Prá galera da Anatomia Vegetal:
Como combinado, estou enviando algumas dicas (roteiro de estudos) para a prova da
nossa primeira unidade.
Primeira aula: na primeira aula, vimos em linhas gerais como as plantas evoluíram de
organismos simples e se diversificaram, tomando como ênfase a invasão do ambiente
terrestre. Vimos que para que isto acontecesse foi necessário que as plantas
desenvolvessem sistemas de tecidos: dérmico, vascular e fundamental; cada qual
constituído por um determinado número de tecidos, conforme o estágio de
desenvolvimento (primário ou secundário), quais sejam: epiderme ou periderme, xilema
e floema primários ou secundários, parênquima, colênquima e esclerênquima,
respectivamente. Vimos, também, como uma planta produz os diferentes tecidos a partir
de uma célula única (o zigoto) envolvendo os fenômenos de divisão, crescimento e
diferenciação celulares e, ainda, de como esse processo se assemelha ao de evolução das
plantas (ontogenia x filogenia). Falamos a respeito da totipotência da célula vegetal e de
como esses fenômenos se relacionam com a biotecnologia vegetal. Desta primeira aula
o importante é dominar os conceitos de diferenciação celular e totipotência. Estes
conceitos estão muito bem explicados nos capítulos 2 e 3 (se não me engano!) do
primeiro volume do livro da Elizabeth Cutter, que pode ser encontrado nas bibliotecas
da Agronomia e do Instituto de Biologia. Perguntas que podem ser feitas: O que é
totipotência? Por que a diferenciação celular é considerada um fenômeno espacial? O
que determina o destino de uma célula em particular dentro de um organismo? Que
fatores influenciam ou determinam a diferenciação celular? No processo de produção de
mudas por estaquia (mudas de galho), por que células que inicialmente somente
expressavam características caulinares passam a produzir raízes? O que garante o
desenvolvimento posterior da muda?
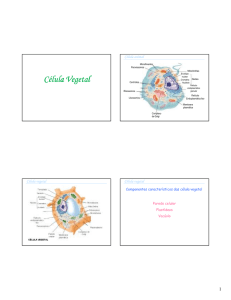
Segunda Aula – Parede celular e vacúolo: na segunda aula vimos que há três
características distintivas principais entre as células vegetais e animais: a parede celular,
o vacúolo e os plastídios. Começando pela parede celular, vimos que ela se diferencia
em primária e secundária e que há semelhanças e diferenças estruturais e químicas entre
elas. Vimos quais são seus componentes químicos, de que forma se apresentam e como
estão arranjados entre si (lembram do exemplo do algodão e da gelatina?). Definimos
apoplasto e simplasto e diferenciamos campos primários de pontoação e pontoações.
Perguntas que podem ser feitas: Quais são e como se arranjam os componentes básicos
da parede celular primária? Quais as diferenças entre as paredes celulares primária e
secundária? O que são apoplasto e simplasto? Qual a diferença entre um campo
primário de pontoação e uma pontoação? Quais as funções da parede celular? O que é
um plasmodesma e qual a sua relação com os campos primários de pontoação e com as
pontoações? Como a presença de água na parede pode definir a consistência (dureza) de
um tecido ou órgão?
Com relação ao vacúolo, vimos que é a organela mais expressiva da célula vegetal,
envolvida por uma membrana, o tonoplasto, que contém uma solução de variadas
substâncias químicas, o suco celular. O vacúolo tem a capacidade de absorver e perder
água muito rapidamente. Por todas essas razões, diferentemente do que se pensava até
muito pouco tempo, o vacúolo possui várias e importantes funções na célula vegetal.
Perguntas que podem ser feitas: Quais a funções do vacúolo? O que é pressão de
turgescência e como ela interfere na sustentação, crescimento e movimentos celulares
(lembram-se da câmara-de-ar no interior do pneu?)? Por que o vacúolo é considerado
como o principal componente do sistema lisossômico da célula vegetal? De que forma o
vacúolo protege o citoplasma contra substâncias tóxicas ao metabolismo?
