FILO NEMATODA
BIOLOGIA E ECOLOGIA
...Neste contexto, os nematódeos adquirem grande importância por serem considerados o
grupo de metazoários mais abundante na biosfera, com estimativa de constituírem até 80%
de todos os metazoários (Bongers, 1988), que incluem muitas formas parasitas de plantas e
animais, inclusive do homem.
O filo NEMATODA abrange os nematóides, vermes não segmentados, de corpo
cilíndrico, alongado, cutícula resistente e estruturas bucais com arranjo trirradial. Com
cerca de 27.000 espécies descritas (e um potencial estimado em 500.000-1.500.000)
constituem uns dos grupos de metazoários mais abundantes. Existem umas 10.700 espécies
de vida livre (solo, água doce e mar) e umas 16.300 parasitas de invertebrados, vertebrados
e plantas. Alguns lodos do fundo do mar têm mais de quatro milhões de nematóides por
metro quadrado, enquanto que um bom solo pode ter 3-9 bilhões por acre (uns 4000 m2).
Sua dramática abundancia levou a Cobb a escrever em 1914: “Se toda a matéria do
universo exceto os nematóides fosse varrida, nosso mundo ainda seria vagamente
reconhecível, e se, como espíritos sem corpo, pudéssemos então pesquisa-lo, deveríamos
encontrar montanhas, vales, rios, lagos e oceanos representados por um fino filme de
nematóides”.
São encontrados em qualquer lugar; talvez nenhum outro grupo taxonômico seja tão
universal quanto ao habitat. Os nematódeos parasitas encontram-se em quase todas as
espécies animais documentadas o que dá uma enorme variedade de habitats (Hodda, 2004).
Muitos são generalistas, mas outros são muito especializados a habitats específicos como
raízes de plantas, sementes de trigo, secreção de feridas de árvores e intestino, sangue e
outros órgãos do corpo dos animais, até chegar ao extremo de uma espécie de nematóide
que só foi encontrada em algumas cidades alemãs debaixo dos descansos para copos de
feltro de canecas de cervejas. A maioria dos nematóides possui menos de cinco cm de
comprimento e muitos são microscópicos (0,08 mm), mas alguns nematóides possuem
tamanhos gigantescos como a espécie parasita da placenta dos cachalotes Placentonema
gigantissima que alcança os nove metros de comprimento.
São conhecidas umas 4000 espécies de nematóides marinhos podendo ser de vida livre ou
endoparasitas. No mar são bentônicos, morando nos espaços intersticiais, algas e
sedimentos principalmente lama e areia. Todo tipo de sedimento pode ser colonizado pelos
nematódeos, desde as secas dunas de areia das praias até os pisos hadais oceânicos. Faltam
na zona pelágica, mas muitas vezes entram em suspensão na coluna de água formando parte
do seston, e também existem espécies associadas às algas flutuantes do gênero Sargassum.
Os nematódeos são o componente principal da meiofauna, representando um 50-90% com
uma densidade até de um milhão de indivíduos por metro quadrado e um tamanho entre
0,5-3 mm. Comumente os nematódeos apresentam uma riqueza de 40-50 espécies por cm2
em ambientes marinhos e 5-15 espécies por cm2 em águas salobras. Embora a biomassa dos
nematódeos marinhos de vida livre pareça baixa (1gr/m2), sua significância em termos de
fluxo de energia, teias alimentares e degradação e mineralização da matéria orgânica é
muito elevada. Em especial o papel dos nematódeos no ciclo do nitrogênio é considerável,
contribuindo com a mineralização de 4-22% do nitrogênio (Griffiths, 2001). A atividade
metabólica da comunidade de nematódeos pode ultrapassar a atividade da macrofauna
associada ao mesmo habitat marinho.
A maioria dos nematódeos marinhos adultos são vermes cilíndricos alongados de 12 mm de comprimento, se bem algumas formas encontradas nas raízes de macroalgas (kelp
holdfast) podem atingir vários milímetros de comprimento.
Os nematódeos são estruturalmente organismos simples, caracteristicamente
pseudocelomados, cobertos por uma cutícula protéica. Os adultos contem aproximadamente
umas 1.000 células somáticas, e potencialmente centenas de células associadas ao sistema
reprodutivo. O mesénquima se reduz quase completamente, de modo que o espaço entre o
intestino e a camada muscular subepitelial está ocupado pela cavidade primaria no limitada
por una membrana (blastoceloma). O desenvolvimento é direto, só nos parasitas aparecem
larvas secundárias. Mostram uma forte tendência à constância no número de células nos
diversos tecidos (eutelia), a conseqüência do qual a capacidade de regeneração fica limitada
a partes de células ou zonas histológicas anucleadas, faltando completamente a reprodução
assexuada. Sem sistema de vasos sanguíneos nem celoma.
A pesar de suas formas de vida muito diversas, conservam uma assombrosa
uniformidade estrutural. São vermes alongados, filiformes de corpo delgado e secção
circular, cilíndricos, as vezes filiformes ou fusiformes. Quase sempre seus extremos se
aguçam gradualmente. Não segmentados (as vezes superficialmente segmentados), pelo
geral transparentes com superfície brilhante. De um modo geral são organismos com
simetria bilateral, seus órgãos se enrolam, as vezes se perde um dos membros e muitos de
vida sedentária tendem à simetria radial. Com ausência total de epitélios ciliados (os cílios
ficam limitados as células sensoriais) e de muitas diversas formas de vida. Muitas vezes
carecem totalmente de apêndices, ainda que algumas espécies de vida livre possuem
prolongações de fixação ou mais freqüentemente cerdas, que aparecem em forma mais ou
menos constante no extremo anterior do corpo. Não existe cabeça diferenciada e existe um
baixo grado de cefalização, o cerebro é anterior e os órgãos dos sentidos se concentram
especialmente em torno da boca. O ânus é ventral e é seguido por uma cauda, as vezes mais
estreita ou encurvada. A cauda contém as glândulas caudais unicelulares, usualmente três,
as quais podem estar totalmente confinadas a cauda ou se estender anteriormente ao
anus/cloaca. As glândulas caudais são responsáveis pela secreção de uma substancia
adesiva que é eliminada por uma estrutura chamada espinereta que desemboca num poro no
extremo posterior do corpo e as vezes assoma ao exterior como um rabo em forma de tubo.
