UMA RETROALIMENTAÇÃO POSITIVA ENTRE A
PRODUÇÃO DE GASES DO EFEITO ESTUFA E O
AUMENTO DE TEMPERATURA TAMBÉM NOS
TRÓPICOS?
Humberto Marotta
Universidade Federal Fluminense – Dep. Geografia/Pólo de Campos dos Goytacazes, Brasil
[email protected]
Luana Pinho
Universidade Federal do Rio de Janeiro – Programa de Pós-Graduação em Ecologia, Brasil
[email protected]
Cristian Gudasz
Universidade de Uppsala – Centro de Biologia e Evolução, Suécia
[email protected]
David Bastviken
Universidade de Linkoping – Centro de Estudos Ambientais e da Água, Suécia
[email protected]
Lars Tranvik
Universidade de Uppsala – Centro de Biologia e Evolução, Suécia
[email protected]
Alex Enrich-Prast
Universidade Federal do Rio de Janeiro – Programa de Pós-Graduação em Ecologia, Brasil
[email protected]
Resumo
Os sedimentos das águas interiores representam um importante sítio sem oxigênio
(anaeróbico) para o acúmulo e a preservação do carbono orgânico em escala global,
estocando muito mais carbono anualmente do que os sedimentos oceânicos. Estudos
recentes nos ecossistemas frios das altas latitudes têm demonstrado uma
retroalimentação positiva entre o aumento da temperatura e da mineralização
orgânica, a qual produz importantes gases causadores do próprio aquecimento global
por Efeito Estufa como o dióxido de carbono (CO2) e o metano (CH4). Por outro lado,
os estoques orgânicos das regiões tropicais naturalmente mais quentes têm sido
menos investigados. No presente estudo, as taxas de liberação de CO2 e CH4 pela
mineralização anaeróbica da matéria orgânica no sedimento lacustre foram
mensuradas em um gradiente experimental de temperatura de 4 a 40 °C. Foram
estudados 9 lagos tropicais (Amazônicos, Brasil) e 8 boreais (Floresta de Coníferas,
Suécia), incluindo representativos tipos de ecossistemas lacustres do planeta. A partir
dos nossos resultados experimentais e assumindo projeções conservativas do IPCC
para o aquecimento regional em 2100 (cenário B1) de 1.8-4.5 0C para a Amazônia e
de 1.6-4.2 0C para a Floresta de Coníferas da Suécia, foi possível estimar um futuro
aumento quase 5 vezes maior das taxas de mineralização da matéria orgânica
anaeróbica nos sedimentos lacustres na Amazônia do que na Floresta de Coníferas,
respectivamente 8.4-23.3 and 1.5-4.2 gC kg sed-1 a-1. Além disso, o aumento na
produção CH4 poderia ser em torno de 4 vezes mais alto nos sedimentos tropicais do
que nos boreais usando estas estimativas, um gás cujo potencial ao aquecimento
global é reconhecidamente mais elevado do que o CO2. Como conclusão, nossos
resultados sugerem que as repostas ao aquecimento nos Trópicos, resultando na
redução dos estoques de carbono orgânico e no aumento das emissões de gases do
Efeito Estufa, podem ser similares ou até mesmo mais elevadas do que nas altas
latitudes.
Agradecimentos: Estudo desenvolvido com recursos da Fundação Sueca para Cooperação
Internacional em Pesquisa e Educação Superior (bolsa de pós-doutorado e financiamento de
pesquisa). Trabalho apresentado no 4º Seminário de pesquisa do Instituto de Ciências da
Sociedade e Desenvolvimento Regional, da Universidade Federal Fluminense - UFF, realizado
em Campos dos Goytacazes, RJ, Brasil, em março de 2011".
Introdução
As florestas tropicais se caracterizam por temperaturas amenas ao longo
do ano inteiro, o que contribui não só para a elevada fixação de carbono (C) em
moléculas orgânicas pela produção primária, mas também para a intensa
mineralização dos substratos orgânicos (Ometto et al. 2005; Rice et al. 2004;
Telles et al. 2003). A menor limitação à decomposição biológica nos trópicos
(Davidson et al. 2000) contrasta com as temperaturas anuais mais frias nas
latitudes elevadas, que apesar de reduzirem a fixação biológica de C também
preservam da degradação importantes estoques globais de carbono orgânico.