Terceira aula – Plastídios e Substâncias Ergásticas: Vimos que os plastídios ou
plastos são, também, organelas muito expressivas das células vegetais, apresentam
formas variadas, são envolvidos por um sistema duplo de membranas que possibilitam
diferentes ambientes químicos no interior das células. Possuem DNA, RNA e
ribossomas próprios sendo, portanto, capazes de síntese protéica e autoduplicação. São
classificados segundo a presença ou não de pigmentos (cromo e leucoplastídios) ou,
ainda, segundo as substâncias que sintetizam e acumulam (amiloplastídios,
proteinoplastídios, etc.). Todas as formas de plastídios são interconversíveis e derivam
de proplastídios (plastídios fundamentais). Dentre todos os plastídios o mais importante
é o cloroplastídio que contém a clorofila e é responsável pela fotossíntese, apresentando,
também, a estrutura mais complexa. O que pode ser perguntado: Como são classificados
os plastídios? Como estão estruturados os cloroplastídios? Qual a importância do
sistema de endomembranas dos cloroplastídios? Do ponto de vista dos plastídios, qual o
fenômeno que está envolvido na maturação dos frutos? Em que plastídios são formados
o amido de assimilação e os grãos de amido? Qual a origem dos tilacóides? Qual a
importância dos cromoplastídios nas flores e frutos?
Os grãos de amido constituem a principal reserva nutritiva das plantas sendo
constituídos de amilose e amilopectina. Da mesma forma que a celulose, são formados
por longas cadeias de glicose que formam moléculas lineares e ramificadas que
apresentam maior ou menor afinidade à água, respectivamente. Embora possam estar
presentes diretamente no citoplasma, geralmente são formados nos amiloplastídios. Ao
microscópio óptico são estruturas bastante simples e, apesar disso, são considerados
como uma característica altamente diagnóstica, pois são exclusivos a cada espécie de
planta. O que pode ser perguntado: Relacione cinco atributos dos grãos de amido que os
tornam úteis ao reconhecimento de plantas ou de fragmentos dessas.
Os cristais são inclusões sólidas comuns nas células e são geralmente encontrados no
interior de vacúolos, apresentando diferentes formas. São formados por sais de cálcio ou
silício e apresentam formas variadas, constituindo estruturas de valor diagnóstico.
Dentre os cristais constituídos de oxalato de cálcio temos: drusas, ráfides, monocristais,
estilóides, e areias cristalinas. Formados por carbonato de cálcio, temos os cistólitos,
muito característicos de umas poucas famílias como Moraceae, Urticaceae e Ulmaceae.
Já os corpos silicosos são formados por óxido de silício e são característicos das
famílias Poaceae (gramíneas) e Cyperaceae. O que pode ser perguntado: Descreva os
seguintes cristais: drusas, ráfides, etc. Por que os cristais são considerados
características diagnóstico? Quais as funções dos cristais?
Quarta aula – Meristemas: Diferentemente dos animais as plantas possuem um tipo
de crescimento denominado aberto, isto é, durante toda a sua vida matêm o potencial de
crescimento. Em uma planta adulta convivem tecidos eternamente embrionários (os
meristemas apicais de caule e raiz e, também, as gemas axilares formadas diretamente
pelos primeiros), tecidos primários formados por estes, tecidos secundários formados a
partir de tecidos maduros que se desdiferenciam e retornam à condição meristemática
para dar origem a novas células que são acrescentadas secundariamente ao corpo do
vegetal. Um meristema, portanto, é uma região localizada do corpo vegetal onde
ocorrem divisão, crescimento e diferenciação celulares. São de dois tipos básicos:
apicais (ou primários) e laterais (secundários), sendo responsáveis pelo crescimento em
extensão ou em diâmetro, respectivamente. Independentemente da categoria a que
pertençam, todos os meristemas são formados por células iniciais e derivadas. As
primeiras são aquelas que após a divisão permanecem no meristema, enquanto as
derivadas, que sofrem o maior número de divisões celulares, são acrescentadas ao corpo
da planta dando origem aos tecidos. O que pode ser perguntado: Quais são os
meristemas primários e qual a origem dos sistemas dérmico, fundamental e vascular
nestes? Por que os meristemas laterais são também chamados de secundários? Quais são
os meristemas laterais e quais tecidos originam? Qual a função do procâmbio? Por que
as células da camada mais externa dos meristemas apicais sofrem somente divisões
anticlinais (perpendiculares à superfície)? Por que as células iniciais sofrem menos
divisões do que as derivadas? Qual a importância dos ápices meristemáticos para
procedimentos de poda ou estaquia (muda de galho)?