O corpo segue o modelo arquitetônico de um tubo dentro de um tubo. O tubo
externo é a parede corporal e o tubo interno conformado pelo tubo digestivo. Entre o tubo
digestivo e a parede corporal se encontra um amplo blastoceloma preenchido de fluido
onde os órgãos reprodutivos estão alocados.
A parede corporal e o tubo digestivo provem vários caracteres úteis na identificação
conjuntamente com o sistema nervoso, excretor e reprodutivo. As quatro características
utilizadas na identificação dos nematódeos marinhos são a cavidade bucal (boca), a forma
da cauda, os anfidios e o padrão da cutícula.
Parede corporal
A parede corporal é um saco músculo-cutâneo, que compreende o aparato excretor e
o sistema nervoso. O saco músculo-cutâneo limita o blastoceloma, que contém ao aparato
digestivo e o reprodutor. O corpo está coberto por uma cutícula resistente, delgada e
quitinosa que freqüentemente tem um relevo em forma de anéis e cobre faringe, digestivo
posterior e outras aberturas corporais. As vezes tem um conjunto variável de proeminências
cuticulares (asas), que podem se encontrar ao longo do corpo, na região cervical ou caudal
do macho (em alguns se encontra uma expansão formada por três lóbulos, com função
copuladora).
A cutícula é um produto elástico das células epidérmicas subjacentes. Entrega um
suporte resistente, protege de alguns compostos tóxicos e permite aumentar o diminuir o
volume corporal sem cambiar a pressão do líquido perivisceral. Na cutícula predominam
proteínas similares ao colágeno, e na camada externa contém uma proteína do tipo das
queratinas, coberta por uma película lipídica. A cutícula consta de três regiões: camada
basal (estriada, laminada ou com fibras helicoidales), camada mediana (que varia entre una
estrutura granular uniforme ate bastonetes, fibrilas ou canais, e as vezes falta) e camada
cuticular externa, cortical ou córtex (freqüentemente anelada e dividida em parte externa e
interna). A camada externa está limitada por uma epicutícula delgada, que pode apresentar
uma coberta de quinona. A cutícula da superfície corporal geralmente apresenta
ornamentações, pontos, verrugas, costelas, espinhos ou cerdas ou outras modificações, que
atuam como órgãos sensoriais ou participam na locomoção. Os diferentes padrões das
cutículas são caracteres diferenciais a nível especifico em nematódeos marinhos.
Nos nematódeos o crescimento é acompanhado normalmente de quatro mudas da
cutícula. A cutícula antiga se separa da epiderme subjacente começando pelo extremo
anterior e o animal segrega a cutícula nova. A cutícula antiga se desprende inteira ou em
fragmentos, nos Adenophorea todas as camadas são mudadas, nos Secernentea a
epicutícula é dissolvida. O animal adulto não realiza mudas, mas a cutícula continua
expandindo-se em quanto o animal cresce.
Sob a cutícula existe uma subcutícula ou epiderme ou hipoderme, geralmente celular,
ainda que em parasitas as vezes é sincicial. A epiderme secreta a cutícula, mantém reservas
de nutrientes, contém fibras de ancoragem que unem a musculatura à cutícula e em algumas
espécies endoparasitas é uma importante superfície de absorção de nutrientes.
A epiderme dos nematódeos se expande ate o interior, formando quatro crestas ou
proeminências longitudinais ao longo das linhas media dorsal, media ventral e medias
laterais (cordões longitudinais). Incluem aos nervos dorsal e ventral e aos canais excretores
laterais, e dividem a musculatura em quatro campos. Geralmente os núcleos epidérmicos
estão restritos estes cordões. As zonas de epiderme situadas entre as protuberâncias são
campos plasmáticos anucleados e delgados. Em muitas espécies parasitas e de vida livre
existem glândulas cutâneas caudais.
O sistema muscular está constituído por células mioepiteliais, as que formam una camada
muscular, que corresponde à musculatura somática geral, e músculos especializados. A
camada muscular está formada por células mioepiteliais, fusiformes, que formam fibras
longitudinais, situadas em quatro bandas ou quadrantes (duas massas musculares dorsais e
duas ventrais) entre os cordões longitudinais. Muitos nematódeos têm só entre dois e cinco
músculos em cada campo (meromiarios), outros tem muitas células musculares em cada
campo (polimiarios). As fibras podem ser compridas e planas, ou bem altas e estreitas,
segundo a espécie. As células musculares dos nematódeos são as únicas que tem extensões
nervosas, cada fibra muscular possui um braço delgado que vai até o cordão nervoso
longitudinal, dorsal o ventral, onde se produz a inervação. Os músculos especializados se
utilizam para o movimento de partes especiais do corpo, por exemplo, os músculos labiais
para a captura do alimento, músculos faríngeos para engolir, músculos retais para a
defecação e músculos copuladores, associados aos órgãos reprodutores, espículas ou vulva.
