1
SISTEMA EXCRETOR
Como foi visto no capítulo sobre Sistema Digestivo, as substâncias que não são absorvidas "para
dentro" de nosso organismo são eliminadas pelas fezes, ou seja, são excretadas, isto é, jogadas para fora
do organismo.
Já estudamos que o metabolismo (degradação) dos carboidratos e lipídios geram CO2 e H2O. O CO2 é
eliminado pelos pulmões enquanto que a água é em grande parte conservada dentro de nosso organismo,
sendo eliminada uma pequena parte dela através da urina, suor, expiração, etc. Como o processo
metabólico ocorre com todos os compostos moleculares existentes nos seres vivos, devemos lembrar que
as proteínas geram catabólitos (resíduos oriundos do metabolismo) que precisam ser eliminados. Esses
catabólitos são eliminados através dos sistemas excretores cujo representante principal é o sistema
urinário, porém a pele, através das glândulas sudoríparas e os pulmões também participam ativamente
desta função.
Assim a eliminação dos catabólitos nitrogenados, ou seja, os produtos gerados a partir do metabolismo
das proteínas (os aminoácidos, que constituem as proteínas possuem um grupo amina: NH2) como a
amônia, uréia, ácido úrico, uratos, serão excretados através do suor (glândulas sudoríparas) e da urina
(pelos rins).
Neste capítulo será estudado a anatomo-fisiologia renal que constitui o principal sistema excretor dos
vertebrados.
HOMEOSTASE: o famoso filósofo francês Claude Bernard disse: "Todos os mecanismos vitais, apesar
de sua diversidade, têm apenas uma finalidade, a de manter constantes as condições de vida no ambiente
interno". Todos os organismos vivos, mantêm, em grau maior ou menor, o estado interno constante, o
que se compreende por homeostase, independente dos extremos do ambiente externo. As atividades
reflexas do sistema nervoso e os hormônios do sistema endócrino constituem as bases do controle da
homeostase. Todas as partes do corpo dos animais, em todas as fases do crescimento e da reprodução,
encontram-se sob sua influência. A situação, mesmo no animal mais simples, é tão complexa e pouco
compreendida, que os processos de regulação são, geralmente, tratados de modo fragmentário, em termos
de alguns aspectos facilmente mensuráveis e não como um todo integrado. A pressão osmótica, a
concentração de íons de hidrogênio e a temperatura são três desses aspectos e cada um deles relaciona-se
intimamente com a água.
OSMORREGULAÇÃO: a água é ingerida juntamente com o alimento e, em formas aquáticas, entra até
certo ponto por absorção. O protoplasma é o solvente e o transportador universal e nenhum organismo
pode prescindir desse líquido essencial. Por sua propriedade de difundir-se através de membranas, a água
é o veículo para a manutenção do estado constante. Os líquidos do corpo de todos os animais são
semelhantes entre si. A regulação da pressão osmótica é simples para a maioria dos invertebrados
marinhos de corpo mole (cartilaginosos), porque seus líquidos do corpo encontram-se em equilíbrio (são
isotônicos) com a água que os circunda; as pressões são iguais interna e externamente e este equilíbrio é
mantido graças a sistemas especiais de controle osmótico existente no organismo desses animais. A água
doce contém apenas cerca de um centésimo da concentração salina da água do mar. Os líquidos do corpo
dos animais de água doce têm conteúdo salino superior ao do meio circundante (hipertônicos) e a água
tende a difundir-se para dentro; o excesso é eliminado de diversos modos. Os protozoários o fazem por
meio dos vacúolos pulsáteis, geralmente ausentes nas formas marinhas. Outros animais de água doce têm
nefrídios, tubos de Malpighi ou rins para excretar o excesso. Esse trabalho de excreção necessita de
energia, o que se comprova pela taxa respiratória mais alta dos animais de água doce em comparação com
espécies marinhas afins.
