Regulação Neuro-humoral da Motilidade
No estudo da motilidade do aparelho digestivo devemos
considerar a inervação intrínseca e a musculatura lisa como
uma única estrutura funcional.
Como os músculos lisos do aparelho digestivo apresentam
diferentes propriedades eletrofisiológicas além de responderem
diferentemente a fármacos, neurotransmissores e hormônios,
será útil separa-los em duas categorias principais:
1. Músculos lisos quiescentes: não apresentam atividade
contrátil espontânea, e entre eles temos o músculo liso do
corpo do esôfago, do fundo gástrico e da vesícula biliar.
2. Músculos lisos com atividade contrátil espontânea: estes
músculos podem gerar sua própria contração, por isso essa
contração e dita de origem miogênica. Como exemplos desse tipo
de músculo liso podemos citar o esfíncter esofágico inferior,
musculatura do corpo, antro e piloro gástricos, dos intestinos
e do esfíncter anal interno.
Do
ponto
de
vista
eletrofisiológico,
a
principal
diferença entre os músculos lisos quiescentes e os de
contração espontânea é que os últimos apresentam a geração dos
potenciais (não confundir com potencial de ação) oscilatórios
conhecidos como ritmo elétrico básico, ou ondas lentas
(qualquer dúvida, consultar o texto sobre músculo liso).
Toda vez que o potencial oscilatório atinge o limiar de
disparo surgem os potenciais de ação e a correspondente
contração muscular.
Outro dado importante é que quando se diz que a
musculatura lisa do intestino, por exemplo, é do tipo que
apresenta contração espontânea, isto não significa que a
inervação excitatória esteja ausente. Embora a musculatura
intestinal produza ondas lentas, recebe também inervação
colinérgica excitatória ao longo de todo o tubo. Esta
inervação pode participar ativamente no processo contrátil.
Controle Nervoso da Motilidade
É importante lembrar que cada parte do tubo digestivo se
apresenta perfeitamente ajustada a sua função, isto leva a uma
heterogeneidade
nas
características
de
cada
segmento
principal,
bem
como
compartimentalização.
Esta
compartimentalização é mantida graças a estruturas que
garantem o isolamento de cada setor do tubo. Essas estruturas
são os esfíncteres, segmentos musculares do tubo digestivo que
se mantém contraídos formando zonas de alta pressão, que
impedem tanto o refluxo de fluidos, bem como garantem que o
trânsito de um segmento para outro ocorra de forma ordenada.
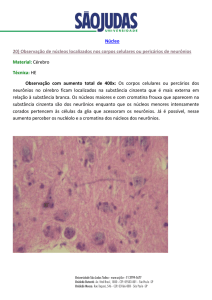
Sistema Nervoso Entérico
O controle nervoso da motilidade se dá através das
inervações extrínsecas e intrínsecas. O sistema nervoso
autônomo
(SNA)
na
suas
duas
divisões,
simpática
e
parassimpática, exerce importante papel na motilidade, porém
toda atividade muscular lisa do tubo digestivo, é mediada
pelos plexos mioentérico e submucoso.
Os plexos intrínsecos apresentam características próprias
que permitem que estes sejam considerados como um sistema
nervoso estruturado para coordenar e programar padrões de
motilidade, mesmo na ausência do sistema nervoso central.
Enquanto o plexo submucoso tem, a sua maior influência nos
processos de transporte através da mucosa dos intestinos, o
plexo mioentérico exerce controle da motilidade. Interconexões
entre os dois plexos seriam responsáveis pela coordenação dos
dois processos. Embora na musculatura estriada do trato
digestivo no terço superior do esôfago a inervação pelo
sistema nervoso parassimpático se dê diretamente sobre as
fibras musculares, na musculatura lisa do esôfago isto não
ocorre. A inervação extrínseca faz sinapse com neurônios do
sistema nervoso entérico, que atua nas células do músculo
liso.
