UNIVERSIDADE FEDERAL DO VALE DO SÃO FRANCISCO
PROGRAMA DE PÓS-GRADUAÇÃO EM RECURSOS NATURAIS DO
SEMIÁRIDO
PALOMA CLEMENTINO DA CRUZ LUBARINO
AVALIAÇÃO FENOTÍPICA E BIOQUÍMICA DE Citrullus lanatus L.
PARASITADAS POR Meloidogyne enterolobii
PETROLINA
2015
PALOMA CLEMENTINO DA CRUZ LUBARINO
AVALIAÇÃO FENOTÍPICA E BIOQUÍMICA DE Citrullus lanatus L.
PARASITADAS POR Meloidogyne enterolobii
Dissertação apresentada à Universidade
Federal do Vale do São Francisco, como
parte das exigências do Programa de
Pós-Graduação em Recursos Naturais
do Semiárido, para obtenção do título de
Mestre em Recursos Naturais
Orientador: Prof. Dr. Leonardo Sousa
Cavalcanti
Co-orientador: Dr. Pedro Martins Ribeiro
Junior
PETROLINA
2015
L926a
Lubarino, Paloma Clementino da Cruz
Avaliação fenotípica e bioquímica de Citrullus lanatus
L. parasitadas por Meloidogyne enterolobii / Paloma
Clementino da Cruz Lubarino. -- Petrolina, 2015.
xvi; 89 f.: il. 29 cm.
Dissertação (Mestrado em Recursos Naturais do
Semiárido) - Universidade Federal do Vale do São
Francisco, Campus Petrolina, Petrolina- PE, 2015.
Orientador: Prof. Dr. Leonardo Sousa Cavalcanti.
1. Nematoide das galhas. 2. Citrullus lanatus.
3. Mecanismos de defesa. I. Título. II. Universidade
Federal do Vale do São Francisco.
CDD 581.19
Ficha catalográfica elaborada pelo Sistema Integrado de Biblioteca SIBI/UNIVASF
Bibliotecário(a): Maria Betânia de Santana da Silva – CRB4-1747.
A Deus, aos meus pais Antonio José Lubarino e
Maria Oneide Clementino da Cruz Lubarino, ao meu
irmão Radamés Clementino da Cruz Lubarino, ao
meu filho Iago Clementino Lubarino Nogueira, ao meu
namorado Manicio Antonio Antas Souza que
acreditaram em mim e me apoiaram em todos os
momentos.
Dedico
AGRADECIMENTOS
Agradeço a Deus, por me manter perseverante em meus propósitos.
Agradeço à Nossa Senhora, por me cobrir com seu manto protetor, fortalecendo- me
mesmo nas dificuldades.
Ao meu orientador, Prof. Dr. Leonardo Sousa Cavalcanti, a quem agradeço pela
atenção e pela oportunidade de ser sua orientada.
Ao meu co-orientador, Dr. Pedro Martins Ribeiro Júnior por todas as contribuições
científicas, pela paciência e técnicas ensinadas.
A minha supervisora da Embrapa, Dr.ª Rita de Cássia Souza Dias, a quem agradeço
pela oportunidade de desenvolvermos os trabalhos de melhoramento, fase
fundamental para seguir com as demais etapas deste trabalho.
Ao Programa de Pós-graduação em Recursos Naturais do Semiárido, aos
professores por me ajudarem como agrônoma, a aprimorar o conhecimento químico
tão relevante para minha formação, a Nancy Freire Cavalcante, pessoa-chave no
colegiado da pós-graduação, que radia simpatia e carisma.
A Embrapa Semiárido pelo suporte para o desenvolvimento deste trabalho, como
casa de vegetação, as sementes dos genótipos do programa de melhoramento de
melancia visando resistência a M. enterolobii, os laboratórios e outros.
As colegas de turma, Amanda Leite Guimarães, Ana Paula de Oliveira, Deize Raquel
dos Reis Cruz, Eriane Dantas Bezerra, Fernanda Granja da Silva Oliveira, Georgia
Sueley Bezerra, Jackeline Ferreira Gomes, Kíscyla Oliveira de Andrade, Naiane
Darklei dos Santos Silva e Sandra Kelle Souza Macedo pelo apoio, momentos de
estudo e amizade.
A Antonio José Lubarino e Maria Oneide Clementino da Cruz Lubarino, meus pais,
por todo amor incondicional e valores ensinados. Por me lembrarem sempre que
Deus está acima de tudo, que Ele é nosso conforto, nossa diretriz e por mais que
existam dificuldades, elas existem para serem superadas.
A Radamés Clementino da Cruz Lubarino, meu irmão, sinônimo de determinação, a
quem agradeço por sempre me ejetar força, coragem, a ele devo a ousadia em
buscar sempre crescer mais e mais.
A Iago Clementino Lubarino Nogueira, meu amado filho, por quem tenho amor maior
e infinito. Esteve presente nas minhas outras formações (Biologia e Agronomia) e,
hoje, mamãe dedica mais um título a você.
A Manicio Antonio Antas de Souza, meu noivo, amigo e parceiro que esteve presente
em algumas etapas deste trabalho, até irrigar as plantinhas, ele irrigou comigo nos
finais de semana. Obrigada, por me transmitir carinho, amor e afetividade, o
sentimento é recíproco.
A todos, o meu “Muito obrigada”!
“Cada sonho que você deixa para trás, é um pedaço
do seu futuro que deixa de existir”.
(Steve Jobs)
RESUMO
O trabalho objetivou selecionar plantas da população de melancia forrageira acesso
BGCIA 240, oriunda do Banco Ativo de Germoplasma de Cucurbitáceas da Embrapa
Semiárido resistentes a Meloidogyne enterolobii, comparar os métodos não
destrutivos com os métodos tradicionais destrutivos; e estudar mecanismos
bioquímicos envolvidos nessa resistência. Para a seleção de plantas resistentes,
foram realizadas avaliações aos 30 dias após a inoculação (DAI) pelos métodos não
destrutivos: desenvolvimento global da parte aérea (DGPA) e de galhas (DGG)
classificando-as com grau de resistência (R),
média resistência (MR)
e
suscetibilidade (S). Após a seleção, as plantas foram transplantadas para vasos de
30 L e avaliadas aos 90 DAI pelos métodos não destrutivos e destrutivos
(desenvolvimento global do sistema radicular (DGSR), de galhas (DGG) e fator de
reprodução (FR). Para o estudo de mecanismos bioquímicos envolvidos na defesa
da melancia ao nematoide foi utilizada um acesso resistente selecionado neste
trabalho comparado com um acesso de melancia BGCIA 875 suscetível a esse
nematoide. Os acessos foram inoculados e avaliados quanto ao número de galhas
(NG), número de ovos por planta (NO), comprimento (CR) e massa fresca de raiz
(MFR) aos 7, 14, 21, 28 e 35 DAI. Avaliou-se também os teores de clorofila e
atividade de enzimas relacionadas à defesa. Pelo método não destrutivo foi
selecionado 46 % de plantas R. A avaliação não destrutiva baseada no DGPA e
DGG das plantas do acesso BGCIA 240 apresentou correlação positiva na
classificação precoce da reação de resistência a M. enterolobii com o FR. Com
relação a reação ao patógeno, o acesso resistente apresentou menor NG e NO, e
aos 14 DAI, apresentou maior atividade das enzimas guaiacol peroxidase, ascorbato
peroxidase, polifenoloxidases e fenilalanina amônia-liase. O uso dos métodos não
destrutivos e o estudo de respostas de defesa na melancia podem auxiliar na
seleção de genótipos visando à resistência a M. enterolobii.
Palavras-chave: nematoide das galhas, Citrullus lanatus, mecanismos de defesa.
ABSTRACT
The study aimed to select plants a population of forage watermelon access BGCIA
240, resistant to Meloidogyne enterolobii from the Cucurbitaceae Germplasm Active
Bank of Embrapa Semi-Arid, methods avoid destructive traditional methods, were
comparad in this, as well as study biochemical mechanisms involved in this
resistance. For the selection of resistant plants an assessments was carried out up
to 30 days after inoculation (DAI) by means of non-destructive approaches evaluating
global development of shoots (DGPA) and galls (DGG) classifying them with degree
of resistance (R), average resistance (MR) and susceptible (S). After selection, these
plants were transplanted and evaluated at 90 DAI by non-destructive and destructive
(overall development of the root system (DGSR), galls (DGG) and reproduction factor
(FR) approaches. For the study of biochemical mechanisms involved in the defense
of watermelon against nematode infection, resistant plants were selected and
compared to a watermelon access BGCIA 875 (susceptible to this nematode). The
plants were inoculated and evaluated as the number of galls (NG), number of eggs
per plant (NO), length (CR) and fresh pasta from root (MFR) at 7, 14, 21, 28 and 35
DAI. The chlorophyll content and enzymes activity related to defense, were also
assessed. The non-destructive method analyses selected 46% of plants as non
susceptible, based on DGPA and DGG analyses. Plant access BGCIA 240 showed a
positive correlation in the early classification of the M. enterolobii resistance reaction
to the FR. Regarding the reaction to pathogen resistant access showed less NG and
NO, and 14 DAI, higher activity of enzymes guaiacol peroxidase, ascorbate
peroxidase, polyphenol and phenylalanine ammonia lyase. The use of nondestructive methods and study biochemical of defense responses in watermelon can
help in the early selection of forage watermelon genotypes for resistance to M.
enterolobii.
Key words: root-knot nematodes, Citrullus lanatus, defense mechanism.
LISTA DE FIGURAS
Figura 1. Espécie vegetal Citrullus lanatus (A), frutos da melancia forrageira (C.
lanatus var. citroides) (B) e da melancia comercial (C. lanatus var. lanatus) (C).......24
Figura 2. Raízes com galhas (A) e parte aérea de Citrullus lanatus var. lanatus
infectada (B) por Meloidogyne enterolobii..................................................................25
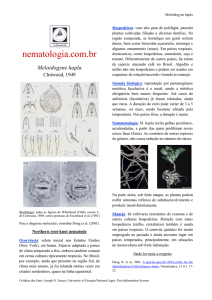
Figura 3. Ciclo de vida do nematoide das galhas, Meloidogyne spp. A terceira fase
juvenil (J3) não está ilustrada na figura. J2 corresponde à segunda fase juvenil e J4
à quarta fase (Fonte: PERRY & MOENS, 2006)........................................................26
Figura 4. Etapas da lavagem de raízes inoculadas do acesso BGCIA 240, para
avaliação aos 30 dias após inoculação. Retirada da planta (A), lavagem e retirada do
excesso de solo nas raízes (B)..................................................................................41
Figura 5. Aspecto geral das plantas em vasos de 30 litros de solo para obtenção de
descendência em condições de telado......................................................................42
Figura 6. Classes do sistema radiculares do acesso BGCIA 240, colhido e avaliado.
Resistente (A), média resistência (B) e suscetível (C) em função do desenvolvimento
global da parte aérea (DGPA) e de galhas (DGG).....................................................47
Figura 7. Número de galhas (A) e número de ovos (B) por planta de dois genótipos
de melancia do Banco Ativo de Germoplasma (BAG) de Cucurbitáceas da Embrapa
Semiárido, acesso BGCIA 240 (resistente) e acesso BGCIA 875 (suscetível)
inoculadas com Meloidogyne enterolobii, avaliadas em diferentes épocas de coleta. *
Significativo a 5% de probabilidade e
ns=
não significativo pelo teste T....................55
Figura 8. Proteínas solúveis totais (A), atividades das enzimas guaiacol peroxidase
(POX) (B), polifenoloxidase (PPO) (C), ascorbato peroxidase (APX) (D), catalase
(CAT) (E) e fenilalanina amônia liase (PAL) (F) de dois genótipos de melancia do
Banco Ativo de Germoplasma (BAG) de Cucurbitáceas da Embrapa Semiárido,
acessos BGCIA 240 e BGCIA 875 inoculadas com Meloidogyne enterolobii, e seus
controles, avaliadas em diferentes épocas de coleta. Barras representam erro
padrão da média.........................................................................................................59
LISTA DE TABELAS
Tabela 1. Caracterização física e química do solo autoclavado utilizado nos
experimentos. Embrapa Semiárido, Petrolina-PE, 2015............................................40
Tabela 2. Avaliação preliminar dos graus de resistência e suscetibilidade em
melancia forrageira acesso BGCIA 240, inoculadas com Meloidogyne enterolobii aos
30 dias após inoculação (DAI) e dados de amplitude do fator de reprodução (FR).
Petrolina, Embrapa Semiárido, 2015..........................................................................47
Tabela 3. Dados de amplitude do acesso BGCIA 240 inoculadas com Meloidogyne
enterolobii avaliadas quanto ao número de ovos/mL de solução do sistema radicular.
Petrolina, Embrapa Semiárido, 2015..........................................................................48
Tabela 4. Valores dos coeficientes de correlação para a associação entre as
variáveis desenvolvimento global de galhas (DGG) e da parte aérea (DGPA) de
planta do acesso BGCIA 240 aos 30 dias após a inoculação. Petrolina, Embrapa
Semiárido, 2015.........................................................................................................49
Tabela 5. Valores dos coeficientes de correlação para a associação entre as
variáveis desenvolvimento global do sistema radicular (DGSR), de galhas (DGG),
fator de reprodução (FR) e desenvolvimento global da parte aérea (DGPA) de
plantas do acesso BGCIA 240 aos 90 dias após a inoculação. Petrolina, Embrapa
Semiárido, 2015.........................................................................................................52
Tabela 6. Comprimento de raiz, massa fresca de raiz, clorofila a e clorofila b de dois
genótipos de melancia do Banco Ativo de Germoplasma de Cucurbitáceas da
Embrapa Semiárido, acessos BGCIA 240 e BGCIA 875 inoculadas e não inoculadas
com Meloidogyne enterolobii em diferentes épocas de avaliação.............................55
LISTA DE ABREVIATURAS E SIGLAS
APX
Ascorbato peroxidase
BAG
Banco Ativo de Germoplasma
CAT
Catalase
CCP
Citocromo c peroxidase
CF
Comprimento da primeira folha verdadeira
CPA
Comprimento da parte aérea
CSR
Comprimento do sistema radicular
cv.
Cultivar
DAI
Dias após a inoculação
DF
Diâmetro da primeira folha verdadeira
DGG
Desenvolvimento global de galhas
DGPA
Desenvolvimento global da parte aérea
DGSR
Desenvolvimento global do sistema radicular
DH
Diâmetro do hipocótilo
DMSO
Dimetilsulfóxido
EAO's
Espécies ativas de oxigênio
EC
Enzyme commission
f. sp.
forma specialis
FC
Final do ciclo
FR
Fator de reprodução
H2O2
Peróxido de hidrogênio
HR
Resposta de hipersensibilidade
IFC
Índice Falker de Clorofila
J2
Juvenil de segundo estágio
J3
Juvenil sedentário
J4
Juvenil no início da fase reprodutiva
Lip
Lignina peroxidase
MDA
Monodehidroascorbato
MFSR
Massa fresca do sistema radicular
MnP
Peroxidase dependente de manganês
MR
Média resistência
NO
Número de ovos
O2-
Oxigênio molecular
OH-
Radical hidroxila
PAL
Fenilalanina amônia liase
PFOs
Polifenoloxidases
POX
Guaiacol peroxidase
PPO
Polifenoloxidase
PR-proteínas
Proteínas relacionadas à patogênese
R
Resistente
S
Suscetível
SAR
Resistência sistêmica adquirida
VSR
Volume do sistema radicular
SUMÁRIO
1. INTRODUÇÃO.......................................................................................................18
2. OBJETIVOS...........................................................................................................21
2.1. Objetivo Geral......................................................................................................21
2.2. Objetivos Específicos..........................................................................................21
3. FUNDAMENTAÇÃO TEÓRICA.............................................................................22
3.1. Botânica de Citrullus lanatus L.............................................................................22
3.2. Nematoides das galhas da melancia...................................................................24
3.3. Recursos genéticos: fontes de resistência de Citrullus lanatus var. citroides.....28
3.4. Melhoramento
genético
de
Citrullus
lanatus
L.
visando
resistência
à
nematoides.................................................................................................................29
3.5. Mecanismos de defesa de plantas à nematoides................................................30
3.5.1. Espécies ativas de oxigênio e enzimas antioxidantes.....................................31
3.5.1.1. Peroxidases...................................................................................................32
3.5.1.2. Ascorbato Peroxidase...................................................................................33
3.5.1.3. Catalase........................................................................................................34
3.5.2. Polifenoloxidase...............................................................................................35
3.5.3. Fenilalanina amônia-liase.................................................................................36
3.6. Proteínas relacionadas à patogênese.................................................................37
3.7. Clorofila............................................................................................................... 38
4. PARTE EXPERIMENTAL.......................................................................................40
4.1. Material vegetal e cultivo das plantas..................................................................40
4.2. Preparo do inóculo e inoculação.........................................................................40
4.3. Seleção de plantas resistentes à M. enterolobii na população do acesso BGCIA
240..............................................................................................................................41
4.4. Avaliação dos métodos não destrutivos com os métodos tradicionais
destrutivos..................................................................................................................42
4.5. Reação de genótipos de melancia de mesa e forrageira ao patógeno...............43
4.6. Análises de clorofila e enzimáticas......................................................................43
4.7. Delineamento experimental e análise estatística................................................44
5. RESULTADOS E DISCUSSÃO.............................................................................46
5.1. Seleção de plantas resistentes à M. enterolobii na população do acesso BGCIA
240..............................................................................................................................46
5.2. Correlação dos métodos não destrutivos com os métodos tradicionais
destrutivos..................................................................................................................49
5.3. Reação de genótipos de melancia de mesa e forrageira ao patógeno...............53
5.4. Respostas bioquímicas de defesa.......................................................................56
6. CONCLUSÕES......................................................................................................60
REFERÊNCIAS..........................................................................................................61
APÊNDICE.................................................................................................................72
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
18
1. INTRODUÇÃO
A melancia forrageira (Citrullus lanatus (Thumb.) Matsum & Nakai var.
citroides (L. H. Bailey) Mansf.) pertence à família Cucurbitaceae, e seu fruto
apresenta casca resistente a impactos e à deterioração, polpa branca
geralmente consistente com baixo teor de sacarose (OLIVEIRA et al., 2000).
Muitas vezes são conservadas por mais de um ano sem perder suas qualidades
nutricionais, sob sol escaldante nas áreas secas do Nordeste brasileiro e sem
necessidade de práticas sofisticadas de armazenamento. A espécie forrageira é
um dos recursos alimentares à criação pecuária da região. Em um hectare, a
depender da quantidade e da distribuição das chuvas, pode chegar a produzir de
25 a 30 toneladas (OLIVEIRA et al., 2000; ARAÚJO et al., 2013).
Alguns estudos também descrevem a importância da espécie C. lanatus
var citroides como porta-enxerto (GAMA et al., 2010; 2013). Na maioria das
vezes, genótipos desta espécie possuem sistema radicular mais vigoroso que o
da planta enxertada, sendo utilizado como alternativa a curto prazo para o
manejo de plantas em solos com problemas de patógenos, salinidade ou mesmo
oscilações de temperatura (RIZZO et al., 2004).
A longo prazo, a melancia forrageira, assim como outras Cucurbitáceas tem
sido utilizada em programas de melhoramento genético visando identificação de
fontes de resistência aos patógenos do solo. Entre estes, o nematoide de galhas
(Meloidogyne spp.) estão entre os patógenos mais comuns que limitam a
qualidade e a produção das Cucurbitáceas. As espécies desse gênero de maior
ocorrência no Brasil são M. incognita (Kofoid & White) Chitwood, M. javanica
(Treub) Chitwood e M. arenaria (Neal) (PEREIRA et al., 2009). Contudo, no
Submédio do Vale do São Francisco a espécie M. enterolobii (sin. M.
mayaguensis) vem, nos últimos anos, causando prejuízos aos produtores de
Cucurbitáceas e de outras culturas (TERAO et al., 2010). O controle desses
nematoides na melancia é difícil devido a ampla gama de plantas hospedeiras e
não existem nematicidas registrados no Brasil para o seu controle (AGROFIT,
2015). Ademais, os nematicidas químicos apresentam alta toxidez ao homem e
ao ambiente e tem alto custo aos produtores. Assim, para o controle destes
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
19
fitopatógenos, a resistência genética tem sido adotada como alternativa mais
econômica para o produtor e mais sustentável para o ambiente (LIMA et al.,
2005).