Quinta aula – Epiderme: A epiderme é um tecido complexo, isto é, constituído por
diferentes tipos celulares que podem ser classificados basicamente em: células
epidérmicas comuns (revestem o corpo do vegetal em estágio primário de crescimento),
células-guarda de estômato, tricomas (ou pêlos) e pêlos radiculares. As células
epidérmicas comuns têm origem na camada mais externa dos meristemas apicais do
caule e raiz (a protoderme) e geralmente apresentam um formato tabular ou
pavimentoso (mais largas do que altas). Quando vistas de frente, podem apresentar
contornos retos ou, sinuosos, à semelhança de peças de quebra-cabeça. Contornos
sinuosos estão geralmente presentes em folhas membranáceas, pois a maior superfície
de contato entre as células garante maior adesão e resistência contra ações mecânicas
(dobramento pelo vento, etc.). De maneira geral as células epidérmicas tendem a
acompanhar a forma das células dos tecidos que lhe são subjacentes, assim as células
sobre as nervuras de uma folha tendem a ter contornos retos e serem alongadas no
mesmo sentido destas, enquanto que, nos espaços entre as nervuras, podem ser sinuosas.
Quando vistas em corte, as células epidérmicas apresentam-se achatadas e suas paredes
são irregularmente espessas: as periclinais externas (paralelas à superfície) mais
espessas do que as anticlinais (perpendiculares à superfície) e periclinais internas. Como
um tecido de revestimento, não há espaços intercelulares e as células estão intimamente
unidas. Também o protoplasto das células epidérmicas é distribuído desigualmente. Um
grande vacúolo posiciona-se junto à parede periclinal externa e todo o citoplasma e
organelas, principalmente o núcleo, são deslocados para a parede periclinal interna;
desta forma a espessa parede periclinal externa e também o vacúolo, que muitas vezes
contém pigmentos como a antocianina, protegem o protoplasto da ação dos raios
ultravioleta. Geralmente os cloroplastídios estão ausentes (a clorofila se degrada com o
excesso de luz), mas estão presentes leucoplastídios e, algumas vezes cromoplastídios
contendo pigmentos carotenóides, que são, também, agentes protetores contra a ação
dos raios “ultraviolentos”. Os vacúolos das células epidérmica costuma apresentar um
potencial hídrico fortemente negativo (a solução vauolar é altamente concentrada de
sais, açucares, etc.) o que garante uma grande pressão de turgecência, contribuindo
assim para a manutenção da arquitetura dos órgãos, especialmente folhas de pequenas
ervas.