Estes estoques de carbono nos ecossistemas boreais são reconhecidamente
sensíveis ao aquecimento (Gudasz et al. 2010; Hashimoto 2005; Zimov et al.
2006).
Dessa forma, grandes estoques orgânicos tanto nas baixas quanto nas
altas latitudes podem ser produzidos nos lagos ou ainda alcançá-los pelo
aporte de rios ou do escoamento e lixiviação terrestre na bacia de drenagem
(Cole et al. 2007), tornando os ecossistemas lacustres componentes
importantes no ciclo global de C (Cole et al. 1994; Duarte and Prairie 2005;
Marotta et al. 2009; Tranvik et al. 2009). Os biomas tropicais e boreais
possuem uma grande parte dos lagos do planeta (Downing et al. 2006), nos
quais o carbono orgânico é mineralizado pelo metabolismo biológico aos mais
importantes gases do efeito estufa (GEE), destacando o dióxido de carbono
(CO2) e o metano (CH4). Os lagos são, portanto, relevantes fontes de CO2
(Cole et al. 1994; Marotta et al. 2009; Tranvik et al. 2009) e CH4 (Bastviken et
al. 2004) à atmosfera, sustentada principalmente pela decomposição biológica
da matéria orgânica (Bastviken et al. 2003; Duarte and Agusti 1998; Duarte and
Prairie 2005), que pode ser especialmente intensa nos tropicais (Marotta et al.
2010). O carbono orgânico remanescente não degradado é, por sua vez,
estocado nos sedimentos lacustres, que constitui um relevante sumidouro
global que remove C da atmosfera por longos períodos de tempo (Downing et
al. 2008; Sobek et al. 2009; Tranvik et al. 2009). Dessa forma, a avaliação dos
efeitos das mudanças de temperatura sobre degradação orgânica é
fundamental para a compreensão dos mecanismos e das retroalimentações do
aquecimento global.
A forte correlação positiva entre o aquecimento global e a mineralização
de carbono orgânico nos sedimentos dos lagos boreais foi recentemente
descrita em condições aeróbias, elevada disponibildade de oxigênio (Gudasz et
al. 2010). Além disso, o rápido consumo do oxigênio nos sedimentos lacustres
ricos em matéria orgânica também poderia aumentar o papel da mineralização
anaeróbica do carbono orgânico (Schulz and Conrad 1996; Sobek et al. 2009).
Sendo assim, os estudos entre diferentes biomas podem revelar importantes
características da sensibilidade de estoques de C ao redor do mundo às
alterações climáticas. No entanto, os efeitos das mudanças de temperatura do
processo de degradação biológica em ambientes quentes tropicais ainda é
incerto (Hashimoto 2005; Holland et al. 2000). No presente estudo,
comparamos a sensibilidade ao aumento de temperatura da mineralização
anaeróbica do carbono orgânico a CO2 e CH4 nos sedimentos amazônicos e
boreais. Testamos a hipótese de que o efeito do aquecimento global sobre a
mineralização orgânica anaeróbica pode ser maior nos sedimentos dos lagos
boreais do que nos amazônicos, resultado da combinação das temperaturas
mais baixas anuais, preservando os substratos orgânicos nas altas latitudes, e
do maior aumento esperado de temperatura pelas previsões do IPCC (IPCC,
2007).
Área de Estudo
Foram amostrados 9 lagos na Floresta Tropical Amazônica (Brasil) e 8 na
Floresta Boreal de Coníferas (Suécia), abrangendo um amplo gradiente de
carbono orgânico dissolvido (COD) nas águas e de conteúdo orgânico no
sedimento lacustres em cada bioma (Tabela 1). As amostras de sedimento
foram coletadas simultaneamente, permitindo a incubação experimental
conjunta na Suécia. Os lagos amazônicos estudados são situados nas
margens do rio Amazonas a oeste da cidade de Santarém (estado do Pará,
Brasil), entre as latitudes 2º 25’ e 2º 31’ S e longitudes 54º 05’ e 54º 57’ W. Os
lagos suecos são situados ao Norte da cidade de Uppsala, entre as latitudes
63º 13’ e 59º 51’ S e longitudes 14º 17’ e 17º 38’ E.
Tabela 1: Carbono e Nitrogênio nas águas e na porção superficial (10 cm) do
sedimento de fundo dos lagos analisados no experimento.