Neste contexto, experimentos mostraram que acessos de Cucurbitáceas
como a bucha (Luffa cylindrica (L.) Roem.), abóbora Goianinha (Cucurbita
moschata Duchesne), abóbora Mini Paulista (Cucurbita moschata Duchesne),
melão Redondo Amarelo (Cucumis melo L. var. inodorus Naud) e melancia
Charleston Gray (Citrullus lanatus (Thunb.) Matsum. & Nakai var. lanatus) foram
resistentes a M. incognita (FRANCO et al., 2008). Em outros trabalhos, Castro et
al. (2011) e Damaceno (2012), relataram a melancia forrageira (Citrullus lanatus
var. citroides) como fonte de resistência contra M. enterolobii, com uso potencial
tanto como porta-enxerto, quanto para cruzamentos com melancia de mesa
(Citrullus lanatus (Thumb.) Matsum & Nakai). Apesar destes relatos, existem
poucos
trabalhos
apresentando
estudos
de
mecanismos
bioquímicos
relacionados à defesa dessas acessos resistentes quando inoculadas com
nematoides do gênero Meloidogyne.
Ao reconhecer o patógeno, em plantas resistentes ocorre o aumento dos
níveis de defesa pela ativação de genes que codificam diversos compostos
como enzimas antioxidantes, proteínas relacionadas à patogênese e enzimas
precursoras de fitoalexinas como a fenilalanina amônia-liase (PAL) (KOVÁCIK et
al, 2007).
Estudos da associação da expressão destes compostos de defesa e de
genótipos resistentes a nematoides em outras olerícolas foram realizados. Na
família das Solanáceas, o tomate e batata tem mostrado associação da enzima
PAL como um fator de resistência a patógenos. Brueske (1980) e Kalaiarasan
(2009) observaram em raízes de tomate resistentes e inoculadas com M.
incognita menor número de galhas por planta e um aumento significativo na
atividade da PAL às 108 horas após a inoculação, superiores à testemunha
suscetível que apresentaram menor atividade dessa enzima. Em raízes de
batatas resistentes e inoculadas com o nematoide Globodera rostochiensis
foram observadas maiores atividades da PAL do que em raízes de plantas
suscetíveis (GIEBEL, 1973).
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
20
De acordo com um levantamento bibliográfico, ainda não foi encontrado
estudo fitoquímico de genótipos de melancia relacionados com a resistentes a
nematoide de galhas.
Diante do exposto, o trabalho tem como objetivos selecionar dentro de
uma população de Citrullus lanatus var. citroides plantas com graus de
resistência a Meloidogyne enterolobii, correlacionando e validando os métodos
de avaliação não destrutivos com os métodos tradicionais destrutivos. Assim
como, avaliar enzimas relacionadas à defesa de plantas à patógenos e clorofilas
em genótipos de melancia forrageira resistente comparando com genótipo de
melancia de mesa suscetível a M. enterolobii.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
21
2. OBJETIVOS
2.1. Geral
Os objetivos gerais deste trabalho foi selecionar plantas de melancia
forrageira com resistência ao nematoide das galhas Meloidogyne enterolobii;
comparar os métodos não destrutivos com os métodos tradicionais destrutivos e
estudar os aspectos bioquímicos relacionados à defesa da planta a esse
patógeno.
2.2. Específicos
Selecionar dentro de uma população de Citrullus lanatus var. citroides
plantas com grau de resistência a M. enterolobii;
Validar métodos não destrutivos na avaliação de resistência a M. enterolobii,
correlacionando-os com os métodos tradicionais destrutivos;
Verificar a resposta de reação dos genótipos de C. lanatus var. lanatus e C.
lanatus var. citroides a M. enterolobii;
Avaliar as enzimas relacionadas à defesa de plantas à patógenos, guaiacol
peroxidase, polifenoloxidase, catalase, ascorbato peroxidase e fenilalanina
amônia-liase e clorofilas em genótipo de melancia forrageira resistente
comparado com genótipo de melancia de mesa suscetível a M. enterolobii;
Contribuir para o estudo fitoquímico de genótipos de melancia relacionados
com a resistencia a nematoide de galhas.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
22
3. FUNDAMENTAÇÃO TEÓRICA
3.1. Botânica de Citrullus lanatus L.
Plantas da família Cucurbitaceae estão amplamente distribuídas pelo
mundo todo. Em regiões da América do Norte, América do Sul, Norte da Ásia,
Sul da Europa, Ásia Tropical e Temperada, ocupa lugar de destaque entre as
hortaliças mais cultivadas (SATURNINO et al., 1982).
A família Cucurbitaceae é constituída por plantas caracterizadas como
ervas anuais ou perenes, subarbustos escandentes ou prostrados. Podem
apresentar gavinhas, que quando presentes são simples ou ramificadas sendo
originadas de modificações dos ramos, ao passo que as folhas são alternas,
simples ou lobadas. As flores são perfeitas ou unissexuadas, com plantas
monoicas, dioicas ou ainda andromonoicas, isoladas ou ordenadas em racemos,
espigas, fascículos ou panículas. A flor feminina apresenta hipâncio geralmente
alongado, ovário ínfero, tricarpelar, unilocular ou dividido em falsos lóculos pela
intrusão de placentas parietais. As flores masculinas contem cinco estames livres
entre si com ou sem estaminódios. O número de estames e a sua disposição,
constituem caráter de valor na sistemática das cucurbitáceas (BARROSO,
1978).
Essa família está classificada dentro da divisão Magnoliophyta, classe
Magnoliopsida, subclasse Rosidae, ordem Cucurbitales (BARROSO, 2001).
Embora não seja a maior família da ordem Cucurbitales, as Cucurbitáceas se
destacam por possuir cerca de 120 gêneros e 820 espécies, dentre as quais
encontra-se espécies de alta relevância econômica, de frutos comestíveis e
amplamente conhecidos, como os jerimuns (Cucurbita spp.), pepino, melão
(Cucumis spp.), e a melancia (Citrullus spp.) (NUEZ et al., 2000).
Esta última pertence ao gênero Citrullus que possui quatro espécies, sendo
três consideradas espécies de melancias selvagens anuais, C. colocynthis (L.)
Schrad, C. ecirrhosus Cogn e C. rehmmi Winter (MEEUSE, 1962) e a C. lanatus
Mansf. (melancia) encontrada em território brasileiro, a qual é consumida como
fruta fresca (ROMÃO, 1995; JOBST et al., 1998).
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
23
Citrullus lanatus (Thunb.) Matsum. & Nakai é uma espécie originária das
regiões quentes da África Tropical e aparentemente os primeiros cultivos
ocorreram a mais de 5000 anos nessa região (WHITAKER; DAVIS, 1962). Hoje,
seu cultivo, tanto quanto seu consumo está difundido nos países de clima quente
em regiões tropicais.
A espécie, ainda apresenta duas variedades botânicas, a melancia
cultivada para consumo humano, Citrullus lanatus var. lanatus e para
alimentação animal, Citrullus lanatus var. citroides (WHITAKER; DAVIS, 1962;
MOHR, 1986). No entanto, existem entre estas duas variedades, características
botânicas e morfológicas que as diferenciam, como por exemplo, o sistema
radicular, que é mais extenso nas variedades citroides justamente por se
tratarem de genótipos adaptados às condições de sequeiro.
Em geral, as folhas dessa espécie apresentam um tamanho médio, são
recortadas, triangulares, lobadas, alternas e de nervura palmeada. Quanto ao
florescimento, as plantas são monoicas, isto é, possuem flores masculinas e
femininas separadas na mesma planta, contudo, algumas populações são
andromonoicas, com flores hermafroditas e masculinas na mesma planta
(SOUZA, 2008). As flores femininas se diferenciam das flores masculinas pela
presença de um ovário súpero, bem visível (CASTELLANE; CORTEZ, 1995).
As flores ocorrem isoladamente, raramente agrupadas, axilares, opostas às
gavinhas, de cálice estrelado, esverdeado e corola dividida em cinco lóbulos, de
coloração amarela ou branca (CASTELLANE; CORTEZ, 1995).
Os frutos são indeiscentes, constituídos por uma baga, de paredes
externas duras e internas carnosas. Quando maduro, a coloração externa varia
de verde cana a verde escuro para as ambas variedades, a interna (parte
comestível) na var. lanatus é geralmente vermelha e macia. Já, a variedade
citroides apresenta a polpa branca e dura (LUBARINO et al., 2012) (Figura 1).
Segundo Nuez et al. (1998), o formato do fruto pode variar entre alongado,
elíptico, deprimido, ovalado e redondo para as ambas variedades.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
24
Figura 1. Espécie vegetal Citrullus lanatus (A), frutos da melancia forrageira (C.
lanatus var. citroides) (B) e da melancia comercial (C. lanatus var. lanatus) (C).
3.2. Nematoides das galhas da melancia
Os nematoides do gênero Meloidogyne, também conhecidos por
nematoides das galhas, estão entre os nematoides mais destrutivos que
infectam as cucurbitáceas (POFU et al., 2011). Esse gênero contém mais de 80
espécies e estão distribuídos em todo o mundo parasitando quase todas as
espécies de plantas superiores (PERRY; MOENS, 2006). As espécies do gênero
Meloidogyne mais importantes em cultivos de cucurbitáceas no Brasil são M.
incognita (Kofoid & White) Chitwood, M. javanica (Treub) Chitwood e M. arenaria
(Neal) Chitwood e são as espécies de maior ocorrência no mundo e no Brasil
(PEREIRA et al., 2009). Outro nematoide desse gênero que vem trazendo
prejuízos aos produtores de cucurbitáceas no Submédio Vale do São Francisco
é o M. enterolobii Yang & Eisenback, 1983 (sin. M. mayaguensis).
O primeiro relato no Brasil de M. enterolobii ocorreu em goiabeira
plantadas em perímetros irrigados de Petrolina (PE) e Juazeiro (BA),
representando ameaça potencial para os produtores de culturas suscetíveis no
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
25
Semiárido brasileiro (CARNEIRO et al., 2001). Nessa região foi relatado um
intenso ataque desse nematoide no cultivo da goiabeira que tomou maiores
proporções com o surgimento de novos plantios, muitos dos quais em áreas
anteriormente utilizadas com culturas suscetíveis como bananeira, tomateiro,
cebola e cucurbitáceas. A disseminação desse nematoide também foi favorecida
pelo uso comunitário de tratores e implementos contaminados com solo
infestado (MARANHÃO et al., 2001).
Pelo comprometimento do sistema radicular, os sintomas reflexos na parte
aérea das plantas ocasionados por patógenos desse gênero podem ser
confundidos com outros problemas como escassez de água ou déficit nutricional,
como amarelecimento e redução do crescimento das plantas.
O sítio de alimentação nematoides do gênero Meloidogyne no hospedeiro
é responsável pelo sucesso do parasitismo. O estabelecimento do sítio se dá
pela formação de três a cinco células gigantes multinucleadas, que funcionam
como drenos de nutrientes do cilindro vascular para o nematoide. As células
gigantes formadoras de galhas são geradas a partir de células vegetais
diferenciadas, situadas na porção anterior ao nematoide, induzidas pela
modificação do ciclo celular, balanço hormonal, sinalização celular, expressão
gênica e, consequentemente, a fisiologia da raiz (WILLIAMSON; GLEASON,
2003; BIRD et al., 2003). As galhas formadas ocasionam a ramificação anormal
das raízes, clorose e amarelecimento das folhas, porte aéreo irregular ou
reduzido, podendo ainda apresentar folhas finas, escassas e murchas (COYNE
et al., 2007; MICHEREF et al., 2005) (Figura 2).
Figura 2. Raízes com galhas (A) e parte aérea (B) de Citrullus lanatus var.
lanatus infectada por Meloidogyne enterolobii.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
26
As espécies de Meloidogyne são consideradas endoparasitas sedentários
e, seu ciclo de vida varia de 28 a 35 dias, dependendo da espécie hospedeira e
das condições ambientais, como a temperatura, tornando-se mais longo em
temperaturas mais baixas ou mais altas (AGRIOS, 2005).
Seguindo a embriogênese desse nematoide, a primeira muda, juvenil (J1)
ocorre dentro do ovo dando origem ao juvenil de segundo estágio (J2) (Figura
3). Quando J2 deixa as massas de ovos, eles infectam as raízes do hospedeiro
e migram pelo cilindro vascular, via força mecânica do estilete e degradação
enzimática da parede celular e lamela média, para formação do sítio de
alimentação (PERRY; MOENS, 2006; GHEYSEN; FENOLL, 2002).
Figura 3. Ciclo de vida do nematoide das galhas, Meloidogyne spp. A terceira
fase juvenil (J3) não está ilustrada na figura. J2 corresponde à segunda fase
juvenil e J4 à quarta fase (Fonte: PERRY & MOENS, 2006).
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
27
A natureza dos estímulos produzidos pelas raízes e percebidos por J2
não é clara, mas sabe-se que raízes de plantas suscetíveis são mais atraentes.
Muitos compostos orgânicos e inorgânicos excretados pelas raízes formam
gradientes da superfície da raiz no solo e podem influenciar os nematoides tanto
na atração como na repelência (PERRY; MOENS, 2006).
Nos estádios J3 e J4, os nematoides já se encontram no interior das
raízes, sendo sedentários e obesos. Após o estádio J4, ocorre à completa
formação do aparelho reprodutor, o que caracteriza o estádio adulto. Cada
fêmea pode produzir massas com aproximadamente 500 a 2.000 ovos
(TIHOHOD, 1993). Destes, apenas 5% sobrevivem para reproduzirem-se em
gerações seguidas, e terão em apenas quatro gerações, respectivamente: 25,
625, 15.625 e 390.625 adultos (CASTAGNONE-SERENO, 2002; TAYLOR;
SASSER, 1978). Em condições favoráveis, o ciclo de vida completo dos
nematoides das galhas ocorre em três a quatro semanas. Contudo, qualquer
espécie reduz ou paralisa por completo as suas atividades vitais em
temperaturas superiores a 40 °C ou inferiores a 5 °C (FERRAZ, 2001).
Nematoides do gênero Meloidogyne, por serem parasitas obrigatórios,
necessitam de plantas suscetíveis para a sua sobrevivência, que podem ser
plantas daninhas. Muitos ovos são mantidos viáveis em massas de ovos,
assegurando rápido crescimento da população quando plantas hospedeiras são
cultivadas (LIMA et al., 1995).
O controle desse nematoide é difícil devido a gama de hospedeiras
alternativas e não existem nematicidas registrados no Brasil para o controle
desse patógeno em melancia (AGROFIT, 2015). Além disso, os nematicidas tem
alto custo, apresentam período residual longo e alta toxidez ao homem e
ambiente. Para amenizar esse problema, a utilização de cultivares resistentes
para controlar este patógeno é a estratégia mais econômica e eficaz para o
produtor, impedindo ou dificultando o estabelecimento do patógeno na área de
cultivo (LIMA et al., 2005).
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
28
3.3. Recursos genéticos: fontes de resistência de Citrullus lanatus var.
citroides
Dentre os estados no Nordeste brasileiro, o Rio Grande do Norte é
considerado como centro secundário de diversidade da melancia forrageira na
agricultura tradicional, uma vez que são encontradas populações adaptadas à
região, e em alguns casos são encontrados tipos superiores que se
desenvolvem quase que totalmente sem agroquímicos (QUEIRÓZ, 1993;
QUEIRÓZ et al., 2004).
Diversos fatores ameaçam a perda desta diversidade genética, como a
substituição de genótipos tradicionais por melhorados e o êxodo rural
(QUEIRÓZ, 1993). Deste modo, foi realizada no final da década de 80
expedições no Nordeste brasileiro sendo coletadas várias amostras de sementes
da agricultura tradicional da região (QUEIRÓZ, 1993). Posteriormente, foram
realizadas etapas como multiplicação dessas sementes, caracterização,
avaliação e conservação, etapas que caracterizam um Banco Ativo de
Germoplasma (BAG) (GIACOMETTI, 1988).
Estes bancos são importantes fontes de variedades crioulas com
resistência que podem ser utilizadas em programas de melhoramento que visem
a resistência das plantas às doenças (BORÉM; MIRANDA, 2009).
Grande parte das cultivares e dos híbridos de melancia disponíveis no
mercado brasileiro não foram desenvolvidas para as condições climáticas do
Brasil, o que contribui para sua suscetibilidade aos principais estresses bióticos
do Semiárido nordestino.
Alguns estudos tem relatado fontes de resistência da melancia ao M.
enterolobii (THIES; LEVI, 2007). Pontes (2009) encontrou resistência a este
patógeno em acesso introduzido dos Estados Unidos, PI 244019, C. lanatus var.
citroides. Em outro trabalho, Castro et al. (2011), ao estudarem a reação de
genótipos de melancia a nematoides verificaram que acessos do BAG de
Cucurbitáceas da Embrapa Semiárido, BGCIA 240, BGCIA 229 e BGCIA TEMP
23, apresentaram os menores números de ovos, promovendo a maior inibição
na reprodução desses nematoides, sendo classificados como muitos resistentes.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
29
Damaceno (2012) verificou que há variabilidade para resistência de
alguns acessos, destacando a acesso BGCIA 240. Costa Filho (2012), ao avaliar
20 acessos de melancia coletados no estado do Rio Grande do Norte quanto à
reação ao M. enterolobii, verificou-se que dois destes acessos apresentaram
como fonte potencial de resistência.
3.4. Melhoramento genético de Citrullus lanatus L. visando resistência à
nematoides
Genes de resistência de plantas a nematoides podem estar presentes em
diversas espécies de culturas cultivadas e parentes selvagens. Sua identificação
e incorporação em cultivares comerciais são fatores importantes para um
programa de melhoramento visando a resistência às doenças (ABREU, 2005;
THURAU et al., 2010).
Esta resistência pode ser monogênica ou poligênica e sofrer influência da
variação de fatores ambientais, como a temperatura principalmente em zonas
tropicais. Neste contexto, foi identificado na cultura do tomate, o gene Mi-1, que
confere resistência contra nematoides das galhas, porém em temperaturas do
solo acima de 28 ºC, este gene é inativado (PERRY; MOENS, 2006).
Em geral, os nematoides provocam danos nas plantas resistentes e
suscetíveis, no entanto, há variação na intensidade dos danos. Na maior parte
dos casos, a resistência e a tolerância a nematoides são características
independentes em plantas. Isso significa que um está relacionado com o
controle da multiplicação do patógeno e outro com danos às plantas,
respectivamente (PERRY; MOENS, 2006).
Sabe-se que existem poucos genótipos de melancia disponíveis aos
produtores no mercado brasileiro. Entretanto, esses genótipos não foram
desenvolvidos para as condições brasileiras e sim de outros países como Japão
e Estados Unidos (COSTA; PINTO, 1977). Isto implica que o cultivo dessas
variedades e híbridos gera gastos com defensivos, uma vez que quando
cultivadas em condições ambientais para as quais não foram desenvolvidas, as
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
30
variedades tornam-se suscetíveis ao ataque de pragas e doenças (SILVEIRA et
al., 2005).
Para isso, é necessário reunir um maior número de informações
importantes sobre o germoplasma selecionado (LORENCETTI et al., 2005;
SOUZA et al., 2004).
No início de um programa de melhoramento genético visando à
incorporação de resistência a fitopatógenos, é necessário conhecer a reação de
acessos da cultura estudada frente a estes agentes para a seleção de fontes de
resistência (WILCKEN et al., 2005). Sendo assim, para que haja sucesso em um
programa de melhoramento genético é fundamental a escolha de potenciais
genitores. Isso permitirá que se obtenham híbridos e, posteriormente,
populações segregantes promissoras.
3.5. Mecanismos de defesa de plantas à nematoides
As plantas são constantemente atacadas por pragas e por patógenos
causando sérios prejuízos aos produtores. Dentre estes patógenos, os
nematoides se destacam e podem parasitar milhares de espécies de plantas.
Entretanto, tem sido relatadas significativas exceções nesse processo de
parasitismo. Assim, com o processo evolutivo, essas plantas desenvolveram
diversos mecanismos de defesa (AGRIOS, 2005) que podem prevenir a atração,
penetração, migração, formação do sítio de alimentação, nutrição, reprodução
ou sobrevivência dos nematoides, podendo ser classificadas em plantas
antagonistas, armadilhas, não hospedeiras ou resistentes (FRAGOSO et al.,
2007).
Nas plantas, os mecanismos de defesa podem ser estruturais e
bioquímicos, ambos podendo ser pré-existentes ou constitutivos (passivos), ou
seja, quando sua expressão independe da presença do patógeno (HUANG,
1985). Nesta categoria, incluem-se os efeitos repelentes e/ou nematicida de
determinadas substâncias químicas presentes na parte aérea ou no exsudato
radicular de algumas plantas. Segundo Rodríguez-Kábana et al. (1994), algumas
plantas contem na parte aérea, compostos nematicidas pré-formados, como
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
31
alcalóides, ácidos graxos, isotiocianatos, glicosídeos acianogênicos, terpenóides
e compostos fenólicos que apresentam toxidez aos nematoides e também
podem ser utilizadas como adubo verde. Em plantas do gênero Tagetes spp. é
produzida a substância química α-tertienil no exsudato radicular, a qual
apresenta efeito nematicida (MARAHATTA et al., 2012).