A característica mais marcante das células epidérmicas comuns (assim como das
células-guarda de estômato e dos tricomas) é a presença da cutícula, que é caracterizada
quimicamente pela presença da cutina (um poliéster de ácidos graxos), seja
impregnando as paredes celulares (camada cutinizada), ou formando, juntamente com as
ceras intracuticulares, uma fina camada externa constituída unicamente de substâncias
lipídicas, a cutícula propriamente dita ou camada cuticularizada. A presença desses
lipídios (substâncias apolares e, portanto, hidrófobas) formando uma camada contínua
sobre as células epidérmicas fazem deste tecido uma camada de resistência contra a
perda de água por transpiração. Assim, quando os estômatos estão fechados, somente a
pouca água que é capaz de atravessar essa camada “impermeabilizante” é perdida para a
atmosfera (transpiração cuticular), cerca de apenas 10% do total da água perdida pela
planta. Por outro lado, sabe-se que não somente a água, mas muitas outras substâncias,
como os herbicidas hidrofílicos a exemplo do glifosato, são capazes de penetrar as
superfícies aéreas das plantas, de forma que essa “impermeabilidade” deve ser
entendida com reservas. Mesmo sendo uma substância lipídica a cutina têm alguma
afinidade à água que depende, entre outros fatores dos seus graus de polimerização e de
metil-esterificação. Assim, a cutina, ao contrário das ceras, é capaz de absorver e reter
alguma quantidade de água expandindo-se. Os polímeros de cutina formam uma rede
tridimensional semelhante a uma bucha (ou esponja) vegetal em que as cavidades estão
preenchidas de ceras hidrófobas. Na presença de umidade atmosférica, o movimento de
expansão da matriz polimérica de cutina não é acompanhado pelas ceras, gerando-se
pequenas cavidades na interface entre as duas substâncias, por onde a água e solutos
podem fluir para o interior da planta. Sob alta demanda evaporativa da atmosfera a
matriz de cutina perde água e se contrai, aderindo-se fortemente às ceras, impedindo o
fluxo de água para o exterior. Desta forma, mais do que uma camada impermeabilizante
a cutícula atua como uma válvula entre os meios interno e externo das plantas.
A cutina é, também altamente resistente ao ataque de agentes físicos e químicos,
fornecendo proteção contra a ação do vento, de agentes químicos (chuva ácida, por
exemplo) e ao ataque de patógenos como fungos e bactérias e vírus.
Os estômatos são poros epidérmicos definidos por duas células epidérmicas especiais,
chamadas células-guarda do estômato. Vistas frontalmente essas células-guarda
geralmente apresentam o formato de rins opostos um ao outro determinando a presença
de um poro entre elas (o ostíolo). Este poro é capaz de abertura e fechamento devido à
ação dos vacúolos sobre as paredes desigualmente espessadas das células-guarda.
Assim, quando a planta perde água, os poros se fecham e a perda de água por
evapotranspiração é evitada, restringindo-se à transpiração cuticular que corresponde a
cerca de 10% do total de água perdida pela planta. Quando as células retomam a sua
pressão de turgescência os poros se abrem, possibilitando, a absorção do CO2,
necessário a fotossíntese, liberando, também o oxigênio.
Tricomas são células epidérmicas que se projetam para além da superfície do vegetal.
Podem ser constituídos de uma única célula, variando em forma desde uma pequena
papila até estruturas complexas constituídas de várias células. São classificados em
tectores (ou de cobertura) ou glandulares (secretam uma gama variada de substâncias).
Dependendo do tipo ou da densidade, os tricomas podem ter variadas funções na planta.
Uma densa cobertura de pêlos mortos na superfície de uma planta mantém predadores
fisicamente afastados dos tecidos vivos, evita a circulação de ar junto à superfície, retém
vapor d’água junto a superfície e proporciona uma camada reflexiva da radiação solar
excessiva. Desta forma estes tricomas contribuem para a proteção da planta contra a
herbivoria (variedades pilosas de soja ou algodoeiro são menos suscetíveis ao ataque da
cigarrinha!) assim como reduzem a perda de água por evapotranspiração. Em alguns
casos, os tricomas podem produzir ganchos ou “espinhos” que danificam o sistema
tegumentar de larvas predadoras ou dificultam sua locomoção, causando-lhes a morte
por trauma, dessecação ou fome. Um caso notável de proteção contra a herbivoria são
os tricomas urticantes da urtiga que, tal como agulhas hipodérmicas, injetam substâncias
letais a muitos predadores. Algumas substâncias produzidas em tricomas glandulares
podem atuar como repelentes químicos contra os predadores ou, algumas vezes, como
atração de agentes polinizadores (como é o caso de tricomas secretores de néctar).