2
Osmorregulação em animais aquáticos: os vertebrados aquáticos mantêm o equilíbrio osmótico de
diferentes maneiras, de acordo com sua história evolutiva. Nos peixes ósseos, as formas de água doce têm
conteúdo salino maior que o do ambiente (hipertônico), o inverso ocorre, porém, com as formas marinhas
(hipotônico). A água entra, nos peixes de água doce, pelas membranas semipermeáveis das brânquias e da
boca, ao passo que tende a deixar o corpo nas formas marinhas. Nas formas de água doce, o excesso de
água é excretado pelos rins e os sais são mantidos nas concentrações apropriadas pela absorção por
células especiais das brânquias. Nas espécies marinhas, a deficiência é evitada pela deglutição e absorção
de água do mar, juntamente com os sais, pelo intestino. O excesso de sais é eliminado por células
secretoras das brânquias. Os rins dos peixes de água doce têm glomérulos bem desenvolvidos, para filtrar
grandes quantidades de água, mas nas formas marinhas os glomérulos são reduzidos e pouca água é
eliminada sob forma de urina. A enguia, o salmão e outros peixes migradores vivem, de modo alternado,
na água doce e na salgada; possivelmente seus glomérulos não sejam funcionais no mar. O sangue dos
peixes elasmobrânquios (tubarões e raias) têm, aproximadamente, o mesmo conteúdo salino que o dos
ósseos, mas contém, também, até 2% de uréia. Esta aumenta a pressão osmótica dos elasmobrânquios
marinhos até pouco acima da do ambiente, de modo que a água tende a entrar e não a sair, como nos
peixes ósseos marinhos. A uréia, nos elasmobrânquios, é retida por um seguimento absorvente especial
do túbulo renal e as brânquias são impermeáveis a ela.
Osmorregulação em animais aéreos: biguás, gaivotas e outras aves marinhas bebem água do mar para
atender às suas necessidades. A água é absorvida pela parede intestinal e o excesso de sais passa à
corrente sangüínea, com o qual é levado a um par de glândulas secretoras de sal situadas perto dos olhos e
com ductos para as narinas. A secreção produzida tem conteúdo salino maior que o da água do mar. Os
répteis marinhos também excretam o sal por glândulas nasais.
Osmorregulação em animais terrestres: os animais terrestres estão sujeitos ao risco da dessecação e
muitos deles têm cutícula impermeável que dificulta a perda de água pela superfície do corpo. Além
disso, desenvolveram-se meios para a reabsorção da água pelos rins e, em alguns casos, por células do
reto. No homem, a excreção ou retenção da água depende do estado de hidratação do corpo como um
todo. A transpiração excessiva diminui o volume de líquido eliminado com a urina, enquanto a ingestão
de grande quantidade de líquido ocasiona um aumento da produção de urina. O equilíbrio de água é
controlado, até certo ponto, pela sede, que varia de modo notável com o estado de hidratação, e pela ação
dos rins, influenciada por sua vez pelo hormônio antidiurético (ADH) secretado pelo lobo posterior da
hipófise. Na ausência desse hormônio, a reabsorção pelos túbulos renais diminui. O mecanismo de
controle é automático, pois um aumento da pressão osmótica do sangue causa aumento da secreção do
hormônio. Este, por sua vez, estimula a reabsorção e, assim, conserva-se a água. O álcool inibe a secreção
do hormônio antidiurético e tem, portanto, ação desidratante. A cafeína é diurética, aumentando a taxa de
filtração glomerular e diminuindo a reabsorção de água pelas células dos túbulos.
EXCREÇÃO DOS COMPOSTOS NITROGENADOS:
A excreção é o principal mecanismo homeostático através da osmorregulação feita por estruturas e
órgãos especializados nos diversos grupos animais.
A excreção é o processo de eliminar do corpo os resíduos do metabolismo. O protoplasma e os líquidos
de um animal, seja ele protozoários ou homem, formam um sistema físico-químico em delicado equilíbrio
e a função do aparelho excretor é manter esse ambiente interno constante. O excesso de água, sais
3
minerais orgânicos, inclusive dos resíduos metabólicos, é excretado, ao passo que as substâncias
essenciais às funções normais são conservadas.