Um fato importante é que o sistema nervoso entérico
apresenta características tanto anátomo-histológicas quanto
funcionais típicas de um sistema nervoso central ("minicérebro").
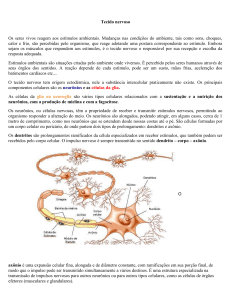
Os gânglios mioentéricos contêm motoneurônios, neurônios
sensoriais
e
interneurônios.
A
maioria
dos
neurônios
mioentéricos
são
neurônios
motores
para
as
camadas
longitudinal e circular. Os motoneurônios excitatórios liberam
acetilcolina para as células do músculo liso. Também podem
liberar a substância P. Já os motoneurônios inibitórios
liberam VIP, e possivelmente óxido nítrico. Outros neurônios
mioentéricos fazem sinapse com neurônios do plexo submucoso.
Um terço dos neurônios mioentéricos são neurônios sensoriais.
Os motoneurônios apresentam receptores nicotínicos, enquanto o
músculo liso receptores muscarínicos.
A maioria dos neurônios do plexo submucoso regula a
atividade secretória de células glandulares e epiteliais.
Neurônios secretores estimulatórios liberam acetilcolina e VIP
para as células glandulares e epiteliais. Neurônios sensoriais
do gânglio submucoso respondem a estímulos químicos ou
deformação mecânica da mucosa. Eles fazem a mediação de
reflexos secretomotores. Outros são interneurônios para outros
neurônios do gânglio submucoso ou para neurônios mioentéricos.
O gânglio submucoso também contém neurônios vasodilatadores
que liberam acetilcolina e VIP para o músculo liso dos vasos
sangüíneos da mucosa.
Desta forma os únicos neurônios que fazem contato com a
musculatura lisa do trato digestivo são os do sistema nervoso
entérico. Para atuar, o sistema nervoso central, depende da
comunicação através dos ramos simpáticos e parassimpáticos,
que
se
interligam
ao
sistema
nervoso
entérico,
que
efetivamente atuará na musculatura lisa do tubo digestivo.
O sistema nervoso entérico é de tal importância que
permite ao trato digestivo atuar de forma ordenada mesmo na
ausência da inervação extrínseca. Porém deve ficar claro que a
inervação extrínseca exerce papel de grande importância no
funcionamento normal do aparelho digestivo, sendo que na sua
ausência,
várias
respostas
reflexas
diminuem
ou
até
desaparecem
completamente.
Estes
reflexos
são
melhor
percebidos nas porções mais proximais do tubo (esôfago e
estômago), contando com a mediação do nervo vago.
Podemos mesmo dizer que o plexo mioentérico do esôfago
contém relativamente poucos gânglios, o mesmo ocorrendo na
região proximal do estômago, enquanto no estômago distal há
uma grande quantidade de gânglios e uma rede neural bem
desenvolvida. É interessante que o intestino delgado também
apresenta os plexos mioentéricos bem desenvolvidos, como no
antro gástrico. No intestino grosso o sistema nervoso entérico
apresenta variações na densidade dos gânglios ao longo de seu
comprimento. No cólon ele é comparável ao do intestino
delgado, porém ao longo de todo o intestino grosso há uma
redução da densidade de gânglios, no sentido próximo-distal,
sendo que no reto poucos gânglios são encontrados. Como era de
se esperar, os ramos simpático e parassimpático do SNA são
mais
importantes
(ou
"atuantes")
nas
regiões
onde
a
manifestação intrínseca é menor.