Águas Lacustres
10 cm no Topo dos Sedimentos Lacustres
N-Total
(mgL-1)
P-Total
(µg L-1)
DOC
(mg L-1)
Densidade
Úmida do
Sedimento
(g/mL)
C
(g kg-1)
N
(g kg-1)
Razão
C:N
AM 1
AM 2
AM 3
AM 4
AM 5
AM 6
AM 7
AM 8
AM 9
0.76
0.98
1.66
0.47
0.53
1.49
0.97
0.25
0.46
106
363
505
57
40
132
137
15
17
1.3
13.1
6.6
1.4
2.1
1.8
2.9
0.6
1.0
1.30
1.43
1.44
1.15
1.05
1.11
1.61
1.02
1.53
10.7
5.0
4.5
13.5
19.0
13.5
4.0
12.6
2.8
0.8
0.5
0.5
1.0
1.2
1.1
0.4
0.9
0.3
12.9
10.0
9.0
13.7
16.2
12.8
10.8
13.9
10.7
BO 1
BO 2
BO 3
BO 4
BO 5
BO 6
BO 7
BO 8
1.16
1.66
0.81
0.95
0.62
0.78
0.20
0.84
24
54
44
42
11
12
13
20
30.4
26.3
10.8
16.4
14.2
17.0
6.0
19.7
0.99
1.05
1.01
1.02
1.00
1.04
1.00
0.95
16.4
11.4
9.3
10.2
14.7
16.0
13.9
13.4
1.2
1.2
1.1
1.2
0.9
0.8
0.9
0.9
13.2
9.9
8.8
8.8
16.6
19.7
15.4
15.5
Métodos
As amostras de sedimento de fundo e de águas superficiais foram
coletadas em cada lago para as incubações experimentais e as análises das
concentrações de nutrientes e de carbono orgânico (APHA 1992). No campo,
perfis verticais de temperatura e oxigênio dissolvido na coluna d’água foram
mensurados com eletrodos previamente calibrados. No laboratório logo após a
coleta, foram realizadas medidas de pH nas amostras de água, usando um
pHmetro com precisão de 0,01 unidades de pH. Amostras de água total e
filtrada (filtro Whatman GF/F, poro de 7 μm) foram congeladas para as análises
de
nitrogênio
e
fósforo
por
cromatografia
iônica
(APHA
1992).
As
concentrações de clorofila-a foram extraídas dos filtros de amostras de 500 mL
de água com etanol no escuro por 24 h e analisadas por espectrofotometria,
usando comprimentos de onda de 433 nm para medidas da excitação e 673 nm
da emissão (Nusch and Palme 1975). Além disso, amostras de água filtrada e
pré-acidificada a pH menor que 2,0 foram analisadas para as concentrações de
COD pelo método da oxidação catalítica em um analisador Shimadzu TOC5000. O conteúdo de água no sedimento foi determinado pela diferença em
uma balança de precisão entre os pesos úmido e seco (liofilizado). Já os
conteúdos de carbono orgânico e nitrogênio no sedimento foram determinados
com um analisador CHN e a composição da matéria orgânica foi avaliada pelo
método de identificação espectrofluorimétrica (Stedmon et al. 2003).
O experimento para testar o efeito da temperatura sobre os diferentes
sedimentos lacustres boreais e tropicais foi baseado no método destrutivo com
frascos de vidro de 25 mL. Estes frascos foram previamente lavados e
hermeticamente lacrados com rolhas massivas de borracha de butil
impermeável a gás (Apodan, Denmark) e um selo de alumínio após a
incubação das amostras. As rolhas usadas foram fervidas pelo menos 5 vezes
para extrair substâncias tóxicas. As amostras de água e sedimento foram
homogeneizadas e bombeadas com N2, isoladamente, no sentido de se
obterem
condições
anóxicas.