A excreção relaciona-se principalmente com a eliminação de substâncias nitrogenadas. Na digestão, as
proteínas, contendo nitrogênio, são reduzidas a aminoácidos e estes absorvidos pelo organismo sendo
parte deles aproveitados pelas células para a síntese de novas proteínas. Nos vertebrados, outros
aminoácidos vão ao fígado, onde são decompostos para serem eliminados pelos rins.
A eliminação dos resíduos nitrogenados está também diretamente relacionada com as quantidades de
água de que dispõe o ser vivo. Os organismos que a possuem em grande quantidade eliminam amônia; os
organismos que precisam exercer um relativo controle sobre a quantidade de água eliminam uréia; e os
organismos que vivem em regime de grande economia de água eliminam ácido úrico.
Nos mamíferos, quelônios, anfíbios e anelídeos (minhoca) o produto final é principalmente a uréia que é
solúvel em água e bem menos tóxica que a amônia.
Nas aves e répteis terrestres e também em alguns caracóis e muitos insetos, o resíduo nitrogenado
compõe-se principalmente do ácido úrico insolúvel já que esses organismos dispõem de muito pouca
água, sendo esse excreta insolúvel em água e muito pouco tóxico. A formação de ácido úrico é importante
no desenvolvimento de embriões de aves e répteis para os quais, nos ovos, o suprimento de água e o
espaço interno são limitados. Assim, o embrião começa excretando amônia; depois a transforma em uréia
que, por sua vez, é transformada em ácido úrico. Os embriões de mamíferos que expelem para o sangue
materno os produtos de sua excreção, eliminam a uréia.
Os peixes teleósteos marinhos excretam até um terço do nitrogênio sob a forma de óxido de
trimetilamina. Muitos invertebrados e vertebrados aquáticos eliminam o nitrogênio como amônia,
composto este altamente tóxico, porém rapidamente eliminado porque há sempre excesso de água.
Alguns animais excretam aminoácidos diretamente e as aranhas excretam nitrogênio como guanina.
EXCREÇÃO NOS INVERTEBRADOS
O método mais simples de excreção é a passagem dos resíduos através da membrana celular diretamente
para a água ambiente, como ocorre em muitos protozoários. Ameba, Paramecium e muitos outros
protozoários da água doce têm um ou mais vacúolos pulsáteis (ou contráteis) que acumulam o excesso de
água do citoplasma e o descarregam periodicamente para o exterior, mantendo o equilíbrio osmótico do
corpo. O principal produto de excreção é a amônia. Os excretos das esponjas e celenterados difundem-se
das células do corpo para a epiderme e, daí, para a água.
Nos insetos e em alguns artrópodes, os principais órgãos excretores são os delgados tubos de malpighi,
ligados à parte anterior do intestino posterior, terminando em fundo cego; esses tubos recolhem as
excretas dos líquidos do corpo e os descarregam no intestino posterior. Tanto os uratos como o dióxido de
carbono são recebidos do sangue em solução; a água e outras substâncias são reabsorvidas na porção
superior dos tubos. Os produtos finais de excreção, incluindo os cristais de ácido úrico, carbonatos,
oxalatos e, às vezes, uréia ou amônia, são eliminados juntamente com as fezes. O exoesqueleto também
tem papel excretor em alguns invertebrados, inclusive nos insetos, pois substâncias nitrogenadas nele
depositadas são eliminadas por ocasião da muda.
Em muitos animais, os órgãos excretores mais comuns são estruturas tubulares, os nefrídios e os
celomodutos, dos quais havia, primitivamente, um par por segmento do corpo. No decurso da evolução,
modificaram-se diversamente. Os nefrídios têm origem ectodérmica. Os platelmintos possuem numerosas
4
células-flama (protonefrídios que terminam internamente em fundo cego) dispersas entre as células do
corpo, das quais os resíduos são recolhidos e lançados num sistema ramificado de ductos. Na minhoca,
cada segmento contém um par de nefrídios (metanefrídios, abertos internamente). A extremidade interna
de cada um deles termina num funil ciliado, o nefróstoma, que drena o celoma. Circundando o longo
túbulo, há vasos sangüíneos, dos quais também são retiradas excretas; o túbulo abre-se externamente por
um pequeno nefridióporo ventral. Os moluscos e os rotíferos têm um ou dois pares de órgãos
semelhantes a nefrídios que drenam o corpo ou o sangue.