Para uma grande parte das funções do estômago e
intestinos o sistema nervoso entérico (SNE) é mais importante
que a inervação parassimpática e simpática. O estômago isolado
e partes isoladas do intestino são capazes de realizar
movimentos peristálticos graças ao SNE. Em condições de jejum
o tubo digestivo segue um padrão complexo de movimentos e
secreções, o complexo motor migratório interdigestivo que é
governado principalmente pelo SNE. As contrações que regulam
em grande parte o esvaziamento gástrico são coordenadas pelo
SNE a partir de informações que recebem do vago. Deve ficar
claro que o esvaziamento gástrico pode contar com a
participação
do
sistema
nervoso
central,
gânglios
paravertebrais e dos plexos intrínsecos.
Em linhas gerais a motilidade tende a ser estimulada pelo
parassimpático e inibida pela estimulação do simpático. A
utilização do termo motilidade se refere aos movimentos do
aparelho digestivo, e não a contração muscular, uma vez que
tanto o simpático quanto o parassimpático podem causar
contração muscular de um segmento e relaxamento de outros.
Outra questão interessante é o modelo proposto para a
contração muscular no intestino delgado. A musculatura deste
segmento gera ondas lentas e apresenta contrações espontâneas.
Estas contrações são impedidas, quando o músculo está
relaxado, por neurônios inibitórios do plexo mioentérico. Os
neurônios inibitórios do plexo mioentérico recebem junções
sinápticas de outros neurônios cuja única função é produzir um
ritmo elétrico básico capaz de manter os neurônios inibitórios
atuando nas células lisas da musculatura circular a ponto de
manter um tônus basal. Os neurônios inibitórios também recebem
conexões sinápticas de outros neurônios que podem produzir uma
hiperpolarização
ou
despolarização.
No
caso
de
haver
hiperpolarização,
estes
neurônios
param
de
liberam
o
neurotransmissor inibitório e o músculo liso se contrai. Caso
o
neurônio
inibitório
se
despolarize
a
liberação
do
neurotransmissor inibitório aumenta e o músculo liso diminui
seu tônus (relaxa).
Desta
forma,
como
veremos
adiante,
o
movimento
peristáltico, por exemplo, seria resultado de uma onda de
hiperpolarização dos neurônios inibitórios, que cessariam de
atuar sobre as células do músculo liso, causando conseqüente
despolarização e contração dessas células por seus próprios
potenciais miogênicos. Isto explica porque destruição de
partes da inervação intrínseca leva a contração da camada
circulas do segmento onde houve a lesão.
Neurotransmissores
controle da motilidade
e
hormônios
que
participam
do
Podemos
dividir
os
hormônios
e
neurotransmissores
envolvidos nas respostas motoras gastrointestinais em dois
grupos: 1) Estimuladores e 2) Inibidores. Porém, cabe aqui uma
ressalva: a resposta de uma célula a um neurotransmissor
depende da própria célula, através de proteínas receptoras de
membrana plasmática (principalmente). Portanto nos referiremos
aqui a estimuladores como aqueles que estimulam fenômenos
motores como um todo, por exemplo: a colecistocinina estimula
a contração da vesícula biliar (excitatória) e relaxa o
esfíncter de Oddi (inibitória) para a liberação da bile;
também aumenta a motilidade intragástrica, embora diminua a
velocidade do esvaziamento do estômago, pelo aumento do tônus
no esfíncter pilórico. Por isso ao falarmos em excitação ou
inibição devemos ter cautela com aparentes paradoxos.
Para ilustrar vamos citar alguns neurotransmissores
encontrados no SNE.
Transmissores excitatórios:
- Acetilcolina (ACh)
- Serotonina (5-hidroxitriptamina, 5-HT)
- Histamina
- Colecistocinina (CCK)
- Motilina
- Gastrina
- Substância P
Transmissores inibitórios:
- Dopamina (DA)
- Noradrenalina (NA)
- Peptídeo Intestinal Vasoativo (VIP)
- Somatostatina
- Encefalinas (ENK)
- Óxido nítrico (NO)
Serotonina - largamente distribuída no trato gastrointestinal,
e parece estar bem estabelecido que atua sobre os neurônios do
plexo mioentérico, de forma estimulatória sobre a motilidade,
atuando em nível neuro-neural.