Posteriormente,
cada
frasco
por
lago,
temperatura e tempo experimental (4 réplicas) foi preenchido com 5 mL de
água total e 5 mL de sedimento, bombeados novamente com N2 antes do
fechamento para evitar qualquer oxigenação, de forma a restarem 15 mL de ar
dentro dos frascos. Os frascos fechados com amostras foram acondicionados
dentro de 6 caixas contendo água com diferentes temperaturas controladas (4,
10, 16, 22, 30, 40 0C). A atividade biológica em cada frasco por lago e
temperatura foi interrompida em cada tempo experimental (0, 3, 6, 10, 20, 30 e
44 dias), acidificando a pH < 1,5 pela adição de 2 mL de ácido sulfúrico 10 %
através das rolhas com uma seringa e agulha e mistura vigorosa. No total,
foram usados 2944 frascos de amostras para 16 lagos, 6 temperaturas e 8
tempos experimentais, incluindo o dia zero antes da manipulação de
temperatura. Após vigorosa homogeneização e coleta com seringa e agulha, as
concentrações de CO2 e CH4 do ar armazenado dentro dos frascos foram
quantificadas em um cromatógrafo gasoso Agilent 6890A acoplado a um
detector FID e metanizador catalítico de níquel.
Resultados e Discussão
Os sedimentos lacustres amazônicos apresentaram uma menor fração
de carbono orgânico por peso seco do que os boreais (média ± SE = 4,4 ± 1,6
e 18,2 ± 2,6 % respectivamente, teste t, p < 0,0005), provavelmente refletindo a
influência dos solos ricos em argila da Amazônia (Telles et al. 2003). No
entanto, no balanço por unidade de volume não houve diferença significativa
dos conteúdos de C e N ou da razão C:N comparando os sedimentos lacustres
dos dois biomas (t-teste, p > 0,05). Este resultado rejeitou a hipótese,
sugerindo que, mesmo influenciado por acentuadas diferenças nas condições
ambientais e nos aportes alóctones, importantes estoques de carbono
orgânico, pelo menos na camada superficial do sedimento, podem ser similares
comparando os lagos amazônicos e boreais.
A mineralização anaeróbica nos sedimentos da maioria dos lagos
estudados (cerca de 70%) foi positivamente relacionada à temperatura
(regressão linear, p <0,05), confirmando a potencial relação positiva também
entre a decomposição anaeróbica e a temperatura (Schulz e Conrad, 1996).
Diversos fatores relacionados à baixa quantidade ou qualidade dos substratos
orgânicos poderia explicar a ausência de respostas significativas com o
aumento de temperatura nos lagos restantes (Davidson e Janssens 2006). No
presente experimento, as taxas médias de aumento da produção anaeróbica
de CO2 e CH4 com o aquecimento experimental foram significativas e similares
comparando ambos os sedimentos tropicais e boreais (ANCOVA, p > 0,05;
Figura 1).
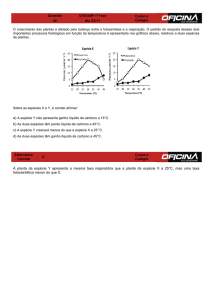
Figura 1: Sensibilidade da produção anaeróbica de CH4, CO2 e da
mineralização total do carbono orgânico ao aumento de temperatura nos
sedimentos lacustres amazônicos e boreais. Cada símbolo representa a média
e as barras o desvio padrão para os lagos amazônicos (quadrados
preenchidos, n = 9) e boreais (quadrados abertos, n = 8), os quais foram
incubados nas mesmas temperaturas. Linhas contínuas e as linhas tracejadas
representam a equação de regressão ajustada (R2> 0,90, p <0,005) para os
sedimentos amazônicos e boreais respectivamente. A inclinação e o intercepto
no eixo Y não foram significativamente diferentes entre os dois biomas para
todas as variáveis estudadas (ANCOVA , P> 0,05): (1A) produção anaeróbica
de CH4 = 0.10e 0.11 * temperatura, (1B) produção anaeróbica de CO2 = 3.71e
0.08Temperatura *
, e (1C), mineralização anaeróbica total do carbono orgânico =
2.35e 0.07 * Temperatura (unidades de cada variável descritas em cada gráfico).
Estudos prévios têm concluído que a mineralização de carbono orgânico
nos solos tropicais naturalmente quentes pode ser muito sensível a pequenas
variações de temperatura (Townsend et al. 1992). Esta sensibilidade à
temperatura pode ser ressaltada especialmente onde nem água ou substratos
orgânicos são limitantes (Hashimoto 2005; Holland et al. 2000), uma condição
que é tipicamente observada nos sedimentos lacustres (Downing et al. 2008;
Tranvik et al. 2009). Nos lagos tropicais, os decompositores anaeróbios
também podem ser estimulados pelo aumento de temperatura (Schulz and
Conrad 1996; Thebrath et al. 1993).