Em alguns anelídeos, moluscos, artrópodes e nos cordados, os principais órgãos excretores são
celomodutos, de origem mesodérmica, derivados possivelmente de ductos genitais, mas diversamente
modificados para remover resíduos da cavidade do corpo. Os crustáceos têm dois pares, as glândulas
"antenais" (verdes) e "maxilares"; ambas têm um sáculo terminal com um ducto que se abre na base de
uma extremidade par. Apenas em raros casos ocorrem ambas bem desenvolvidas no mesmo estágio de
uma mesma espécie. As aranhas têm glândulas coxais derivadas de celomodutos.
EXCREÇÃO NOS VERTEBRADOS
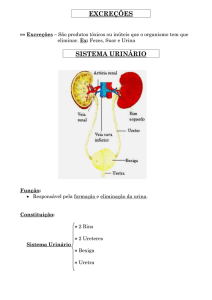
Os principais órgãos excretores de um vertebrado são os dois rins, onde ocorrerá a filtração do sangue,
num processo que resulta na eliminação de cerca de 2 litros de líquido, conhecido como urina. Na figura
1, pode ser visualizado as partes que compõe o sistema excretor urinário no homem, composto pelos rins,
ureteres, bexiga urinária e uretra.
5
Fig. 1 - Esquema geral do aparelho excretor no homem
Os rins são compostos, em geral, por uma massa de celomodutos que se abrem num ducto coletor. Os rins
nos peixes e salamandras estendem-se ao longo de quase toda a cavidade do corpo. Os rins dos
vertebrados inferiores, de ciclóstomos a anfíbios, e os embrionários dos grupos superiores desenvolvemse segmentarmente, um par por segmento do corpo (pronefro, mesonefro); alguns dos túbulos têm
nefróstomas abertos para o celoma, assemelhando-se, assim, aos nefrídios das minhocas. Os rins adultos
(metanefros) dos répteis, aves e mamíferos não são segmentares e drenam excretas unicamente do sangue.
Ver fig.2 que ilustra os principais tipos de rins dos vertebrados e o quadro 1 que mostra a seqüência de
ocorrência desses rins nos animais.
6
Fig.2-Esquema dos padrões básicos do aparelho excretor dos vertebrados.
O pronefro é o mais primitivo e, embora presente no desenvolvimento embrionário de todos os
vertebrados, não é funcional no adulto de nenhum deles. O rim funcional dos peixes é do tipo
mesonéfrico. Ele consiste de uma série de túbulos renais, que apresentam, no desenvolvimento inicial,
uma disposição segmentar, que desaparece posteriormente. Cada túbulo é enrolado tanto na porção
proximal como na distal, e dirige-se para um ducto coletor longitudinal comum chamado de ducto
arquinéfrico. Este, por sua vez, comunica-se com o meio exterior, em geral, pela cloaca recebendo
produtos dos sistemas digestivos e urogenital. A porção proximal de cada túbulo termina numa cápsula
hemisférica, conhecida como cápsula de Bowman, na qual existe um novelo vascular ou glomérulo do
sistema circulatório. A cápsula e o glomérulo formam juntos o corpúsculo renal. A figura 3 ilustra com
detalhes um rim humano e, ampliado, esta esquematizado os néfrons e os túbulos coletores.
7
Fig. 3 - Estrutura do Rim
De cada rim, qualquer que seja seu tipo, parte um ducto coletor comum, o ureter, que conduz as excretas
para a bexiga. Nos anfíbios, répteis e aves, os dois ureteres desembocam na cloaca, já nos anfíbios e
8
alguns répteis o ureter se liga a uma bexiga urinária. A excreta, ou urina, é líquida, exceto nos répteis e
aves, onde as excretas semi-sólidos (ácido úrico) são eliminados como uma pasta branca (guano)
juntamente com as fezes. Na maioria dos mamíferos, os ureteres comunicam-se diretamente com a
bexiga, da qual sai um ducto mediano, a uretra, que conduz o excreta ao exterior. Os aparelhos excretor e
reprodutor dos vertebrados, relacionados entre si, são freqüentemente reunidos sob o nome de aparelho
urogenital.