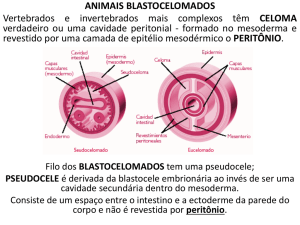
Blastoceloma
O blastoceloma ou pseudocele é una cavidade derivada do blastocele, localizada entre as
vísceras e a parede corporal, não está coberta por peritônio e possui líquido perivisceral em
seu interior. A cavidade se estende desde a musculatura até o tubo digestivo, e rodeia aos
órgãos reprodutores. Sua alta pressão hidrostática (forte turgescência), conjuntamente com
a cutícula, atua como antagonista elástico (hidroesqueleto) da camada longitudinal
muscular, o que se correlaciona funcionalmente com a ausência de músculos circulares. O
líquido perivisceral contém metabólitos orgânicos, incluindo hemoglobina em algumas
espécies. Em nematódeos parasitas acostuma ter substancias tóxicas, especialmente
hemolíticas para o hospede. Nas paredes do blastoceloma existem células fagocitárias fixas,
importantes na defesa interna.
Locomoção
Devido a falta de músculos circulares e a abundância de músculos longitudinais, se
locomovem curvando-se e retorcendo-se, para frente e atrás, mediante ondulações
serpenteantes no plano dorso-ventral, pela contração alternada de fibras musculares
longitudinais ventrais e dorsais, com ajuda do hidroesqueleto formado pelo fluido do
blastoceloma. O antagonista é a cutícula. Devido ao blastoceloma no segmentado, os
movimentos implicam sempre o corpo todo. Os anéis cuticulares podem contribuir na
flexibilidade, enquanto que fibras helicoidais cruzadas impedem a flexão e formação de
hérnias quando o corpo se dobra e aumenta a pressão hidrostática. Os movimentos
ondulatórios dos nematódeos de vida livre são eficazes para avançar só quando se realizam
sobre partículas do substrato ou contra a tensão superficial da película de água, o qual
permite nadar a muitas espécies. Muitos nematódeos nadam intermitentemente em
distancias curtas, por exemplos as espécies que habitam nos musgos, quando a planta está
alagada. Algumas espécies podem rastejar. As ornamentações da cutícula e as cerdas
anteriores ajudam a segurar-se ao substrato. A cutícula atua opondo-se a ação da
musculatura longitudinal. A glândula caudal ou espinereta, que se encontra na maioria das
espécies marinhas, se usa na fixação temporal, em movimentos de retração com fixação
pela cauda e no movimento de chicote.
Nutrição
Muitas espécies de vida livre são carnívoras e outras fitófagas. Formas marinhas e de
água doce se alimentam de diatomáceas, algas, fungos e bactérias. Abundam as espécies
terrestres que perfuram raízes vegetais para sugar seu conteúdo. Estes nematódeos
produzem grandes perdas comerciais. Também existem muitas espécies que ingerem
partículas de substrato (sedimentívoras), que ao igual que as que vivem na matéria orgânica
morta (estrume, cadáveres) se nutrem na realidade de bactérias e fungos. Algumas espécies
são saprófagas, se alimentam sugando cadáveres de pequenos animais ou plantas mortas, ou
seus restos em diversos estados de decomposição. Os nematódeos são o grupo de
consumidores de bactérias e fungos más abundante e cosmopolita, pelo que têm grande
importância nas teias tróficas. Segundo Moens et al (1999) os nematódeos de vida livre
marinhos apresentam quatro estratégias alimentares: comedores de depósitos seletivos,
comedores de depósitos não seletivos, comedores de epistratos, predadores verdadeiros e
predadores facultativos.
O aparato digestivo es quase retilíneo, raramente ondulado. Estende-se entre a abertura
oral (antero-terminal) e a abertura anal (subterminal), que pode faltar. Está formado por um
estomodeo (boca, cavidade bucal e faringe), um mesenterion (um intestino médio) e um
proctodeo (intestino terminal, que pode ser reto ou cloaca).
Na região bucal se manifestam as maiores variações. A boca carece de proboscíde, mas
comumente está muito diferenciada e recoberta pela cutícula. A superfície cuticular pode
espessar-se e estar reforçada por bordas, bastonetes ou placas, ou levar dentes afiados,
pontiagudo (onchia). A boca está rodeada por um número variável de lóbulos salientes ou
de lábios e cerdas e papilas de vários tipos. Nas espécies marinhas geralmente a boca está
rodeada por 6 lóbulos em forma de lábio, 3 a cada lado. Devido a fusão, as formas terrestres
e parasitas acostumam ter só 3 lábios. Primitivamente os lábios e a superfície anterior
externa possuem 18 cerdas. A boca conduz a uma cavidade bucal ou estoma, más o menos
tubular e recoberta por cutícula. Os detalhes estruturais da cavidade bucal estão
relacionados com os hábitos alimentares e são importantes na identificação das espécies. A
cavidade bucal pode ser um tubo estreito ou um espaço oval ou com forma de xícara.
Quando a cavidade bucal está muito especializada, pode dividir-se em una câmara anterior,
fechada pelos lábios; um prostoma, comprido, e um telostoma.
Em alguns carnívoros e vegetarianos, na cavidade bucal existe um comprido estilete oral
o arpão bucal, oco ou maciço, que pode sair da boca mediante ação muscular. Os estiletes
servem para punçar a presa, e o estilete oco atua, ademais, como um tubo por onde a
faringe suga. O estilete as vezes se origina em uma modificação do epitélio da cavidade
bucal (estomatostilo) e outras vezes por una importante modificação de um dente
(odontostilo). Os nematódeos que vivem no interior de tecidos animais e os saprófagos de
vida livre se alimentam predominantemente de líquidos e sua região bucal se reduz a um
poro diminuto que conduz à faringe. Nos nematódeos carnívoros existem freqüentemente
dentes, protuberâncias grandes, placas cortantes, raspas ou dentículos pequenos e
abundantes. Detrás da boca pode haver uma cápsula bucal com dentes na sua base.