Alguns tricomas podem atuar, também, como estruturas de absorção de água e
nutrientes aéreos, substituindo total ou parcialmente o papel das raízes (é o caso da
barba-de-pau ou do cravo-do-mato que crescem inclusive sobre fios elétricos e
alambrados). Os pêlos radiculares nada mais são do que células da rizoderme que se
projetam em direção ao solo em busca de fontes de água, formando longos fios ou
túbulos que ampliam grandemente a superfície de contato com o solo e,
consequentemente, a capacidade de absorção de água e nutrientes. Estritamente, são
também tricomas, mas que são classificados separadamente em razão de sua natureza e
funções precípuas. Micorrizas são associações simbióticas entre fungos e as raízes. Os
fungos sabidamente são os maiores organismos terrestres, são decompositores de
matéria orgânica e produtores de substâncias antibióticas como a penicilina. O corpo
vegetativo de muitos fungos são formados por finos filamentos celulares que se
espalham em forma de círculo no solo (ou outros substratos como uma tora de madeira)
atingindo diâmetros de até 3 km. Assim, as micorrizas substituem com vantagens o
papel dos pêlos radiculares, ampliando em muito a superfície de contato com o solo e
aumentando a capacidade da planta em adquirir água e, também, nutrientes minerais, já
que decompõem e mineralizam a matéria orgânica solo, tornando, desta forma, os
nutrientes disponíveis às raízes. É sabido, também, que as associações micorrízicas,
protegem as plantas de patógenos, não somente por produzirem plantas bem nutridas e
saudáveis, mas, talvez, pela produção de antibióticos que seriam compartilhados com a
planta. O que pode ser perguntado: Quais as funções da epiderme? Qual a principal
característica química e estrutural da epiderme? De que forma a cutícula pode atuar
como uma válvula entre os meios interno e externo das plantas no que se refere ao
transporte de água e outras substâncias? Por que os herbicidas são aplicados no início da
manhã ou à tardinha? O que são estômatos e quais são as suas funções? De que forma a
abertura e o fechamento dos estômato é regulada? Quais as principais características
citológicas das células epidérmicas comuns? O que são e para que servem os tricomas?
De que forma as micorrizas substituem com vantagens os pêlos radiculares?
Este é apenas um roteiro de estudos, não significando que serão estas as questões
da prova ou que sejam tal e qual estão colocadas aqui. A propósito, será dez o
número de questões e elas serão variadas, desde escolha simples, correlação de
colunas, falso e verdadeiro até questões descritivas, todas elas valendo um ponto.
Bibliografia
APPEZZATO-DA-GLÓRIA, B. & CARMELLO-GUERREIRO, S. (Ed.) Anatomia
vegetal. Viçosa: Editora da Universidade Federal de Viçosa. 2003.
CUTTER, E. G. Anatomia vegetal: parte I – células e tecidos. São Paulo: Rocca, 1986.
_____ Anatomia vegetal: parte II – órgãos, experimentos e interpretação. São Paulo:
Rocca, 1987.
ESAU, K. Anatomia das plantas com sementes. São Paulo: Edgar Blücher, 1974.
OLIVEIRA, E. C. de Introdução à biologia vegetal. São Paulo: Editora da
Universidade de São Paulo, 1996. (Acadêmica; 7)
RAVEN, P. H.; EVERT, R. F. & EICHHORN, S. E. Biologia vegetal. Rio de Janeiro:
Guanabara Koogan S.A., 2001.
Obs.: os livros grifados em vermelho são facilmente encontrados nas bibliotecas da
Agronomia e do Instituto de Biologia. Qualquer um deles é indicado para estudo.***