Nesse sentido, as relações significativas exponenciais da produção
anaeróbica de CO2 e CH4 com o aquecimento experimental revelou que,
apesar de não haver diferença significativa nas taxas de incremento médio
comparando os lagos das latitudes baixas e altas (ANCOVA, p> 0,05;. Figura
1), os valores máximos foram observados nos sedimentos tropicais. Estes
resultados sugerem que o efeito do aumento de temperatura sobre a
mineralização orgânica pode ser significativo não só nos sedimentos lacustres
das altas latitudes ou em condições aeróbias como previamente evidenciado
(Gudasz et al., 2010), mas também nas regiões tropicais e sob condições
anóxicas fortemente dominante nestes estoques de C ao redor do mundo.
Portanto, a nossa hipótese sobre a maior sensibilidade à temperatura nos
sedimentos lacustres boreais do que nos amazônicos também foi refutada.A
partir das relações entre a mineralização e a temperatura descritas na figura 1,
estimamos
a
mineralização
do
carbono
orgânico
anaeróbio
sob
as
temperaturas anuais médias atualmente observadas no campo por cada bioma.
Os sedimentos lacustres amazônicos apresentaram taxas de mineralização
anaeróbica do carbono orgânico (média ± SE de inclinação e intercepto-Y),
quase seis vezes maior do que as dos lagos boreais, respectivamente, 61,5
(43,2-87,3) e 11,1 (9,3-13,2) mg kg C sed-1 a-1 . Este resultado confirmou o
metabolismo biológico mais intenso em condições mais quentes (Brown et al.
2004), além da potencial natureza exponencial da sensibilidade das mudanças
bióticas ou da degradação orgânica ao aquecimento (Davidson and Janssens
2006; Dillon et al. 2010).
Nesse contexto, foram também estimados os possíveis aumentos das
emissões anaeróbicas de CO2 e CH4 pelos sedimentos lacustres (Tabela 2),
assumindo os cenários regionais do IPCC mais conservadores para o
aquecimento global ao longo deste século (cenário B1: 1,6-4,2 0C para a
Amazônia e 1,8-4,5 0C para o Norte da Europa; IPCC, 2007). A estimativa de
incremento médio das taxas de mineralização biológica anaeróbica com o
aumento de temperatura foi substancialmente mais elevada nos sedimentos
amazônicos do que nos suecos. Além disso, a proporção da taxa de aumento
da produção de CH4 em relação àquela de CO2 foi quase quatro vezes maior
nos sedimentos lacustres tropicais que nos boreais (Tabela 2). Este resultado
provavelmente refletiu a alta contribuição da metanogênese ou da redução de
CO2 a CH4 nestes ecossistemas tropicais quentes (Conrad et al. 2010), pois a
última etapa da degradação da matéria orgânica produzindo CH 4 é fortemente
limitada em baixas temperaturas (Nguyen et al. 2010; Schulz and Conrad
1996), enquanto a oxidação do CH4 não é dependente da temperatura (Nguyen
et al., 2010). A molécula de CH4 apresenta uma propriedade de efeito estufa
cerca de 21 vezes mais intensa do que a de CO2, tornando esta mais alta
produção de CH4 observada nos sedimentos amazônicos potencialmente
relevante ao aquecimento global.
Tabela 2: Estimativas de aumento da mineralização anaeróbica total do
carbono orgânico e da produção anaeróbica de CO2 e CH4 pelos sedimentos
lacustres (valores em g C kg sed -1 a-1).
Sedimentos
Lacustres
Mineralização
orgânica
anaeróbica total
Produção
anaeróbica de
CO2
Prdoução
anaeróbica de
CH4
Média* Limítrofe* Média* Limítrofe* Média* Limítrofe*
Amazônico
15.9
8.4-23.3
13.6
7.3-19.8
3.2
1.6-4.7
Boreal
2.9
1.5-4.2
3.5
1.9-5.0
0.2
0.1-0.3
Amazônico/
Boreal
5.6
-
3.9
-
14.3
-
* - O aumento dessas taxas foi calculado utilizando a previsão mais conservadora do
IPCC para o aquecimento regional projetada para 2100 (cenário B1). Esta previsão foi
simulada por um conjuntos de dados multi-modelos que incorpora causas conhecidas
na Amazônia (1,6-4,2 0C) e na Suécia (1,8-4,5 0C; IPCC , 2007). Foram calculadas as
médias entre as duas estimativas dentro do intervalo de probabilidade de cada
intervalo de aquecimento projetado.