A cavidade bucal se abre para uma faringe tubular denominada esôfago ou faringeesôfago. O lúmem da farínge é trirradiado em secção transversal, contém fibras musculares
radiais e também está revestida por cutícula. A parede tem células mioepiteliales e
glandulares. Em alguns casos é um tubo não especializado, mas geralmente está
especializada por regiões e varia amplamente em quanto a sua forma e à proporção relativa
de tecidos glandulares e musculares. Muitos nematódeos vegetarianos possuem um
engrossamento oval em torno ao centro da faringe. Entre os zôoparasitas a faringe pode
apresentar diferentes características. Alguns apresentam um ventrículo glandular não
muscular e posterior, que pode esticar-se, ou possuem a faringe dividida em una porção
anterior curta e una porção muscular posterior más comprida. As glândulas faríngeas
secretam enzimas que iniciam a digestão do alimento ou ajudam na penetração dos
nutrientes a través da parede celular. As secreções enzimáticas se eliminam fora da boca
realizando uma pré-digestão dos alimentos, previa a sua ingestão. Alguns nematódeos
possuem uma ou mais protuberâncias musculares (bulbos ou cegos) no extremo posterior
da faringe, que funcionam como bombas que levam o alimento líquido para o intestino.
Freqüentemente existem válvulas. Enquanto a faringe se enche, uma válvula faringointestinal está fechada, e quando os músculos faríngeos empurram o alimento para o
intestino, se abre a válvula e a da boca se fecha.
A digestão começa no lúmem do intestino e se completa intracelularmente. A digestão
intracelular, quando existe, é pouco importante. O epitélio intestinal secreta enzimas
digestivas. O intestino armazena alimentos e participa na síntese de vitelo. As reservas de
glicogênio e gorduras das células intestinais se consomem durante o jejum e a muda. As
vezes a seu entorno se encontra una capa muscular. Uma válvula em cada extremo do
intestino impede que o alimento seja expulso pela pressão do líquido blastocelómico. Desde
a faringe sai um intestino médio tubular, formado por una camada de células epiteliais, e
depois segue um reto ou intestino terminal, curto e aplanado, que está recoberto por
cutícula e pode conter glândulas retais unicelulares.
O intestino médio é um tubo simples com alguma escassa especialização regional.
Apresenta poucas variações, ainda que o número de células que o compõem varia
enormemente. Em algumas espécies, apresenta em seu extremo anterior umas evaginações
cegas, e na superfície pode estar pregada. Em alguns casos se pode distinguir,
histológicamente, uma região anterior ventricular, uma região media e uma pré-retal.
O intestino terminal ou reto deriva da ectoderme e está coberto por cutícula. Estende-se
desde a válvula intestino-retal até o ânus, que está na linha media ventral, antes do extremo
do corpo, e tem forma de fenda. O lábio do ânus e a parede retal se levantam pela ação de
um músculo que ajuda na defecação. A forca de expulsão deriva da pressão do blastocele.
Em espécies parasitas são freqüentes as glândulas retais grandes e unicelulares. Nos
machos, o conduto reprodutor se une ao reto formando uma câmara chamada cloaca, ainda
que nela não desembocam as vias excretoras.
Respiração
Os nematódeos carecem de órgãos respiratórios diferenciados. Os adultos que vivem
como parasitas intestinais são principalmente anaeróbios, neles falta o ciclo de Krebs e o
sistema de citocromos, mas todos podem utilizar o oxigênio si está disponível. Alguns
nematódeos de vida livre e os estados livres de alguns parasitas são aeróbios obrigados,
possuindo o ciclo de Krebs e o sistema de citocromos.
Excreção
Os nematódeos excretam resíduos em forma de amônio, que difunde a través da parede
corporal. O sistema excretor carece de células flama e de protonefridios. Uns poucos
nematódeos carecem de qualquer sistema de excreção. A osmorregulação, a regulação
iónica e, talvez, a excreção de outros resíduos, se associam geralmente com estruturas
especializadas particulares: uma célula ou células glandulares excretoras (glândula ventral),
sistema de canais excretores (condutos aqüíferos), ou ambos. Estes sistemas aparecem só
em nematódeos e dentro do grupo constituem os órgãos más desenvolvidos. Em ambos
casos de trata de formações unicelulares. Quando se apresentam tanto sistema de canais
como glândulas, compartem o mesmo poro para o exterior.
A célula glandular excretora ou glândula renete é grande, se encontra ao nível da união
de faringe com o intestino médio, ventralmente no pseudocele e tem um conduto em forma
de colo que desemboca medioventralmente em um poro. Seu papel excretor é incerto,
foram sugeridas outras funciones, como secreção de matriz gelatinosa em torno aos ovos,
secreção de uma coberta de glicoproteínas sobre a cutícula e produção de exoenzimas para
iniciar a digestão dos tecidos do hospedeiro. O órgão excretor de tipo glândula ventral se
encontra especialmente em formas de vida livre, assim os nematódeos marinhos acostumam
ter uma simples célula excretora (as vezes possuem duas) saculiforme com seu colo aberto
para o poro excretor.
O sistema de canais excretores, sem cílios, se encontra dentro de uma célula, a célula más
grande do corpo do animal, geralmente tem forma de agá, os dois canais compridos se
situam nos cordões epidérmicos laterais e se unem por um ou vários canais situados na
barra transversal. Este sistema serve especialmente para a regulação iônica. Do conduto
transversal sai um conduto curto impar que desemboca na superfície ventral do corpo em
um poro da região faríngea. As vezes o conduto se alarga e forma una ampola que se enche
e esvazia ritmicamente. O sistema de canais que correm ao longo dos cordões laterais se
encontra em muitos parasitas e em espécies do solo. Em alguns casos este tipo de sistema
excretor apresenta um par de glândulas cervicais.