Algumas
plantas
também
apresentam
mecanismos
estruturais
e
bioquímicos pós-formados (induzidos) que são ativados ou aumentam o nível de
compostos pré-existentes após a infecção pelo patógeno (MICHEREFF et al.,
2005). Por exemplo, em plantas resistentes, após o reconhecimento do
patógeno, há o aumento dos níveis de defesa da planta pela ativação de genes
que codificam diversos compostos em resposta a fatores externos bióticos e/ou
abióticos (MICHEREFF et al., 2005). Estas respostas de defesa bioquímicas
podem estar associadas às espécies reativas de oxigênio (EAO´s), proteínas
relacionadas à patogênese e fitoalexinas, dentre outros (AGRIOS, 2005).
3.5.1. Espécies ativas de oxigênio e enzimas antioxidantes
Após o reconhecimento do patógeno pela planta, a primeira resposta de
defesa ativada é a explosão oxidativa ou produção de EAO´s (ex: H2O2, O2- e
OH-) (RESENDE et al., 2003). As EAO’s ocorrem normalmente na célula vegetal,
mas quando estas se acumulam podem se tornar tóxicas à própria célula da
planta. A resposta hipersensitiva é um mecanismo bioquímico pós-formado de
defesa, caracterizado pela morte de células do hospedeiro próximas aos
patógenos biotróficos, o que resulta na interrupção da infecção e na
consequente resistência da planta à doença pelo acúmulo de EAO´s. A indução
da resposta de hipersensibilidade tem sido associada à interação molecular
específica entre uma proteína de avirulência do patógeno, que sinaliza sua
presença para a planta com uma proteína de resistência, que faz a ligação de
reconhecimento proteína-proteína e desencadeia a resposta hipersensitiva
(BAKER et al., 1997; PARKER et al., 2000), conhecida como hipótese de
resistência gene-a-gene.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
32
No entanto, para evitar um maior dano nos tecidos do hospedeiro, além
do ponto de infecção dos patógenos, as plantas possuem um conjunto de
enzimas antioxidantes, tais como guaiacol peroxidase, ascorbato peroxidase,
superóxido dismutase e catalase, que impedem a ação tóxica das EAOs
protegendo as células da planta (RESENDE; SALGADO; CHAVES, 2003).
3.5.1.1. Peroxidases
As peroxidases são glicoproteínas antioxidantes, que catalisam a
oxidação do substrato concomitantemente à redução do peróxido de hidrogênio
(CHOI et al., 2007). Elas estão presentes em fungos, bactérias, plantas e
animais (HIRAGA et al., 2001). Nas plantas, as peroxidases são subdivididas em
três classes. A classe I é representada por enzimas intracelulares, como a
citocromo c peroxidase; vale salientar que, cada enzima recebe uma abreviação
do nome e classificação numérica baseado nas reações químicas que catalisam,
assim o código EC (Enzyme commission) e abreviação da enzima acima
descrita é E.C 1.11.1.5, CCP, repectivamente; e a ascorbato peroxidase (E.C
1.11.1.11, APX). A classe II é composta por enzimas extracelulares, como a
lignina peroxidase (E.C 1.11.1.14, LiP) e peroxidase dependente de manganês
(E.C 1.11.1.13, MnP). A classe III é formada por peroxidases (E.C 1.11.17, POX),
que são secretadas no apoplasto ou acumuladas no vacúolo, apresentam uma
ampla variedade de substratos e são responsáveis pela maioria das reações de
oxirredução estimuladas pelas variações ambientais, ferimentos ou reações
infecciosas (HIRAGA et al., 2001).
Na reação da atividade do guaiacol peroxidase (1), o guaiacol reage com
o peróxido de hidrogênio na presença da peroxidase formando o tetraguaiacol
(HIRAGA et al., 2001). Nesta reação, o peróxido de hidrogênio é reduzido e o
guaiacol, oxidado, atua como doador de prótons.
Peroxidase
4 Guaiacol + 4 H2O2
Tetraguaiacol (alaranjado) + 8 H2O
(1)
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
33
Em um estudo sobre interação de nematoide das galhas com plantas de
tomate resistentes e infectadas pelo fungo Fusarium oxysporum f.sp.
Lycopersici, foi mostrado que a atividade específica de POX aumentou em
plantas inoculadas com o fungo até o quinto dia após a infecção. Enquanto nas
plantas inoculadas com nematoides, a atividade da enzima aumentou apenas
até ao segundo dia e, em seguida, diminuiu. Atividade da enzima POX foi
significativamente menor nos tratamentos (nematoide + fungos), em comparação
com as plantas de controle inoculados com o fungo. Foi observado também, que
em todas as fases biológicas do nematoide há uma supressão da atividade de
peroxidase em raízes de tomateiro, mas esta é acentuada na formação de
adultos do que nas outras fases da vida do nematoide, incluindo formação de
células gigantes, postura de ovos e penetração (SAHEBANI et al., 2008).
3.5.1.2. Ascorbato Peroxidase
A enzima ascorbato peroxidase (E.C 1.11.1.11, APX) engloba uma família
de isoenzimas importantes no sistema de detoxificação de espécies reativas de
oxigênio nas plantas, catalisando a conversão do peróxido de hidrogênio
(H2O2) em água (GUZZO; HARAKAVA, 2007).
Esta enzima constitui parte do ciclo conhecido como ascorbato/glutationa.
Esta via faz uso de antioxidantes não enzimáticos doadores de elétrons como o
ascorbato e a glutationa. Estes oxidantes são responsáveis pela remoção de
metabólitos de peróxido onde a enzima catalase não consegue alcançar, como
por exemplo, no cloroplasto (POLLE, 2001).
Na reação (2), observa-se que o H2O2 pode ser reduzido e removido pela
APX
através
do
ascorbato
como
redutor,
formando
o
radical
monodehidroascorbato (MDA) (YOSHIMURA et al., 2000).
APX + H2O2
Composto I + H2O
Composto I + Ascorbato
Composto II + MDA
Composto II + Ascorbato
APX + MDA. + H2O
(2)
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
34
Após o reconhecimento da planta ao patógeno, ocorre a sinalização de
respostas de defesa, onde o H2O2 age como um sinalizador. A partir desse
momento, sucede-se explosão oxidativa e indução de genes de proteção celular
nas células vizinhas restringindo o desenvolvimento da lesão em função da
resposta de hipersensibilidade (HR), que é, então, acompanhada pelo
desenvolvimento de uma resistência sistêmica adquirida (SAR) (RESENDE et
al., 2003).
Em raízes de soja inoculadas com nematoides das galhas (Meloidogyne
javanica e Meloidogyne incognita) foi expresso, após 12 horas da inoculação, um
aumento desta enzima nas plantas inoculadas com o patógeno quando
comparadas com plantas controles não inoculadas (SANTANA, 2012). Logo, a
APX está envolvida nos mecanismos de defesa vegetal contra os danos
provocados pelo estresse oxidativo.
3.5.1.3. Catalase
As catalases (E.C 1.11.1.6, CAT) são metaloproteína constituídas por
quatro grupos heme que convertem H2O2 em H2O e O2 (3), sendo encontradas
em organismos aeróbicos (SCANDALIOS, 2005). Nas células vegetais, estão
presentes nos peroxissomos, glioxissomas, mitocôndrias e dispersos no citosol
(RESENDE et al., 2003). Esta enzima decompõe peróxido de hidrogênio a um
ritmo extremamente rápido, correspondente uma atividade de centro catalítico de
cerca de 107 min-1, apresentando baixa constante de Michaelis (Km) que é a
concentração de substrato necessária para obter a metade da velocidade
máxima.
Catalase
H2O2
H2O +
O2
(3)
A CAT está relacionada com a expressão de genes em plantas, com o
aumento da resistência das mesmas durante estresses abióticos e estresse
biótico. Para este último, foi observada a atividade dessa enzima em diferentes
interações planta-patógeno. Em plantas de tomate inoculadas com Meloidogyne
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
35
spp. houve maior atividade de CAT, 50 dias após a inoculação, em relação às
plantas não inoculadas (MOLINARI; MIACOLA, 1997). E ao estudar sobre
resistência sistêmica adquirida, foi relatada que a infecção de plantas de algodão
pelo nematoide Rotylenchulus reniformis aumenta a atividade da catalase tanto
nas cultivares suscetível (B2RF), quanto nas resistentes (LONREN – 1), o
mesmo não ocorre com o controle, o qual foi isento do estresse causado pelo
patógeno (ARYAL et al, 2011).
3.5.2. Polifenoloxidase
As polifenoloxidases (E.C 1.10.3.1, PFOs) são enzimas que reagem (4)
junto com a agente oxidante O2, inserindo um átomo de oxigênio em um grupo
hidroxila existente nos compostos fenólicos, seguida pela oxidação do o-difenol
na quinona correspondente (MAYER et al., 2006).
O2 + Catecol (o-difenol)
o-quinona + H2O2
(4)
Desta forma, o papel fisiológico da polifenoloxidase (PPO) em plantas tem
sido relacionado à resistência ao ataque de insetos e microorganismos. Ao
passo que, o dano causado por esse ataque rompe as células dos cloroplastos,
e a enzima que se encontrava latente, torna-se ativa quando liberada na
membrana dos tilacóides e assim ocorre a reação de oxidação, que tem como
resultado a produção de quinonas, compostos que são bastante reativos
(YORUK; MARSHALL, 2003). Além disso, as quinonas são mais tóxicas a
microrganismos do que os fenóis em sua forma original (AGRIOS, 2005).
Não se conhece nenhum trabalho que relacione a PPO em melancia
como forma de defesa a nematoides. No entanto, alguns estudos tem mostrado,
para plântulas de soja e inoculadas com J2 de M. javanica, um aumento na
atividade da PPO após 72 h da inoculação na variedade Hartwig (resistente) em
comparação com raízes de plantas não infectadas e com raízes de Cristalina
(suscetível). No caso da soja inoculada com J2 de Heterodera glycines, houve
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
36
aumento na atividade de PPO na variedade resistente às 48 h após a
inoculação. Acredita-se que a atividade provocada por H. glycines foi detectada
mais precocemente pela forma de infecção e estabelecimento na raiz, uma vez
que este nematoide provoca mais lesões no tecido da raiz que M. javanica
(SHIMIZU, 2004).
3.5.3. Fenilalanina amônia-liase
A fenilalanina amônia-liase (E.C 4.3.1.5, PAL) é uma enzima chave do
metabolismo vegetal estando envolvida na biossíntesse dos fenilpropanoides,
com participação de fenilalanina catalisando a reação de conversão de Lfenilalanina em amônia e ácido trans-cinâmico, além de ser o ponto de
ramificação entre o metabolismo primário (via do ácido chiquímico) e secundário
(HERRMANN; WEAVER, 1999).
A PAL é uma enzima tetramérica composta por quatro subunidades
diferentes e está presente nas plantas, localizada no citoplasma das células,
mas também pode estar associada a estruturas membranosas (DIXON; PAIVA,
1995). O papel da PAL como um fator de resistência de plantas a patógenos é
bem conhecido, assim como o papel desta em plantas infectadas com
nematoides das galhas. Muitos estudos a este respeito foram descritos em
algumas plantas da família da Fabaceae e Solanaceae, o mesmo não é
compreendido entre as Cucubitáceas.
Janegitz (2012), ao estudar a indução de compostos de defesa pela
aplicação de cis-jasmone e danos causados por M. javanica em genótipos de
soja resistente e suscetível, observou que a pulverização foliar com cis-jasmone
induziu sistemicamente a atividade da PAL, após 120h da aplicação. Já, a
pulverização foliar com cis-jasmone e inoculação aumentaram a atividade da
PAL, 72h após a inoculação. Estes resultados sugerem que a indução de forma
sistêmica, nas raízes de soja, por cis-jasmone assim como em resposta ao
parasita, podem estar relacionados à resistência da soja à M. javanica. Em
avaliação de genótipos de tomate resistentes e suscetíveis a M. incógnita,
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
37
observou-se que os genótipos resistentes possuíam atividade de PAL superior
quando comparado a genótipos suscetíveis (KALAIARASAN, 2009).
Diversos trabalhos demonstraram a indução de fitoalexinas por plantas
em defesa ao ataque de nematoides do gênero Meloidogyne. Em raízes de
plantas de soja resistente e inoculada com M. javanica foi observada a produção
da fitoalexina coumestrol, induzidas pela via do ácido jasmônico (JANEGITZ et
al., 2012). Outra substância química conhecida pelo seu papel na defesa das
plantas e encontradas na soja é a fitoalexina gliceolina, que pode ser produzida
em resposta ao ataque por M. incognita (KAPLAN et al., 1980).
Na família das Solanáceas, o tomate e a batata tem mostrado associação
da atividade da enzima PAL como um fator de resistência a patógenos (CAMM;
TOWERS, 1973; BRUESKE, 1980; KALAIARASAN, 2009). Nesse sentido, foi
observado em raízes de tomate resistentes e inoculadas com M. incognita menor
número de galhas por planta e um aumento significativo na atividade da PAL às
108 horas após a inoculação, superiores às plantas suscetíveis inoculadas com
este nematoide (BRUESKE, 1980, KALAIARASAN, 2009). Em raízes de batatas
resistentes e inoculadas com o nematoide dos cistos, Globodera rostochiensis,
foram encontradas maiores atividades da PAL e tirosina amônia-liase (TAL) do
que em raízes de plantas suscetíveis (GIEBEL, 1973).
Plantas resistentes expostas às proteínas dos patógenos ativam seus
mecanismos de defesa, tais como a enzima PAL e a atividade das enzimas
antioxidantes, catalase e peroxidase (RYAN; JAGENDORF, 1995; MARIUTTO et
al., 2011; TRIPATHI, 2006).
3.6. Proteínas relacionadas à patogênese
As PR-proteínas são proteínas localizadas intra e extracelularmente que
se acumulam em tecidos vegetais ou em cultura de células após o tratamento
com eliciadores ou sob ataque de patógenos. Estas proteínas apresentam
algumas características físico-químicas e imunológicas comuns que permitiram
que elas fossem classificadas em 17 famílias, variando de PR-1 até PR-17.
Destas, as mais estudadas são as peroxidases, quitinases e β-1,3-glucanases
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
38
(VAN LOON et al., 2006). A PR- 9 é um tipo específico de peroxidase que atua
no reforço da parede celular da planta, catalisando a lignificação e aumentando a
resistência contra vários patógenos (PASSARDI et al., 2004).
Assim, as PR-Proteínas podem estar presentes na planta em baixos
níveis, e seus níveis são aumentados quando as plantas são submetidas a
condições de estresses. Nessa condição, os genes correspondentes são
ativados, vindo a ser detectadas nos tecidos vegetais após injúrias de origem
biótica (pragas e doenças) e/ou abiótica (salinidade, seca, baixas/altas
temperaturas, etc) (BERNARDS et al., 1999; MARTINS-MIRANDA, 2002).
Muitas PR-Proteínas possuem atividades antifúngica, antibacteriana in
vitro e antinematicida como, por exemplo, quitinases, glucanases e proteínas
que se ligam à quitina. Pesquisas avaliando cultivares resistentes de algodão e
café infectadas com M. incognita e M. paranaensis, respectivamente,
identificaram proteínas relacionadas com a patogenicidade como quitinase e
quinona redutase (FRANCO et al., 2010). O reconhecimento da presença dos
nematoides resulta em mudanças conformacionais das proteínas que sinalizam
a resposta de defesa. No entanto, não se sabe ao certo se as proteínas que
reconhecem a presença do patógeno o fazem por meio do reconhecimento
direto do produto de uma molécula do nematoide ou pelas mudanças na célula
hospedeira mediada pelos genes do patógeno (BELKHADIR et al., 2004).
3.7. Clorofila
A clorofila está diretamente relacionada com a atividade fotossintética nas
plantas (TAIZ & ZEIGER, 2006). Assim, a sua quantificação é relevante no
estudo de fatores externos que interferem na qualidade e na quantidade da
clorofila no tecido foliar (LARCHER, 1986).
Diversas formas podem ser trabalhadas para se obter dados de clorofila
nas folhas, estas podem ser diretas (método destrutivo ou clássico) ou indiretas
(método não destrutivo). Alguns estudos tem mostrado a correlação positiva
entre os dois métodos. De tal forma, Rigon et al. (2012) conduziram um
experimento para estabelecer a relação entre os pigmentos fotossintéticos
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
39
extraídos em dimetilsulfóxido (DMSO) e as leituras obtidas no clorofilômetro
portátil ClorofiLOG® 1030. As leituras foram realizadas em discos foliares com
diferentes tonalidades de verde, sendo feita, nesses mesmos discos, a
determinação da clorofila pelo método clássico. Os resultados indicaram que o
clorofilômetro portátil ClorofiLOG® 1030, associado a modelos matemáticos,
permitiu estimar a concentração dos pigmentos fotossintéticos, exceto a clorofila
b, com alta precisão, com economia de tempo e com reagentes normalmente
utilizados nos procedimentos convencionais.
Alguns estudos demonstram a importância da relação da clorofila com as
doenças de plantas. Na avaliação de cultivares de soja quanto à resistência
parcial à P. pachyrhizi e o teor de clorofila com o auxílio de um clorofilômetro,
Polizel et. al. (2011) observaram que o teor de clorofila apresentou uma relação
inversamente proporcional à severidade da ferrugem asiática. Em outro trabalho,
Lubarino (2012) ao realizar avaliações aos 60 dias após a inoculação com M.
enterolobii em genótipos de melancia do BAG de Cucurbitacéas da Embrapa
Semiárido observou que os índices de clorofila a e b foram menores nas plantas
inoculadas quando comparadas com seus controles não inoculados. Este estudo
mostrou que embora existam genótipos com maior ou menor presença de galhas
nas raízes, nas duas situações há uma diminuição da clorofila presente na folha.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
40
4. PARTE EXPERIMENTAL
4.1. Material vegetal e cultivo das plantas
Os experimentos foram realizados na casa de vegetação da Embrapa
Semiárido, entre os meses de junho a agosto, de 2013 e 2014. No primeiro
experimento foi utilizada uma população de plantas do acesso BGCIA 240 de
melancia forrageira (C. lanatus var. citroides) proveniente do Banco Ativo de
Germoplasma (BAG) de Cucurbitáceas da Embrapa Semiárido previamente
selecionada pelo programa de melhoramento visando resistência a patógenos
de solo desta Unidade. No segundo experimento foram utilizadas as plantas do
acesso BGCIA 240 classificadas no experimento anterior como resistente a M.
enterolobii, mais as plantas de um acesso BGCIA 875 de melancia de mesa (C.
lanatus var. lanatus), suscetível ao patógeno. Para a produção das mudas, as
sementes foram plantadas em copos descartáveis de polipropileno (uma
semente por copo) contendo 500 mL de solo autoclavado, cujos resultados das
análises química e física realizadas estão apresentadas na Tabela 1.
Tabela 1. Caracterização física e química do solo autoclavado utilizado nos
experimentos. Embrapa Semiárido, Petrolina-PE, 2015.
Análise Física do Solo
Areia
Silte
Porosidade Total
47,5
M.O
PO4 ++
kg-1
dm-3
g
-
mg
5
Argila
g kg-1
(%)
K+
759,9
177,6
Análise Química do Solo
Ca2+
Mg2+
Na+
Al+ H
--------------cmolcdm-3
0,1
0,04
1,6
62,6
pH
mS cm -1
-------------
1,1
CE
1
6,2
0,54
4.2. Preparo do inóculo e inoculação
Para a produção e manutenção do inóculo, foram coletadas fêmeas de
uma população de M. enterolobii de raízes de goiabeira (Psidium guajava L. cv.
Paluma) no município de Petrolina, PE, que foram previamente identificadas
utilizando-se o padrão de isoenzimas (CARNEIRO et al., 2001) e multiplicado em
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
41
mudas de tomateiro cv. Santa Clara. Para o preparo do inóculo, os ovos foram
extraídos das plantas de tomateiro infectadas de acordo com a metodologia
proposta por Hussey e Barker (1973). A inoculação foi realizada 12 dias após a
germinação das sementes, quando surgi a primeira folha expandida, com
suspensão de 500 ovos de M. enterolobii por planta, utilizando-se 1 mL da
suspensão, sendo depositados 500 µL de cada lado do colo das plantas.
4.3. Seleção de plantas resistentes a M. enterolobii na população do acesso
BGCIA 240
Afim de se visualizar os sintomas iniciais da infecção causado pelo M.
enterolobii em plantas de melancia, foi realizado aos 30 dias após a inoculação,
lavagem das raízes (Figura 4) e avaliações individuais nas plantas pelos
métodos não destrutivos, como tais variáveis:
Figura 4. Etapas da lavagem de raízes inoculadas da Acesso BGCIA 240, para
avaliação aos 30 dias após inoculação. Retirada da planta (A), lavagem e
retirada do excesso de solo nas raízes (B).
A) Desenvolvimento global da parte aérea (DGPA): comprimento da parte
aérea (CPA), diâmetro do hipocótilo (DH), comprimento (CF) e diâmetro da
primeira folha verdadeira (DF).
