizada acima dos rins, possui região cortical e
medular (gânglio simpático: recebe diretamente do SNC uma inervação
simpática). Essas duas regiões secretam hormônios diferentes.
A região cortical possui 3 regiões diferenciadas pela atividade das
enzimas:
-glomerulosa: mineralocorticóides (aldosterona)
-reticulada: glicocorticóides (cortisol)
-fasciculada: hormônios sexuais, produção irrelevante. É importante em
doenças.
Aldosterona: regulação da concentração plasmática e PA.
Os hormônios corticais derivam do colesterol, esse se transforma em
pregnolona que pode se transformar em 17-OH pregnolona (glicocorticóides),
17-OH progesterona (mineralocorticóides) ou progesterona.
O que vai diferenciar uma região ou outra da zona cortical é a atividade
de enzimas e essas enzimas vão determinar a especificidade das células.
O receptor para aldosterona está no citosol. O complexo hormônio
receptor (H-R) migra para o núcleo, estimulando a síntese protéica. Esse
complexo H-R ativa a síntese de uma proteína que são canais para Na +. O Na+
entra da luz para a célula, então, a concentração de Na+ intracelular é muito alta
e se abrem as bombas de Na+/K+ para que o excesso de Na+ saia para o
interstício.
Portanto, o ADH regula a concentração de água e a aldosterona regula a
concentração de sal (Na+).
Regulação da aldosterona: é feita pela concentração de Na +, portanto não é
hipofisária nem hipotalâmica. A regulação também é feita pela presença da
angiotensina II (ver produção de aldosterona na parte do sistema respiratório).
A concentração de K+ é reguladora, mas não é importante, assim como a
presença de ACTH. O ACTH só regula a produção de aldosterona se estiver em
muita quantidade.
Corticóide: glicocorticóide é liberado em situações de stress. É um hormônio
catabólico, utiliza as reservas, quebra o glicogênio e a glicose que vão para o
plasma. Nesse caso, o organismo não tem glicose disponível na célula para
produzir ATP. Essa glicose é liberada para que o corpo possa usa-la onde
necessitar. Como não tem glicose para formar ATP, o corpo usa reservas de
gordura. O corpo vai usar carboidratos e aa para produzir glicose.
O corticóide tem o mesmo efeito cetogênico que a insulina. Ele é
vasodilatador, então passa mais sangue nos vasos e o plasma extravaza mais
para o interstício, levando consigo células de defesa num processo inflamatório.
Os corticóides permitem a expressão de receptores adrenérgicos.
Regulação da liberação de corticóide: é importante manter uma quantidade
normal de corticóides para regular os receptores para adrenalina. E se não há
adrenalina em uma situação de emergência, o corpo entra em choque. Quem
toma muito corticóide por muito tempo pode ter uma crise psicótica.
REGULAÇÃO DO Ca2+ EXTRACELULAR
É muito importante manter o cálcio extracelular, para isso é necessário ter
uma boa dieta onde exista cálcio. Além da dieta de cálcio, outra fonte de cálcio
no corpo são os ossos. Se não há uma boa quantidade de cálcio na dieta, o corpo
o retira dos ossos.
Absorção intestinal: absorve de acordo com a necessidade de cálcio.
Pouco cálcio, absorve mais, muito cálcio, absorve menos.
Absorção renal: ocorre igualmente à intestinal, dependendo da
necessidade de cálcio no corpo.
Absorção óssea: o osso é tampão do cálcio. Se não existe uma boa dieta
de cálcio, o corpo retira dos ossos.
Regulação do cálcio: é feita pelo PTH, vitamina D e pela calcitonina.
-Vitamina D: o precursor da vitamina D chamado dehydrocolesterol (derivado
do colesterol), se acumula na pele. Quando batem os raios solares, esse
dehydrocolesterol se transforma em colecalciferol (D3) que é uma vitamina D
inativa e esse passa no fígado e se transforma em 25 OH D 3 que ao passar pelos
rins se transforma em 1,25 (OH)2 D3 que é a vitamina D ativa. A vitamina D
ativa receptores nas microvilosidades do intestino, estimulando a síntese
(absorção) de cálcio no corpo.
Como a vitamina D é lipossolúvel, ela entra no núcleo das células e ativa
a transcrição de proteínas que são receptores para o cálcio.
-PTH: paratormônio que atua na transformação de colecalciferol em 25 OH D 3.
Sem PTH, o corpo produz pouca vitamina D, por isso é importante ter PTH a
vida toda. O PTH tem efeito indireto na absorção de cálcio por causa do seu
efeito. Com muito PTH, tem pouco cálcio no corpo e é necessário que o corpo
não perca cálcio, o PTH então, atua no rim, ativando a absorção de cálcio no
corpo.
O PTH atua também na atividade osteoclástica, aumentando a atividade
dos osteoclastos de digerir o osso.
Aumento de PTH: hiperglicemia
Diminuição de PTH hipoglicemia
-Calcitonina: faz mais ou menos o contrário que o PTH. A calcitonina inibe a
atividade dos osteoclastos. Os osteoclastos digerem o osso para liberar cálcio. A
calcitonina atua quando há pouco PTH.
HORMÔNIOS SEXUAIS MASCULINOS
A testosterona é o principal representante dos hormônios masculinos. Os
hormônios sexuais vão promover o amadurecimento de estruturas sexuais.
-Espermatogênese: resume-se em divisão celular, gerando espermátides que se
transformam em espermatozóides. A espermatogênese é regulada por
hormônios. Células responsáveis: células de Leydig – produção de testosterona;
e células de Sertoli – nutrição e absorção das células germinativas. As células
de Sertoli também produzem estrógeno, mas é uma produção mínima e ela não
cai na corrente sangüínea.
-Ato sexual: a testosterona têm função na atração (desejo) sexual. Simpático:
ereção e parassimpático: ejaculação.
Os hormônios sexuais atuam diretamente no SNC.
-Características sexuais:
Primária – formação de próstata, vesícula seminal, testículos e pênis.
Enfim, está relacionada com a formação da genitália masculina.
Secundária – formação de pêlos, deposição de gordura no abdome,
mudança da voz que são determinados pela presença dos hormônios
masculinos.
A calvície é determinada pelos baixos níveis de testosterona.
Os hormônios sexuais são anabólicos, ou seja, aumentam a síntese
protéica.
Regulação de hormônios sexuais
GnRH (hipotálamo)
+
FSH
Estrógeno
(Sertoli)
LH
Testosterona
(Leydig)
Gonadotrofina coriônica: produzida pela placenta, atua sobre o feto para
a produção de testosterona se o feto for XY.