Sistema nervoso
O sistema nervoso é intraepitelial, localizado na epiderme, a faringe e no digestivo
posterior. Existe um cérebro anterior em forma de um anel nervoso circum-entérico, com
gânglios dorsais, ventrais e laterais associados, que rodeia ao intestino anterior e sobre o
qual as células nervosas se distribuem geralmente em forma difusa, e vários cordões
longitudinais. O anel nervoso circum-entérico está formado principalmente por fibras
nervosas, os corpos neurais se localizam em gânglios. Do anel nervoso circum-entérico
saem para a frente nervos que inervam os órgãos sensoriais, especialmente as cerdas e os
anfidios. Para a parte posterior saem troncos nervosos dorsais, laterais e ventrais dentro dos
cordões longitudinais, unidos por comissuras. Alguns são motores (mediodorsal,
medioventral, sublaterais), os outros são sensitivos (laterais). Os extremos das células
musculares contatam com os nervos dorsais e ventrais; os nervos laterais se associam com
os canais excretores. O cordão nervoso dorsal, predominantemente motor, se encontra no
cordão epidérmico dorsal. Os troncos nervosos laterais, predominantemente sensitivos, se
encontram nos cordões epidérmicos laterais, atravessando os gânglios lombares e
terminando no ânus. O nervo principal é o tronco ventral, que freqüentemente tem raízes
pares e provavelmente deriva da fusão dos nervos. Existe um sistema simpático faríngeo.
Os nervos viscerais inervam o intestino. Nas vias nervosas existem gânglios, concentrados
especialmente cerca do extremo posterior, onde existe um gânglio anal ou caudal, simples
ou duplo.
Órgãos dos sentidos
Os órgãos sensoriais são relativamente simples. Estão concentrados no extremo corporal
anterior. As formas livres possuem um maior número que as parasitas. No tipo fundamental
se situam em três círculos e em um par de fossetas laterais. Os círculos constam de 4 ou 6
órgãos sensoriais. Os receptores tácteis se apresentam sob a forma de papilas ou de cerdas,
os quimiorreceptores, como os anfidios, variam desde um simples poro ate uma complexa
estrutura tortuosa e espiralada. Os órgãos sensoriais levam um dendrito ciliado englobado
em uma parte especializada da parede corporal. Nas papilas, cerdas e anfidios existe um
dendrito com um cílio distal.
Papilas labiais e cefálicas: pequenas salientes da cutícula nos lábios e na cabeça.
Prováveis mecanorreceptores. Os deiridios ou papilas cervicais se encontram em muitas
espécies edáficas ou parasitas, mas acostumam faltar nas formas aquáticas.
Cerdas: cerdas aguçadas, na cabeça e corpo. São mecanorreceptores que quando se
estimulam fazem que o animal se afaste do estímulo.
Anfidios e fasmídios: são estruturas sensoriais pares quimiorrecptoras exclusivas do Filo
Nematoda. Os anfidios se localizam lateralmente na cabeça, atrás das cerdas cefálicas, que
desembocam ao exterior por um poro cuticular que expõe cílios modificados ao ambiente
(até a descoberta dos cílios modificados por microscopia eletrônica, os nematódeos eram
considerados o único filo no qual os cílios estavam ausentes). Os anfidios alcançam seu
máximo desenvolvimento nos nematódeos marinhos, são invaginações cegas, tubulares ou
saculiformes da cutícula, que contém um poro exterior, um ducto e uma bolsa anfidial
ligada ao nervo do anfidio. Em alguns nematódeos, como Ancelostoma, os anfidios se
conectam com grandes glândulas anfidiales (glândulas cefálicas), estendidas na cavidade
corporal. Estas estruturas bilateralmente simétricas são de grande importância taxonômica.
Basicamente tem dos tipos: espiral ou não-espiral. Usualmente a forma não-espiralada tem
uma estrutura em forma de bolso onde a abertura externa é em forma de fenda que conduz a
uma cavidade (fovea) preenchida com substancia gelatinosa (corpus gelatum). O muco é
essencialmente glicoproteina e sua composição difere entre espécies. Nos afidios
espiralados a fovea é aberta e alongada, e pode dar uma volta a partir do nervo ou várias
voltas (fovea multiespiralada). Os fasmídios ou glândulas precaudais são um par de órgãos
glândulo-sensoriais unicelulares que desembocam separadamente a cada lado da cauda,
especialmente em espécies parasitas. Constam de uma glândula, um poro e terminações
nervosas. Os fasmídios se relacionam com a reprodução e mostram dimorfismo sexual. Os
nematódeos marinhos de vida livre (Classe Adenophorea) carecem de fasmídios (=
Aphasmida).
Ocelos (manchas oculares): pouco freqüentes, uno a cada lado da faringe em nematódeos
aquáticos. No nematódeo marinho Deontostoma, cada ocelo consiste de um cálice
pigmentado contendo lamelas empilhadas derivadas de uma única célula.
Receptores de estiramento: em cordões epidérmicos, provavelmente regulam os
movimentos locomotores.
Reprodução
A reprodução é sempre sexual e a fecundação interna. Quase todos os nematódeos são de
sexos separados (dioicos), e na maioria dos casos o macho é menor que a fêmea. Os
machos apresentam caracteres sexuais secundários, tais como glândulas ventrais e lóbulos
caudais.
Existem alguns poucos nematódeos terrestres que são hermafroditas o partenogenéticos.