B) Grau de resistência da planta estimado a partir do desenvolvimento global
de galhas (DGG), obtido através de uma escala de nota, a qual variou de 0 a 5,
em função do número de galhas onde 0 = ausência de galhas a 1 = 1 a 2 galhas
(Resistente, R); 2 = 3 a 10 galhas a 3 = 11 a 30 (Média Resistência, MR); e 4 =
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
42
31 a 100 galhas a 5 = mais de 100 galhas (Suscetível, S), adaptada conforme
proposta de Taylor & Sasser (1998).
4.4. Avaliação dos métodos não destrutivos com os métodos tradicionais
destrutivos
Após a seleção nas avaliações anteriores baseado no DGPA e DGG, as
plantas classificadas preliminarmente como R, MR e S, foram transplantadas
para vasos de 30 litros de solo, submetidas ao manejo recomendado para
melancia (DIAS et al., 2010). Afim de se observar o máximo da infecção causada
pelo M. enterolobii nas plantas inoculadas do acesso BGCIA 240, foram
realizadas avaliações ao final do ciclo (FC), aos 90 DAI (Figura 5).
Figura 5. Aspecto geral das plantas em vasos de 30 litros de solo para obtenção
de descendência em condições de telado.
As avaliações foram feitas pelos métodos não destrutivos, como tais
variáveis:
A) Desenvolvimento global da parte aérea (DGPA): (exceto CF e DF, e
adicionou-se os dados de clorofilas A e B, na região apical, mediana e basal da
planta a partir do clorofilômetro marca ClorofiLOG® modelo CFL 1030).
B) Desenvolvimento global de galhas (DGG): conforme Taylor & Sasser
(1998).
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
43
E pelos métodos destrutivos, que são as avaliações do sistema radicular
realizadas após eliminação da parte aérea baseadas no:
A) Desenvolvimento global do sistema radicular (DGSR): comprimento do
sistema radicular (CSR), massa fresca do sistema radicular (MFSR) e volume do
sistema radicular (VSR).
B) Fator de reprodução (FR): através do número de ovos (NO) é definido a
razão entre a população final (que foi recuperada no sistema radicular) e a inicial
(utilizada na inoculação). Plantas com FR < 1 são consideradas resistentes,
enquanto que aquelas com FR ≥ 1 são consideradas suscetíveis (MOURA;
REGIS, 1987).
4.5. Reação de genótipos de melancia de mesa e forrageira ao patógeno
Para a avaliação da reação dos genótipos de melancia de mesa suscetível
(BGCIA 875) e de forrageira resistente (BGCIA 240) ao nematoide, foram
realizadas avaliações aos 7, 14, 21, 28 e 35 dias após a inoculação, do número
de galhas (NG) e número de ovos por planta (NO) (COOLEN; D’HERDE, 1972).
Além disso, avaliou-se também o comprimento do sistema radicular (CSR) e
massa fresca do sistema radicular (MFSR). Para a realização das análises, as
raízes das plantas foram retiradas dos recipientes e lavadas com água.
4.6. Análises de clorofila e enzimáticas
Para a avaliação das clorofilas e enzimática, foi instalado outro
experimento paralelo ao do item 4.5.
Antes das coletas das raízes, foram realizadas as avaliações das clorofilas
a e b (IFC - Índice Falker de Clorofila) das folhas utilizando-se o clorofilômetro
marca ClorofiLOG® modelo CFL 1030. Para tanto, foram realizadas três leituras
com o clorofilômetro, sempre na primeira folha completamente expandida (do
topo do dossel para a base) aos 7, 14, 21, 28 e 35 dias após a inoculação (DAI).
Para as análises enzimáticas, foram realizadas coletas nos mesmos
tempos citados anteriormente, onde as raízes dos genótipos foram lavadas
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
44
cuidadosamente com água corrente. Posteriormente, as amostras radiculares
foram envolvidas em papel alumínio, identificadas, mergulhadas em nitrogênio
líquido e armazenadas em freezer (-80 °C) até o momento das análises. Para a
obtenção do extrato protéico, tecidos radiculares foram triturados em nitrogênio
líquido, com almofariz e pistilo, até a obtenção de um pó fino. Em seguida, foi
adicionado tampão fosfato de potássio 100 mM pH 7,0 (3 mL de tampão por
grama de material triturado) e homogeneizou-se com o pistilo por 2 min. Após
esse processo, a suspensão foi centrifugada a 12.000 g por 15minutos (4 oC) e o
sobrenadante foi utilizado como fonte enzimática.
As proteínas solúveis totais contidas nos extratos foram quantificadas com
base no ensaio de Bradford (1976) utilizando-se uma curva padrão de albumina
sérica bovina (BSA).
A atividade do guaiacol peroxidase (POX) foi determinada pelo aumento na
absorbância a 470 nm durante 2 min em um meio de reação contendo fosfato de
potássio 100 mM (pH 7,0), guaiacol 50 Mm e H2O2 150 mM (URBANEK et al.,
1991).
A atividade da catalase (CAT) foi estimada pelo decréscimo na
absorbância a 240 nm durante 3 min em um meio de reação contendo fosfato de
potássio 100 mM (pH 7,0) e H2O2 250 mM (AZEVEDO et al.,1998).
A atividade do ascorbato peroxidase (APX) foi medida pelo decréscimo na
absorbância a 290 nm durante 3 min em um meio de reação contendo fosfato de
potássio 100 mM (pH 7,0), ácido ascórbico 10 Mm e H2O2 2 mM (NAKANO;
ASADA, 1981).
A atividade de polifenoloxidase (PPO) foi estimada pelo aumento na
absorbância a 420 nm durante 3 min em um meio de reação contendo fosfato de
potássio 100 mM (pH 6,5) e pirogalol 100 mM (GAUILLARD et al., 1993). A
reação ocorreu pela transformação do pirogalol (fenol) em quinona.
A atividade da enzima fenilalanina amônia liase (PAL) foi ensaiada através
da reação entre 90 μL de tampão 100 mM Tris-HCl (pH 8,8), 50 μL de 40 mM de
fenilalanina e 50 μL do extrato enzimático (MORI et al.,2002).
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
45
4.7. Delineamento experimental e análise estatística
No experimento de seleção de plantas resistentes e validação dos
métodos de avaliação não destrutivo foi utilizado uma população de 37 plantas.
O delineamento utilizado foi o inteiramente casualizado, onde cada planta
correspondeu a uma unidade experimental. O estudo da correlação entre as
variáveis estudadas foi baseado no método do coeficiente de correlação de
Pearson, através do software GENES (CRUZ, 2006).
No experimento de reação dos genótipos ao patógeno e análises
enzimáticas e de clorofila, o delineamento utilizado foi o inteiramente
casualizado, com três repetições e parcela experimental composta por três
plantas. Os dados foram submetidos à análise de variância utilizando-se o
programa Sisvar (FERREIRA, 2006). As médias do número de galhas e número
de ovos por planta foram comparadas pelo teste T a 5% de probabilidade. As
médias do comprimento, massa fresca de raiz e clorofilas a e b foram
comparadas pelo teste de Tukey 5% de probabilidade. Para melhor visualização
dos resultados das análises enzimáticas ao longo do tempo, plotou-se a curva
de progresso da atividade das enzimas com os respectivos erros padrões.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
46
5. RESULTADOS E DISCUSSÃO
5.1. Seleção de plantas resistentes a M. enterolobii na população do acesso
BGCIA 240
Na Tabela 2, é observada uma semelhança nos resultados referentes à
resistência das plantas com relação à avaliação preliminar do grau de resistência
das plantas e o fator de reprodução (FR) de M. enterolobii. Assim, verificou-se
que das 37 plantas do acesso BGCIA 240 que germinaram, 17 foram
classificadas preliminarmente como R, correspondendo a 46 %, 32 % do
estande foram MR e 22 % das plantas foram classificadas como S. Segundo
Oliveira et al. (2000), a melancia forrageira apresenta um período de dormência
fisiológica, de aproximadamente dois meses e atingem o máximo de germinação
entre 90 e 100 dias após a colheita, explicando assim a dificuldade em se obter
um maior estande de plantas de melancia forrageira para as avaliações, já que a
qualidade de germinação varia entre os genótipos de C. lanatus var. citroides.
Segundo Almeida et al. (2011) e Coyne et al. (2007) à medida que as
galhas se formam, há uma menor disponibilidade de nutrientes para a planta,
interferindo no desenvolvimento da parte aérea. No presente trabalho, o alto
percentual de acerto (78,33 %) na caracterização precoce da reação ao
nematoide (R, MR e S) (Figura 6), no qual se considerou o DGPA e DGG está de
acordo com o reportado pelos citados autores.
Por meio dos dados de amplitude do fator de reprodução (FR), verificouse uma variação de 0,04 – 1,62 para plantas R (Tabela 2), correspondendo a
94,12 % das plantas avaliadas preliminarmente como R, enquanto que se
observou a amplitude de 0,05 – 2,78 para as plantas MR, e de 0,23 – 2,03 para
as S. Isto indica uma porcentagem de acerto (%), validada pelo FR < 1, de 91,66
% das plantas MR. Enquanto isso, a porcentagem de acerto correspondente as
planta avaliadas como S, foi de 87,5 %. Este resultado, provavelmente, foi
influenciado pelo menor número de plantas classificadas como S (8 plantas),
ocasionando uma margem de erro maior. Contudo, a média de porcentagem de
acerto foi de 91,09 %, indicando a efetividade dos parâmetros considerados na
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
47
avaliação preliminar como ferramenta alternativa e não destrutiva na
discriminação precoce do grau de resistência de genótipos a M. enterolobii.
Tabela 2. Avaliação preliminar dos graus de resistência e suscetibilidade em
melancia forrageira acesso BGCIA 240, inoculadas com Meloidogyne enterolobii
aos 30 dias após inoculação (DAI) e dados de amplitude do fator de reprodução
(FR), avaliado aos 90 DAI. Petrolina, Embrapa Semiárido, 2015.
Época de avaliação
Amplitude do Fator de
reprodução (FR) *
Grau de resistência
Resistente
Média resistência
Suscetível
Total
Média
Média FR
30 DAI
Mínima
Máxima
17
12
8
37
0,04
0,05
0,23
1,62
2,78
2,03
0,62
0,87
1,24
0,11
2,14
0,91
*. FR < 1: Plantas resistentes são consideradas aquelas com número de ovos menor que 500 por
sistema radicular e FR ≥ 1: Plantas suscetíveis são as plantas que apresentaram número de
ovos maior e igual que 500 por sistema radicular.
Figura 6. Classes do sistema radiculares do acesso BGCIA 240, colhido e
avaliado. Resistente (A), média resistência (B) e suscetível (C) em função do
desenvolvimento global da parte aérea (DGPA) e de galhas (DGG).
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
48
De acordo com dados de amplitude do acesso BGCIA 240 (Tabela 3)
observou-se que a avaliação preliminar para plantas designadas como
resistentes (R), média resistência (MR) e suscetíves (S) apresentaram médias
do número de ovos/mL dentro do resultado esperado, < 500 ovos encontrados
para plantas classificadas como R e MR, bem como > 500 ovos para plantas S.
De acordo com os dados de amplitude, o número de ovos/mL variou de 20 a 808
para plantas R (observando-se que das 17 plantas avaliadas como R, somente
uma planta apresentou o número de ovos > 500). Este dado corresponde ao
encontrado por Castro et al. (2011), estudando a reação de genótipos de
melancia a M. enterolobii, verificaram que o acesso BGCIA 240 teve o menor
número de ovos (NO) em relação aos demais genótipos estudados, com apenas
45 ovos por sistema radicular. Estas progênies promoveram a maior inibição na
reprodução dos nematoides sendo classificados como muito resistente. Para
plantas avaliadas como S, somente uma, das oito plantas classificadas
inicialmente, apresentou NO < 500. Portanto, a avaliação preliminar baseada no
desenvolvimento global da parte aérea e de galhas, mostrou-se confiável, tendo
em vista à necessidade de seleção de caraterísticas não destrutivas,
possibilitando assim, o transplantio de mudas inoculadas com M. enterolobii com
potencial para seguir nos trabalhos de melhoramento de melancia visando
resistência ao referido patógeno do solo.
Tabela 3. Dados de amplitude do acesso BGCIA 240 inoculadas com
Meloidogyne enterolobii avaliadas quanto ao número de ovos/mL de solução do
sistema radicular. Petrolina, Embrapa Semiárido, 2015.
Avaliação Preliminar
Amplitude número de ovos
Mínima
Máxima
Média
Resistente
20
808*
310
Média resistência
25
1391
436
Suscetível
113
1017**
622
*. Somente uma planta apresentou valor acima de 500 ovos.
**. Somente uma planta apresentou valor abaixo de 500 ovos.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
49
5.2. Correlação dos métodos não destrutivos com os métodos tradicionais
destrutivos
Em relação às correlações entre os caracteres, observou-se que das 30
combinações possíveis entre os quatro caracteres e os três grupos de plantas
classificadas quanto a diferentes graus de resistência (R, MR e S) foram
altamente significativas 11 combinações e três significativas a 5 % (Tabela 4).
Ao analisar o desenvolvimento global da parte aérea (CPA, DH, CF e DF)
das plantas R verificou-se que não houve correlações destas variáveis com o
desenvolvimento global de galhas (DGG), devido a variável constante. No
entanto, foi observado nas plantas S uma correlação negativa do compriemento
de folha (CF) e o DGG de -0,97.
Ao avaliar as demais correlações de desenvolvimento global da parte
aérea (DGPA) das plantas R, observou-se correlações significativamente
positiva entre elas.
Tabela 4. Valores dos coeficientes de correlação para a associação entre as
variáveis desenvolvimento global de galhas (DGG) e da parte aérea (DGPA) de
planta do acesso BGCIA 240 aos 30 dias após a inoculação. Petrolina, Embrapa
Semiárido, 2015.
DGG
CPA
DH
CF
R
MR
S
R
MR
S
R
MR
S
R
MR
S
CPA
1,00**b
0,85 ns
-0,82 ns b
DH
1,00**b
0,35 ns
-0,93 ns b
1,00**
0,26 ns
0,65 ns
CF
1,00**b
-0,31 ns
-0,97* b
1,00**
-0,41 ns
0,66 ns
1,00**
-0,94 ns
0,98*
DF
1,00**b
-0,03 ns
-0,91 ns b
1,00**
-0,74 ns
0,57 ns
1,00**
-0,94 ns
0,99**
1,00**
0,93 ns
0,98*
*. A correlação é significativa no nível 0,05 (2 extremidades)
**. A correlação é significativa no nível 0,01 (2 extremidades)
b
. Não é possível calcular porque pelo menos uma das variáveis é constante.
DGPA: comprimento da parte aérea (CPA), diâmetro do hipocótilo (DH), comprimento (CF) e
diâmetro da primeira folha verdadeira (DF).
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
50
Em relação às correlações existentes na Tabela 5, observou-se que das
234 combinações possíveis entre as treze caracteres e seus graus de
resistência (R, MR e S), 27 combinações foram altamente significativas e
significativas a 5 %.
Ao analisar o coeficiente de galhas de plantas R com o desenvolvimento
global do sistema radicular (CSR, MFSR e VSR), observou-se correlações
significativamente positivas. Nas plantas resistentes, é preservada a estrutura
morfológica do sistema radicular e uma baixa população de M. enterolobii, no
entanto, este patógeno determina a retenção de água e nutrientes nas galhas
implicando em um maior volume e massa fresca da raiz.
Ao analisar o coeficiente de galhas de plantas suscetíveis, verificou-se
uma correlação significativamente positiva com o fator de reprodução (FR). Este
resultado afirma a idéia de que quanto mais galhas houver no sistema radicular
de plantas suscetíveis, maior será seu fator de reprodução. Esta característica
confirma a credibilidade dos dados aqui apresentados de acordo com as
encontradas
na
literatura
(TAYLOR;
SASSER,
1978).
Ademais,
serão
necessárias avaliações futuras para uma boa caracterização de outros
parâmetros envolvidos no desenvolvimento do sistema radicular, tais como
ramificação (secundária, terciária etc.), que provavelmente indiquem a
capacidade de regeneração do sistema radicular frente à destruição feita pelo
nematoide das galhas.
No entanto, ao correlacionar galhas com o desenvolvimento global da
parte aérea (CPA, DH, CAA e CBA) das plantas avaliadas, observou-se
correlações significativamente positivas, onde esperava-se que houvesse
correlação negativa. A única correlação significativamente negativa desse
caracter foi para o coeficiente de CAB (-0,95) de plantas R. Provavelmente,
como as plantas R tem um maior DGPA, há maior demanda por nutrientes que
as plantas S. Como no presente ensaio, houve apenas uma adubação de
plantio, alguns nutrientes como nitrogênio das folhas mais velhas podem ter sido
carreadas para as folhas mais jovens. Assim, é sugerido para os demais
experimentos, empregar a adubação de cobertura das plantas.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
51
Quanto ao FR de plantas R, observou-se correlação significativamente
positiva com MFSR (0,97) e com VSR (0,96). Po outro lado, Damaceno (2012)
ao avaliar alguns genótipos do BAG de Cucurbitáceas da Embrapa Semiárido,
observou essa mesma correlação entre massa fresca das raízes e fator de
reprodução, porém em plantas suscetíveis e inoculadas com M. enterolobii.
E para o DGPA, foi constatado correlação positiva com CPA de plantas
suscetíveis (0,96) e correlação negativa com CBA de plantas de média
resistência (-0,99).
Os dados acima relatados mostram que é possível realizar uma seleção
precoce de plantas com potencial de resistência a M. enterolobii, através de
variáveis que inicialmente mensurem o desenvolvimento global da parte aérea e
de galhas, e, posteriormente confirmem com o FR da população selecionada.
Entretanto, a busca de marcadores bioquímicos e/ou moleculares devem ser
estudadas.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L. parasitadas por Meloidogyne enterolobii
52
Tabela 5. Valores dos coeficientes de correlação para a associação entre as variáveis desenvolvimento global do sistema radicular (DGSR), de galhas (DGG), fator
de reprodução (FR) e desenvolvimento global da parte aérea (DGPA) de plantas do acesso BGCIA 240 aos 90 dias após a inoculação. Petrolina, Embrapa
Semiárido, 2015.
CSR
MFSR
VSR
DGG
FR
CPA
DH
CAA
CBA
CAM
CBM
CAB
R
MR
S
R
MR
S
R
MR
S
R
MR
S
R
MR
S
R
MR
S
R
MR
S
R
MR
S
R
MR
S
R
MR
S
R
MR
S
R
MR
S
MFSR
0,98*
-0,52 ns
-0,72 ns
VSR
0,99*
-0,61 ns
-0,97*
0,99**
0,99**
0,87 ns
DGG
0,99*
-0,44 ns
0,83 ns
0,99**
0,63 ns
-0,21 ns
0,99**
0,64 ns
-0,66 ns
FR
0,91 ns
0,31 ns
0,75 ns
0,97**
0,57 ns
-0,08 ns
0,96*
0,48 ns
-0,56 ns
0,94 ns
-0,06 ns
0,99**
CPA
0,99**
-0,42 ns
0,91 ns
0,99**
0,45 ns
-0,36 ns
1,0**
0,47 ns
-0,77 ns
0,99**
0,97*
0,99*
0,95 ns
-0,26 ns
0,96*
DH
0,97**
-0,41 ns
0,71 ns
0,96*
0,58 ns
-1,00**
0,97*
0,59 ns
-0,87 ns
0,99*
1,00**
0,20 ns
0,88 ns
-0,10 ns
0,07 ns
0,97*
0,98*
0,35 ns
CAA
0,99*
-0,56 ns
0,17 ns
0,99*
-0,19 ns
0,57 ns
1,0**
-0,10 ns
0,10 ns
1,00**
0,45 ns
0,68 ns
0,93 ns
-0,91 ns
0,77 ns
0,99**
0,61 ns
0,56 ns
0,99*
0,48 ns
-0,58 ns
CBA
0,83*
-0,44 ns
0,88 ns
0,86 ns
-0,49 ns
-0,29 ns
0,88 ns
-0,39 ns
-0,72 ns
0,90 ns
0,04 ns
1,00**
0,80 ns
-0,99*
0,98*
0,85 ns
0,23 ns
1,00**
0,94 ns
0,07 ns
0,28 ns
0,90 ns
0,91 ns
0,62 ns
CAM
0,71 ns
-0,69 ns
0,83 ns
0,62 ns
-0,25 ns
-0,98*
0,61 ns
-0,14 ns
-0,95 ns
0,58 ns
-0,10 ns
0,38 ns
0,52 ns
-0,79 ns
0,26 ns
0,65 ns
0,20 ns
0,52 ns
0,53 ns
-0,10 ns
0,98*
0,59 ns
0,73 ns
-0,41 ns
0,20 ns
0,88 ns
0,46 ns
CBM
0,56 ns
-0,56 ns
0,67 ns
0,43 ns
-0,41 ns
-1,00**
0,43 ns
-0,30 ns
-0,84 ns
0,42 ns
-0,16 ns
0,15 ns
0,31 ns
-0,87 ns
0,02 ns
0,48 ns
-0,07 ns
0,30 ns
0,38 ns
-0,14 ns
1,0**
0,43 ns
0,76 ns
-0,62 ns
0,04 ns
0,93 ns
0,23 ns
0,97*
0,98*
0,97*
CAB
-0,99*
-0,34 ns
0,98*
-0,94 ns
0,08 ns
-0,85 ns
-0,95 ns
0,12 ns
-1,00**
-0,95*
-0,61 ns
0,70 ns
-0,84 ns
0,18 ns
0,59 ns
-0,96*
-0,69 ns
0,80 ns
-0,94 ns
-0,66 ns
0,84 ns
-0,96*
-0,32 ns
-0,49 ns
-0,77 ns
-0,05 ns
0,75 ns
-0,78 ns
0,40 ns
0,93 ns
-0,66 ns
0,31 ns
0,81 ns
CBB
-0,98*
-0,35 ns
0,77 ns
-0,92 ns
0,58 ns
-1,00**
-0,93 ns
0,10 ns
-0,91 ns
-0,94 ns
-0,62 ns
0,29 ns
-0,81 ns
0,15 ns
0,16 ns
-0,95 ns
-0,69 ns
0,43 ns
-0,93 ns
-0,66 ns
1,0**
-0,95
-0,29 ns
-0,50 ns
-0,75 ns
-0,02 ns
0,36 ns
-0,79 ns
0,43 ns
0,99**
-0,69 ns
0,34 ns
0,99**
1,00**
1,00**
0,89 ns
*. A correlação é significativa no nível 0,05 (2 extremidades); **. A correlação é significativa no nível 0,01 (2 extremidades); DGSR: comprimento do sistema
radicular (CSR), massa fresca do sistema radicular (PSR) e volume do sistema radicular (VSR); DGPA: comprimento da parte aérea (CPA), diâmetro do hipocótilo
(DH), clorofilas A e B, na região apical (CAA e CBA), mediana (CAM e CBM) e basal (CAB e CBB).