Quadro 1 - Tipos de rins nos animais
Concentração
sanguínea em
relação ao meio
Concentração
das excreções
em relação ao
sangue
Órgãos
excretores
Osmorregulação
Animal
Habitat
Produto de
excreção
Platelmintes
Água doce
Amoníaco
-
Hipotônica
Protonefrídios
com células
flama
Não bebem
água
Anelídeos
Água doce ou
terrestre
Amoníaco
Hipertônica
Hipotônica
Metanefrídeos
Não bebem
água
Insectos
Terrestre
Ácido úrico
-
Hipertônica
Túbulos de
Malpighi
Bebem água
Peixes
cartilagíneos
Água salgada
Uréia
Isotônica
Isotônica
Rins mesonefros
Não bebem
água, uréia
ajuda a reter
água.
Água salgada
Amoníaco
Hipotônica
Isotônica
Rins mesonefros
e brânquias
Bebem água e
excretam sal
Água doce
Amoníaco
Hipertônica
Hipotônica
Rins mesonefros
Não bebem
água e
absorvem sal
Água doce
terrestre
Amoníaco ou
uréia
Hipertônica
Hipotônica
Rins mesonefros
e pele
Não bebem
água e
absorvem sal
Água salgada
Uréia e ácido
úrico
Hipotônica
Hipertônica
Rins metanefros
Bebem água e
excretam sal
Terrestre
Ácido úrico
-
Hipertônica
Rins metanefros
Bebem água
Água salgada
Uréia
Hipotônica
Hipertônica
Rins metanefros
Não bebem
água
Terrestre
Uréia
-
Hipertônica
Rins metanefros
Bebem água
Peixes ósseos
Anfíbios
Répteis e Aves
Mamíferos
9
EXCREÇÃO NO HOMEM
No homem, cada rim contém cerca de um milhão de unidades filtradoras, chamadas néfrons. Os néfrons
consistem de um fino túbulo excretor que se origina numa estrutura chamada cápsula de Bowman. A
cápsula de Bowman envolve um novelo capilar chamado glomérulo. Cada túbulo termina em ductos
coletores que desembocam numa cavidade do rim partindo daí para um ureter. Uma grande quantidade de
sangue flui continuamente pelos rins. Cada rim recebe sangue da aorta pela artéria renal. Esta artéria se
ramifica em milhares de arteríolas, que vão formar os glomérulos e os capilares, que envolvem os túbulos
dos néfrons. Os capilares se reúnem em vênulas, que se juntam na veia renal, a qual sai do rim. O sangue
que entra no glomérulo está sob alta pressão. Essa pressão força a água e pequenas moléculas do plasma
sangüíneo para dentro da cápsula de Bowman. Este filtrado compõe-se de sais, glicose, aminoácidos,
uréia e outras substâncias de baixo peso molecular. Essa composição é igual à do plasma sangüíneo,
exceto pela falta de proteínas. Forma-se cerca de meio copo de filtrado por minuto nos dois rins.
Entretanto, a composição desse fluido se modifica bastante, e o seu volume é muito reduzido antes de
chegar ao ureter, pois à medida que o fluido percorre os túbulos dos néfrons, sofre um processo de
reabsorção (fig. 4). Ocorre uma reabsorção ativa de sais, glicose e aminoácidos e uma reabsorção passiva
de água. A volta de água para a corrente sangüínea se deve à diferença de pressão osmótica entre o túbulo
e os capilares sangüíneos que o envolvem. Cerca de 99% da água filtrada pelo glomérulo é reabsorvida
para o sangue nos túbulos. Só depois da reabsorção é que podemos falar em urina. A diferença entre o
filtrado e a urina é, então, praticamente quantitativa. Cada litro de urina formado, resulta de 180 litros de
plasma reciclado, filtrado pelos néfrons. A urina dos mamíferos contêm um resíduo nitrogenado menos
tóxico que a amônia: a uréia. Cada molécula de uréia formada no fígado, a partir da amônia, proveniente
da degradação de aminoácidos, é lançada no sangue e eliminada através dos rins.