Existem casos em que se desconhecem os machos. As espécies hermafroditas são
protándricas, e apresentam um ovo-testículo. Em geral se autofecundam. Os
espermatozóides se desenvolvem primeiro e são estocados nas vesículas seminais. A
autofecundação ocorre depois da formação e maduração dos óvulos. Periodicamente surge
um pequeno número de machos que fecundam cruzadamente às hermafroditas. Em algumas
espécies terrestres ocorre partenogênese.
O sistema reprodutor é geralmente par. As gônadas, em número de uma ou duas, se
comunicam com o exterior por um poro único, a cloaca, nos machos, e um gonoporo ou
vulva nas fêmeas. A posição da vulva varia, sendo a vezes posterior e outras vezes anterior.
Nos machos, existe um testículo tubular, com forma de um cordão maciço. Em alguns
nematódeos existem dois testículos, orientados geralmente em forma oposta. Os testículos
se convertem imperceptivelmente em um comprido espermiducto ou ducto deferente. Cada
espermiducto se alarga no extremo posterior formando uma comprida vesícula seminal,
onde se acumulam os espermatozóides. Um ducto ejaculador muscular, com glândulas
prostáticas, conecta as vesículas seminais com a cloaca. As secreções prostáticas são
adesivas e possivelmente facilitam a cópula. A vesícula seminal desemboca no reto,
modificado em uma cloaca. A parede da cloaca está evaginada formando dos sacos que se
unem antes de desembocar na câmara cloacal.
A região posterior dos machos apresenta uma considerável variação. Acostuma estar
curvada em forma de gancho ou a cutícula alargada em expansões laterais (asas) com forma
de abanico, constituindo um acessório copulador chamado bursa. As vezes apresentam
papilas pedunculadas, cerdas sensoriais ou expansões a modo de ventosas. O poro genital
masculino está situado muito perto do ânus e têm ganchos cuticulares (espículas
copuladoras), bastonetes utilizados para segurar a fêmea durante a cópula e para manter
aberto o gonoporo feminino durante a transmissão dos espermatozóides. Cada saco contém
uma espícula, que geralmente é curta, com forma de folia aguçada e curva. As espículas
assomam a través da cloaca e saem pelo ânus, mediante músculos especiais, podem ser
evaginadas e retraídas na bolsa cutânea. As espículas copuladoras do macho assomam pela
cloaca e o ânus. Os espermatozóides, de distintas formas (redondos, cônicos, sinuosos ou
alargados), se podem mover lentamente em forma amebóide e carecem de flagelo. Os
espermatozóides podem estar formados por cabeça e cauda, a cauda acostuma possuir uma
comprida mitocôndria central com microtúbulos laterais.
Pode haver um ou dois ovários, tubulares, tipicamente pares. Normalmente uma gônada
está orientada para a parte anterior e a outra para a parte posterior, com seus extremos
opostos enfrentados. Em muitas espécies cada gônada se prega sobre si mesma e em
algumas espécies parasitas cada gônada é comprida e enrolada em espiral. A parte
germinativa é terminal, e nas espécies maiores as células acostumam agrupar-se em torno
de um cordão nutricional central (raquis). A célula mais interna de cada gônada, a célula do
extremo distal, secreta uma substancia promotora da mitose, que produz a proliferação de
núcleos de células germinais. Cada ovário se prolonga pouco a pouco convertendo-se
primeiro em oviducto tubular e depois em um útero comprido e muito amplio. Em alguns
casos existe um ovário único e um só oviducto. O extremo superior do útero pode funcionar
como receptáculo seminal. Cada útero desemboca em um tubo muscular curto comum,
denominado vagina. A vagina desemboca ao exterior pelo poro sexual geralmente impar
(vulva), situado ventralmente, geralmente na zona media do corpo. O poro sexual feminino
está situado na parte ventral do extremo anterior do corpo, ainda as vezes se traslada para as
proximidades do ânus.
Os ovos são pequenos, geralmente alongados e estão rodeados por envolturas muito
duras, que permitem esperar indefinidamente a aparição de condiciones ambientais
adequadas. Existem três cobertas: uma lipídica, outra cuticular e uma terceira protéica, com
ornamentações. São numerosos nas espécies parasitas. Por exemplo, una fêmea de Ascaris
coloca muitos milhões de ovos. Conhecem-se casos de fêmeas que produziram 27 milhões
de ovos, expulsando 200.000 diariamente. As espécies marinhas de vida livre raramente
colocam mais de 50 ovos, os quais são freqüentemente postos em agregados.
As fêmeas de algumas espécies produzem um feromônio que atrai aos machos. A
fecundação é interna. Durante a cópula, as espículas cloacais do extremo posterior em
forma de gancho do macho são expulsadas pela abertura cloacal, se enredam em torno à
região dos poros genitais da fêmea e se espetam no gonoporo feminino, mantendo-lhe
aberto. Os espermatozóides ameboides migram para a vagina e se dirigem ao receptáculo
seminal, no extremo superior do útero, onde ocorre a fecundação. O óvulo fecundado
secreta una grosa membrana de fecundação, que se endurece. A esta camada se agrega
outra coberta externa, secretada pelas paredes uterinas, que freqüentemente apresenta
estruturas características. A superfície dos ovos está esculpida de diferentes formas
específicas para cada espécie. Os ovos são retidos no útero durante algum tempo antes de
serem depositados. As vezes o desenvolvimento começa quando os ovos ainda estão dentro
da fêmea.