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
53
5.3. Reação de genótipos de melancia a Meloidogyne enterolobii
Plantas do acesso de melancia forrageira BGCIA 240, considerada
resistente, e do acesso melancia de mesa BGCIA 875, considerada suscetível,
apresentaram ao longo das avaliações número de galhas por planta
semelhantes estatisticamente (Figura 7A). Entretanto, aos 35 dias após a
inoculação com M. enterolobii, plantas do acesso suscetível apresentou 87,5%
mais galhas que a acesso resistente. Segundo Taylor e Sasser (1978) uma
planta que apresenta índice de galhas entre 31 a 100, é considerada altamente
suscetível.
Dos 7 aos 28 dias após a inoculação, não se observou efeito dos acessos
para o número de ovos por planta. Aos 35 dias após a inoculação, plantas
suscetíveis apresentaram maior número de ovos por planta do que plantas do
acesso
resistente,
diferindo
estatisticamente.
Observou-se
nas
plantas
suscetíveis 8,08 vezes mais ovos que nas plantas resistentes (Figura 7B). O
maior número de ovos observados no acesso BGCIA 875 indica seu estado de
suscetibilidade. Por outro lado, mesmo havendo formação de galhas em raízes
do acesso BGCIA 240, observou-se uma baixa produção de ovos de M.
enterolobii. Embora as plantas resistentes sejam invadidas pelos nematoides, o
parasitismo não é bem sucedido devido a mecanismos de resistência das
plantas (TRUDGIL; BLOK, 2001). Assim, plantas resistentes podem reconhecer
substâncias liberadas pelo patógeno e desencadear respostas de defesa contra
o invasor (FRAGOSO et al., 2007).
Para comprimento de raiz, observou-se que a inoculação não afetou o
crescimento da mesma, não havendo diferenças significativas em cada acesso
de melancia inoculada e não inoculada. Em média, o acesso de melancia
forrageira apresentou maior comprimento do que a melancia de mesa (Tabela 6).
O comprimento da raiz pode estar relacionado com estratégias de sobrevivência
desenvolvida pelas variedades de melancias forrageiras. A melancia forrageira
adaptou-se bem às condições semiáridas, assim como a estresse hídrico,
nutricional ou ataque de patógenos do solo (QUEIRÓZ, 1993; ROMÃO, 1995).
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
54
Com isso, supõem-se que a medida que as galhas foram se desenvolvendo, as
raízes secundárias ficaram impedidas de absorver nutrientes induzindo o
alongamento axial da raiz. O mesmo não foi observado para o acesso BGCIA
875, talvez por se tratar de uma variedade comercial suscetível. Na avaliação da
massa fresca de raiz, foi observado comportamento semelhante ao do
comprimento de raiz. Não foi observado efeito significativo da inoculação na
massa fresca das raízes em cada acesso de melancia (Tabela 6). Este resultado
provavelmente está associado com a formação das galhas nas raízes das
plantas inoculadas aumentando inicialmente a massa nas plantas inoculadas.
Em média, a massa fresca foi maior no acesso de melancia forrageira.
Afim de compreender o mecanismo de infecção de M. javanica em plantas
de soja, Fragoso et al. (2007) demonstraram que a superexpressão do gene de
corismato mutase I do fitonematoide altera a via do xiquimato por causa da
degradação do corismato citoplasmático, que resulta num fenótipo de redução
ou aborto das radícolas laterais, provavelmente como consequência da redução
dos níveis de auxinas nos plastídios. Assim, no presente trabalho foi observado
para a acesso BGCIA 240 uma recuperação do sistema radicular aos 28 e 35
DAI e para o acesso BGCIA 875 uma contínua queda dos valores de massa
fresca de raiz.
Os dados de clorofila a e b para as épocas estudadas não diferiram
estatisticamente entre as plantas inoculadas e não inoculadas, nem entre os
genótipos (Tabela 6). Provavelmente, esse fato ocorreu devido ao pouco tempo
após a inoculação. Os sintomas reflexos na parte aérea da planta pela presença
do nematoide na raiz aparecem num estágio mais avançado da infecção e na
fase de produção, caracterizando um maior dreno de nutrientes para os frutos.
Por sua vez, Lubarino (2012) ao estudar a relação do teor de clorofila com a
ação do M. enterolobii em plantas inoculadas de melancia, observou, aos 60
dias após a inoculação, um menor teor de clorofila em plantas inoculadas com
relação as não inoculadas. Em geral, o índice médio de clorofila a foi maior que
o de clorofila b, que ocorre devido a uma maior proporção do fotossistema I que
é mais rico em clorofila a, em condições normais de luminosidade,
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
55
(CRITCHELEY, 1999) e menor clorofila b, uma vez que, esta tem uma maior
proporção em ambientes sombreados (SOUZA et al., 2011).
A
N° de galhas/planta
50
ns
40
30
20
ns
10
ns
0
7
B
ns
ns
14
21
28
Dias após inoculação
400
BGCIA240R+
N° de ovos/planta
35
BGCIA 875+
*
300
200
100
ns
ns
ns
ns
14
21
28
0
7
35
Dias após inoculação
BGCIA240
BGCIA 875
Figura 7. Número de galhas (A) e número de ovos (B) por planta de dois
genótipos
de
melancia
do
Banco
Ativo
de
Germoplasma
(BAG)
de
Cucurbitáceas da Embrapa Semiárido, acessos BGCIA 240 (resistente) e BGCIA
875 (suscetível) inoculadas com Meloidogyne enterolobii, avaliadas em
diferentes épocas de coleta. * Significativo a 5% de probabilidade e
significativo pelo teste T.
ns=
não
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
56
Tabela 6. Comprimento de raiz, massa fresca de raiz, clorofila a e clorofila b de
dois genótipos de melancia do Banco Ativo de Germoplasma de Cucurbitáceas
da Embrapa Semiárido, acessos BGCIA 240 e BGCIA 875 inoculadas e não
inoculadas com Meloidogyne enterolobii em diferentes épocas de avaliação.
Comprimento de raiz (cm)
Acessos
Dias após inoculação
7
14
21
28
35
BGCIA 240+
16,25 a
12,90 a
14,30 ab
21,75 a
21,17 a
BGCIA 240
17,00 a
15,50 a
16,17 a
13,40 b
9,33 a
BGCIA 875+
10,25 b
10,50 a
10,50 b
9,83 b
12,25 a
BGCIA 875
10,73 b
10,87 a
10,83 b
11,80 b
11,75 a
Massa fresca de raiz (g)
7
14
21
28
35
BGCIA 240+
2,66 a
1,78 a
1,46 ab
1,61 a
1,73 a
BGCIA 240
2,54 ab
1,85 a
2,22 a
1,24 a
1,14 a
BGCIA 875+
0,61 b
0,63 a
0,63 b
0,29 b
0,60 a
BGCIA 875
0,99 ab
0,69 a
1,00 ab
0,35 b
1,11 a
Clorofila a (IFC)
7
14
21
28
35
BGCIA 240+
24,90 a
32,07 a
31,80 a
28,03 a
24,57 b
BGCIA 240
26,00 a
33,90 a
34,47 a
27,73 a
28,93 ab
BGCIA 875+
24,10 a
28,90 a
31,80 a
28,40 a
28,57 ab
BGCIA 875
26,33 a
30,40 a
36,17 a
30,03 a
30,63 a
Clorofila b (IFC)
7
14
21
28
35
BGCIA 240+
9,33 a
14,67 a
15,37 a
14,90 a
10,83 a
BGCIA 240
12,90 a
15,07 a
16,83 a
13,33 a
11,07 a
BGCIA 875+
12,33 a
11,87 a
12,67 a
12,10 a
9,27 a
BGCIA 875
12,53 a
13,27 a
18,73 a
13,40 a
11,97 a
+= genótipos inoculados; IFC= Índice Falker de Clorofila. Médias com mesma
letra nas colunas não diferem entre si de acordo com o teste de Tukey (P <
0,05).
5.4. Respostas bioquímicas de defesa
As proteínas presentes na planta podem estar envolvidas no processo de
defesa bioquímica contra os danos causados pelo patógeno (BERNARDS et al.,
1999). Além disso, espécies vegetais sensíveis ao nematoide das galhas podem
apresentar a quantidade ou qualidade das proteínas alteradas. Não foi
observado efeito da inoculação com o M. enterolobii nos teores de proteínas
solúveis totais nos dois acessos estudados nos tempos avaliados, exceto aos 35
dias após a inoculação que o genótipo suscetível (BGCIA 875) inoculado
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
57
apresentou menor teor de proteínas que os não inoculados, época de maior
formação de galhas e número de ovos observado neste genótipo (Figura 8- A).
Para a guaiacol peroxidase, foi observado que não ocorreram diferenças
na atividade desta enzima em plantas resistentes e suscetíveis infectadas e não
infectadas com o M. enterolobii até os 14 dias após a inoculação (Figura 8-B). A
acesso com maior nível de resistência (BGCIA 240) inoculada com o nematoide
apresentou maior atividade dessa enzima, especialmente nos tempos de 21 e 28
dias após a inoculação, enquanto que a não inoculada apresentou atividade
semelhante à do acesso suscetível BGCIA 875 inoculada e não inoculada. No
presente estudo, foi observado nas plantas resistentes aumento da atividade da
enzima guaiacol peroxidase somente aos 21 DAI, o que significa dizer que
nesse momento há uma interação de reconhecimento planta-patógeno,
desencadeando a resposta de defesa da planta contra os danos causados pelo
nematoide.
A atividade da polifenoloxidase foi maior nas plantas do acesso resistente,
tanto para plantas inoculadas como para não inoculadas somente aos 14 dias
após a inoculação (Figura 8-C). Não há relatos que correlacionem o aumento da
atividade de polifenoloxidase com a defesa das plantas de melancia aos
nematoides. Entretanto, para outras culturas como a soja estudos tem mostrado
que plântulas de soja de variedade resistente e inoculadas com J2 de M.
javanica está associada a um aumento na atividade da PPO 72 horas após a
inoculação (SHIMIZU, 2004).
A atividade da ascorbato peroxidase foi maior nas plantas do acesso
resistente (BGCIA 240), tanto inoculada como não inoculada com M. enterolobii,
até os 21 dias após a inoculação, superiores à acesso suscetível (Figura 8-D).
Não foi observado diferença nas atividades da catalase entre os acessos
suscetíveis (BGCIA 875) e resistentes de melancia (Figura 8-E).
O resultado de não haver diferença entre plantas inoculadas e não
inoculadas do BGCIA 240 para a atividade da PPO até 14 DAI e APX, indica o
possível uso destas enzimas como marcadores de seleção de genótipos, pois
diferentes quantidades são observadas entre os dois acessos, BGCIA 240 e
BGCIA 875.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
58
Para a atividade da fenilalanina amônia-liase, aos 14 dias após a
inoculação, foi observado aumento significativo nas raízes de plantas do acesso
BGCIA 240, inoculadas, em relação às plantas do acesso suscetível, BGCIA
875, também inoculadas (Figura 8-F). Este aumento de atividade provavelmente
reflete o aumento da utilização de compostos fenólicos em raízes resistentes. A
partir desse momento houve queda na atividade dessa enzima para as plantas
do acesso BGCIA 240 inoculadas e não inoculadas. Kalaiarasan (2009) ao
avaliar genótipos de tomate resistentes e suscetíveis a M. incognita quanto aos
danos provocados pelo patógeno observou por avaliação bioquímica, que os
genótipos resistentes possuíam maior atividade de fenilalanina amônia-liase que
os genótipos suscetíveis.
Estes resultados mostram o potencial do uso de enzimas na seleção por
tempo de infecção, uma vez que, que o acesso resistente apresentou
precocemente maior atividade da enzima PAL.
Os resultados deste trabalho possibilitam um maior conhecimento dos
mecanismos de defesa envolvidos na resistência da melancia ao nematoide M.
enterolobii. Entre as respostas bioquímicas na defesa das plantas contra
patógenos, as peroxidases, catalases, polifenoloxidases e fenilalanina amônialiases tem recebido destaque em diversos estudos (KALAIARASAN, 2009) pois
estes compostos podem atuar na mitigação dos danos causados pelo patógeno,
ou impedir o avanço do mesmo nos tecidos do hospedeiro (RESENDE et al.,
2003). Estas enzimas podem atuar de forma diferente em diferentes relações
planta-nematoide e, neste trabalho, observou-se que o aumento na atividade das
enzimas peroxidases, polifenoloxidase e fenilalanina amônia-liase indica um
efeito de resposta da planta após o reconhecimento do patógeno.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
59
D
A
200
APX (mM min-1 mg P-1 )
0,80
Proteína (mg mL-1 )
0,70
0,60
0,50
0,40
0,30
0,20
0,10
160
120
80
40
0,00
0
7
14
21
28
35
7
14
Dias após inoculação
BGCIA240+
BGCIA240
BGCIA875+
BGCIA875
B
28
35
BGCIA240+
BGCIA240
BGCIA875+
BGCIA875
E
40000
4,00
3,50
CAT (mM min -1 mg P-1 )
POX (mM min -1 mg P-1 )
21
Dias após inoculação
30000
20000
10000
3,00
2,50
2,00
1,50
1,00
0,50
0
0,00
7
14
21
28
35
7
14
21
Dias após inoculação
28
35
Dias após inoculação
BGCIA240+
BGCIA240
BGCIA240R+
BGCIA240R
BGCIA875+
BGCIA875
BGCIA875+
BGCIA875
C
F
0,25
PAL (mM min-1 mg P-1 )
PPO (mM min -1 mg P-1 )
20
16
12
8
0,20
0,15
0,10
0,05
0,00
4
7
14
21
28
BGCIA875+
7
14
21
28
35
Dias após inoculação
Dias após inoculação
BGCIA240+
35
BGCIA240
BGCIA240R+
BGCIA240R
BGCIA875
BGCIA875+
BGCIA875
Figura 8. Proteínas solúveis totais (A), atividades das enzimas guaiacol peroxidase
(POX) (B), polifenoloxidase (PPO) (C), ascorbato peroxidase (APX) (D), catalase
(CAT)(E) e fenilalanina amônia liase (PAL) (F) de dois genótipos de melancia do Banco
Ativo de Germoplasma (BAG) de Cucurbitáceas da Embrapa Semiárido, acessos
BGCIA 240 e BGCIA 875 inoculadas com Meloidogyne enterolobii, e seus controles,
avaliadas em diferentes épocas de coleta. Barras representam erro padrão da média
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
60
6. CONCLUSÕES
A avaliação baseada nos métodos não destrutivos permitiu selecionar
plantas resistentes dentro da população do acesso BGCIA 240, num percentual
de 46%.
Os parâmetros utilizados, desenvolvimento global da parte aérea e
número de galhas, apresentaram correlação positiva com o FR e um elevado
percentual de acerto na classificação precoce e não destrutiva do grau de
resistência de genótipos de melancia a Meloidogyne enterolobii.
Os métodos não destrutivos utilizados foram validados pelos métodos
tradicionais destrutivos indicando ser uma ferramenta alternativa não destrutiva
importante para os futuros trabalhos de melhoramento genético de plantas
resistentes à Meloidogyne spp.
Com relação a reação ao patógeno, o acesso BGCIA 240 resistente
apresentou menor número de galhas e de ovos, aos 14 dias após inoculação.
Este estudo demonstrou, que a presença do patógeno induziu, no acesso
resistente inoculada com M. enterolobii, maior valor de comprimento de raiz aos
28 DAI e menor massa fresca de raiz aos 21 DAI.
O acesso BGCIA 240 resistente apresentou maior atividade das enzimas
guaiacol peroxidase, ascorbato peroxidase, polifenoloxidases e fenilalanina
amônia-liase.
As enzimas POX, APX, PPO e PAL indicaram ser compostos potenciais
para serem utilizados como marcadores bioquímicos de resistência a M.
enterolobii em melancia.
O uso da atividade de enzima, também se mostrou promissora como
marcadores na seleção por tempo de infecção, uma vez que, o acesso resistente
apresentou precocemente maior atividade da enzima PAL, e na seleção de
genótipos, uma vez que, não houve diferença entre plantas inoculadas e não
inoculadas dentro do acesso, e sim entre os acessos.
Este trabalho também trouxe dados inéditos na literatura científica, com
os primeiros relatos de estudos fitoquímicos em raízes de Citrullus lanatus L.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
61
RERERÊNCIAS
ABREU, F.A. Herança de resistência Phytophthora infestans, de
características de frutos e seleção de genótipos resistentes em geração F5
de cruzamentos interespecíficos em tomateiro. 2005. 107f. Tese (Doutorado
em Genética e Melhoramento). Universidade Federal de Viçosa, Viçosa, 2005.
AGRIOS, G.N. Plant Pathology. 5th ed, San Diego: Elsevier Academic Press, p.
922, 2005.
AGROFIT: http://www.agrofit.com.br/novoportal/. acessado em 26 jan 2015.
ALMEIDA, E. J.; ALVES, G. C. S.; SANTOS, J. M.; MARTINS, A. B. G.
Assinalamentos de Meloidogyne enterolobii em goiabeira e em plantas invasoras
no estado de São Paulo, Brasil. Nematologia Brasileira, Piracicaba-SP, Brasil.
v. 35 (1-2), p. 50-52, 2011.
ARAÚJO, D. A. L. de; FERREIRA, M. A. J. F.; NASCIMENTO, T. L. do; ARAÚJO,
C. de L.; ARAÚJO, L. S.; SENA, E. M. N. de; LIRA, I. C. de S. A. L.; PASSOS, L.
R. G. Seleção recorrente em melancia forrageira para as condições
Semiáridas. In: Congresso Brasileiro de Melhoramento de Plantas. Uberlândia.
Variedade melhorada: a força da nossa agricultura. Anais... Viçosa, MG: SBMP,.
p. 2869-2871, 2013
ARYAL, S.K.; DAVIS, R.F.; STEVENSON, K.L.; TIMPER, P.; JI, P. Influence of
infection of cotton by Rotylenchulus reniformis and Meloidogyne incognita on the
production of enzymes involved in systemic acquired resistance. Journal of
Nematology. v. 43, 3-4, p. 152-159, 2011.
AZEVEDO, R. A.; ALAS, R. J.; SMITH, R. J.; LEA, P. J. Response of antioxidant
enzymes to transfer from elevated carbon dioxide to air ozone fumigation, in the
leaves and roots of wild-tye and a catalase-deficient mutant of barley. Physiol.
Plant. 104, 280-292, 1998.
BERNARDS, M. A.; FLEMING, D. W.; LLEWELLYN, D. B.; PRIEFER, R.; YANG,
X.; SABATINO, A.; PLOURDE, G. L. Biochemical characterization of the
suberisation-associated anionic peroxidase of potato. Plant Physiology,
Bethesda, 121, 135-146, 1999.