Dopamina - Catecolamina largamente distribuída
produzida e estocada em neurônios entéricos.
no
SNE,
é
Encefalinas e endorfinas - Estes neuropeptídeos são opiáceos
endógenos.
A
alteração
da
motilidade
digestiva
por
encefalinas, endorfinas e drogas do tipo morfina está bem
determinada.
Opiáceos
parecem
inibir
a
liberação
de
acetilcolina
do
plexo
mioentérico.
Isto
tem
levado
a
especulação que opiáceos endógenos poderiam estar atuando como
moduladores, e não como transmissores. Porém há evidências
claras que os opiáceos alteram a excitabilidade do corpo
celular
dos
neurônios
mioentéricos,
levando-os
a
hiperpolarização por aumento da condutância ao potássio. Isto
seria devido a um efeito direto na membrana da célula póssináptica, e não pela interferência com a transmissão
sináptica entre neurônios. Esta evidência coloca estes
neuropeptídeos como transmissores e não como moduladores
apenas.
Substância P - Peptídeo com 11 aminoácidos que como as
encefalinas apresenta-se amplamente distribuído. É encontrada
principalmente no sistema nervoso entérico, embora também
tenha sido encontrada em células endócrinas.
A substância P apresenta vários efeitos biológicos. Na
musculatura intestinal estimula a potência contrátil. A
substância P pode despolarizar certos neurônios mioentéricos.
Somatostatina - Tetradecapeptídeo que se encontra dentro de
neurônios mioentéricos (e de outras partes do sistema nervoso,
como gânglios prevertebrais, e parte do encéfalo, como
hipotálamo). Há sugestões de que os neurônios entéricos que
possuem somatostatina sejam interneurônios.
Peptídeo Intestinal Vasoativo (VIP) - É um peptídeo de 27
aminoácidos estruturalmente relacionado à secretina, glucagon
e peptídeo inibidor gástrico (que serão discutidos quando
tratarmos da motilidade em cada segmento do tubo digestivo). É
encontrado em neurônios ao longo do todo o trato digestivo,
bem como gânglio paravertebral e muitas partes do cérebro. É
forte estimulador das secreções, é dilatador de vasos
esplâncnicos e salivares. Sua ação na musculatura lisa
intestinal é inibitória, e seu papel em sinapses neuro-neurais
não está ainda claro.
Colecistocinica-pancreozimina (CCK-PZ) - Peptídeo secretado
pela mucosa duodenal que apresenta grande semelhança com a
gastrina. Devido a essa semelhança estrutural tendem a atuar
de forma parecida, com ação estimulatória, sobre a motilidade
gastroduodenal como veremos adiante.
A CCK-PZ é mais especificamente indutora da secreção
pancreática (protéica) e da contração da vesícula biliar,
enquanto a gastrina é especificamente indutora da secreção
gástrica.
A CCK-PZ também já foi encontrada em neurônios cerebrais,
concentrada em vesículas sinápticas.
Motilina - Secretada pelo duodeno, estimula movimentos
característicos da fase interdigestiva. Sua concentração
sangüínea parece acompanhar as variações de movimento típicas
dessa fase.
Secretina - Secretada pela mucosa duodenal, este hormônio
protéico inibe contrações antrais e estimula a contração do
esfíncter pilórico. Também estimula o pâncreas a secretar
solução rica em bicarbonato.
A secretina é liberada principalmente em resposta a
presença de ácido no duodeno.
Óxido nítrico - O óxido nítrico é um gás que tem papel
neurotransmissor no sistema nervoso e é conhecido relaxante
(inibidor) de músculo liso em geral. Seu papel é mais óbvio
como neurotransmissor inibitório no esôfago, onde juntamente
com o VIP desempenha importante papel no trânsito esofágico,
como veremos adiante.
A atuação das principais substâncias transmissoras bem
como outras não comentadas até aqui, serão discutidas ao longo
do estudo da atividade motora de cada órgão.