Desenvolvimento e Ciclo biológico
Alguns nematódeos são ovíparos, outros são ovovivíparos. O tempo necessário para
alcançar a etapa adulta varia desde uns poucos dias nos nematódeos livres, até mais de um
ano em alguns parasitas. O desenvolvimento é direto e estritamente determinado. O ovo
fecundado pode ser depositado sem haver começado a segmentação (por exemplos em
Ascaris), quando a segmentação se ha iniciado (por exemplo, em Ancelostoma), quando é
muito avançada (por exemplo em Enterobius), o quando o embrião está completamente
formado (por exemplo em Wuchereria). A clivagem prematura segue um padrão
assimétrico que pronto passa a ser bilateral, mas mostra por seu estreito determinismo leves
semelhanças com a segmentação espiral dos anelídeos, moluscos e alguns platelmintos. O
desenvolvimento embrionário leva à formação de três capas germinativas (ectoblasto,
mesoblasto, endoblasto), o blastoceloma surge em um espaço limitado por endoblasto e
ectoblasto. Os diversos órgãos apresentam um número relativamente fixo de células, o qual
se alcança no momento da eclosão. O desenvolvimento é direto. Dentro da envoltura do
ovo, a fase juvenil (denominada geralmente larva) realiza uma ou duas mudas. Existe um
incremento limitado do número de células durante as etapas juvenis, quase todo o
crescimento é conseqüência do incremento do tamanho celular. Os juvenis têm quase todas
as estruturas do adulto, exceto as partes do aparato reprodutor. O crescimento se
acompanha de quatro mudas da cutícula. A terceira fase é em muitas espécies a fase de
dispersão. Os adultos no mudam, mas alguns continuam crescendo.
Muitos nematódeos são capazes de suspender seus processos vitais quando as
condições ambientais são desfavoráveis, nesses estados resistentes eles podem sobreviver
situações extremas de falta de umidade, calor o frio, e voltar a viver quando as condições
favoráveis retornarem. Este processo é denominado criptobiosis, e é uma característica
compartida com rotíferos e tardígrados.
Parasitismo
Dentro dos nematódeos marinhos existe uma grande variedade de espécies parasitas de
mamíferos marinhos, peixes e invertebrados, e ocasionalmente o homem (zoonosis).
Geralmente estas espécies têm quatro estádios larvais que podem alternar em vários
hospedeiros intermediários. A maioria dos nematódeos marinhos parasitas pertencem a
classe Secernentea (= Phasmida), uma dos grupos mais estudados é a dos nematódeos
ascaroideos da familia Anisakidae. A anisakiosis e uma parasitose que pode ter ao homem
como um hospedeiro ocassional (antropozoonose) (Cabrera & Altamirano, 2004)
De um modo geral as formas adultas habitam no trato digestivo (estômago e intestino) de
mamíferos marinos. Na mucosa gástrica, os vermes se agrupam entre si produzindo uma
úlcera crateriforme muito característica. Os ovos, quando se expulsam pelas fezes, no estão
embrionados (L1), madurecem no mar até formar-se a larva de segundo estádio (L2) que
fica livre na água ao quebrar-se o ovo. Esta larva se for ingerida por pequenos crustáceos
do plâncton, primeiro hospede intermediário, muda à larva L3, alcançando um tamanho de
uns 3 mm. A ingestão dos crustáceos infectados por peixes marinhos e cefalópodes permite
que a L3 migre do intestino à cavidade peritoneal, e de lá aos tecidos, crescendo ate uns 2-3
cm. Por depredação, podem ter vários passos de um peixe ou cefalópode a outro, mas não
completam o ciclo a menos que cheguem ao estômago dos mamíferos marinos. Neste
hospede definitivo a L3 se adere à parede gástrica evolucionando ao quarto estádio larvário
(L4) e depois a adulto, fechando-se o ciclo. Em resumo, os mamíferos marinhos albergam
L3, L4 e adultos em seu trato digestivo. Neste ciclo se intercalam diversas espécies de
peixes e cefalópodes que atuam como hospedadores paraténicos o de transporte. Neles as
L3, liberadas em seu trato digestivo, se encapsulam no tecido muscular e vísceras,
conservando sua capacidade infestante.
O homem é um hospede acidental que se interpõe no ciclo quando ingere peixes ou
cefalópodes crus ou pouco cozidos (sushi, sashimi, seviche), assim como defumados,
semiconservas, salgados, peixe seco ou em vinagre que albergam as L3 de estes
nematódeos. As espécies de peixes descritas como parasitadas são muitas, incluindo o
bacalhau, cavala, anchova, sardinha, arenque, salmão, bonito e no Brasil pargo, dourado e
peixe espada (Barros & Matos, 1993). Entre os cefalópodes, os mais parasitados são a lula e
a sépia. Os hospedeiros definitivos são cetáceos ou pinípedes de águas frias tais como
baleias, golfinhos, belugas, focas y leões marinhos, entre outros. De entre as diferentes
espécies que causam a anisakiosis a mais freqüente no homem é Anisakis simplex, um
trabalho recente de Barros & Cavalcanti (2005) realizado no Nordeste do Brasil destacou o
gênero Contracaecum como o principal nematódeo parasita em pargos e dourados.
Importância econômica
São poucos os animais ou plantas que carecem de nematódeos parasitas. Incidem no ser
humano, como parasitas próprios, de seus animais domésticos e de seus cultivos. Existem
numerosos nematódeos parasitas do ser humano que causam enfermidades que não foram
ainda controladas pela medicina, e os que parasitam aos animais domésticos causam
perdidas por valores de milhares de milhões de dólares. Considera-se que unas 15 espécies
de nematódeos parasitam habitualmente ao ser humano, mas outras 15 adicionais que se
encontram habitualmente em outros animais, podem parasitar-lo ocasionalmente
(zoonosis). Existem vários nematódeos pequenos (1-2 mm de largo) que produzem lesões
em vegetais úteis, como herbáceas, flores, frutos e árvores. Calcula-se que consomem
aproximadamente um 10% dos cultivos. Prejudicam a seu hóspede por sua ação corrosiva,
porque esgotam a seiva da planta, lesam os pontos de crescimento, e pela transmissão de
vírus ou ao abrir caminhos de entrada a bactérias ou fungos. Os principais médios de
controle são a rotação de cultivos, a esterilização da terra e o desenvolvimento de
variedades de plantas resistentes.