Fig.4 - Filtração Renal
(http://curlygirl.no.sapo.pt/excrecao.htm)
10
A produção de urina é controlada de diversas maneiras. A filtração é diretamente influenciada pela
pressão sangüínea, que por sua vez, é influenciada pela epinefrina da medula adrenal que constringe os
vasos sangüíneos dos glomérulos.
Os hormônios produzidos pelo córtex da adrenal (aldosterona e cortisol), influenciam a reabsorção de
sódio, cloreto e glicose pelos túbulos renais e a eliminação de potássio. A reabsorção da água é o processo
mais importante da função renal. Cerca de 80% da água do líquido tubular é, provavelmente, recapturada
por difusão direta para os capilares e, destes, para o sistema venoso, pela atração dos colóides do sangue.
Outra porção da água é reabsorvida na alça de Henle e na parte distal do túbulo, por um mecanismo
controlado pelo hormônio antidiurético (ADH) secretado pela hipófise.
No fluxograma 2 está esquematizado o mecanismo de regulação do volume urinário. O volume urinário
é o reflexo do volume sangüíneo e pressão osmótica.
POR QUÊ O PH DA URINA É ÁCIDO?
Secreção de H+: as células dos túbulos proximais e distais, como as células das glândulas gástricas,
secretam íons hidrogênio. A acidificação também ocorre nos ductos coletores. A reação que é
primariamente responsável pela secreção de H+ no túbulo proximal é a troca Na+ por H+. Isto é um
exemplo de transporte ativo secundário. A saída de Na+ das células para espaços intercelulares é mediada
por uma enzima chamada Na+-K+ ATPase, que diminui o Na+ intracelular e isto causa entrada de Na+ nas
células do lúmem tubular com saída de H+. O H+ vem da dissociação intracelular do H2CO3, em HCO3- e
H+, que se difunde para o líquido intersticial.
Anidrase Carbônica: catalisa a formação de H2CO3 e drogas que inibem a anidrase carbônica deprimem
ambas, a secreção do ácido pelo túbulo proximal e reações que dependem dela.
Destino do H+ na Urina: a quantidade de ácido secretada depende de fatos subseqüentes que ocorrem na
urina tubular. O gradiente máximo de H+ contra o qual o mecanismo de transporte pode secretar, nos
homens, corresponde ao pH da urina de cerca 4, 5, isto é, a concentração de H+ na urina é 1000 vezes a
concentração no plasma; pH 4, 5, é o pH-limitante. Se não existissem tampões ou outras substâncias que
"fixam" H+ na urina, este pH poderia ser alcançado rapidamente, e a secreção de H+ parada. No entanto,
três importantes reações no líquido tubular removem H+ livre, permitindo que mais ácido seja secretado.
Estas são as reações com HCO3- para formar CO2 e H2O, com HPO42- para formar H2PO4- e com NH3
para formar NH4+.
Rins artificiais: têm sido construídos e estão disponíveis em muitos hospitais para o atendimento de
casos de doenças renais agudas ou de envenenamento do sangue. O sangue é desviado de uma artéria,
passando por um conjunto de tubos plásticos com membranas semipermeáveis banhados por um líquido
hipotônico em relação ao sangue e depois devolvido a uma veia. As membranas dos tubos plásticos tem
poros de diâmetro semelhante ao dos capilares dos glomérulos, de modo que as substâncias difundir-se-ão
numa direção ou noutra, dependendo das concentrações de cada uma no banho e no sangue. Dosando-se a
concentração de substâncias do banho, consegue-se adicionar algumas ao sangue, ou dele retirá-las, de
acordo com a necessidade.
11