BAKER, B.; ZAMBRYSKI, P.; STASKAWICZ, B.; DINESH-KUMAR, S. P.
Signaling in plant-microbe interactions. Science, v. 276, n. 5313, p. 726- 733,
1997
BARROSO, G.M. Sistemática das Angiospermas do Brasil. SP. EDUSP/U.F.
Viçosa. vol. 1,2,3. 2001, 1984, 1986.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
62
BARROSO, G.M. Sistemática de Angiospermas do Brasil. Livro Técnico e
Científico. EDUSP, São Paulo, p. 255, 1978.
BELKHADIR, Y.; SUBRAMANIAM, R.; DANGL, J.L. Plant disease resistance
protein signaling: NBS-LRR proteins and their partners. Current Opinion in
Plant Biology, v.7, n.4, p.391-399, 2004.
BERNARDS, M. A.; FLEMING, D. W.; LLEWELLYN, D. B.; PRIEFER, R.; YANG,
X.; SABATINO, A.; PLOURDE, G. L. Biochemical characterization of the
suberisation-associated anionic peroxidase of potato. Plant Physiology,
Bethesda, v. 121, p. 135-146, 1999.
BIRD, D. M.; OPPERMAN, C. H.; DAVIES, K. G. Interactions between bacteria
and plant-parasitic nematodes: now and then. International Jornal for
Parasitology, v. 33, n. 11, p. 1269-1276, 2003
BORÉM, A.; MIRANDA, G.V. Melhoramento de plantas. 5.ed. Viçosa: UFV, p.
529, 2009.
BRADFORD, M. M. A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye binding. Anal.
Biochem., 72, 248-254, 1976
BRUESKE, C.H. Phenylalanine ammonia-lyase activity in tomato roots infected
and resistant to the root knot nematode, Meloidogyne incognita. Physiologycal
Plant Pathology, v. 16, p. 409-414, 1980.
CAMM, E.L.; TOWERS, G.H. Review article-Phenylalanine ammonia lyase.
Phylochemistry, v.12, p. 961-973, 1973.
CARNEIRO, R.M.D.G; MOREIRA, W. A; ALMEIDA, M. R A. Primeiro registro de
Meloidogyne mayaguensis em goiabeira no Brasil. Soc. Bras. Nematol.,
Brasília, v. 25, p. 223-228, 2001
CASTAGNONE-SERENO, P. Genetic variability in parthenogenetic root-knot
nematodes, Meloidogyne spp., and their ability to overcome plant resistance
genes. Nematology, Leiden, v. 4, n. 5, p. 605-608, 2002.
CASTELLANE, P.D.; CORTEZ, G.E.P. A cultura da melancia. Jaboticabal:
FUNEP, p. 1-2, 1995.
CASTRO, J.M.C.; DIAS, R.C.S.; TEIXEIRA, F.A.; DAMACENO, L.S.; BARBOSA,
G.S. Reação de genótipos de melancia à nematoides. In: CONGRESSO
BRASILEIRO DE OLERICULTURA, 51. Anais... Viçosa: ABH. p. 3373-3379,
2011.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
63
CHOI, H.W.; KIM, Y.J.; LEE, S.C.; HONG, J.K.; HWANG, B.K. Hydrogen peroxide
generation by pepper extracellular peroxidase CaPO2 activates local and
systemic cell death and defense response to bacterial pathogens. Plant
Physiology, Rockville, v. 145, p. 890-904, 2007.
COOLEN, W. A.; D’HERDE, C. J. Method for the quantitative extraction of
nematode plant tissue. Ghent State Agriculture Research Center, Ministry of
Agriculture, p. 777, 1972
COSTA, C.P. da; PINTO, C.A.B.P. Melhoramento de hortaliças: Revisão.
Piracicaba: USP-ESALQ, p. 313, 1977.
COSTA FILHO, J.H. Coleta e reação de acessos melancia a Meloidogyne
enterolobii. 2012. 55 f. Dissertação (Mestrado em Fitotecnia). Universidade
Federal Rural do Semi-Árido. 2012.
COYNE, D.L.; NICOL, J.M.; CLAUDIUS-COLE, B. Nematologia prática: Um guia
de campo e de laboratório. SP-IPM Secretariat, International Institute of
Agriculture (IITA), Cotonou, Benin. p. 82, 2007
CRITCHLEY,
C.
Concepts
in
photobiology:
photosinthesis
and
photomorphogenesis. Singhal, G.S., Renger, G., Sopory, S. K., Irrgang, K. D.,
Govindjee, eds.; New Delhi: Narosa publishing House, 587, 1999.
CRUZ, C.D. Programa Genes: Biometria. Editora UFV. Viçosa (MG). p. 382,
2006
DAMACENO LS. Reação de genitores de melancia a Meloidogyne
enterolobii. 2012. 79f. Dissertação (Mestrado em Agronomia). Departamento de
Tecnologia e Ciências Sociais. Universidade do Estado da Bahia, Juazeiro. 2012.
DIAS, R. de C. S.; SILVA, A. F.; COSTA, N. D.; RESENDE, G. M.; SOUZA, F. de
F; ALVES, J. C. Da S. F. de. Sistema de produção da melancia. 2010.
http://sistemasdeproducao.cnptia.embrapa.br. acessado em 29 set. 2014.
DIXON, R.A.; PAIVA, N. Stress-induced phenylpropanoid metabolism. The Plant
Cell, Baltimore, v. 7, p. 1085-1097, 1995.
FERRAZ, L. C. C. B. As meloidoginoses da soja: passado, presente e futuro.
In: SILVA, J. F. V. (Org.). Relações parasito-hospedeiro nas meloidoginoses da
soja. Londrina: EMBRAPA Soja; Sociedade Brasileira de Nematologia, p. 15-38,
2001.
FERREIRA, D. F. Análises estatísticas por meio do SISVAR (Sistema para
análise de variância) para Windows versão 4.0. Reunião anual da Região
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
64
Brasileira da Sociedade Internacional de Biometria, Universidade Federal de São
Carlos, Brasil, 2000.
FRAGOSO, F.R.; LOURENÇO, I.T.; VIANA, A.M.B.; SOUZA, D.S.L.; ANDRADE,
R.V.; MEHTA, A.; BRASILEIRO, A.C.M.; PINTO, E.R.C.; LIMA, L.M.; ROCHA,
T.L.; SÁ, M.F.G. Interação Molecular Planta-Nematoide. Documentos 198
Embrapa: Brasília. p. 56, 2007.
FRANCO, A.J.; CHARLO, H.C.O.; GALATTI, F.S.; BRAZ, L.T. Avaliação de
genótipos de cucurbitáceas quanto à resistência à Meloidogyne incognita.
Resumos do 48° Congresso Brasileiro de Olericultura, Maringá, Paraná,
Brasil, 2008.
FRANCO, O.L.; PEREIRA, J.L.; COSTA, P.H.A.; ROCHA, T.L.; ALBUQUERQUE,
E.V.S.; GROSSI-DE-SA, M.F.; CARNEIRO, R.M.D.G.; CARNEIRO, R.G.;
MEHTA, A. Methodological evaluation of 2-DE to study root proteomics during
nematode infection in cotton and coffee plants. Preparative Biochemistry &
Biotechnology, v.40, p.52-163, 2010.
GAMA, R. N. C de S.; DIAS, R. de C. S.; ALVES, J. C. da S. F.; DAMACENO, L.
S.; TEIXEIRA, F. A.; AMARAL, C. M. do. Seleção de porta-enxertos para
cultivares de melancia. In: Congresso Brasileiro de Recursos Genéticos. Anais...
Salvador, 2010.
GAMA, R. N. C de S.; DIAS, R. de C. S.; ALVES, J. C. da S. F.; DAMACENO, L.
S.; TEIXEIRA, F. A.; BARBOSA, G. da S. Taxa de sobrevivência e desempenho
agronômico de melancia sob enxertia. Horticultura Brasileira, v. 31, p. 128-132,
2013.
GAUILLARD, F.; FORGET, F. R.; NICOLAS, J. New spectrophotometric assay for
polyphenol oxidase activity. Anal. Biochem., 215, 59-65, 1993.
GHEYSEN, G.; FENOLL, C. Gene expression in nematode feeding sites. Annual
Review of Phytopathology, v. 40, p. 191-219, 2002.
GIACOMETTI, D.C. Descritores para caracterização e avaliação de
germoplasma. Anais do I Encontro Sobre Recursos Genéticos, Jaboticabal,
Brasil, p.129- 147, 1988.
GIEBEL, J. Phenylalanine and tyrosine ammonia-lyase activities in potato roots
and their significance in potato resistance to Heterodera rostochiensis.
Nematologica, v.19, p. 1-6, 1973.
GUZZO, S.D.; MARTINS, E.M.F. Local and systemic induction of β-1,3glucanase and chitinase in coffee leaves protected against Hemileia vastatrix by
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
65
Bacillus thuringiensis. Journal of Phytopathology, Berlin, v. 144, n. 9-10, p.
449-454, 1996.
HERRMANN, K.M.; WEAVER, L.M. The shikimate pathway. Annual Review of
Plant Physiology and Plant Molecular Biology, Palo Alto, v. 50, p. 473-503,
1999.
HIRAGA, S.; SASAKI, K.; ITO, H.; OHASHI, Y. MATSUI, H. A large family of class
III plant peroxidases. Plant and Cell Physiology, Oxford, UK, v. 42, n. 2, p. 462468, 2001.
HUANG, J. S. Mechanisms of resistance to root-knot nematodes. In:
SASSER, J. N.; CARTER, C. C. An advanced treatise on Meloidogyne:
Biology and control. Raleigh: North Caroline State University Graphics, v. 1,
p.11-17, 1985.
HUSSEY, R.S.; BARKER, K.R. A comparison of methods of collecting inoculated
of Meloidogyne spp., including a new technique. Plant Disease Reporter,
Beltsville, v.57, n.4, p.1025-1028, 1973.
JANEGITZ, T. Efeitos de cis-jasmone na indução de compostos fenólicos
em genótipos de soja, inoculados ou não com Meloidogyne javanica. 2012.
62f. Dissertação (Mestrado em Agronomia). Universidade Estadual de Maringá,
UEM, Brasil. 2012.
JANEGITZ, T.; VIEIRA, S. S.; GRAÇA, J. P.; UEDA, T.; SALVADOR, M. C.;
LEMES, T.; OLIVEIRA, M. C. N.; DIAS, W.; FERRARESE-FILHO, O.;
HOFFMANN-CAMPO, C. B. Indução de resistência de genótipos de soja a
Meloidogyne javanica. VI Congresso Brasileiro de Soja. Anais... Cuiabá - MT.
2012.
JOBST, J.; KING, K.; HEMLEBEN. V. Molecular evolution of the internal
transcribed spacers (ITS1 and ITS2) and phylogenetic relationships among
species of the family Cucurbitaceae. Molecular Phylogenetics and Evolution,
v. 9, p. 204-219, 1998.
KALAIARASAN, P. Biochemical markers for identification of root knot nematode
(Meloidogyne incognita) resistance in tomato. Karnataka Journal of
Agricultural Sciences, v. 22, n. 3, p. 471-475, 2009.
KAPLAN, D.T.; KEEN, N.T.; THOMASON, I.J. Studies on the mode of acyion of
glyceollin in soybean. Incompatibility to the root knot nematode, Meloidogyne
incognita. Physiological and Molecular Plant Pathology, v. 16, p. 319-325,
1980.
KOVÁCIK, J.; KLEJDUS, B.; BACKOR, M.; REPCAK, M. Phenylalanine
ammonia-lyase activity and phenolic compounds accumulation in nitrogen-
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
66
deficient Matricaria chamomilla leaf rosettes. Plant Science, v. 172, p. 393-399,
2007.
LARCHER, W. Ecofisiologia Vegetal. Editora Pedagógica Universitária Ltda.
São Paulo. p. 319, 1986.
LIMA, R.D.; DIAS, W.P.; CASTRO, J.M. da C. Doenças causadas por
nematoides em cucurbitáceas. Informe Agropecuário, Belo Horizonte, v. 17, n.
182, p. 57-59, 1995.
LIMA, G.S.A.; ASSUNÇÃO, I.P.; VALLE, L.A.C. Controle genético de doenças
radiculares. In: MICHEREFF, S.J.; DOMINGOS, E.G.T.; ANDRADE, M.M.
Ecologia e manejo de patógenos radiculares em solos tropicais. Recife: UFRP. p.
248 – 272. 2005.
LORENCETTI, C.; CARVALHO, F. I. F.; BENIN, G.; MARCHIORO, V. S.;
OLIVEIRA, A. C.; SILVA, J. A. G.; HARTWIG, I.; SCHMIDT, D. A. M.; VALÉRIO, I.
P. Capacidade combinatória e heterose em cruzamento dialélico de aveia (Avena
sativa L.). Revista Brasileira de Agrociência, Pelotas, v.11, n. 2, p. 143-148,
2005
LUBARINO, P. C. da C. Caracterização e avaliação de genótipos de Citrullus
lanatus L. quanto à resistência a nematoide das galhas, Meloidogyne enterolobii.
Trabalho de Conclusão de Curso apresentado ao Departamento de Tecnologia e
Ciências Sociais, da Universidade do Estado da Bahia, 2012.
LUBARINO, P. C. da C.; DIAS, R. C. S.; COSTA, J. da A.; SANTOS, J. S. dos;
SOUZA, R. R. C. de; SANTOS, S. de S. Avaliação morfoagronômica de acessos
de melancia do BAG de Cucurbitáceas da Embrapa Semiárido. In: II Congresso
Brasileiro de Recursos Genéticos, 2012, Belém-PA. II Congresso Brasileiro de
Recursos Genéticos, 2012.
MARAHATTA, S.P.; WANG, K.H.; SIPES, B.S.; HOOKS, C.R. Effects of Tagetes
patula on Active and Inactive Stages of Root-Knot Nematodes. The Journal of
Nematology, v. 44, n.1, p. 26-30, 2012.
MARANHÃO, S. R. V. L.; MOURA, R. M. de; PEDROSA, E. M. R. Reação de
Indivíduos Segregantes de Goiabeira a Meloidogyne incognita raça 1 e M.
mayaguensis. Nematologia Brasileira, Pelotas - RS, v. 25, n.2, p. 191-195,
2001.
MARIUTTO, M.; DUBY, F.; ADAM, A.; BUREAU, C.; FAUCONNIER, M. L.;
ONGENA, M.; THONART, P.; DOMMES, J. The elicitation of a systemic
resistance by Pseudomonas putida BTP1 in tomato involves the stimulation of
two lipoxygenase isoforms. BMC Plant Biology, v. 11, n. 29, 2011.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
67
MARTINS-MIRANDA, A.S. Atividade de enzimas relacionadas com estresses
bióticos e abióticos em plântulas de feijão-de-corda [Vigna unguiculata (L.)
Walp.] expostas à salinidade e deficiência hídrica. 2002. 85f. Dissertação
(Mestrado em Bioquímica) – Departamento de Bioquímica e Biologia Molecular,
Universidade Federal do Ceará, Fortaleza. 2002.
MAYER, A.M. Polyphenol oxidases in plants and fungi: going places? A review.
Phytochemistry, Oxford, UK, v. 67, p. 2318-2331, 2006.
MEEUSE A. D. J. The Cucurbitaceae of southern Africa. Bothalia, v.8, p. 1–111,
1962.
MICHEREFF, S. J.; ANDRADE, D. E. G. T.; MENEZES, M. Ecologia e Manejo de
Patógenos Radiculares em Solos tropicais. Recife: UFRPE, p. 4-310, 2005
MOHR, H.C. Watermelon Breeding. In: BASSET, M.L. (Ed) Breeding vegetables
crops. Westport A vi, p. 33-66, 1986.
MORI, T.; SAKURAI, M.; SAKUTA, M. E. Vects of conditioned medium on
activities of PAL, CHS, DAHP synthase (DS-Co and DS-Mn) and anthocyanin
production in suspension cultures of Fragaria ananassa. Plant Science, 160,
355-360, 2001
MOLINARI, S.; MIACOLA, C. Catalase induction in galls produced by
Meloidogyne-tomato root interactions in vitro. Nematologia Mediterranea, v. 25,
n. 2, p. 299-303, 1997
MOURA, R.M.; REGIS, E.M.O. Reação de cultivares de feijoeiro comum
(Phaseolus vulgaris) em relação ao parasitismo de Meloidogyne javanica e M.
incognita (Nematoda: Heteroderidae). Nematologia Brasileira, v.11, p.215-225,
1987.
NAKANO, Y.; ASADA, K. Hydrogen peroxide is scavenged by ascobate-specific
peroxidase in spinach chloroplasts. Plant Cell Physiol., 22, 867-880, 1981.
NUEZ, F.; PHOHENS, A; et al. Catálogo de Simillas de Sandía. Banco de
Germoplasma de la Universidad Politécnica de Valência. Madrid: p. 16-17,
25- 28, 1998.
OLIVEIRA, M. C. de; BERNARDINO, F. A. Melancia forrageira, um novo recurso
alimentar para a pecuária das regiões secas do Nordeste do Brasil. Petrolina:
Embrapa Semiárido, 2000. p.17. (Circular técnica, 49).
PARKER, J. E.; FEYS, B. J.; VAN DER BIEZEN, E. A.; NÖEL, L.; ARTS, N.
Unravelling R gene-mediated resistance pathways in Arabidopsis. Molecular
Plant Pathology, v. 1, p. 17-24, 2000.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
68
PASSARDI F.; PENEL C.; DUNAND C. Performing the paradoxical:Howplant
peroxidases modify the cell wall. Trends Plant Sci. v. 9, p. 534–40, 2004.
PEREIRA F. M., SOUZA, R. M., SOUZA, P. M., DOLINSKI, C.; SANTOS, G. K.
Estimativa do impacto econômico e social direto de Meloidogyne mayaguensis
na cultura da goiaba no Brasil. Soc. Bras. Nematol., v. 33, p. 176–181, 2009
PERRY, R. N; MOENS, M. Plant Nematology. CABI, 2nd Edition, 2006.
POLIZEL, A. C.; MENEZES, P. C.; SILVA, A. R. B.; GUIMARÃES, S. L.; SILVA,
M. A P. Teor de clorofila e severidade da ferrugem asiatica em funcao de
cultivares, aplicacao de fungicida e epoca de semeadura da soja. Enciclopédia
biosfera, v. 7, p. 1-13, 2011.
POLLE, A. Dissecting the Superoxide Dismutase-Ascorbate-Glutathione-Pathway
in Chloroplasts by Metabolic Modeling: Computer Simulations as a Step towards
Flux Analysis. Plant Physiology, v.126, p. 445-462, 2001.
POFU, K.M.; MASHELA, P.W.; MPHOSI, M.S. Management of Meloidogyne
incognita in nematode susceptible watermelon cultivars using nematode resistant
Cucumis africanus and Cucumis myriocarpus rootstocks. Afr. J. Biotech., 10,
8790-8793, 2011.
PONTES, M. F. C. Resistência de melancieira a Meloidogyne mayaguensis e
avaliação dos mecanismos envolvidos. 2009. Tese (Doutorado em Fitopatologia).
Universidade Federal Rural de Pernambuco, UFRPE, Brasil, 2009.
QUEIRÓZ, M. A. Potencial do germoplasma de Cucurbitáceas no Nordeste
brasileiro. Horticultura Brasileira. Brasília-DF, v. 11, n. 1, p. 7-9, 1993.
QUEIRÓZ, M. A. Germplasm of Cucurbitaceae in Brazil. Crop Breeding and
Applied Biotechnology, Viçosa, MG, v. 4, n.4, p. 377-383, 2004.
RIZZO AAN; CHAVES FCM; LAURA VA; GOTO R. Avaliação de métodos de
enxertia e porta-enxertos para melão rendilhado. Horticultura Brasileira, v. 22,.
p. 808-810, 2004.
RESENDE, M. L. V.; SALGADO, S. M. L.; CHAVES, Z. M. Espécies ativas de
oxigênio na resposta de defesa de plantas a patógenos. Reactive oxygen
species on plant defense responses to pathogens. Fitopatologia Brasileira,
v.28, n.2, p.123-130, 2003.
RIGON, J. P.G. ; CAPUANI, S. ; BELTRÃO, N. E. de M. ; BRITO NETO, J. F. ;
SOFIATTI, V. ; SILVA, F. V. F. Non-destructive determination of photosynthetic
pigments in the leaves of castor oil plants. Acta Scientiarum. Agronomy
(Impresso), v. 34, p. 325-329, 2012.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
69
RODRÍGUEZ-KÁBANA, R.; KOKALIS-BURELLE, N.; ROBERTSON, D. G.;
KING, P. S.; WELLS, L. W. Rotations with coastal bermudagrass, cotton and
bahiagrass for management of Meloidogyne arenaria and southern blight in
peanut. Journal of Nematology, v. 26, p. 665-668, 1994.