Extrapolando para a área ocupada pelos lagos nas zonas tropicais e
boreais e assumindo as previsões de aquecimento do cenário B1 do IPCC
(IPCC 2007), o aumento relacionado apenas à camada superficial de 10 cm do
sedimento de fundo já poderia representar um significativo efluxo de C destes
estoques orgânicos nos lagos (0,60-1,67 e 0,19-0,53 Tg C a-1 para os
sedimentos tropicais e boreais, respectivamente). Estas estimativas poderiam
ser ainda substancialmente maiores se incluissem as camadas mais profundas
do sedimento e não apenas os 10 centímetros iniciais ou ainda de 2 a 5 vezes
maiores assumindo outros cenários menos conservadores do IPCC para o
aquecimento global (como A1B e B2).
Mesmo com uma área estimada quase 2,5 maior (Molot e Dillon 1996)
do que a dos lagos tropicais (Marotta et al. 2009), os sedimentos de lagos
boreais apresentaram um aumento na mineralização anaeróbica cerca de dois
terços menor. Estes resultados confirmam que, apesar do menor aquecimento
esperado que os ecossistemas do norte (IPCC, 2007), as águas tropicais
também poderiam apresentar importantes mudanças bióticas na decomposição
aquática e na produção de GEE pelo aquecimento, como previamente
evidenciado para as taxas metabólicas terrestres nos trópicos em função de
efeitos não lineares da temperatura sobre o metabolismo (Dillon et al. 2010) .
Portanto, nossos resultados sugerem consistentemente uma potencial
contribuição significativa tanto de baixas quanto de altas latitudes para uma
retroalimentação positiva entre o aquecimento global e a mineralização
orgânica nos sedimentos lacustres. Grandes estoques orgânicos tropicais
submersos por águas aquecidas nos ecossistemas aquáticos e nas áreas de
inundação poderiam constituir sítios ativos potenciais para mineralização
orgânica a gases do efeito estufa por aumentos de temperatura, uma
retroalimentação positiva a ser considerada em qualquer cenário de
aquecimento global.
Bibliographic References
APHA. 1992. Standard methods for the examination of water and wastewater. ,
18 ed. American Public Health Association.
Bastviken, D., J. Cole, M. Pace, and L. Tranvik. 2004. Methane emissions from
lakes: Dependence of lake characteristics, two regional assessments, and a
global estimate. Global Biogeochemical Cycles 18.
Bastviken, D., J. Ejlertsson, I. Sundh, and L. Tranvik. 2003. Methane as a
source of carbon and energy for lake pelagic food webs. Ecology 84: 969981.
Brown, J. H., J. F. Gillooly, A. P. Allen, V. M. Savage, and G. B. West. 2004.
Toward a metabolic theory of ecology. Ecology 85: 1771-1789.
Cole, J. J., N. F. Caraco, G. W. Kling, and T. K. Kratz. 1994. Carbon-Dioxide
Supersaturation in the Surface Waters of Lakes. Science 265: 1568-1570.
Cole, J. J. and others 2007. Plumbing the global carbon cycle: Integrating inland
waters into the terrestrial carbon budget. Ecosystems 10: 171-184.
Conrad, R., M. Klose, P. Claus, and A. Enrich-Prast. 2010. Methanogenic
pathway, C-13 isotope fractionation, and archaeal community composition in
the sediment of two clear-water lakes of Amazonia. Limnology and
Oceanography 55: 689-702.
Davidson, E. A., and I. A. Janssens. 2006. Temperature sensitivity of soil
carbon decomposition and feedbacks to climate change. Nature 440: 165173.
Davidson, E. A., L. V. Verchot, J. H. Cattanio, I. L. Ackerman, and J. E. M.
Carvalho. 2000. Effects of soil water content on soil respiration in forests and
cattle pastures of eastern Amazonia. Biogeochemistry 48: 53-69.