Entre os aspectos positivos que tem os nematódeos para o ser humano, deve considerar-se
que são parcialmente responsáveis da destruição de muitos cadáveres, atacam a muitos
animais e vegetais prejudiciais para o ser humano e que as atividades dos nematódeos do
solo facilitam a aeração do solo e a circulação e mineralização de componentes minerais e
orgânicos.
Como uma alternativa ao uso de inseticidas químicos se propuz o controle biológico com
nematódeos parasitas. Baseia-se na aplicação de um biolarvicida a base de nematódeos
parasitas Romanomermis culicivorax e Romanomermis iyengari. Estes biolarvicidas se
caracterizam por ser específicos para larvas de mosquitos, não afetando flora, fauna e o
homem. Outra característica de estes biolarvicidas é que se podem obter ou produzir a
baixo custo, com matérias primas locais. Desde o ponto de vista científico, deve destacar-se
que o nematódeo Caenorhabditis elegans é um dos animais mais estudados. Conhece-se o
destino de cada uma de suas células durante o curso do desenvolvimento e seu genoma é
um dos melhores conhecidos. Estudos genéticos sob esta espécie deu o Premio Nobel de
Medicina do ano 2002 aos pesquisadores Brenner, Horvitz & Sulston.
Os projetos de pesquisa modernos que visam o estudo da biodiversidade marinha como
base para entender a estrutura, dinâmica e resilencia do ecossistema estão centralizando
seus focos de estudos nos nematódeos. Este grupo poderia ser um médio factível para
entender os mecanismos que geram e mantém a diversidade biológica. Como primeiro
ponto, a maioria dos nematódeos são morfologicamente uniformes, e, conseqüentemente,
sua identificação é dificultosa. Os estilos de vida dos nematódeos aquáticos, de qualquer
modo mostram uma ampla variação. Existem consumidores de algas seletivos e
consumidores de detrito/bactérias, predadores e espécies (as vezes sem boca) que se
alimentam dos nutrientes dissolvidos na água. Dentro de uma pequena amostra podem
ocorrer espécies com um ciclo de vida de poucos dias e outras que completam seu ciclo
vital em um ano. Devido à grande variedade de estilos de vida, estudos ecológicos que
tratam aos nematódeos como um grupo uniforme não tem muito valor no presente; por tal
ração muitos grupos de pesquisa estão trabalhando simultaneamente em diferentes projetos
de pesquisa sobre taxonomia-anatomia e ecologia-fisiologia dos nematódeos marinhos.
(Riemann, 2003).
A classificação mais utilizada por especialistas atualmente considera a existência de
duas classes:
CLASSE SECERNENTEA (= Phasmida)
1. Anfídeos labiais, aberturas em forma de poro ou fenda.
2. Fasmídios presentes, geralmente posteriores.
4. Sistema excretor tubular
5. Cutícula com 2 a 4 camadas, estriada.
6. Esôfago variado com 3 glândulas esofágicas
7. Macho geralmente com um testículo
8. Asa caudal freqüente
9. Papilas sensoriais cefálicas, e caudais em alguns machos.
Compreendem parte das espécies que habitam o solo, a maioria das parasitas
animais e quase todos as parasitas dos vegetais, raramente aquáticos. Entre os fásmidos
encontram-se os Ascaris, oxiúros e filarias.
Subclasse Rhabditia: livres terrestre e parasitas animais (Mamíferos marinhos, pinípedes)
Subclasse Spiruria: todos parasitas animais
Ordem Ascaridida (parasitas de peixes e mamíferos marinhos família
Anisakidae)
Ordem Spirurida (parasitas de mamíferos marinhos, pinípedes, belugas,
baleias e cachalotes, Placentonema gigantissima)
Ordem Camallanida – (parasitas de vertebrados terrestres e aquáticos, com
copépodos como hospedeiros secundários) Condrictios principalmente tubarões
Subclasse Diplogasteria. Livres terrestes, parasitas de insetos e plantas.
CLASSE ADENOPHOREA (= Aphasmida)
1. Anfídeos pós-labiales de formas variáveis (poro a elaborada).
2. Fasmídeos ausentes.
3. Sistema excretor simples, não tubular, células glandulares.
4. Macho geralmente com dois testículos
5. Asa caudal geralmente ausente
6. Papilas sensoriais cefálicas e ao longo do corpo.
Maioria de vida livre. Compreendem quase todas as espécies aquáticas, tanto de
água doce como oceânicas, parte das espécies do solo, parte das parasitas animais e alguns
parasitas dos vegetais. 12 ordens.
Subclasse Enoplia: de vida livre e parasita, terrestre, marinho e raramente de água doce.
Parasitas de invertebrados marinhos (poliquetas, priapúlidos, crustáceos,
holoturias)
Subclasse Chromadoria todos de vida livre, a maioria marinhos
No Brasil segundo o estudo da Avaliação do estado do conhecimento da diversidade
biológica do Brasil de Invertebrados Marinhos (Migotto & Marques, 2003) Brasil tem entre
200-430 espécies de nematódeos marinhos. Existindo pouquíssimos especialistas no Brasil:
Liliana Rúbia de A. Medeiros (Universidade de Ibirapuera) e Thais Navajas Corbisier
(Instituto de Oceanografia da USP), e nos últimos anos na UFPE o laboratório de
Meiofauna coordenado pela Prof. Verônica Genovois vem trabalhando com taxonomia de
nematódeos livres marinhos.