ROMÃO, R. L. Dinâmica evolutiva e variabilidade genética de populações
de melancia [Citrullus lanatus (Thumb.) Matsum & Nakai] em três regiões
do Nordeste brasileiro. 1996. 75 f. Dissertação de mestrado, Escola Superior
de Agricultura “Luiz de QUEIRÓZ”, São Paulo. 1995.
RYAN, C. A.; JAGENDORF, A. Self defense by plants. Proceedings of the
National Academy of Sciences of the United States of America, v.92, p.
4075, 1995.
SAHEBANI, N.; ZAD J.; SHARIFI TEHRANI, A.; KHEYRI, A. A. Study of changes
in peroxidase activity in interaction between root-knot nematode (Meloidogyne
javanica) and tomato fusarium wilt agent (Fusarium oxysporum f.sp. lycopersici).
Iranian Journal of Agricultural Sciences (Journal of Agriculture), v 39, n. 1, p.
127 – 138, 2008.
SANTANA, F.A. Análise proteômica de raízes de soja em resposta à
inoculação com nematoides de galha. 2012. Tese (Doutorado em Genética e
Melhoramento), Universidade Federal de Viçosa, MG, 2012.
SATURNINO, H.M.; PAIVA, B.M.; GOTIJO, V.P.M; FERNANDES, D.P.
Cucurbitáceas: aspectos estatísticos. Informe Agropecuário, v. 8, p. 3-21, 1982.
SCANDALIOS, J.G. Oxidative stress: molecular perception and transduction of
signals triggering antioxidant gene defenses. Brazilian Journal of Medical and
Biological Research, Ribeirão Preto, v. 38, n. 7, p. 995-1014, 2005
SHIMIZU, Miton Massao. Polifenoloxidase como fator de resistência da soja
a nematóides e na oxidação do palmito. 2004. 108f. Tese (Doutorado em
Biologia Vegetal). Universidade Estadual de Campinas, UNICAMP, Brasil. 2004.
SILVEIRA, L. M. da; QUEIRÓZ, M. A. de; LIMA, J. A de A.; NEGREIROS, M. Z.
de; RAMOS, N. F.; NASCIMENTO, A. K. Q. do. Seleção de acessos e progênies
de Citrullus spp. para resistência a três potyvirus. Fitopatologia Brasileira, v.30,
n.4, p. 394-399, 2005.
SOUZA, F. F.; GAMA, F. C.; QUEIRÓZ, M. A. Análise da capacidade de
combinação em cruzamentos dialélicos de três genótipos de melancia.
Horticultura Brasileira, Brasília, DF, v.22, n.4, p.789-793, 2004.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
70
SOUZA, F. F. de; DIAS, R. de C. S.; QUEIRÓZ, M. A. de. Aspectos botânicos. In:
SOUZA, F. de F. (Ed.). Cultivo da melancia em Rondônia. Porto Velho: Embrapa
Rondônia, p. 82-95, 2008.
SOUZA, G. S.; SILVA, J. S.; SANTOS, A. R.; GOMES, D. G.; OLIVEIRA, U. C.
Crescimento e produção de pigmentos fotossintéticos em alfavaca cultivada sob
malhas coloridas e adubação fosfatada. Enciclopédia Bioesfera, Goiânia, 7,
2011
TAIZ, L.; ZEIGER, E. Plant Physiology. 4th. ed. Sunderland: Sinauer
Associates, p. 764, 2006.
TAYLOR, A. L.; SASSER, J.N. Biology, identification and control of root-knot
nematodes, Meloidogyne species. Raleigh: North Carolina State University
Graphics, p. 111, 1978.
TERAO et al., Sistema de produção da melancia. 2010. Disponível em:
http://sistemasdeproducao.cnptia.embrapa.br. Acesso em: 29 set. 2014.
THIES, J. A.; LEVI, A. Characterization of watermelon (Citrullus lanatus var.
citroides) germplasm for resistance to root-knot nematodes. HortScience 42, p.
1530–1533, 2007.
THURAU T; W, CAI D. Insect and Nematode Resistance. In: Widholm JM,
Nagata T (eds) Biotechnology in Agriculture and Forestry. Heidelberg Berlin
:Springer-Verlag, p. 177-198, 2010.
TIHOHOD, D. Nematologia agrícola aplicada. Jaboticabal: FUNEP, p. 372,
1993.
TRIPATHI, R. D. Plant response to environmental stress. International Book
Distributing Company. p. 476, 2006.
TRUDGILL, D. L.; BLOK, V. C. Apomictic, polyphagous root-knot nematodes:
exceptionally successful and damaging biotrophic root pathogens. Annu. Rev.
Phytopathol., 39, 53-77, 2001.
URBANEK, H., E. KUZNIAK-GEBAROWSKA; HERKA, K.. Elicitation of defence
responses in bean leaves by Botrytis cinereapolygalacturonase. Acta Physiol.
Plant. 13, 43-50, 1991.
VAN LOON, L.C.; REP, M.; PIETERSE, C.M.J. Significance of Inducible Defenserelated Proteins in Infected Plants. Annu. Rev. Phytopathol. v. .44, p. 35-162,
2006.
LUBARINO, P. C. da C. Avaliação fenotípica e bioquímica de Citrullus lanatus L.
parasitadas por Meloidogyne enterolobii
71
WHITAKER, T.W.; DAVIS, G.N. Cucurbits: botany, cultivation and utilization.
London: Leonard Hill Books, p. 250, 1962.
WILCKEN, S. R. S.; GARCIA, M. J. M.; SILVA, N. Resistência de alface tipo
americana a Meloidogyne incognita raça 2. Nematologia brasileira, Piracicaba,
v. 29, n. 2, p. 267-271, 2005.
YORUK, R.; MARSHALL, M.R. Physicochemical properties and function of plant
polyphenol oxidase: a review. Journal of Food Biochemistry, Malden, v. 27, p.
361-422, 2003.
YOSHIMURA, K.; YABUTA, Y.; ISHIKAWA, T.; SHIGEOKA, S. Expression of
spinach ascorbate peroxidase isoenzimes in response to oxidative stresses.
Plant Physiology, v. 123, p. 223-233, 2000.
WILLIAMSON, V. M.; GLEASON, C. A. Plant-nematode interactions. Current
Opinion in Plant Biology, v. 6, n. 4, p. 327-333, 2003
72
APÊNDICE
Artigo submetido à:
Revista: Química Nova
Título: Respostas bioquímicas associadas à defesa em melancia contra
Meloidogyne enterolobii
Autores: Paloma C. da C Lubarino, Leonardo Sousa Cavalcanti, Pedro M.
Ribeiro Junior e Rita de Cássia S. Dias
73
RESPOSTAS BIOQUÍMICAS ASSOCIADAS À DEFESA EM MELANCIA
CONTRA MELOIDOGYNE ENTEROLOBII
Paloma C. da C Lubarinoa,*, Leonardo Sousa Cavalcantia, Pedro M. Ribeiro Juniorb
e Rita de Cássia S. Diasb
a
Universidade Federal do Vale do São Francisco, 56.304-205, Petrolina, Pernambuco,
Brasil
b
Embrapa Semiárido, BR 428, Km 152, Zona Rural, 56302-970, Petrolina, Pernambuco,
Brasil
-----------------------------------------------------------------------------------------------------------( ) Manuscrito com material suplementar
(X) Manuscrito sem material suplementar
-----------------------------------------------------------------------------------------------------------*e-mail: [email protected]
BIOCHEMICAL RESPONSES ASSOCIATED WITH DEFENSE IN
WATERMELON AGAINST MELOIDOGYNE ENTEROLOBII
Este estudo objetivou investigar a reação de linhas de melancia resistente (BGCIA 240) e
suscetível (BGCIA 875) ao Meloidogyne enterolobii, assim como avaliar atividade de
enzimas relacionadas à defesa e os teores de clorofilas nessas plantas. As linhas foram
inoculadas com o nematoide e avaliadas quanto ao número de galhas (NG) e número de
ovos por planta (NO), comprimento (CR) e massa fresca de raiz (MFR), os teores de
clorofila e à atividade de enzimas relacionadas à defesa, guaiacol peroxidase (POX),
polifenoloxidases (PPO), ascorbato peroxidase (APX), catalase (CAT) e fenilalanina
amônia-liase (PAL). Para as análises, foram coletadas amostras de raízes aos 7, 14, 21, 28
e 35 dias após a inoculação (DAI). A presença do nematoide induziu na linha resistente
maior valor CR aos 28 DAI e menor MFR aos 21 DAI. A linha suscetível também
apresentou menor MFR, porém maior formação de galhas e maior NO. Aos 14 DAI,
houve maior atividade de POX, APX, PPO e PAL na linha resistente inoculada. Estes
resultados sugerem que uma combinação destas respostas de defesa na melancia podem
74
contribuir para a seleção de genótipos em programas de melhoramento visando
resistência da melancia contra M. enterolobii.
Keywords: nematoide das galhas, Citrullus spp., mecanismos de defesa.
This study aimed to investigate the reaction of resistant (BGCIA 240) and susceptible
(BGCIA 875) watermelon lines to Meloidogyne enterolobii, and to evaluate the activity
of enzymes related to the defense and the content of chlorophyll. The lines were
inoculated with the pathogen and evaluated for reaction by the number of galls (NG) and
number of eggs per plant (NO), length (CR) and fresh weight of roots (MFR) and
analyzed for chlorophyll content and activity of defense-related enzymes, guaiacol
peroxidase (POX), polyphenol oxidases (PPO), ascorbate peroxidase (APX), catalase
(CAT) and phenylalanine ammonia lyase (PAL). For analysis, samples were collected
from roots 7, 14, 21, 28 and 35 days after inoculation (DAI). The pathogen induced in the
resistant line highest value CR at 28 DAI and lower MFR at 21 DAI. The susceptible line
also showed less MFR, but most galling and increased NO. At 14 days, a higher enzyme
activity in the resistant line inoculated for POX, APX, PPO and PAL. These results
suggest that a combination of these defense responses in watermelon can help in the
selection of genotypes in breeding programs for watermelon resistance against M.
enterolobii attack.
Keywords: root-knot nematodes, Citrullus spp., defense mechanism.
75
INTRODUÇÃO
Os nematóides de galhas (Meloidogyne spp.) estão entre os patógenos mais comuns
que limitam a qualidade e a produção das Cucurbitáceas As espécies desse gênero de
maior ocorrência no Brasil são M. incognita (Kofoid & White) Chitwood, M. javanica
(Treub) Chitwood e M. arenaria (Neal) (PEREIRA et al., 2010) Contudo, no Submédio
do Vale do São Francisco a espécie M. enterolobii (sin. M. mayaguensis) vêm, nos
últimos anos, causando prejuízos aos produtores de cucurbitáceas e de outras culturas
(TERAO et al., 2010) O controle desses nematoides nas cucurbitáceas é difícil devido a
ampla gama de hospedeiras e não existem nematicidas registrados no Brasil para o seu
controle (AGROFIT, 2015). Ademais, os nematicidas químicos apresentam alta toxidez
ao homem e ao ambiente e tem alto custo aos produtores. Nesse contexto, outras
estratégias devem ser adotadas no controle destes fitopatógenos na cultura da melancia,
como a resistência genética que é a forma de controle mais econômica para o produtor e
mais sustentável para o ambiente (LIMA et al., 2005)
Alguns estudos vêm sendo realizados visando à identificação de linhas de
melancia e outras cucurbitáceas com resistência a Meloidogyne spp. Experimentos
mostraram que linhas de Cucurbitáceas como a bucha, abóbora Goianinha, abóbora Mini
Paulista, melão Redondo Amarelo e melancia Charleston Gray foram classificadas como
resistentes a M. Incognita (FRANCO et al., 2008) Melancias forrageiras (Citrullus
lanatus var. citroides) tem sido relatadas como fonte de resistência contra M. Enterolobii
(DAMACENO, 2012; CASTRO et al., 2011) e podem ser utilizadas tanto como portaenxerto quanto fontes de genes de resistencia para cruzamentos com melancia de mesa
(Citrullus lanatus (Thumb.) Matsum & Nakai). Apesar destes relatos, existem poucos
trabalhos apresentando estudos de mecanismos bioquímicos relacionados à defesa dessas
linhas resistentes quando inoculadas com nematoides do gênero Meloidogyne.
Ao reconhecer o patógeno, em plantas resistentes ocorre o aumento dos níveis de
defesa pela ativação de genes que codificam diversos compostos de defesa como a
explosão oxidativa, enzimas antioxidantes, proteínas relacionadas à patogênese e enzimas
precursoras de fitoalexinas como a fenilalanina amônia-liase (PAL) (KOVÁCIK et al,
2007).
76
Estudos da associação da expressão destes compostos de defesa e de genótipos
resistentes a nematois em outras olerícolas foram realizados. Na família das Solanáceas, o
tomate e batata tem mostrado associação da enzima PAL como um fator de resistência a
patógenos (CAM; TOWRES, 1973; BRUESKE, 1980; KALAIARASAN, 2009). Em
outro estudo foi observado em raízes de tomate resistentes e inoculadas com M. incognita
menor número de galhas por planta e um aumento significativo na atividade da PAL às
108 horas após a inoculação, superiores à testemunha suscetível que apresentaram menor
atividade de dessa enzima (BRUESKE, 1980; KALAIARASAN, 2009). Em raízes de
batatas resistentes e inoculadas com o nematóide Globodera rostochiensis foram
observadas maiores atividades da PAL do que em raízes de plantas suscetíveis (GIEBEL,
1973).
Diante do exposto, este estudo objetivou investigar a reação de uma linha de
melancia forrageira resistente e uma linha de melancia de mesa suscetível ao M.
enterolobii. Objetivou também avaliar a atividade de enzimas relacionadas à defesa e os
teores de clorofilas nesses genótipos inoculados com M. enterolobii.
PARTE EXPERIMENTAL
Material vegetal
Os experimentos foram realizados na Embrapa Semiárido com genótipos de
melancia provenientes do Banco Ativo de Germoplasma (BAG) de Cucurbitáceas dessa
Unidade, sendo uma Linha BGCIA240 de melancia forrageira (C. lanatus var. citroides)
resistente a M. enterolobii e uma Linha BGCIA875 de melancia de mesa (C. lanatus var.
lanatus), suscetível ao patógeno. Para a produção das mudas, as sementes foram
plantadas em copos descartáveis de polipropileno (uma semente por copo) contendo 500
mL de solo autoclavado.
Preparo do inóculo e inoculação
Para a produção e manutenção do inóculo, foram coletadas fêmeas de uma
população de M. enterolobii de raízes de goiabeira (Psidium guajava L. cultivar. Paluma)
77
no município de Petrolina, PE, que foram previamente identificadas utilizando-se o
padrão de isoenzima (CARNEIRO; ALMEIDA, 2001). O inóculo foi multiplicado em
mudas de tomateiro Santa Clara. Para o preparo do inóculo, os ovos foram extraídos das
raízes das plantas de tomateiro infectadas de acordo com a metodologia proposta por
Hussey e Barker (HUSSEY; BARKER, 1973). A inoculação foi realizada 12 dias após a
germinação das mudas, com suspensão de 500 ovos de M. enterolobii por planta,
utilizando-se 1 mL da suspensão distribuídos no vaso ao redor do colo das plantas.
Reação dos genótipos de melancia de mesa e forrageira ao Meloidogyne enterolobii
Para a avaliação da reação dos genótipos de melancia ao nematoide, foram
realizadas avaliações aos 7, 14, 21, 28 e 35 dias após a inoculação do número de galhas
(NG) e número de ovos por planta (NO) (COOLEN; D’HERDE, 1972). Além disso,
avaliou-se também o comprimento de raiz (CR)e massa fresca de raiz (MFR). Para a
realização das análises, as raízes das plantas foram retiradas dos recipientes lavadas com
água.
Análises de clorofila e enzimáticas
Para a avaliação das clorofilas e enzimática, foi realizado outro experimento nas
mesmas condições do experimento anterior.
Antes das coletas das raízes, foram realizadas as avaliações das clorofilas a e b
(IFC - Índice Falker de Clorofila) das folhas utilizando-se o clorofilômetro marca
ClorofiLOG® modelo CFL 1030. Para tanto, foram realizadas três leituras com o
clorofilômetro, sempre na primeira folha completamente expandida (do topo do dossel
para a base) aos 14, 21, 28 e 35 dias após a inoculação (DAI).
Para as análises enzimáticas, foram realizadas coletas nos mesmos tempos citados
anteriormente, onde as raízes dos genótipos foram lavadas cuidadosamente com água
corrente. Posteriormente, as amostras radiculares foram envolvidas em papel alumínio,
identificadas, mergulhadas em nitrogênio líquido e armazenadas em freezer (-80 °C) até o
momento das análises. Para a obtenção do extrato protéico, tecidos radiculares foram
triturados em nitrogênio líquido, com almofariz e pistilo, até a obtenção de um pó fino.
78
Em seguida, foi adicionado tampão fosfato de potássio 100 mM pH 7,0 (3 mL de tampão
por grama de material triturado) e homogeneizou-se com o pistilo por 2 min. Após esse
processo, a suspensão foi centrifugada a 12.000 g por 15minutos (4 oC) e o sobrenadante
foi utilizado como fonte enzimática.
As proteínas solúveis totais contidas nos extratos foram quantificadas com base no
ensaio de Bradford (1976) utilizando-se uma curva padrão de albumina sérica bovina
(BSA).
A atividade do guaiacol peroxidase (POX) foi determinada pelo aumento na
absorbância a 470 nm durante 2 min em um meio de reação contendo fosfato de potássio
100 mM (pH 7,0), guaiacol 50 Mm e H2O2 150 mM (URBANEK et al., 1991).
A atividade da catalase (CAT) foi estimada pelo decréscimo na absorbância a 240
nm durante 3 min em um meio de reação contendo fosfato de potássio 100 mM (pH 7,0)
e H2O2 250 mM (AZEVEDO et al.,1998).
A atividade do ascorbato peroxidase (APX) foi medida pelo decréscimo na
absorbância a 290 nm durante 3 min em um meio de reação contendo fosfato de potássio
100 mM (pH 7,0), ácido ascórbico 10 Mm e H2O2 2 mM (NAKANO; ASADA, 1981).
A atividade de polifenoloxidase (PPO) foi estimada pelo aumento na absorbância
a 420 nm durante 3 min em um meio de reação contendo fosfato de potássio 100 mM (pH
6,5) e pirogalol 100 mM (GAUILLARD et al., 1993). A reação ocorreu pela
transformação do pirogalol (fenol) em quinona.
A atividade da enzima fenilalanina amônia liase (PAL) foi ensaiada através da
reação entre 90 μL de tampão 100 mM Tris-HCl (pH 8,8), 50 μL de 40 mM de
fenilalanina e 50 μL do extrato enzimático (MORI et al.,2002).
Delineamento experimental e análise estatística
Os delineamentos experimentais utilizados nos experimentos foram o inteiramente
casualizado, com três repetições e parcela experimental composta por três plantas. Os
dados foram submetidos à análise de variância utilizando-se o programa Sisvar
(FERREIRA, 2006). As médias do número de galhas e número de ovos por planta e fator
foram comparadas pelo teste T a 5% de probabilidade. As médias do comprimento, massa
fresca de raiz e clorofilas a e b foram comparadas pelo teste de Tukey 5% de
79
probabilidade. Para melhor visualização dos resultados das análises enzimáticas ao longo
do tempo, plotou-se a curva de progresso da atividade das enzimas com os respectivos
erros padrões.
RESULTADOS E DISCUSSÃO
Reação de genótipos de melancia a Meloidogyne enterolobii
Plantas da linha de melancia forrageira BGCIA 240, considerada resistente, e da
linha melancia de mesa BGCIA 875, considerada suscetível, apresentaram ao longo das
avaliações número de galhas por planta semelhantes estatisticamente (Figura 1A).
Entretanto, aos 35 dias após a inoculação com M. enterolobii, a linha suscetível
apresentou 87,5% mais galhas que a linha suscetível. Segundo Taylor e Sasser (1978)
uma planta que apresenta índice de galhas entre 31 a 100, é considerada altamente
suscetível.
Dos 7 aos 21 dias após a inoculação, não observou-se efeito das linhas para o
número de ovos por planta. Aos 35 dias após a inoculação, plantas suscetíveis
apresentaram maior número de ovos por planta do que plantas da linha resistente,
diferindo estatisticamente. Observou-se nas plantas suscetíveis 8,08 vezes mais ovos que
nas plantas resistentes (Figura 1B). O maior número de ovos observados na linha BGCIA
875 indica seu estado de suscetibilidade. Por outro lado, mesmo havendo formação de
galhas em raízes da linha BGCIA 240, observou-se uma baixa produção de ovos de M.
enterolobii. Embora as plantas resistentes sejam invadidas pelos nematóides, o
parasitismo não é bem sucedido devido a mecanismos de resistência das plantas
(TRUDGIL; BLOK, 2001). Assim, plantas resistentes podem reconhecer substâncias
liberadas pelo patógeno e desencadear respostas de defesa contra o invasor (FRAGOSO
et al., 2007).