Dillon, M. E., G. Wang, and R. B. Huey. 2010. Global metabolic impacts of
recent climate warming. Nature 467: 704-U788.
Downing, J. A. and others 2008. Sediment organic carbon burial in agriculturally
eutrophic impoundments over the last century. Global Biogeochemical Cycles
22.
Downing, J. A. and others 2006. The global abundance and size distribution of
lakes, ponds, and impoundments. Limnology and Oceanography 51: 23882397.
Duarte, C. M., and S. Agusti. 1998. The CO2 balance of unproductive aquatic
ecosystems. Science 281: 234-236.
Duarte, C. M., and Y. T. Prairie. 2005. Prevalence of heterotrophy and
atmospheric CO2 emissions from aquatic ecosystems. Ecosystems 8: 862870.
Gudasz, C., D. Bastviken, K. Steger, K. Premke, S. Sobek, and L. J. Tranvik.
2010. Temperature-controlled organic carbon mineralization in lake
sediments. Nature 466: 478-U473.
Hashimoto, S. 2005. Temperature sensitivity of soil CO2 production in a tropical
hill evergreen forest in northern Thailand. Journal of Forest Research 10:
497-503.
Holland, E. A., J. C. Neff, A. R. Townsend, and B. Mckeown. 2000.
Uncertainties in the temperature sensitivity of decomposition in tropical and
subtropical ecosystems: Implications for models. Global Biogeochemical
Cycles 14: 1137-1151.
IPCC. 2007. Climate change 2007: The physical science basis, p. 18.
Marotta, H., C. M. Duarte, F. Meirelles-Pereira, L. Bento, F. A. Esteves, and A.
Enrich-Prast. 2010. Long-term variability of CO2 in two shallow tropical lakes
experiencing episodic eutrophication and acidification events. Ecosystems
13: 382–392.
Marotta, H., C. M. Duarte, S. Sobek, and A. Enrich-Prast. 2009. Large CO2
disequilibria in tropical lakes. Global Biogeochemical Cycles 23.
Nguyen, T. D., P. Crill, and D. Bastviken. 2010. Implications of temperature and
sediment characteristics on methane formation and oxidation in lake
sediments. Biogeochemistry 100: 185-196.
Nusch, E. A., and G. Palme. 1975. Biologische Methoden für die Praxis der
Gewässeruntersuchung.
Bestimmung
des
Chlorophyll-a
und
Phaeopigmentgehaltes in Oberflächenwasser. GWF-Wasser/Abwasser 116:
562-565.
Ometto, J., A. D. Nobre, H. R. Rocha, P. Artaxo, and L. A. Martinelli. 2005.
Amazonia and the modern carbon cycle: lessons learned. Oecologia 143:
483-500.
Rice, A. H. and others 2004. Carbon balance and vegetation dynamics in an
old-growth Amazonian forest. Ecological Applications 14: S55-S71.
Schulz, S., and R. Conrad. 1996. Influence of temperature on pathways to
methane production in the permanently cold profundal sediment of Lake
Constance. Fems Microbiology Ecology 20: 1-14.
Sobek, S. and others 2009. Organic carbon burial efficiency in lake sediments
controlled by oxygen exposure time and sediment source. Limnology and
Oceanography 54: 2243-2254.
Stedmon, C. A., S. Markager, and R. Bro. 2003. Tracing dissolved organic
matter in aquatic environments using a new approach to fluorescence
spectroscopy. Marine Chemistry 82: 239-254.
Telles, E. D. C. and others 2003. Influence of soil texture on carbon dynamics
and storage potential in tropical forest soils of Amazonia. Global
Biogeochemical Cycles 17.
Thebrath, B., F. Rothfuss, M. J. Whiticar, and R. Conrad. 1993. Methane
production in littoral sediment of Lake Constance. Fems Microbiology
Ecology 102: 279-289.
Townsend, A. R., P. M. Vitousek, and E. A. Holland. 1992. Tropical soils could
dominate the short-term carbon-cycle feedbacks to increased global
temperatures. Climatic Change 22: 293-303.
Tranvik, L. J. and others 2009. Lakes and reservoirs as regulators of carbon
cycling and climate. Limnology and Oceanography 54: 2298-2314.
Zimov, S. A., E. a. G. Schuur, and F. S. Chapin. 2006. Permafrost and the
global carbon budget. Science 312: 1612-1613.