Para comprimento de raiz observa-se que a inoculação não afetou o crescimento
da mesma, não havendo diferenças significativas em cada linha de melancia inoculada e
não inoculada. Em média, a linha de melancia forrageira apresentou maior comprimento
do que a melancia de mesa (Tabela 1). O comprimento da raiz pode estar relacionado
com estratégias de sobrevivência desenvolvida pelas variedades de melancias forrageiras.
80
A melancia forrageira adaptou-se bem às condições semiáridas, assim como a estresse
hídrico, nutricional ou ataque de patógenos do solo (QUEIROZ, 1993; ROMÃO, 1995).
Com isso, supõem-se que a medida que as galhas foram se desenvolvendo, as raízes
secundárias ficaram impedidas de absorver nutrientes induzindo o alongamento axial da
raiz. O mesmo não foi observado para a linha BGCIA 875, talvez por se tratar de uma
variedade comercial suscetível. Na avaliação da massa fresca de raiz, foi observado
comportamento semelhante ao do comprimento de raiz. Não foi observado efeito
significativo da inoculação na massa fresca das raízes em cada linha de melancia (Tabela
1). Este resultado provavelmente está associado com a formação das galhas nas raízes das
plantas inoculadas aumentando a massa nas plantas inoculadas. Em média, a massa fresca
foi maior na linha de melancia forrageira
Afim de compreende o mecanismo de infecção de M. javanica em plantas de soja,
foi demonstrado que a superexpressão do gene de corismato mutase I do fitonematoide
altera a via xiquimato por causa da degradação do corismato citoplasmático, que resulta
num fenótipo de redução ou aborto das radícolas laterais, provavelmente como
conseqüência da redução dos níveis de auxinas nos plastídios (FRAGOSO et al., 2007).
Assim observou-se para a linha BGCIA 240 uma recuperação do sistema radicular aos 28
e 35 DIA e para a linha BGCIA 875 uma contínua queda dos valores de massa fresca de
raiz.
Os dados de clorofila a e b para as épocas estudadas não diferiram
estatisticamente entre as plantas inoculadas e não inoculadas, nem entre os genótipos
(Tabela 1). Provavelmente, esse fato ocorreu devido ao pouco tempo após a inoculação.
Os sintomas reflexos na parte aérea da planta pela presença do nematoide na raiz
aparecem num estágio mais avançado da infecção e na fase de produção, caracterizando
um maior dreno de nutrientes para os frutos. Por sua vez, Lubarino (2012) ao estudar a
relação do teor de clorofila com a ação do M. enterolobii em plantas inoculadas de
melancia observou aos 60 dias após a inoculação um menor teor de clorofila em plantas
inoculadas com relação as não inoculadas. Em geral, o índice médio de clorofila a foi
maior que o de clorofila b, que ocorre devido a uma maior proporção do fotossistema I
que é mais rico em clorofila a, em condições normais de luminosidade, (CRIRCHELEY,
1999) e menor clorofila b, uma vez que, esta tem uma maior proporção em ambientes
sombreados (SOUZA et al., 2011).
81
A
N° de galhas/planta
50
ns
40
30
20
ns
10
ns
0
7
B
ns
ns
14
21
35
Dias após inoculação
400
BGCIA240R+
N° de ovos/planta
28
BGCIA 875+
*
300
200
100
ns
ns
ns
ns
14
21
28
0
7
35
Dias após inoculação
BGCIA240
BGCIA 875
Figura 1. Número de galhas (A) e número de ovos (B) por planta de dois genótipos de
melancia do Banco Ativo de Germoplasma (BAG) de Cucurbitáceas da Embrapa
Semiárido, Linha BGCIA 240 (resistente) e Linha BGCIA 875(suscetível) inoculadas com
Meloidogyne enterolobii, avaliadas em diferentes épocas de coleta. * Significativo a 5%
de probabilidade e ns= não significativo pelo teste T.
Tabela 1. Comprimento de raiz, massa fresca de raiz, clorofila a e clorofila b de dois
genótipos de melancia do Banco Ativo de Germoplasma de Cucurbitáceas da Embrapa
Semiárido, Linha BGCIA 240 e Linha BGCIA 875 inoculadas e não inoculadas com
Meloidogyne enterolobii em diferentes épocas de avaliação.
Linhas
BGCIA 240+
BGCIA 240
BGCIA 875+
BGCIA 875
7
16,25 a
17,00 a
10,25 b
10,73 b
7
Comprimento de raiz (cm)
Dias após inoculação
14
21
28
12,90 a
14,30 ab
21,75 a
15,50 a
16,17 a
13,40 b
10,50 a
10,50 b
9,83 b
10,87 a
10,83 b
11,80 b
Massa fresca de raiz (g)
14
21
28
35
21,17 a
9,33 a
12,25 a
11,75 a
35
82
BGCIA 240+
BGCIA 240
BGCIA 875+
BGCIA 875
2,66 a
2,54 ab
0,61 b
0,99 ab
1,78 a
1,85 a
0,63 a
0,69 a
1,46 ab
1,61 a
1,73 a
2,22 a
1,24 a
1,14 a
0,63 b
0,29 b
0,60 a
1,00 ab
0,35 b
1,11 a
Clorofila a (IFC)
7
14
21
28
35
BGCIA 240+
24,90 a
32,07 a
31,80 a
28,03 a
24,57 b
BGCIA 240
26,00 a
33,90 a
34,47 a
27,73 a
28,93 ab
BGCIA 875+
24,10 a
28,90 a
31,80 a
28,40 a
28,57 ab
BGCIA 875
26,33 a
30,40 a
36,17 a
30,03 a
30,63 a
Clorofila b (IFC)
7
14
21
28
35
BGCIA 240+
9,33 a
14,67 a
15,37 a
14,90 a
10,83 a
BGCIA 240
12,90 a
15,07 a
16,83 a
13,33 a
11,07 a
BGCIA 875+
12,33 a
11,87 a
12,67 a
12,10 a
9,27 a
BGCIA 875
12,53 a
13,27 a
18,73 a
13,40 a
11,97 a
+= genótipos inoculados; IFC= Índice Falker de Clorofila. Médias com mesma letra nas
colunas não diferem entre si de acordo com o teste de Tukey (P < 0,05 ).
Respostas bioquímicas de defesa
As proteínas presentes na planta podem estar envolvidas no processo de defesa
bioquímica contra os danos causados pelo patógeno (BERNARDS et al., 1999). Além
disso, espécies vegetais sensíveis ao nematoide das galhas podem apresentar a quantidade
ou qualidade das proteínas alteradas. Não foi obeservado efeito da inoculação com o M.
enterolobii nos teores de proteínas solúveis totais nas duas linhas estudadas nos tempos
avaliados, exceto aos 35 dias após a inoculação que o genótipo suscetível (BGCIA 875)
inoculado apresentou menor teor de proteínas que os não inoculado, época de maior
formação de galhas e número de ovos observado neste genótipo (Figura 1- A).
Para a guaiacol peroxidase, foi observado que não ocorreram diferenças na
atividade desta enzima em plantas resistentes e suscetíveis infectadas e não infectadas
com o M. enterolobii até os 14 dias após a inoculação (Figura 1-B). A linha com maior
nível de resistência (BGCIA 240) inoculada com o nematoide apresentou maior atividade
dessa enzima, especialmente nos tempos de 21 e 28 dias após a inoculação, enquanto que
a não inoculada apresentou atividade semelhante à da linha suscetível BGCIA875
inoculada e não inoculada. No presente estudo, foi observado nas plantas resistentes
aumento da atividade da enzima guaiacol peroxidase somente aos 21 DAI, o que significa
83
dizer que nesse momento há uma interação de reconhecimento planta-patógeno,
desencadeando a resposta de defesa da planta contra os danos causados pelo nematóide.
A atividade da polifenoloxidase foi maior nas plantas da linha resistente, tanto
para plantas inoculadas como para não inoculadas somente aos 14 dias após a inoculação
(Figura 1C). Não há relatos que correlacionem o aumento da atividade de
polifenoloxidases com a defesa DAE plantas de melacia aos nematoides. Entretanto, para
outras culturas como a soja estudos têm mostrado que plântulas de soja de variedade
resistente e inoculadas com J2 de M. javanica está associada a um aumento na atividade
da PPO 72 horas após a inoculação (SHIMIZU, 2004).
A atividade da ascorbato peroxidase foi maior nas plantas da linha resistente
(BGCIA 240), tanto inoculada como não inoculada com M. enterolobii, até os 21 dias
após a inoculação, superiores à linha suscetível (Figura 1D). Não foi observado diferença
nas atividades da catalase entre as linhas suscetíveis (BGCIA 875) e resistentes de
melancia.
Para a atividade da fenilalanina amônia-liase , aos 14 dias após a inoculação, foi
observado aumento significativo nas raízes de plantas da linha BGCIA 240, inoculadas,
em relação às plantas da linha suscetível, BGCIA 875, também inoculadas (Figura 2F).
Este aumento de atividade provavelmente reflete o aumento da utilização de compostos
fenólicos em raízes resistentes. A partir desse momento houve queda na atividade dessa
enzima para as plantas da linha BGCIA 240 inoculadas e não inoculadas. Kalaiarasan
(2009) ao avaliar genótipos de tomate resistentes e suscetíveis a M. incognita quanto aos
danos provocados pelo patógeno observou por avaliação bioquímica que os genótipos
resistentes possuíam maior atividade de fenilalanina amônia-liase que os genótipos
suscetíveis.
Os resultados deste trabalho possibilitam um maior conhecimento dos
mecanismos de defesa envolvidos na resistência da melancia ao nematoide M.
enterolobii. Entre as respostas bioquímicas na defesa das plantas contra patógenos, as
peroxidases, catalase, polifenoloxidase e fenilalanina amônia-liase têm recebido destaque
em diversos estudos (KALAIARASAN, 2009) pois estes compostos podem atuar na
mitigação dos danos causados pelo patógeno, ou impedir o avanço do mesmo nos tecidos
do hospedeiro (RESENDE et al., 2003). Estas enzimas podem atuar de forma diferente
em diferentes relações planta-nematóide e, neste trabalho, observou-se que o aumento na
84
atividade das enzimas peroxidases, polifenoloxidase e fenilalanina amônia-liase indica
um efeito de resposta da planta após o reconhecimento do patógeno.
D
A
200
APX (mM min-1 mg P-1 )
0,80
Proteína (mg mL-1 )
0,70
0,60
0,50
0,40
0,30
0,20
0,10
160
120
80
40
0,00
0
7
14
21
28
35
7
14
Dias após inoculação
28
BGCIA240+
BGCIA240
BGCIA240+
BGCIA240
BGCIA875+
BGCIA875
BGCIA875+
BGCIA875
B
35
E
40000
4,00
3,50
CAT (mM min -1 mg P-1 )
POX (mM min -1 mg P-1 )
21
Dias após inoculação
30000
20000
10000
3,00
2,50
2,00
1,50
1,00
0,50
0
0,00
7
14
21
28
35
7
14
21
Dias após inoculação
28
35
Dias após inoculação
BGCIA240+
BGCIA240
BGCIA240R+
BGCIA240R
BGCIA875+
BGCIA875
BGCIA875+
BGCIA875
C
F
0,25
PAL (mM min-1 mg P-1 )
PPO (mM min -1 mg P-1 )
20
16
12
8
0,20
0,15
0,10
0,05
0,00
4
7
14
21
28
BGCIA875+
7
14
21
28
Dias após inoculação
Dias após inoculação
BGCIA240+
35
BGCIA240
BGCIA240R+
BGCIA240R
BGCIA875
BGCIA875+
BGCIA875
35
85
Figura 2. Proteínas solúveis totais (A), atividades das enzimas guaiacol peroxidase
(POX) (B), polifenoloxidase (PPO) (C), ascorbato peroxidase (APX) (D), catalase
(CAT)(E) e fenilalanina amônia liase (PAL) (F) de dois genótipos de melancia do Banco
Ativo de Germoplasma (BAG) de Cucurbitáceas da Embrapa Semiárido, Linha BGCIA
240 e Linha BGCIA 875 inoculadas com Meloidogyne enterolobii, e seus controles,
avaliadas em diferentes épocas de coleta. Barras representam erro padrão da média
CONCLUSÃO
Este estudo demonstrou, em alguns casos, que a presença do patógeno induziu na
linha resistente inoculada com M. enterolobii, maior valor de comprimento de raiz aos 28
DAI e menor massa fresca de raiz aos 21 DAI. A massa fresca também foi menor para a
linha suscetível inoculada, apresentando-se mais acentuado, por estar associado com a
formação das galhas nessas raízes.
Nas análises bioquímicas a maior concentração de compostos de defesa na linha
resistente inoculada foi aos 14 DAI. As enzimas relacionadas a defesa da melancia a M.
enterolobii nesse trabalho foi a POX, APX, PPO e PAL. Desta forma nota-se que estes
compostos são potenciais para serem utilizados como marcadores bioquímicos de
resistência a M. enterolobii em melancia.
AGRADECIMENTOS
À Embrapa Semiárido pela utilização das instalações (casa de vegetação e
laboratórios) e ao Programa de Pós-graduação em Recursos Naturais do Semiárido da
Universidade Federal do Vale do São Francisco pelo suporte financeiro.
REFERÊNCIAS
1. Pereira F. M., Souza, R. M., Souza, P. M., Dolinski, C.; Santos, G. K. Estimativa do
impacto econômico e social direto de Meloidogyne mayaguensis na cultura da goiaba no
Brasil. Soc. Bras. Nematol., 2009, 33, 176–181.
2.
Terao
et
al.,
Sistema
de
produção
da
melancia.
http://sistemasdeproducao.cnptia.embrapa.br. acessada em 29 set. 2014.
2010:
86
3. Agrofit: http://www.agrofit.com.br/novoportal/. acessada em 26 jan 2015.
4. Lima, G.S.A.; Assunção, I.P.; Valle, L.A.C. Controle genético de doenças radiculares.
Em Michereff, S.J.; Domingos, E.G.T.; Andrade, M.M. Ecologia e manejo de patógenos
radiculares em solos tropicais. Recife: UFRP, 2005, 248-272.
5. Franco, A.J.; Charlo, H.C.O.; Galatti, F.S.; Braz, L.T. Avaliação de genótipos de
cucurbitáceas quanto à resistência à Meloidogyne incognita. Resumos do 48° Congresso
Brasileiro de Olericultura, Maringá, Paraná, Brasil, 2008.
6. Damaceno, L.S. Reação de genitores de melancia a Meloidogyne enterolobii.
Dissertação de Mestrado. Universidade do Estado da Bahia, Brasil. 2012.
7. Castro, J. M. C; Dias, R. C. S; Teixeira, F. A; Damaceno, L. S; Barbosa, G. S. Reação
de genótipos de melancia à nematoides. Resumos do 51° Congresso Brasileiro de
Olericultura, Viçosa, Minas Gerais, Brasil, 2011.
8. Kovácik, J.; Klejdus, B.; Backor, M.; Repcak, M. Phenylalanine ammonia-lyase
activity and phenolic compounds accumulation in nitrogen-deficient Matricaria
chamomilla leaf rosettes. Plant Science 2007, 172, 393-399.
9. Camm, E. L.; Towers, G.H. Review article-Phenylalanine ammonia lyase.
Phylochemistry 1973, 12, 961-973.
10. Brueske, C. H. Phenylalanine ammonia-lyase activity in tomato roots infected and
resistant to the root knot nematode, Meloidogyne incognita. Physiologycal Plant
Pathology 1980, 16, 409-414.
11. Kalaiarasan, P. Biochemical markers for identification of root knot nematode
(Meloidogyne incognita) resistance in tomato. Karnataka Journal of Agricultural
Sciences 2009, 22, 471-475.
87
12. Giebel, J. Phenylalanine and tyrosine ammonia-lyase activities in potato roots and
their significance in potato resistance to Heterodera rostochiensis. Nematologica 1973,
19, 1-6.
13. Carneiro, R.M.D.G; Moreira, W. A; Almeida, M. R A. Primeiro registro de
Meloidogyne mayaguensis em goiabeira no Brasil. Soc. Bras. Nematol., Brasília, 2001,
25, 223-228.
14. Hussey, R. S.; Barker, K. R. A comparison of methods of collecting inoculated of
Meloidogyne spp., including a new technique. Plant Dis. Rep. 1973, 57, 1025-1028.
15. Coolen, W. A.; D’Herde, C. J. Method for the quantitative extraction of nematode
plant tissue. Ghent State Agriculture Research Center, Ministry of Agriculture 1972, 777.
16. Bradford, M. M. A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976,
72, 248-254.
17. Urbanek, H., E. Kuzniak-Gebarowska; K. Herka. Elicitation of defence responses in
bean leaves by Botrytis cinereapolygalacturonase. Acta Physiol. Plant. 1991,13, 43-50.
18. Azevedo, R. A.; Alas, R. J.; Smith, R. J.; Lea, P. J. Response of antioxidant enzymes
to transfer from elevated carbon dioxide to air ozone fumigation, in the leaves and roots
of wild-tye and a catalase-deficient mutant of barley. Physiol. Plant. 1998, 104, 280-292.
19. Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascobate-specific
peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867-880.
20. Gauillard, F.; Forget, F. R.; Nicolas, J. New spectrophotometric assay for polyphenol
oxidase activity. Anal. Biochem. 1993, 215, 59-65.
88
21. Mori, T.; Sakurai, M.; Sakuta, M. E. Vects of conditioned medium on activities of
PAL, CHS, DAHP synthase (DS-Co and DS-Mn) and anthocyanin production in
suspension cultures of Fragaria ananassa. Plant Science 2001, 160, 355-360.
22. Ferreira, D. F. Análises estatísticas por meio do SISVAR (Sistema para análise de
variância) para Windows versão 4.0. Reunião anual da Região Brasileira da Sociedade
Internacional de Biometria, Universidade Federal de São Carlos, Brasil, 2000.
23. Taylor, A. L.; Sasser, J.N. Biology, identification and control of root-knot nematodes,
Meloidogyne species. Raleigh: North Carolina State University Graphics, 1978, 111.
24. Trudgill, D. L.; Blok, V. C. Apomictic, polyphagous root-knot nematodes:
exceptionally successful and damaging biotrophic root pathogens. Annu. Rev.
Phytopathol., 2001, 39, 53-77.
25. Fragoso, F.R.; Lourenço, I.T.; Viana, A.M.B.; Souza, D.S.L.; Andrade, R.V.; Mehta,
A.; Brasileiro, A.C.M.; Pinto, E.R.C.; Lima, L.M.; Rocha, T.L.; Sá, M.F.G. Interação
Molecular Planta-Nematóide. Documentos 198, Embrapa: Brasília, 2007, 56.
26. Queiróz, M.A. de. Potencial do germoplasma de Cucurbitáceas no Nordeste
brasileiro. Hortic. Bras., 1993, 11, 7-9.
27. Romão, R.L. Dinâmica evolutiva e variabilidade de populações de melancia Citrullus
lanatus (Thunb.) Matsum & Nakai em três regiões do Nordeste brasileiro. Dissertação de
Mestrado: USP- ESALQ, Piracicaba, 1995.
28. LUBARINO, P. C. da C. Caracterização e avaliação de genótipos de Citrullus lanatus
L. quanto à resistência a nematoide das galhas, Meloidogyne enterolobii. Trabalho de
Conclusão de Curso apresentado ao Departamento de Tecnologia e Ciências Sociais, da
Universidade do Estado da Bahia, 2012.
89
29. Critchley, C. Concepts in photobiology: photosinthesis and photomorphogenesis.
Singhal, G.S., Renger, G., Sopory, S. K., Irrgang, K. D., Govindjee, eds.; New Delhi:
Narosa publishing House, 1999, 587.
30. Souza, G. S.; Silva, J. S.; Santos, A. R.; Gomes, D. G.; Oliveira, U. C. Crescimento e
produção de pigmentos fotossintéticos em alfavaca cultivada sob malhas coloridas e
adubação fosfatada. Enciclopédia Bioesfera, Goiânia, 2011, 7.
31. Bernards, M. A.; Fleming, D. W.; Llewellyn, D. B.; Priefer, R.; Yang, X.; Sabatino,
A.; Plourde, G. L. Biochemical characterization of the suberisation-associated anionic
peroxidase of potato. Plant Physiology, Bethesda, 1999, 121, 135-146.
32. Shimizu, M. M. Polifenoloxidase como fator de resistência da soja a nematóides e na
oxidação do palmito. Tese de Doutorado. Universidade Estadual de Campinas,
UNICAMP, Brasil. 2004.
33. Resende, M. L. V.; Salgado, S. M. L.; Chaves, Z. M. Espécies ativas de oxigênio na
resposta de defesa de plantas a patógenos. Fitopatol. bras., Brasília, 2003, 28.